
Genome-Wide Identification, Characterization and Expression Analysis of the TaDUF724 Gene Family in Wheat (Triticum aestivum)


 and
and
Abstract
:1. Introduction
2. Results
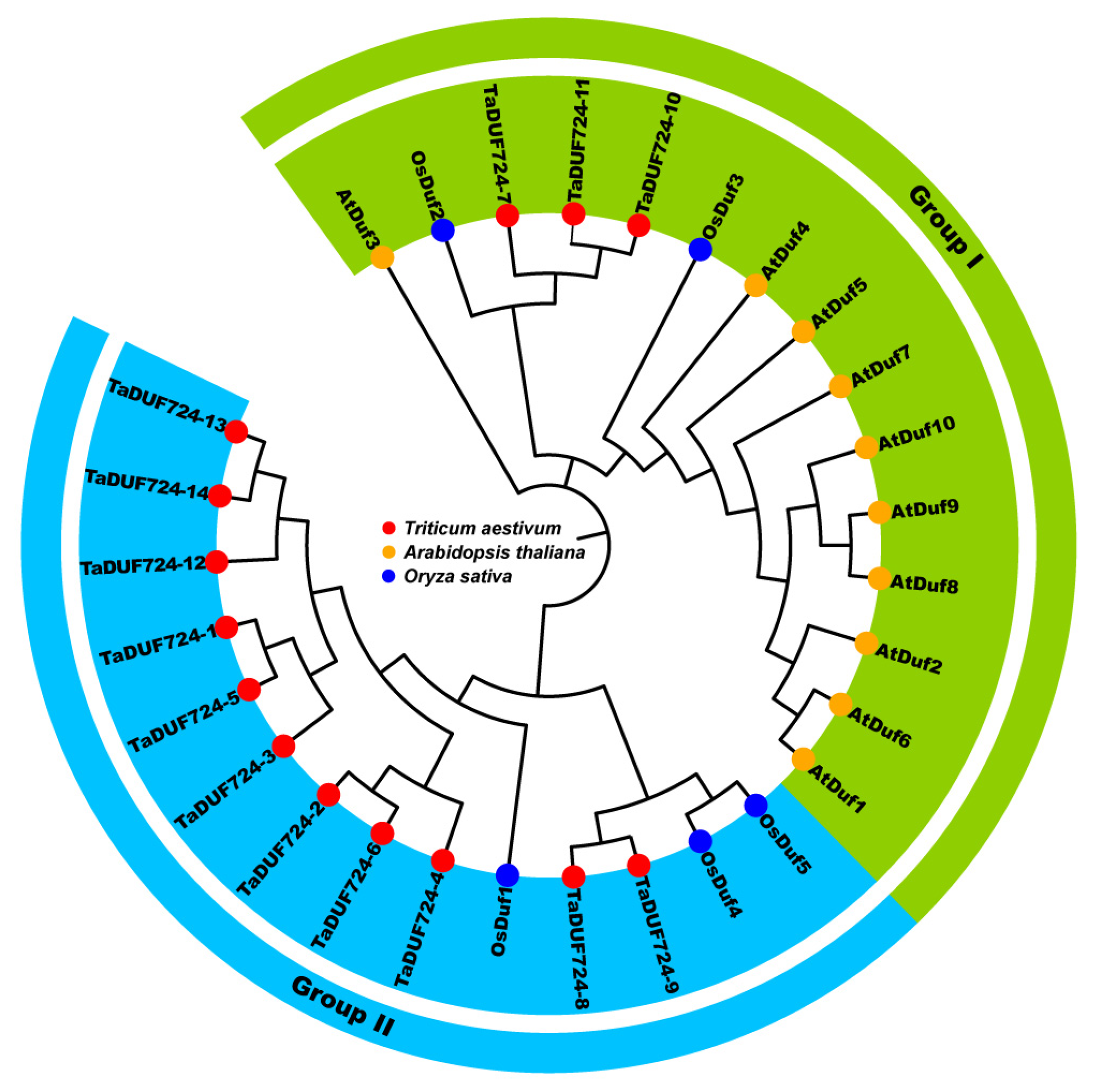
2.1. Identification and Phylogenetic Analysis of TaDUF724s
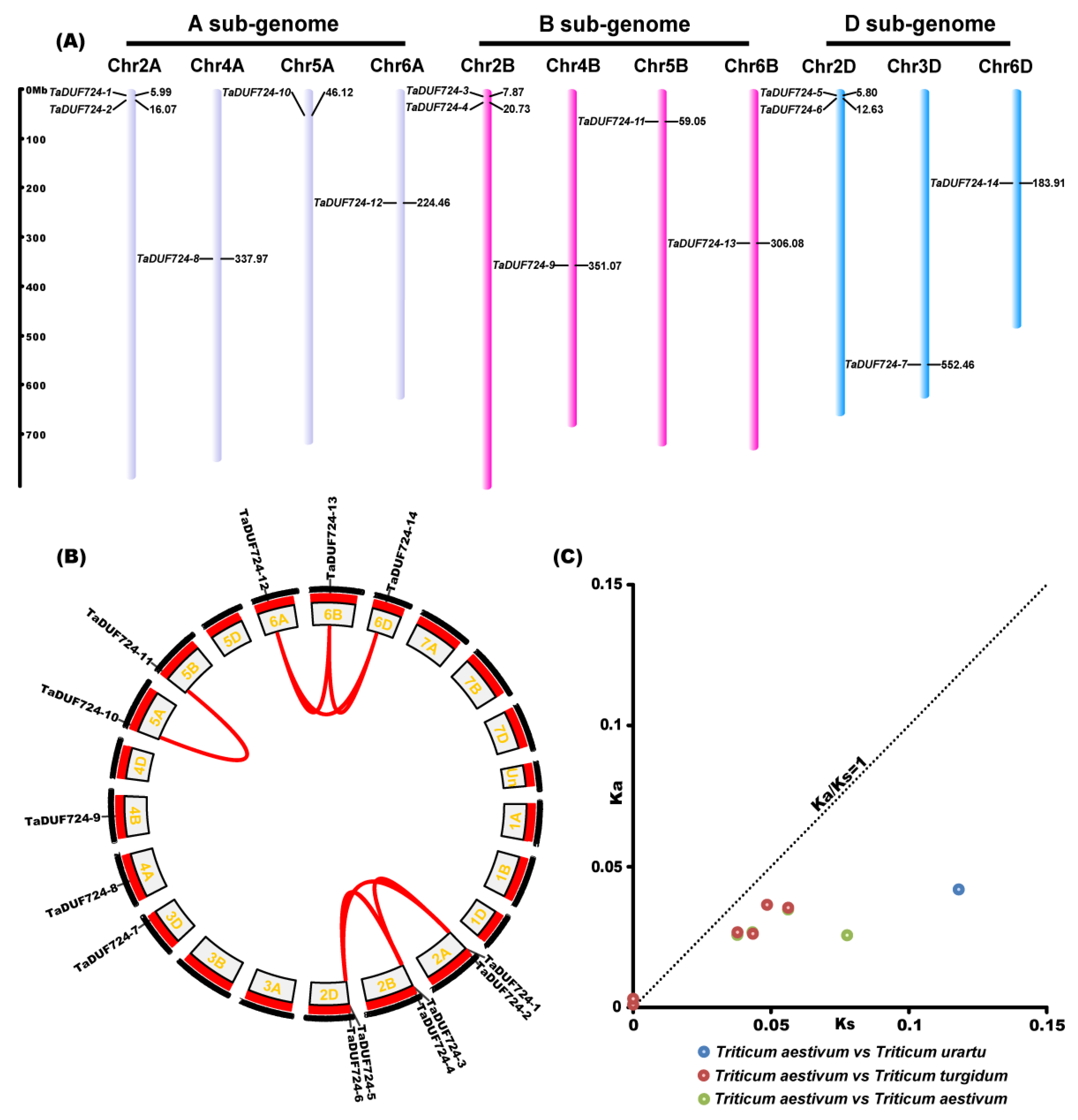
2.2. Chromosome Localization, Evolution Analysis of TaDUF724s
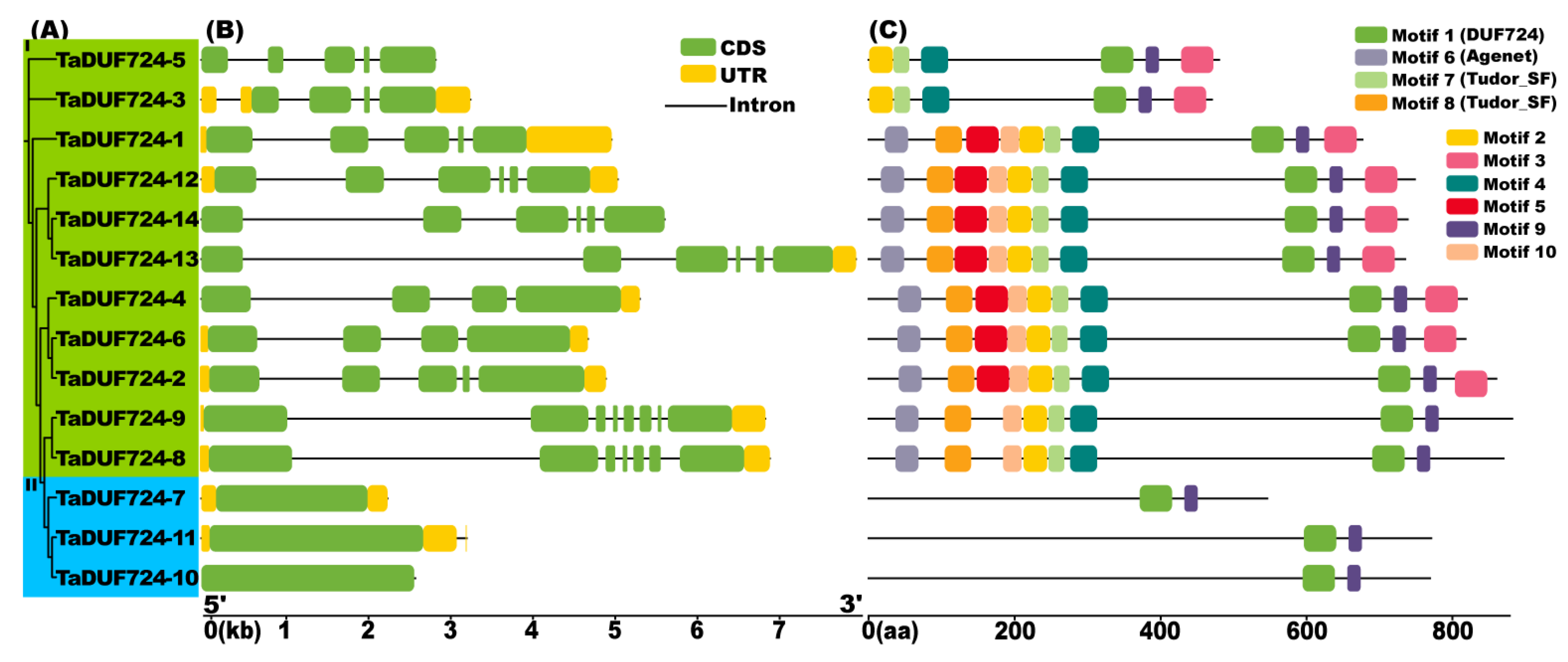
2.3. Gene Structure and Conserved Motif Analysis of TaDUF724s
2.4. Analysis of Protein Properties of TaDUF724s
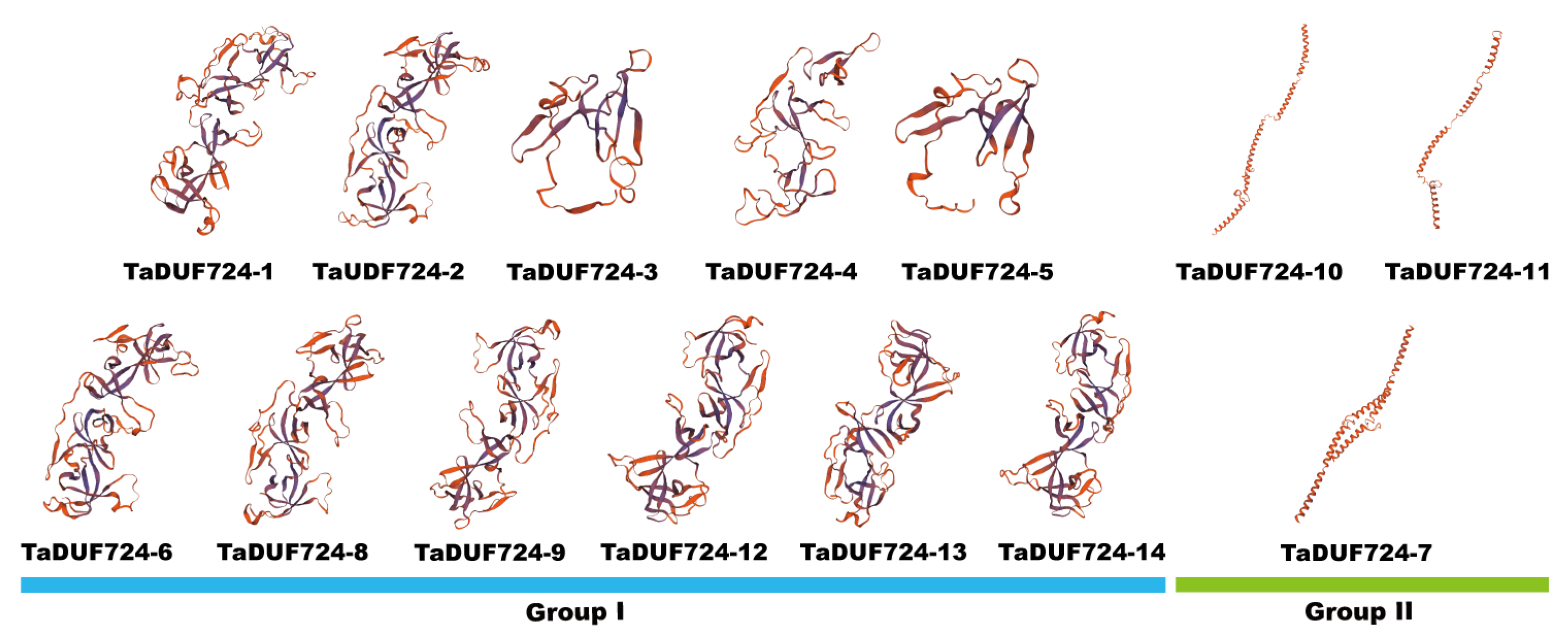
2.5. 3D-Structure Analysis of TaDUF724 Protein
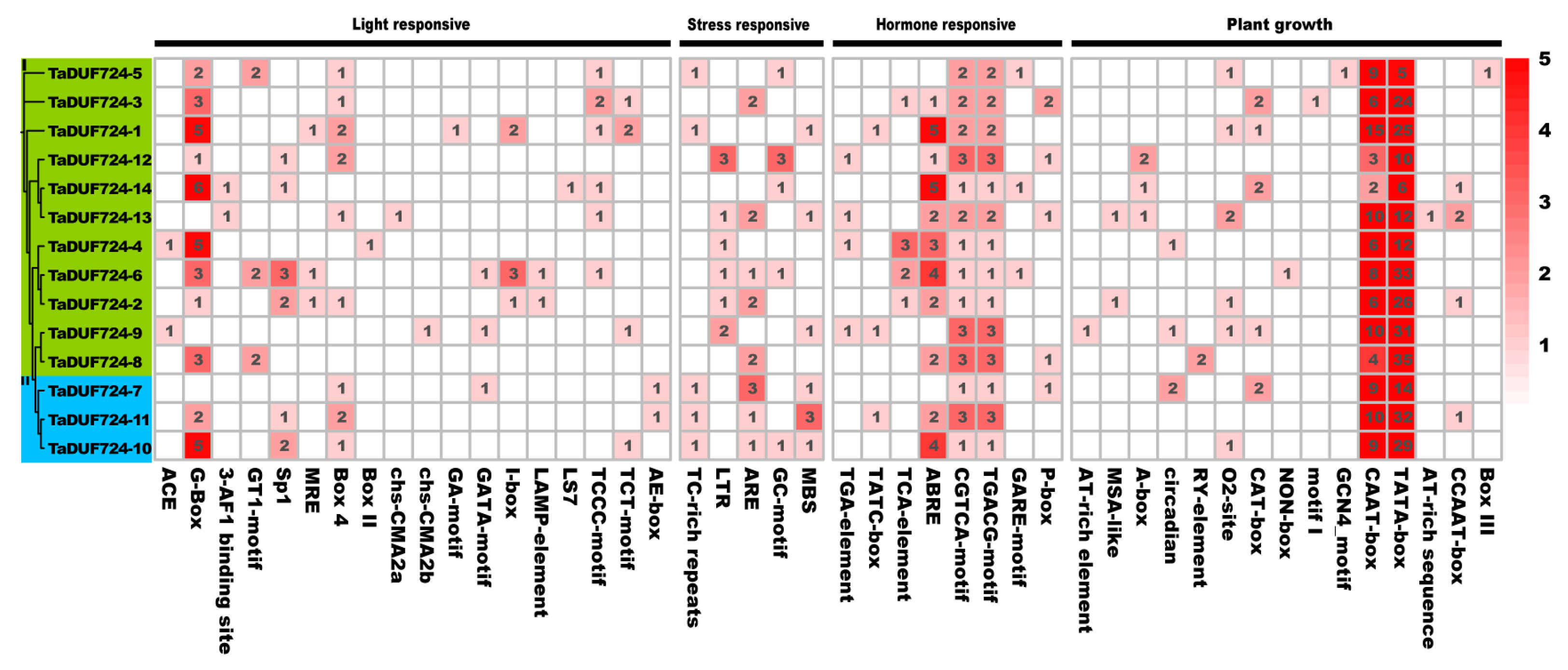
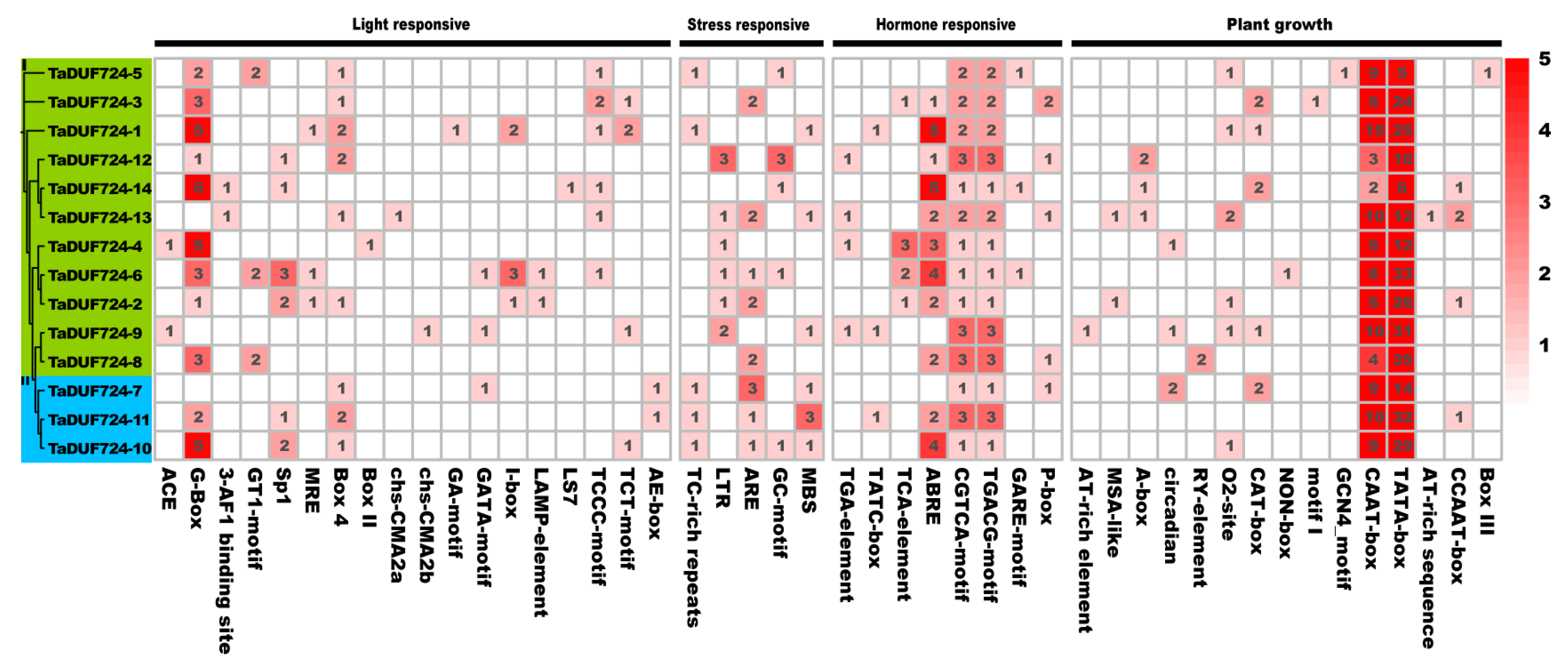
2.6. Promoter Cis-Element Analysis of TaDUF724s
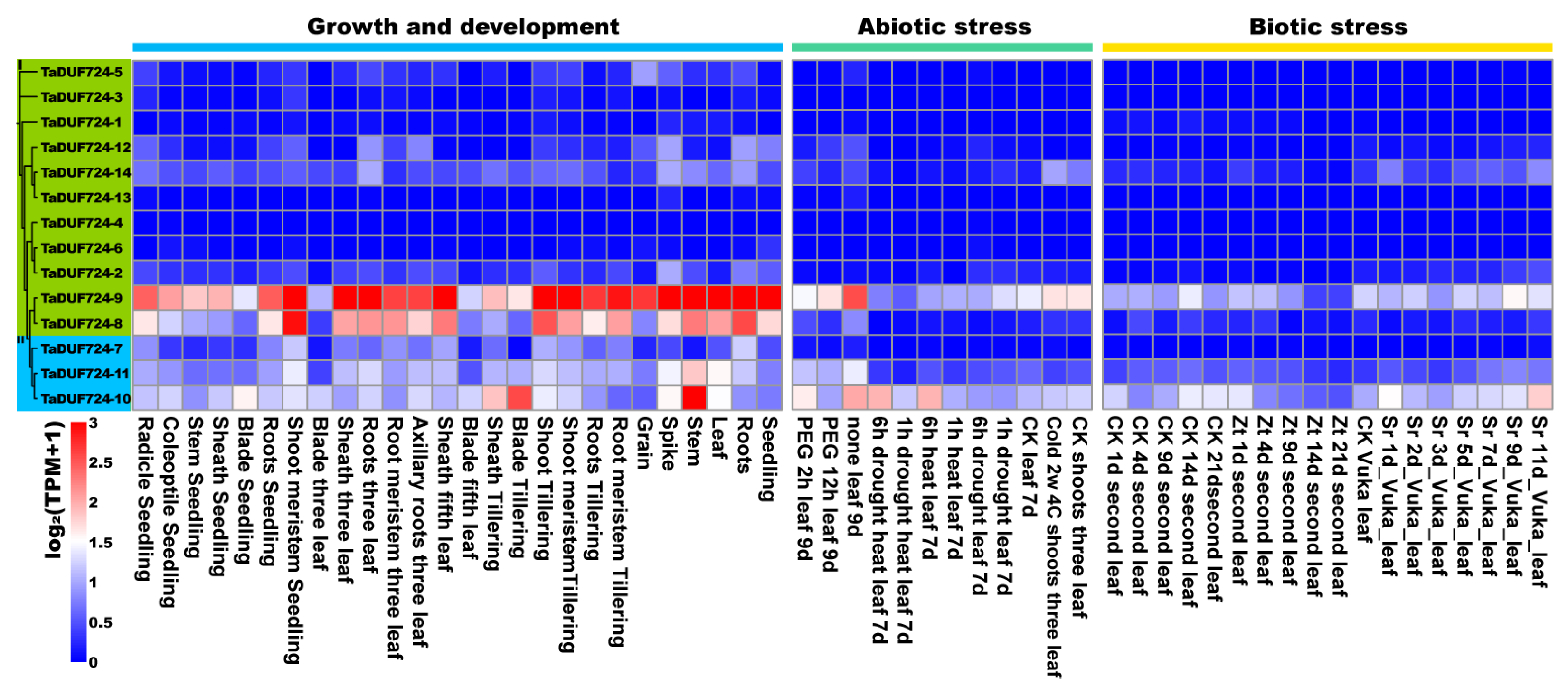
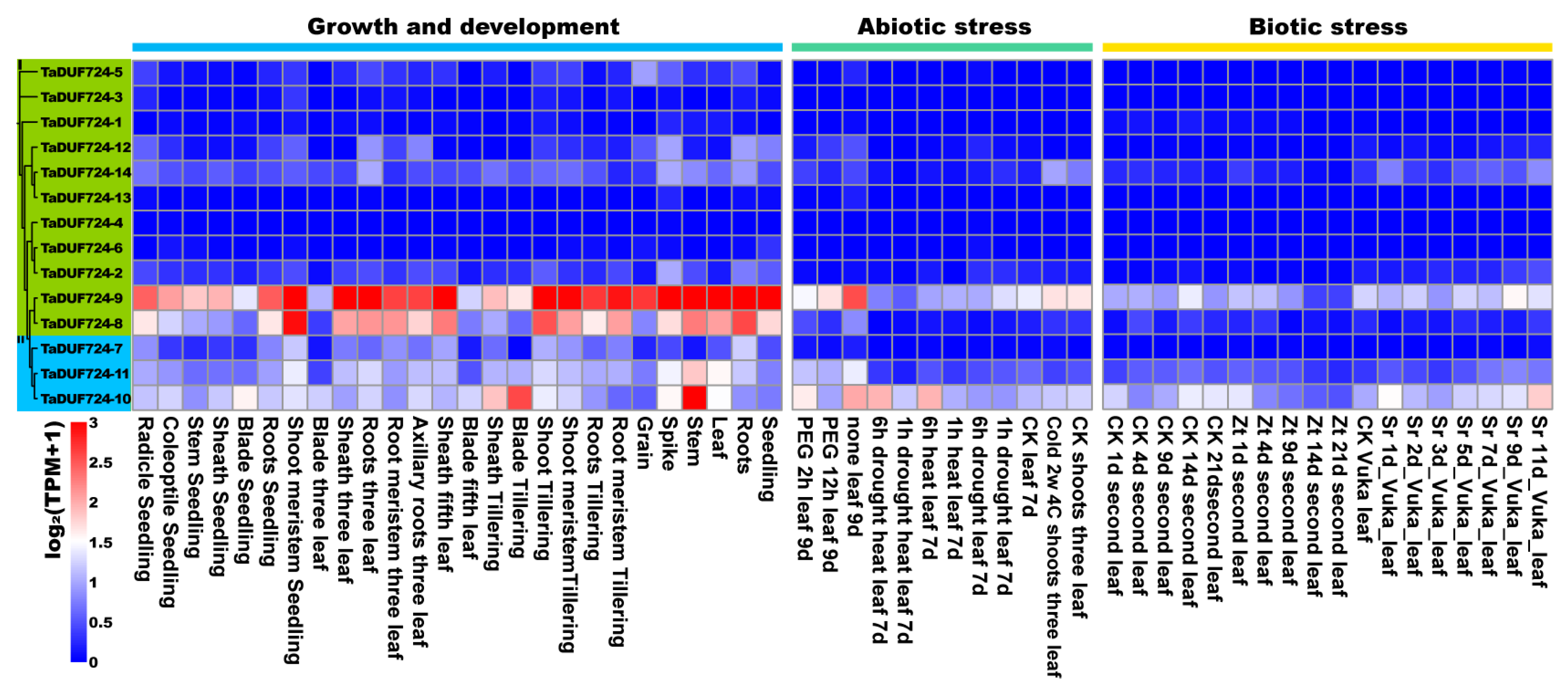
2.7. Analysis of TaDUF724 Gene Family Expression Pattern
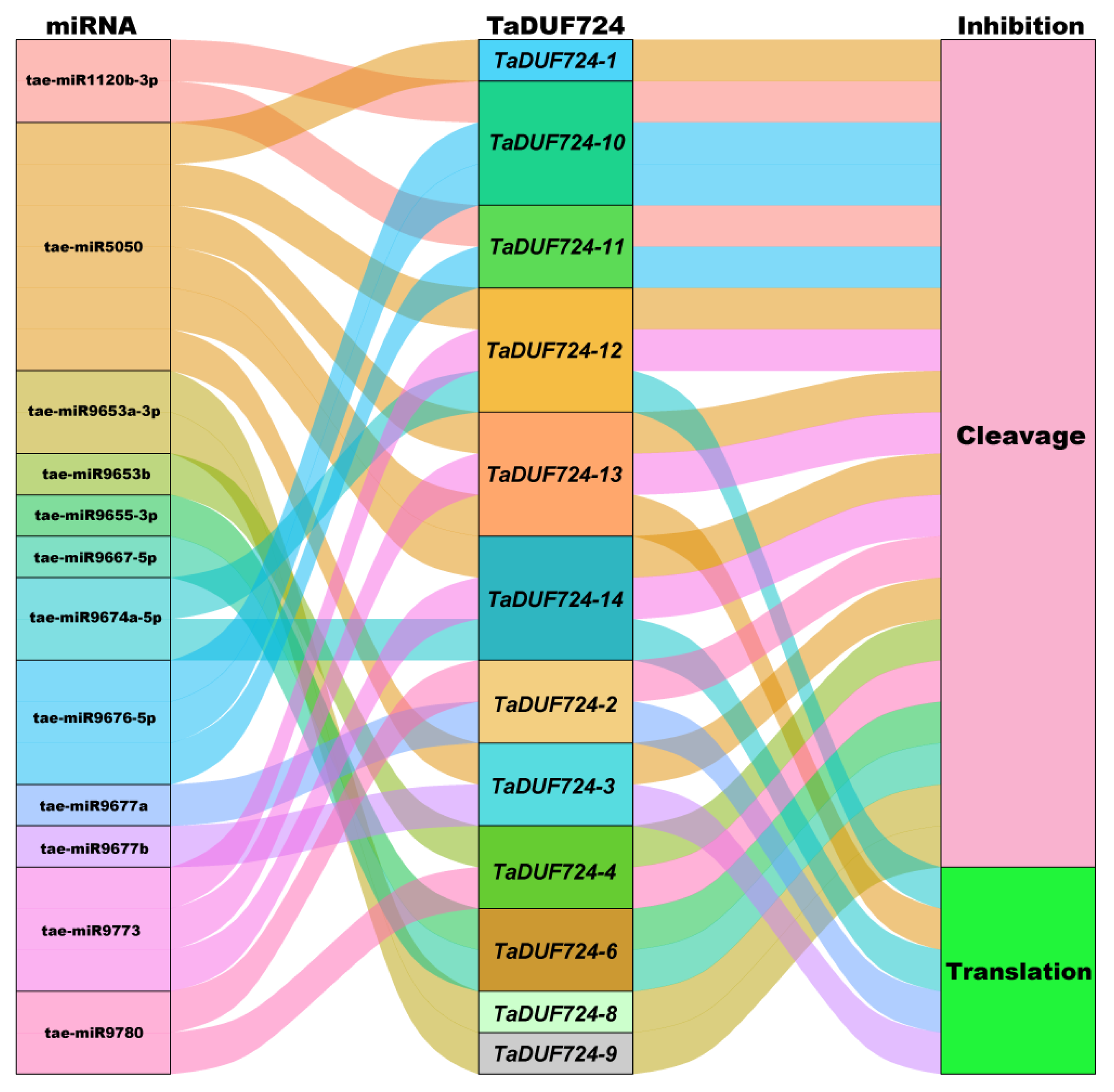
2.8. Post-Transcriptional Regulation of TaDUF724s by miRNA
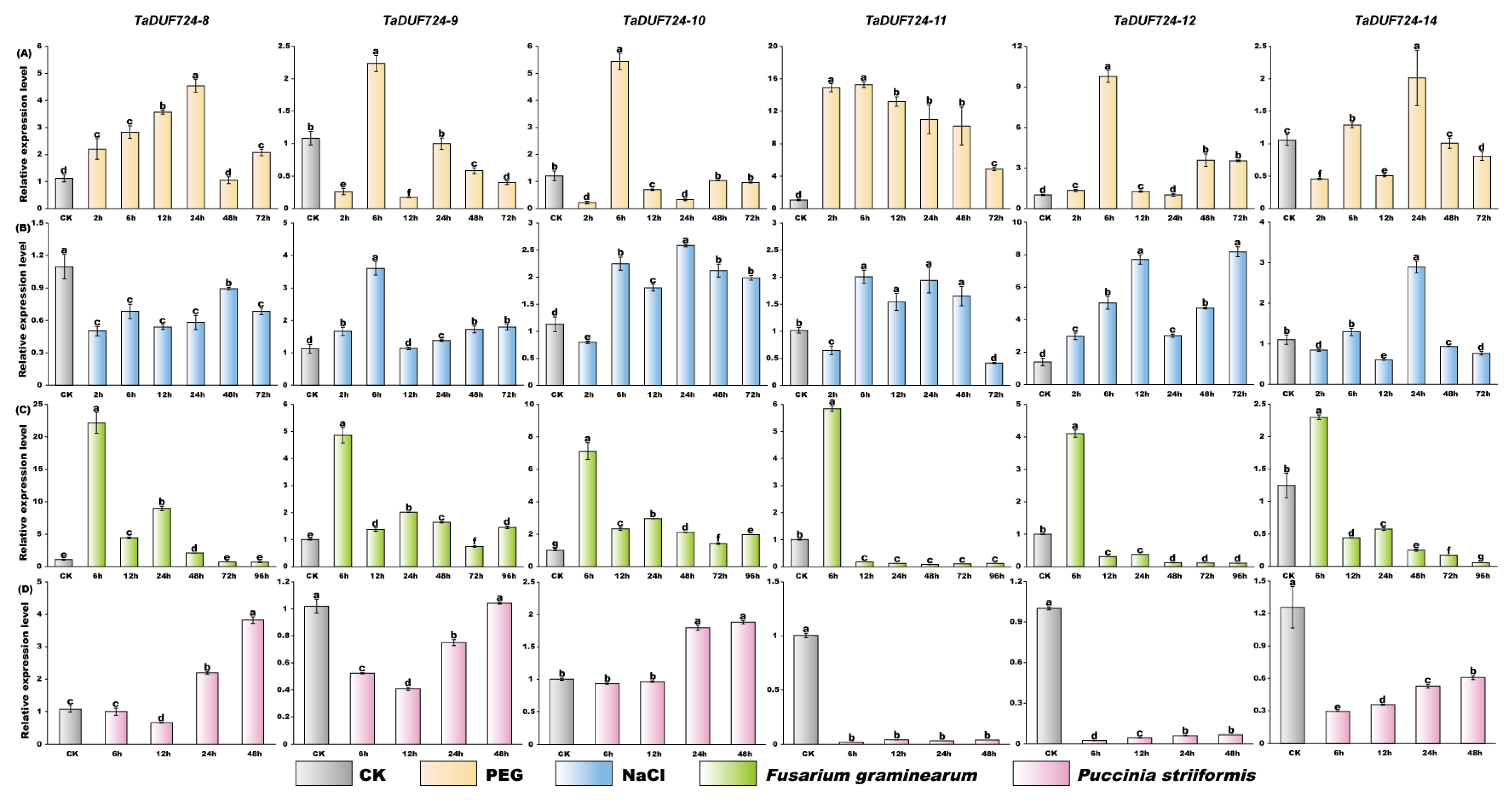
2.9. Quantitative Real-Time PCR and Data Analysis
3. Discussion
4. Materials and Methods
4.1. Identification of DUF724 Gene Family Members in Wheat
4.2. Multiple Sequence Alignment and Phylogenetic Trees
4.3. Chromosome Localization, Collinearity and Ka/Ks Analysis
4.4. Analysis of Protein Conserved Motifs and Gene Structure
4.5. Characteristic Analysis of TaDUF724 Protein
4.6. Promoter Cis-Acting Element Analysis
4.7. Expression Pattern Analysis of TaDUF724s
4.8. Prediction of Targeting Relationship between miRNA and TaDUF724s
4.9. Wheat Materials and Handling
4.10. RNA Extraction and RT-qPCR
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, Y.; Xi, K.; Liu, X.; Han, S.; Han, X.; Li, G.; Yang, L.; Ma, D.; Fang, Z.; Gong, S.; et al. Silica nanoparticles promote wheat growth by mediating hormones and sugar metabolism. J. Nanobiotechnol. 2023, 21, 2. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.; Rizvi, A.; Saif, S.; Zaidi, A. Phosphate-solubilizing microorganisms in sustainable production of wheat: Current perspective. Probiot. Agroecosyst. 2017, 51–81. [Google Scholar] [CrossRef]
- Jiang, W.; Geng, Y.; Liu, Y.; Chen, S.; Cao, S.; Li, W.; Chen, H.; Ma, D.; Yin, J. Genome-wide identification and characterization of SRO gene family in wheat: Molecular evolution and expression profiles during different stresses. Plant Physiol. Biochem. 2020, 154, 590–611. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.L.; Fang, Z.W.; Sun, C.; Zhang, P.; Zhang, X.; Lu, C.; Wang, S.P.; Ma, D.F.; Zhu, Y.X. Rapid identification of a stripe rust resistant gene in a space-induced wheat mutant using specific locus amplified fragment (SLAF) sequencing. Sci. Rep. 2018, 8, 3086. [Google Scholar] [CrossRef]
- Yin, X.; Yuan, Y.; Han, X.; Han, S.; Li, Y.; Ma, D.; Fang, Z.; Gong, S.; Yin, J. Genome-Wide Identification, Characterization, and Expression Profiling of TaDUF668 Gene Family in Triticum aestivum. Agronomy 2023, 13, 2178. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.; Tosatto, S.C.; Paladin, L.; Raj, S.; Richardson, L.J. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Lv, P.; Wan, J.; Zhang, C.; Hina, A.; Al Amin, G.; Begum, N.; Zhao, T. Unraveling the Diverse Roles of Neglected Genes Containing Domains of Unknown Function (DUFs): Progress and Perspective. Int. J. Mol. Sci. 2023, 24, 4187. [Google Scholar] [CrossRef]
- Zhou, X.; Zhu, X.; Shao, W.; Song, J.; Jiang, W.; He, Y.; Yin, J.; Ma, D.; Qiao, Y. Genome-wide mining of wheat DUF966 gene family provides new insights into salt stress responses. Front. Plant Sci. 2020, 11, 569838. [Google Scholar] [CrossRef]
- Zhong, H.; Zhang, H.; Guo, R.; Wang, Q.; Huang, X.; Liao, J.; Li, Y.; Huang, Y.; Wang, Z. Characterization and functional divergence of a novel DUF668 gene family in rice based on comprehensive expression patterns. Genes 2019, 10, 980. [Google Scholar] [CrossRef]
- Yang, Q.; Niu, X.; Tian, X.; Zhang, X.; Cong, J.; Wang, R.; Zhang, G.; Li, G. Comprehensive genomic analysis of the DUF4228 gene family in land plants and expression profiling of ATDUF4228 under abiotic stresses. BMC Genom. 2020, 21, 12. [Google Scholar] [CrossRef]
- Maurer-Stroh, S.; Dickens, N.J.; Hughes-Davies, L.; Kouzarides, T.; Eisenhaber, F.; Ponting, C.P. The Tudor domain ‘Royal Family’: Tudor, plant agenet, chromo, PWWP and MBT domains. Trends Biochem. Sci. 2003, 28, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Brasil, J.N.; Cabral, L.M.; Eloy, N.B.; Primo, L.M.; Barroso-Neto, I.L.; Grangeiro, L.P.P.; Gonzalez, N.; Inzé, D.; Ferreira, P.C.; Hemerly, A.S. AIP1 is a novel Agenet/Tudor domain protein from Arabidopsis that interacts with regulators of DNA replication, transcription and chromatin remodeling. BMC Plant Biol. 2015, 15, 270. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, M.; Yang, C.; Zhao, L.; Qin, G.; Peng, L.; Zheng, Q.; Nie, W.; Song, C.P.; Shi, H. SWO1 modulates cell wall integrity under salt stress by interacting with importin α in Arabidopsis. Stress Biol. 2021, 1, 22. [Google Scholar] [CrossRef] [PubMed]
- Alpatov, R.; Lesch, B.J.; Nakamoto-Kinoshita, M.; Blanco, A.; Chen, S.; Stützer, A.; Armache, K.J.; Simon, M.D.; Xu, C.; Ali, M.; et al. A Chromatin-Dependent Role of the Fragile X Mental Retardation Protein FMRP in the DNA Damage Response. Cell 2014, 157, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Yang, K.-Z.; Xia, C.; Zhang, X.-Q.; Chen, L.-Q.; Ye, D. Characterization of DUF724 gene family in Arabidopsis thaliana. Plant Mol. Biol. 2010, 72, 61–73. [Google Scholar] [CrossRef]
- Ru, J.-N.; Hou, Z.-H.; Zheng, L.; Zhao, Q.; Wang, F.-Z.; Chen, J.; Zhou, Y.-B.; Chen, M.; Ma, Y.-Z.; Xi, Y.-J. Genome-wide analysis of DEAD-box RNA helicase family in wheat (Triticum aestivum) and functional identification of TaDEAD-box57 in abiotic stress responses. Front. Plant Sci. 2021, 12, 797276. [Google Scholar] [CrossRef]
- Lin, M.; Dong, Z.; Zhou, H.; Wu, G.; Xu, L.; Ying, S.; Chen, M. Genome-Wide Identification and Transcriptional Analysis of the MYB Gene Family in Pearl Millet (Pennisetum glaucum). Int. J. Mol. Sci. 2023, 24, 2484. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Z.; Vang, S.; Yu, J.; Wong, G.K.-S.; Wang, J. Correlation between Ka/Ks and Ks is related to substitution model and evolutionary lineage. J. Mol. Evol. 2009, 68, 414–423. [Google Scholar] [CrossRef]
- Lei, P.; Wei, X.; Gao, R.; Huo, F.; Nie, X.; Tong, W.; Song, W. Genome-wide identification of PYL gene family in wheat: Evolution, expression and 3D structure analysis. Genomics 2021, 113, 854–866. [Google Scholar] [CrossRef]
- Xu, G.; Guo, C.; Shan, H.; Kong, H. Divergence of duplicate genes in exon-intron structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1187–1192. [Google Scholar] [CrossRef]
- Jeffares, D.C.; Penkett, C.J.; Bähler, J. Rapidly regulated genes are intron poor. Trends Genet. 2008, 24, 375–378. [Google Scholar] [CrossRef] [PubMed]
- Yao, F.; Song, C.; Wang, H.; Song, S.; Bai, T. Genome-Wide Characterization of the HSP20 Gene Family Identifies Potential Members Involved in Temperature Stress Response in Apple. Front. Genet. 2020, 11, 609184. [Google Scholar] [CrossRef]
- Verma, S.; Bhatia, S. Analysis of genes encoding seed storage proteins (SSPs) in chickpea (Cicer arietinum L.) reveals co-expressing transcription factors and a seed-specific promoter. Funct. Integr. Genom. 2019, 19, 373–390. [Google Scholar] [CrossRef] [PubMed]
- Gagneux, P. Protein Structure and Function. J. Hered. 2004, 95, A67–A75. [Google Scholar] [CrossRef]
- Su, D.; Xiang, W.; Wen, L.; Lu, W.; Li, Z. Genome-wide identification, characterization and expression analysis of BES1 gene family in tomato. BMC Plant Biol. 2021, 21, 161. [Google Scholar] [CrossRef]
- Garcia, C.M.H.; Finer, J.J. Identification and Validation of Promoters and Cis-acting Regulatory Elements. Plant Sci. 2014, 217, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Xiao-Hui, Z. Analysis on TATA-box, GC-box and CAAT-box in Eukaryotic Promoters. J. Anhui Agric. Sci. 2008, 36, 1380. [Google Scholar]
- Li, S.; Xu, C.; Yang, Y.; Xia, G. Functional analysis of TaDi19A, a salt-responsive gene in wheat. Plant Cell Environ. 2010, 33, 117–129. [Google Scholar]
- Gong, S.; Jin, X.; Xiao, Y.; Li, Z. Ocean acidification and warming lead to increased growth and altered chloroplast morphology in the thermo-tolerant alga Symbiochlorum hainanensis. Front. Plant Sci. 2020, 11, 585202. [Google Scholar] [CrossRef]
- BB, B. Overexpression of the nuclear protein gene AtDUF4 increases organ size in Arabidopsis thaliana and Brassica napus. J. Genet. Genom. 2018, 45, 459e462. [Google Scholar]
- Cai, Y.; Yu, X.; Hu, S.; Yu, J. A Brief Review on the Mechanisms of miRNA Regulation. Genom. Proteom. Bioinform. 2009, 7, 8. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.-F.; Wang, Y.-K.; Wang, P.; Duan, W.-J.; Yuan, S.-H.; Sun, H.; Yuan, G.-L.; Ma, J.-X.; Wang, N.; Zhang, F.-T. Uncovering male fertility transition responsive miRNA in a wheat photo-thermosensitive genic male sterile line by deep sequencing and degradome analysis. Front. Plant Sci. 2017, 8, 1370. [Google Scholar] [CrossRef] [PubMed]
- Akdogan, G.; Tufekci, E.D.; Uranbey, S.; Unver, T. miRNA-based drought regulation in wheat. Funct. Integr. Genom. 2016, 16, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Yanfei, D.; Yueliang, T.; Cheng, Z. Emerging roles of microRNAs in the mediation of drought stress response in plants. J. Exp. Bot. 2013, 64, 3077–3086. [Google Scholar]
- Luo, L.; Bai, J.; Yuan, S.; Guo, L.; Liu, Z.; Guo, H.; Zhang, T.; Duan, W.; Li, Y.; Zhao, C. Genome Wide identification and characterization of wheat GH9 genes reveals their roles in pollen development and anther dehiscence. Int. J. Mol. Sci. 2022, 23, 6324. [Google Scholar] [CrossRef]
- Han, Y.; Zhao, Y.; Wang, H.; Zhang, Y.; Ding, Q.; Ma, L. Identification of ceRNA and candidate genes related to fertility conversion of TCMS line YS3038 in wheat. Plant Physiol. Biochem. PPB 2021, 158, 190–207. [Google Scholar] [CrossRef]
- Hongxia, L.; Jinglei, G.; Chengyang, Z.; Weijun, Z.; Yulong, S.; Yu, W. Identification of Differentially Expressed miRNAs between a Wheat K-type Cytoplasmic Male Sterility Line and Its Near-Isogenic Restorer Line. Plant Cell Physiol. 2019, 60, 1604–1618. [Google Scholar]
- Pomeranz, M.C.; Hah, C.; Lin, P.C.; Kang, S.G.; Finer, J.J.; Blackshear, P.J.; Jang, J.C. The Arabidopsis Tandem Zinc Finger Protein AtTZF1 Traffics between the Nucleus and Cytoplasmic Foci and Binds Both DNA and RNA. Plant Physiol. 2010, 152, 151–165. [Google Scholar] [CrossRef]
- Xiao, M.; Li, J.; Li, W.; Wang, Y.; Yu, W. MicroRNAs activate gene transcription epigenetically as an enhancer trigger. RNA Biol. 2017, 14, 1326–1334. [Google Scholar] [CrossRef]
- Alaux, M.; Rogers, J.; Letellier, T.; Flores, R.; Alfama, F.; Pommier, C.; Mohellibi, N.; Durand, S.; Kimmel, E.; Michotey, C. Linking the International Wheat Genome Sequencing Consortium bread wheat reference genome sequence to wheat genetic and phenomic data. Genome Biol. 2018, 19, 1–10. [Google Scholar] [CrossRef]
- Peng, S.; Li, X.; Jiang, W.; Wang, Y.; Xiang, G.; Li, M.; Wang, Y.; Yang, Z.; Li, Y.; Liu, X.; et al. Identification of two key UDP-glycosyltransferases responsible for the ocotillol-type ginsenoside majonside-R2 biosynthesisin Panax vietnamensis var. fuscidiscus. Planta 2023, 257. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.J.; Yang, L.Y.; He, Y.H.; Zhang, H.Z.; Yin, J.Y. Genome-wide identification and transcriptional expression analysis of superoxide dismutase (SOD) family in wheat (Triticum aestivum). PeerJ 2019, 7, e8062. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Wang, L.; Zhao, J.; Li, Y.; Huang, R.; Jiang, X.; Zhou, X.; Zhu, X.; He, Y.; He, Y.; et al. Genome-wide characterization of the C2H2 zinc-finger genes in Cucumis sativus and functional analyses of four CsZFPs in response to stresses. BMC Plant Biol. 2020, 20, 359. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Li, Y.; Liu, X.; Xiao, Y.; Wen, Y.; Li, K.; Ma, Z.; Yang, L.; Zhu, Y.; Yin, J. Genome-wide characterization and function analysis uncovered roles of wheat LIMs in responding to adverse stresses and TaLIM8-4D function as a susceptible gene. Plant Genome 2022, 15, e20246. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Zhu, Y.X.; Yang, L.; Liu, N.; Yang, J.; Zhou, X.K.; Xia, Y.C.; He, Y.; He, Y.Q.; Gong, H.J.; Ma, D.F. Genome-wide identification, structure characterization, and expression pattern profiling of aquaporin gene family in cucumber. BMC Plant Biol. 2019, 19, 345. [Google Scholar] [CrossRef]
- Tian, J.; Li, Y.; Hu, Y.; Zhong, Q.; Yin, J.; Zhu, Y. Mining the Roles of Cucumber DUF966 Genes in Fruit Development and Stress Response. Plants 2022, 11, 2497. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Fang, Z.-W.; He, Y.-Q.; Liu, Y.-K.; Jiang, W.-Q.; Song, J.-H.; Wang, S.-P.; Yin, J.-L. Bioinformatic identification and analyses of the non-specific lipid transfer proteins in wheat. J. Integr. Agric. 2020, 19, 1170–1185. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Yin, J.; Wang, L.; Xi, K.; Zhu, X.; Li, G.; Zhu, Y.; Liu, Y. Identification and evolutionary analysis of the metal-tolerance protein family in eight Cucurbitaceae species. Plant Genome 2022, 15, e20167. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Guo, G.; Ni, Z.; Sunkar, R.; Du, J.; Zhu, J.-K.; Sun, Q. Cloning and characterization of microRNAs from wheat (Triticum aestivum L.). Genome Biol. 2007, 8, R96. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhu, Y.; Ma, D.; Xu, W.; Zhou, J.; Yan, H.; Yang, L.; Yin, J. Screening, identification, and optimization of fermentation conditions of an antagonistic endophyte to wheat head blight. Agronomy 2019, 9, 476. [Google Scholar] [CrossRef]
- Yang, X.; Ma, J.; Li, H.; Ma, H.; Yao, J.; Liu, C. Different genes can be responsible for crown rot resistance at different developmental stages of wheat and barley. Eur. J. Plant Pathol. 2010, 128, 495–502. [Google Scholar] [CrossRef]
- Zhan, C.; Li, Y.; Li, H.; Wang, M.; Gong, S.; Ma, D.; Li, Y. Phylogenomic analysis of phenylalanine ammonia-lyase (PAL) multigene family and their differential expression analysis in wheat (Triticum aestivum L.) suggested their roles during different stress responses. Front. Plant Sci. 2022, 13, 982457. [Google Scholar] [CrossRef]
- Sahin, U.; Ekinci, M.; Kiziloglu, F.M.; Yildirim, E.; Turan, M.; Kotan, R.; Ors, S. Ameliorative effects of plant growth promoting bacteria on water-yield relationships, growth, and nutrient uptake of lettuce plants under different irrigation levels. HortScience 2015, 50, 1379–1386. [Google Scholar] [CrossRef]
- Yin, J.; Han, X.; Zhu, Y.; Fang, Z.; Gao, D.; Ma, D. Transcriptome profiles of circular rnas in common wheat during fusarium head blight disease. Data 2022, 7, 121. [Google Scholar] [CrossRef]
- Yin, J.; Hou, L.; Jiang, X.; Yang, J.; He, Y.; Zhou, X.; Zhu, X.; Gong, A.; Zhu, Y.; Chen, Z. Identification and validation of reference genes for quantitative real-time PCR studies in alligatorweed (Alternanthera philoxeroides). Weed Sci. 2021, 69, 404–411. [Google Scholar] [CrossRef]
- Yin, J.; Liu, M.; Ma, D.; Wu, J.; Li, S.; Zhu, Y.; Han, B. Identification of circular RNAs and their targets during tomato fruit ripening. Postharvest Biol. Technol. 2018, 136, 90–98. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene ID | Len | MW | Pi | Ins | Sta | GRAVY | Sig | Sub |
|---|---|---|---|---|---|---|---|---|---|
| TaDUF724-1 | TraesCS2A03G0023500.1 | 752 | 82.4 | 5.8 | 65 | not | −0.436 | no | Nuc |
| TaDUF724-2 | TraesCS2A03G0056500.1 | 956 | 106 | 5 | 51.4 | not | −0.465 | no | Nuc |
| TaDUF724-3 | TraesCS2B03G0029300.1 | 523 | 57.8 | 6.1 | 63 | not | −0.458 | no | Nuc |
| TaDUF724-4 | TraesCS2B03G0083400.1 | 911 | 101 | 4.7 | 54 | not | −0.451 | no | Nuc |
| TaDUF724-5 | TraesCS2D03G0020600.1 | 534 | 58.5 | 6.2 | 58.3 | not | −0.354 | no | Chl. Nuc |
| TaDUF724-6 | TraesCS2D03G0057200.1 | 909 | 100.5 | 4.9 | 50.3 | not | −0.464 | no | Nuc |
| TaDUF724-7 | TraesCS3D03G0969100.1 | 607 | 69.6 | 5.1 | 55.8 | not | −0.804 | no | Nuc |
| TaDUF724-8 | TraesCS4A03G0406500.1 | 967 | 107.5 | 7.6 | 58 | not | −0.62 | no | Nuc |
| TaDUF724-9 | TraesCS4B03G0462800.3 | 980 | 108.9 | 7.6 | 55.4 | not | −0.61 | no | Chl. Nuc |
| TaDUF724-10 | TraesCS5A03G0123500.1 | 855 | 96.4 | 5.8 | 58.3 | not | −0.529 | no | Chl. Nuc |
| TaDUF724-11 | TraesCS5B03G0134600.1 | 857 | 96.9 | 5.5 | 58.8 | not | −0.536 | no | Chl |
| TaDUF724-12 | TraesCS6A03G0468800.1 | 832 | 92.1 | 6.1 | 55.8 | not | −0.524 | no | Chl |
| TaDUF724-13 | TraesCS6B03G0586100.1 | 817 | 90.6 | 6 | 56.7 | not | −0.478 | no | Chl |
| TaDUF724-14 | TraesCS6D03G0391800.1 | 821 | 90.9 | 6.4 | 56.8 | not | −0.481 | no | Chl |
| Name | Alpha Helix (%) | Extended Strand (%) | Beta Turn (%) | Random Coil (%) |
|---|---|---|---|---|
| TaDUF724-1 | 36.44 | 11.97 | 7.18 | 44.41 |
| TaDUF724-2 | 34.52 | 13.81 | 5.44 | 46.23 |
| TaDUF724-3 | 44.36 | 9.37 | 7.27 | 39.01 |
| TaDUF724-4 | 33.37 | 16.03 | 6.70 | 43.91 |
| TaDUF724-5 | 43.63 | 9.18 | 6.37 | 40.82 |
| TaDUF724-6 | 32.45 | 15.95 | 7.04 | 44.55 |
| TaDUF724-7 | 44.48 | 10.71 | 5.60 | 39.21 |
| TaDUF724-8 | 35.68 | 12.62 | 5.27 | 46.43 |
| TaDUF724-9 | 35.31 | 14.29 | 6.53 | 43.88 |
| TaDUF724-10 | 44.68 | 10.06 | 5.03 | 40.23 |
| TaDUF724-11 | 44.81 | 9.22 | 5.37 | 40.61 |
| TaDUF724-12 | 33.77 | 11.42 | 5.17 | 49.64 |
| TaDUF724-13 | 35.25 | 12.36 | 5.02 | 47.37 |
| TaDUF724-14 | 35.57 | 12.79 | 5.24 | 46.41 |
| Gene Name | Forward Primer | Reverse Primer |
|---|---|---|
| Ta2291 | GCTCTCCAACAACATTGCCAAC | GCTTCTGCCTGTCACATACGC |
| TaDUF724-8 | CCAAGGTCGCCATTTCTGTC | GGGGAACCACGAGTAGCCATA |
| TaDUF724-9 | CAAGCATTGGAGGATTGTAAGTC | TTTTTGTGGCTGGAGATTGTG |
| TaDUF724-10 | ATGGTTGAATGGGCACTTTG | ACATTTATCACCTGCCGTATCTG |
| TaDUF724-11 | AAGAGGGTGGAAGAAAGCAAGA | TTGTTCACCAACCCTCTGTAATG |
| TaDUF724-12 | TTGCTTGAGGCAGATGGTTT | TTCTTGGTCCTTGGTTTCTTTG |
| TaDUF724-14 | GTATTAAGCGGTCAGGGTAAGC | CCTAGAGCCACTGCTGTTTGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, Y.; Yin, X.; Han, X.; Han, S.; Li, Y.; Ma, D.; Fang, Z.; Yin, J.; Gong, S. Genome-Wide Identification, Characterization and Expression Analysis of the TaDUF724 Gene Family in Wheat (Triticum aestivum). Int. J. Mol. Sci. 2023, 24, 14248. https://doi.org/10.3390/ijms241814248
Yuan Y, Yin X, Han X, Han S, Li Y, Ma D, Fang Z, Yin J, Gong S. Genome-Wide Identification, Characterization and Expression Analysis of the TaDUF724 Gene Family in Wheat (Triticum aestivum). International Journal of Molecular Sciences. 2023; 24(18):14248. https://doi.org/10.3390/ijms241814248
Chicago/Turabian StyleYuan, Yi, Xiaohui Yin, Xiaowen Han, Shuo Han, Yiting Li, Dongfang Ma, Zhengwu Fang, Junliang Yin, and Shuangjun Gong. 2023. "Genome-Wide Identification, Characterization and Expression Analysis of the TaDUF724 Gene Family in Wheat (Triticum aestivum)" International Journal of Molecular Sciences 24, no. 18: 14248. https://doi.org/10.3390/ijms241814248
APA StyleYuan, Y., Yin, X., Han, X., Han, S., Li, Y., Ma, D., Fang, Z., Yin, J., & Gong, S. (2023). Genome-Wide Identification, Characterization and Expression Analysis of the TaDUF724 Gene Family in Wheat (Triticum aestivum). International Journal of Molecular Sciences, 24(18), 14248. https://doi.org/10.3390/ijms241814248