

Chemical Systems for Wetware Artificial Life: Selected Perspectives in Synthetic Cell Research


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
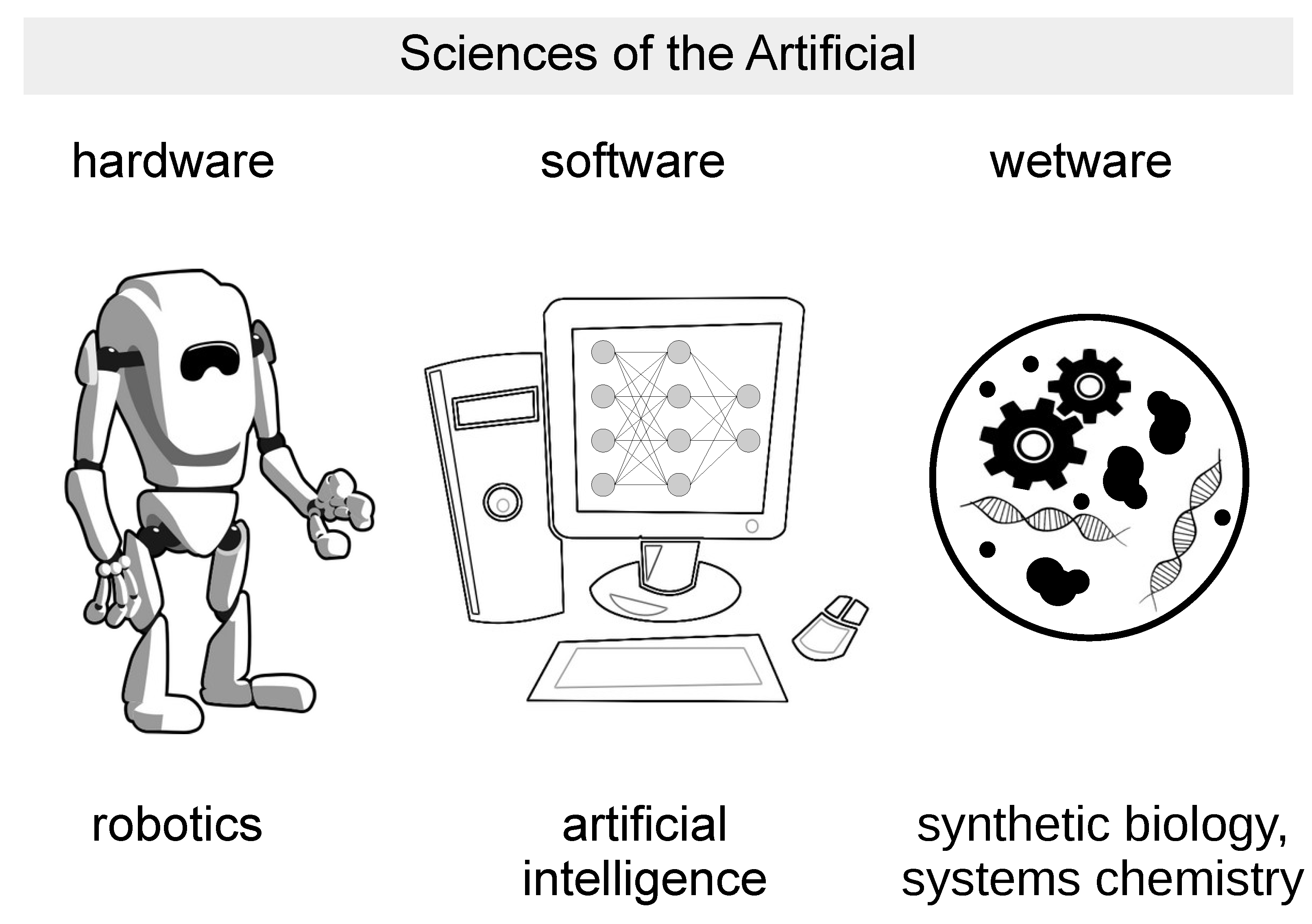
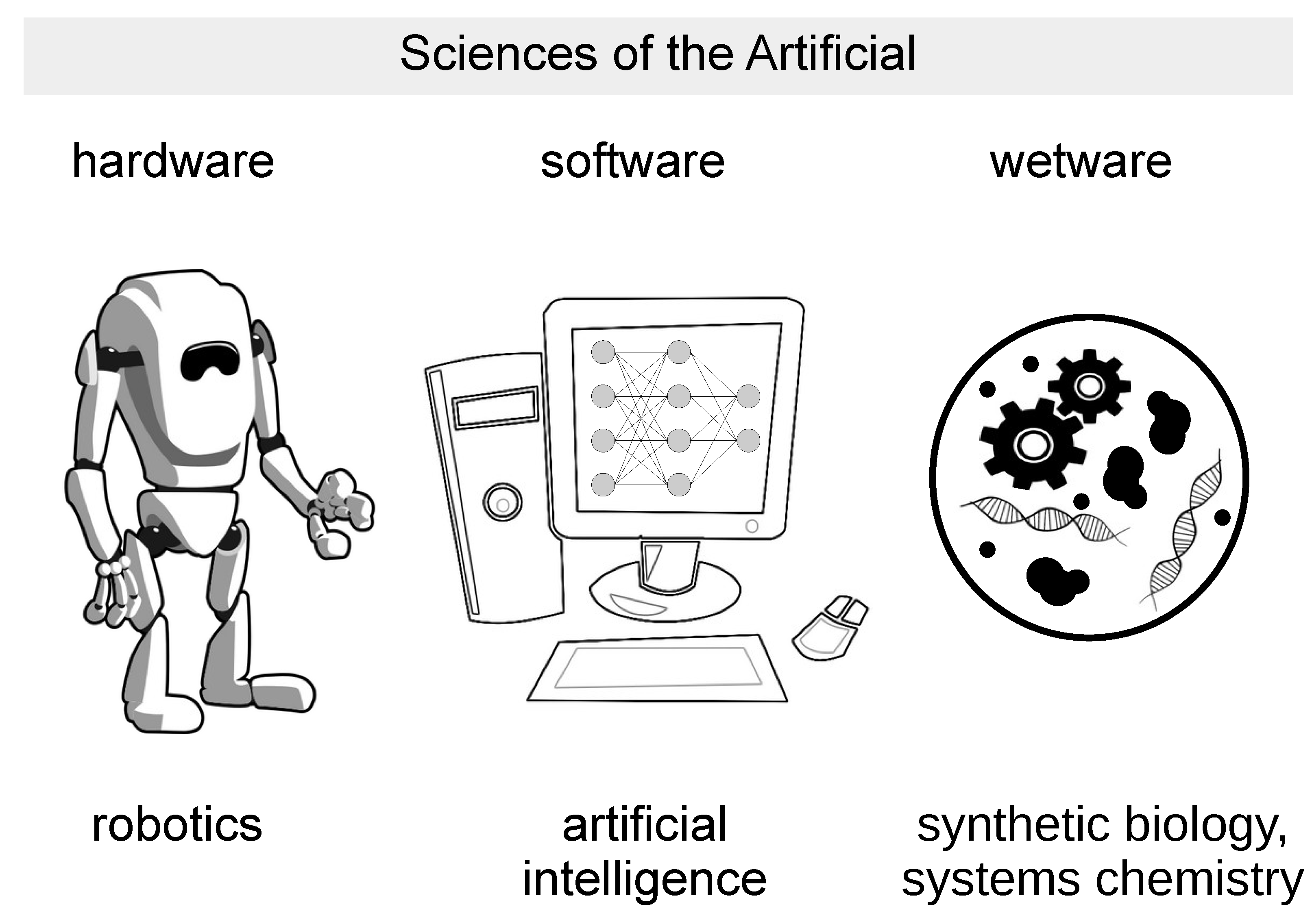
:1. Wetware Artificial Life as the Third Pillar of the Sciences of the Artificial
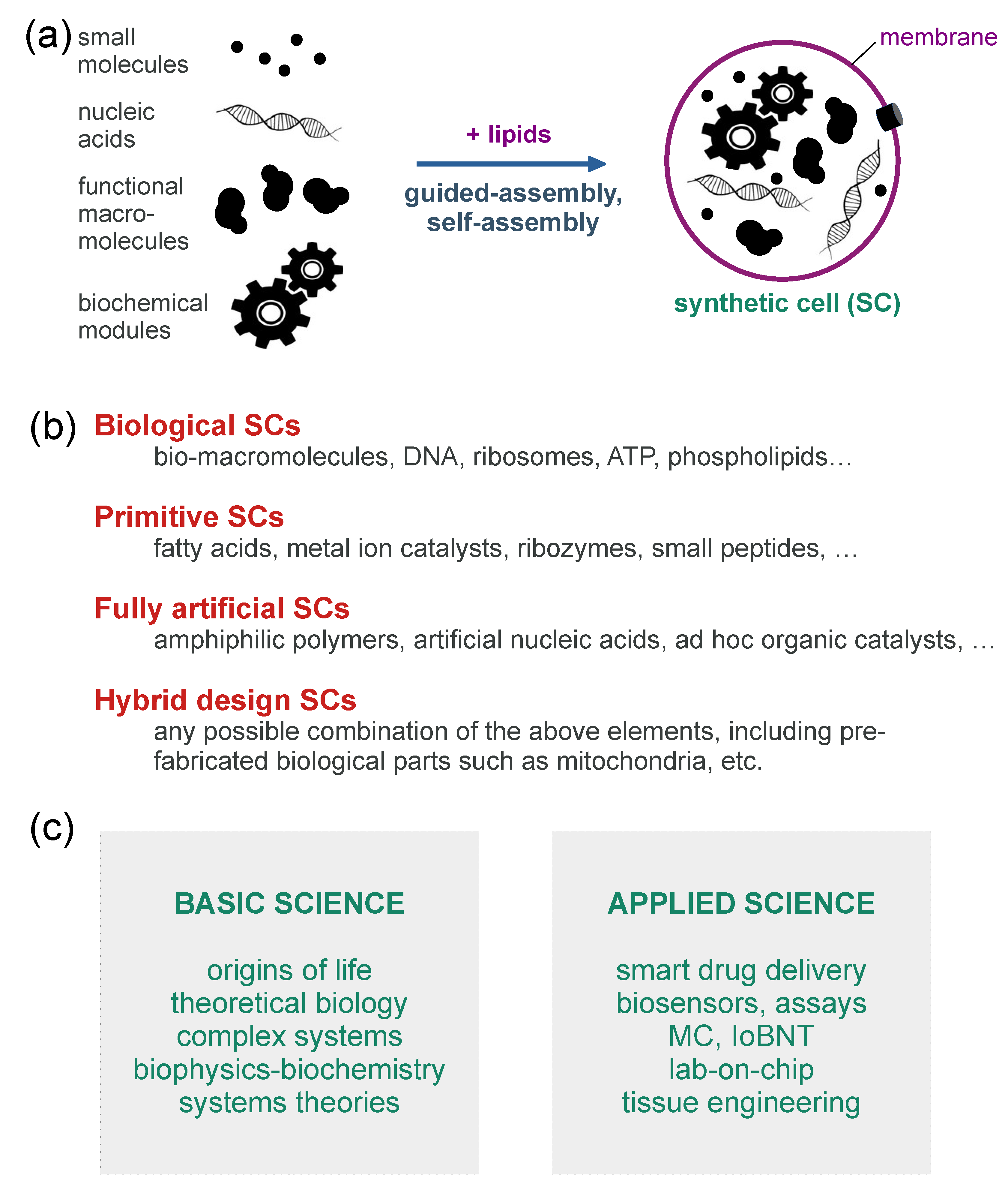
2. Synthetic Cells as Tools for Exploring Artificial Life and Artificial Intelligence
2.1. Recognizing the Twofold Value of Wetware Approaches
2.2. Machine-like SCs
2.3. Organism-like SCs
2.4. Between Machine-like and Organism-like SCs
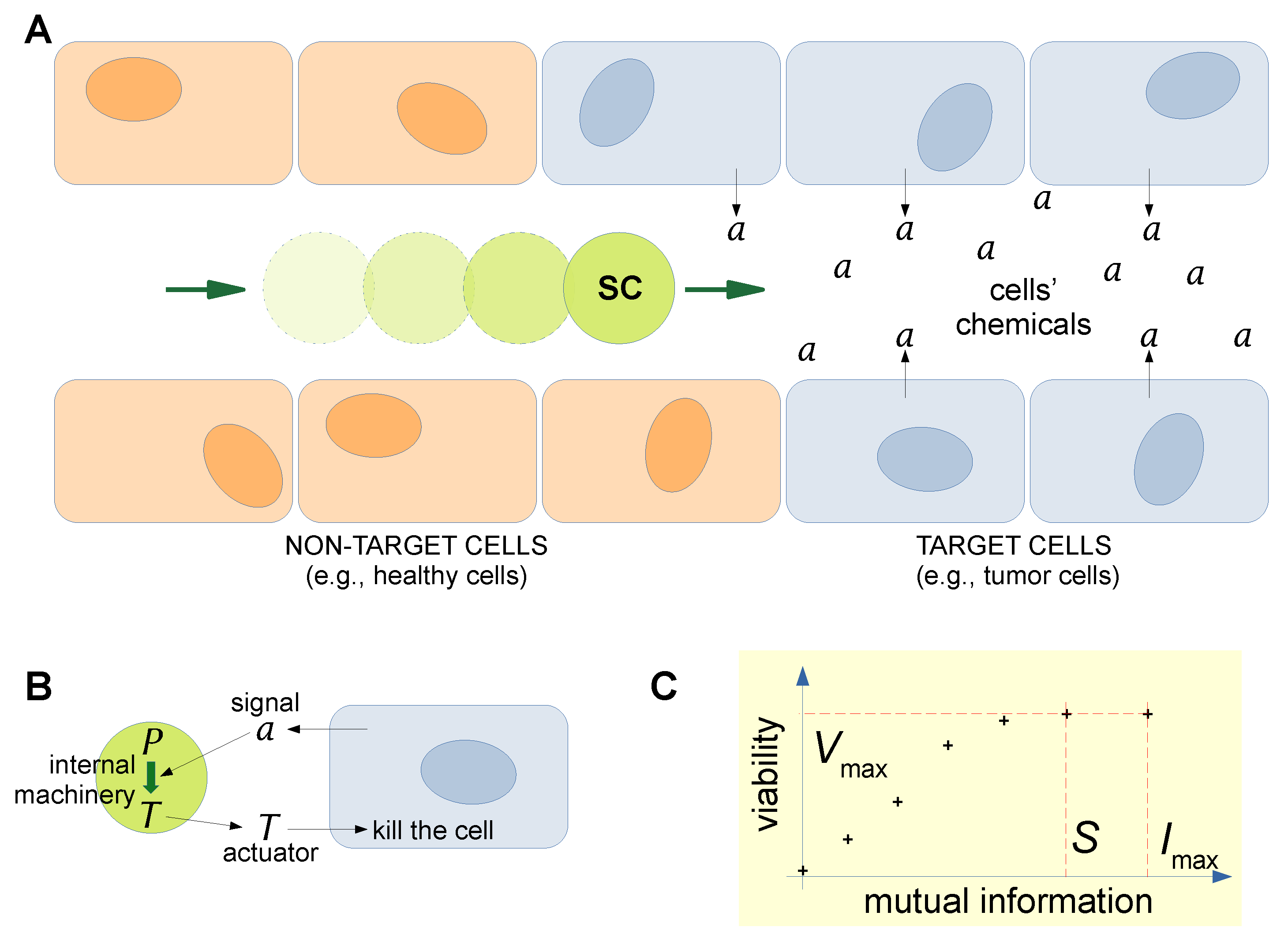
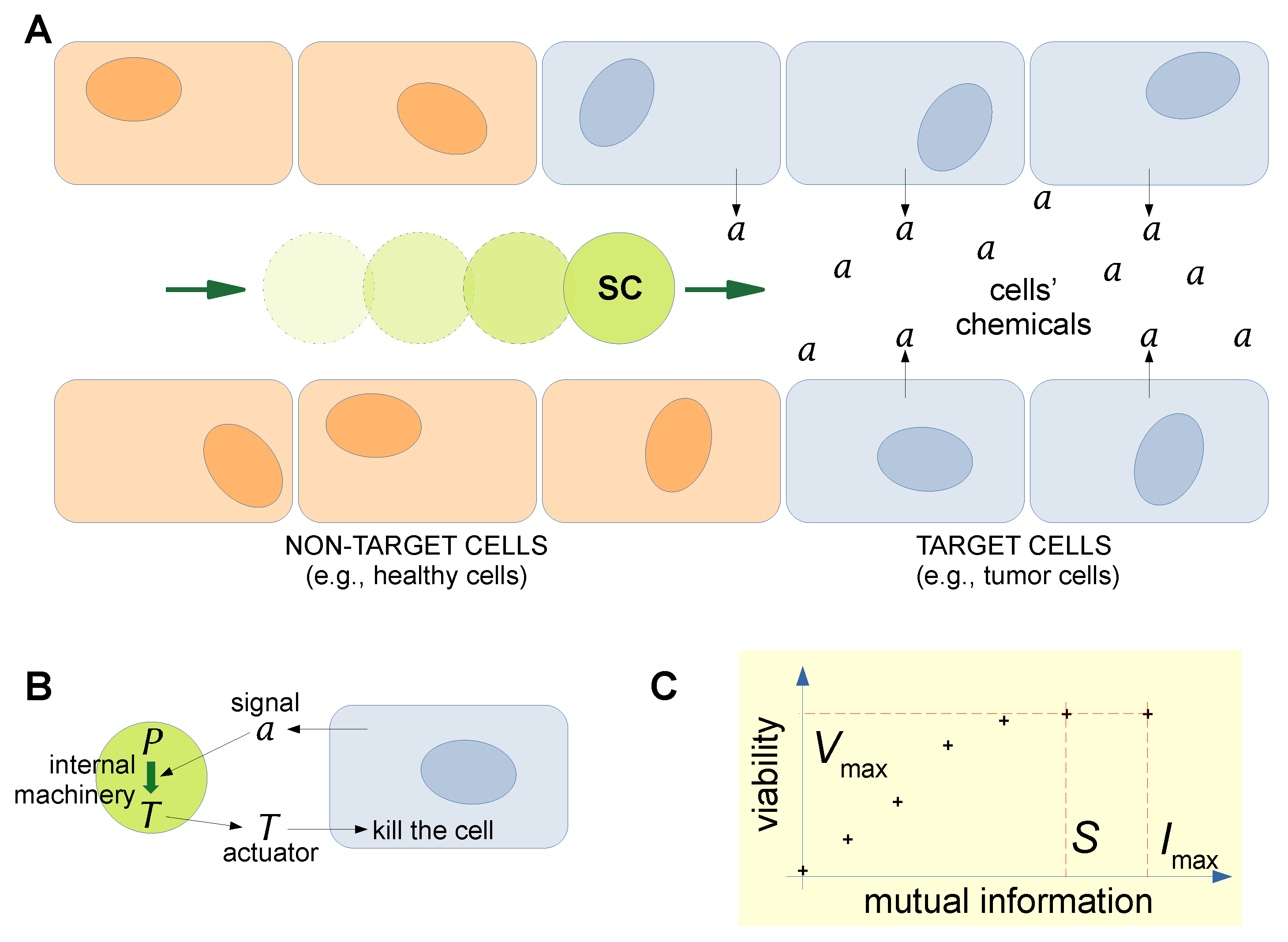
3. Estimating Semantic Information in Situated Synthetic Cells: A Case Study
3.1. The Kolchinsky–Wolpert Approach
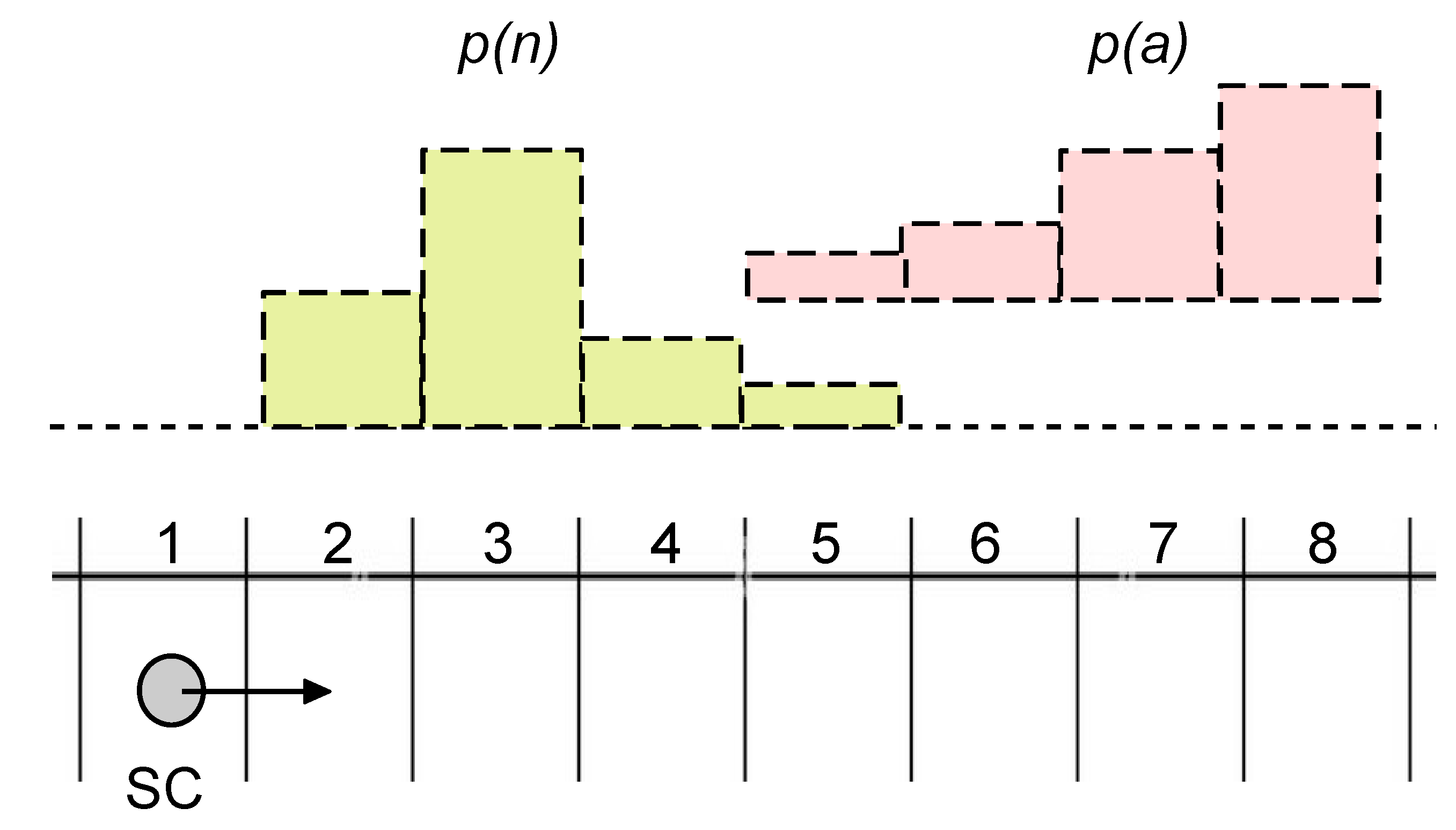
3.2. Stored and Observed Semantic Information Calculated for a Situated SC
3.3. Towards the Understanding of Semantic Information and Meaning for SCs
3.4. Can SCs Generate Meaning from the Interaction with Their Environment?
“[a] cognitive structure […] selects, and retroactively alters, the stimuli to which it is sensitive. By this combination of choice and feed-back, the organic structure determines (in a way moulds) its own specific environment; and the environment in turn brings the cognitive organization to its full development. The system and the environment make one another: cognition according to Maturana and Varela is a process of co-emergence” (italics in the original text) [90].
4. Concluding Remarks
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AI | Artificial Intelligence |
| AL | Artificial Life |
| E | Environment |
| I | Mutual Information |
| IoBNT | Internet of Bio-Nano Things |
| KW | Kolchinsky–Wolpert |
| MC | Molecular Communication |
| Region of Viability | |
| S | System |
| S | Semantic Information (Figure 3C) |
| SB | Synthetic Biology |
| SC | Synthetic Cell |
| S-E | Super-system formed by the union of S and E |
| Transfer Entropy | |
| TX-TL | Transcription-translation |
| V | Viability; value of the viability function |
| Organization |
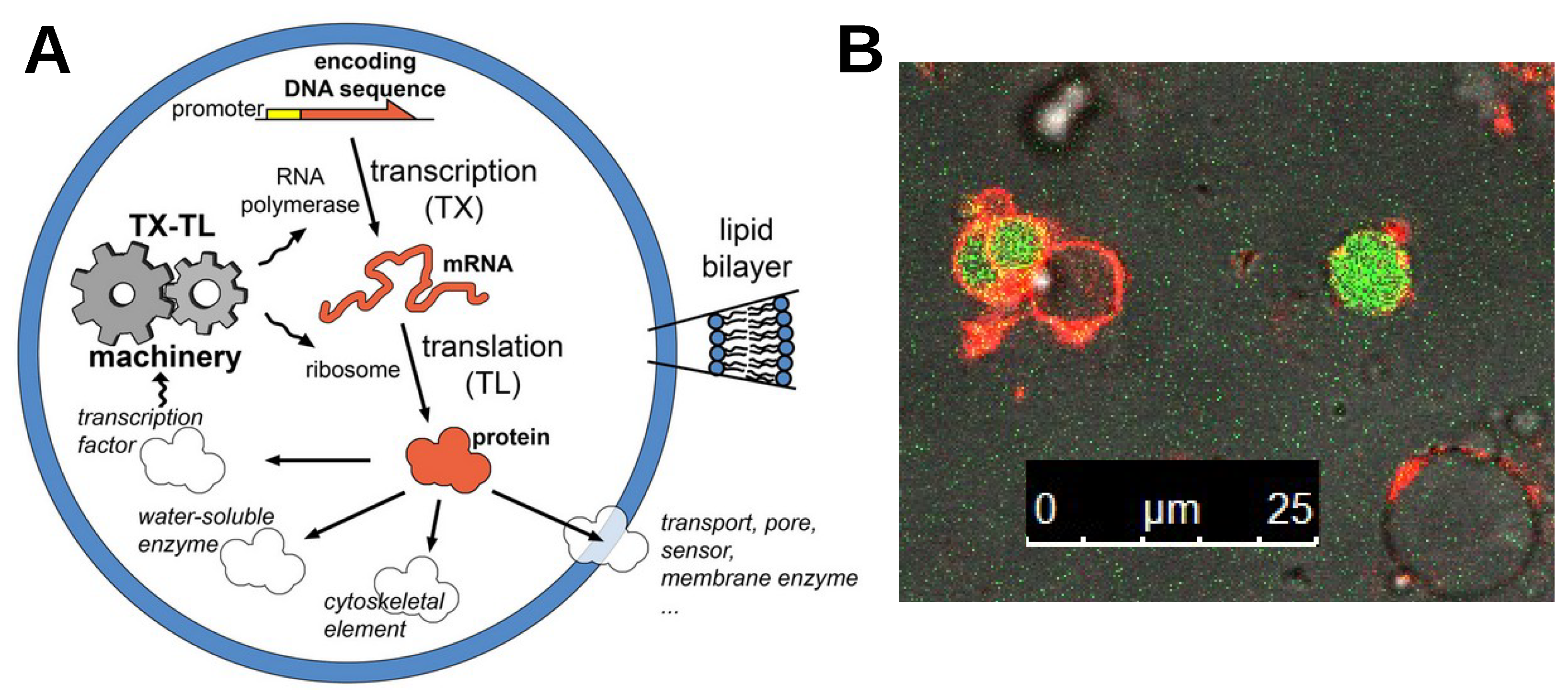
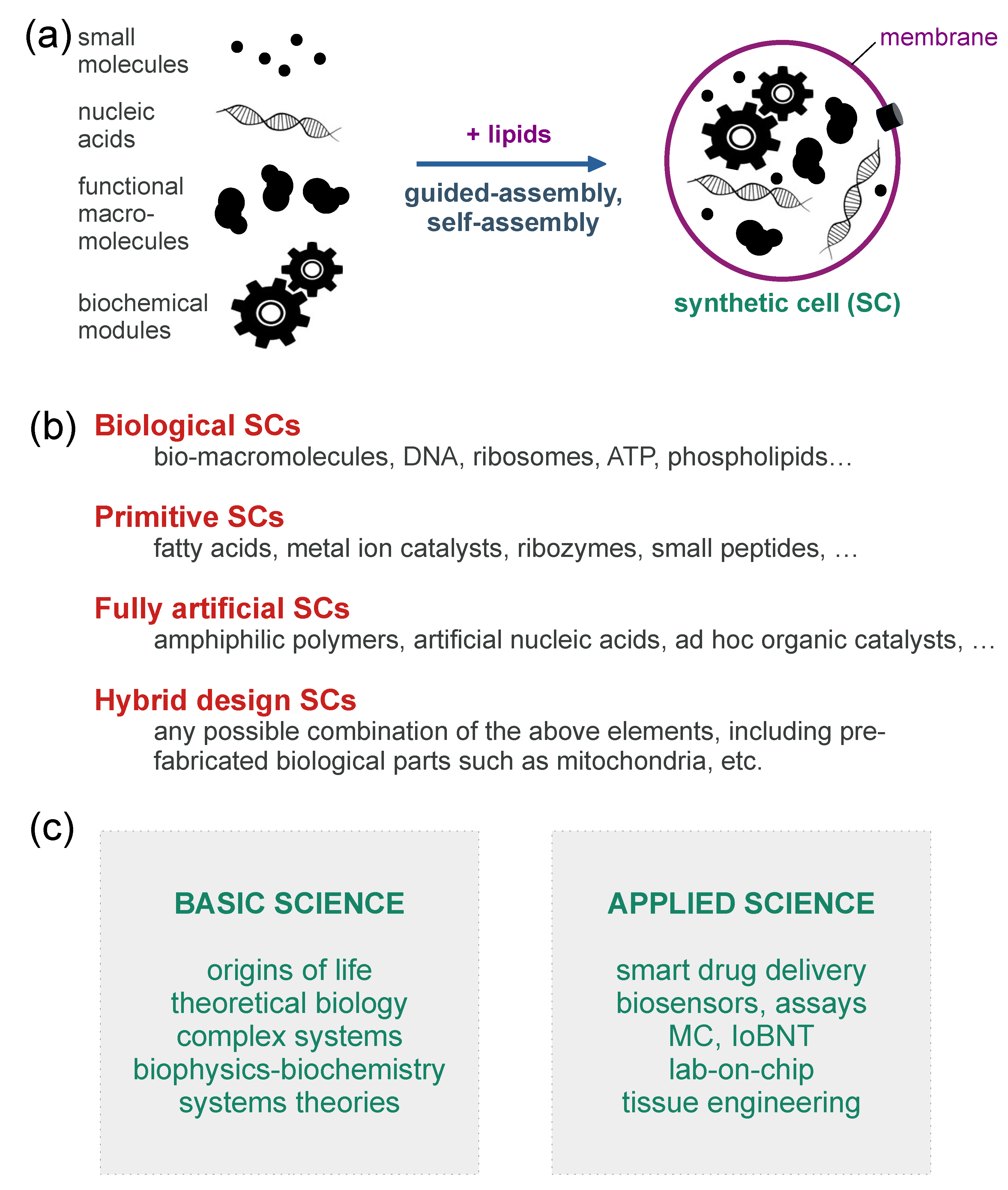
Appendix A. More Information on Bottom-Up Synthetic Cells
Appendix A.1. Pioneering Works on Bottom-Up Synthetic Cells
Appendix A.2. Synthetic Cells Based on the Encapsulation of Biomacromolecules inside Giant Vesicles: A Popular Model

Appendix A.3. A Specific Example of Communicating SC: Bidirectional Communication between Synthetic Cells and Bacteria [25]

References
- Langton, C.G. (Ed.) Artificial Life: Proceedings of an Interdisciplinary Workshop on the Synthesis and Simulation of Living Systems; Addison-Wesley Longman Publishing Co.: Boston, MA, USA, 1989. [Google Scholar]
- Langton, C.G. Artificial life. In The Philosophy of Artifcial Life; Boden, M.A., Ed.; Oxford University Press: Oxford, UK, 1996; pp. 39–94. [Google Scholar]
- Bedau, M.A. Artificial life: Organization, adaptation and complexity from the bottom up. Trends Cogn. Sci. 2003, 7, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Maturana, H.R.; Varela, F.J. De Máquinas y Seres Vivos: Una Teoría Sobre la Organizacíon Biológica; Editorial Universitaria: Santiago, Chile, 1973. [Google Scholar]
- Maturana, H.R.; Varela, F.J. Autopoiesis and Cognition: The Realization of the Living, 1st ed.; D. Reidel Publishing Company: Dordrecht, The Netherlands, 1980. [Google Scholar]
- Varela, F.J. Principles of Biological Autonomy; The North-Holland Series in General Systems Research; Elsevier North-Holland, Inc.: New York, NY, USA, 1979. [Google Scholar]
- Cordeschi, R. The Discovery of the Artificial. Behavior, Mind and Machines Before and Beyond Cybernetics; Studies in Cognitive Systems; Springer: Dordrecht, The Netherlands, 2002. [Google Scholar]
- Cariani, P. To evolve an ear. Epistemological implications of Gordon Pask’s electrochemical devices. Syst. Res. 1993, 10, 19–33. [Google Scholar] [CrossRef]
- Harnad, S. The symbol-grounding problem. Phys. D Nonlinear Phenom. 1990, 42, 335–346. [Google Scholar] [CrossRef]
- Floridi, L. The Philosophy of Information; Oxford University Press: Oxford, UK, 2011. [Google Scholar]
- Hoffmann, P.M. Life’s Ratchet. How Molecular Machines Extract Order from Chaos, 1st ed.; Basic Books. A Member of the Perseus Books Group: New York, NY, USA, 2012. [Google Scholar]
- Phillips, R.; Quake, S.R. The Biological Frontier of Physics. Phys. Today 2006, 59, 38–43. [Google Scholar] [CrossRef]
- Stano, P.; Gentili, P.L.; Damiano, L.; Magarini, M. A Role for Bottom-Up Synthetic Cells in the Internet of Bio-Nano Things? Molecules 2023, 28, 5564. [Google Scholar] [CrossRef]
- Stankiewicz, J.; Eckardt, L.H. Chembiogenesis 2005 and Systems Chemistry Workshop. Angew. Chem. Int. Edit. 2006, 45, 342–344. [Google Scholar] [CrossRef]
- Ruiz-Mirazo, K.; Briones, C.; de la Escosura, A. Prebiotic systems chemistry: New perspectives for the origins of life. Chem. Rev. 2014, 114, 285–366. [Google Scholar] [CrossRef]
- Ashkenasy, G.; Hermans, T.M.; Otto, S.; Taylor, A.F. Systems chemistry. Chem. Soc. Rev. 2017, 46, 2543–2554. [Google Scholar] [CrossRef]
- Ludlow, R.F.; Otto, S. Systems chemistry. Chem. Soc. Rev. 2007, 37, 101–108. [Google Scholar] [CrossRef]
- Čejková, J.; Cartwright, J.H.E. Chemobrionics and Systems Chemistry. ChemSystemsChem 2022, 4, e202200002. [Google Scholar] [CrossRef]
- Luisi, P.L. Toward the engineering of minimal living cells. Anat. Rec. 2002, 268, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Stano, P. Is Research on “Synthetic Cells” Moving to the Next Level? Life 2019, 9, 3. [Google Scholar] [CrossRef] [PubMed]
- Stano, P. Commentary: Rapid and facile preparation of giant vesicles by the droplet transfer method for artificial cell construction. Front. Bioeng. Biotechnol. 2022, 10, 1037809. [Google Scholar] [CrossRef] [PubMed]
- Stano, P. Exploring information and communication theories for synthetic cell research. Front. Bioeng. Biotech. 2022, 10, 927156. [Google Scholar] [CrossRef] [PubMed]
- Stano, P. A four-track perspective for bottom-up synthetic cells. Front. Bioeng. Biotechnol. 2022, 10, 1029446. [Google Scholar] [CrossRef]
- Kolchinsky, A.; Wolpert, D.H. Semantic information, autonomous agency and non-equilibrium statistical physics. Interface Focus 2018, 8, 20180041. [Google Scholar] [CrossRef]
- Lentini, R.; Martín, N.Y.; Forlin, M.; Belmonte, L.; Fontana, J.; Cornella, M.; Martini, L.; Tamburini, S.; Bentley, W.E.; Jousson, O.; et al. Two-Way Chemical Communication between Artificial and Natural Cells. ACS Cent. Sci. 2017, 3, 117–123. [Google Scholar] [CrossRef]
- Altamura, E.; Albanese, P.; Marotta, R.; Milano, F.; Fiore, M.; Trotta, M.; Stano, P.; Mavelli, F. Chromatophores efficiently promote light-driven ATP synthesis and DNA transcription inside hybrid multicompartment artificial cells. Proc. Natl. Acad. Sci. USA 2021, 118, e2012170118. [Google Scholar] [CrossRef]
- Hirschi, S.; Ward, T.R.; Meier, W.P.; Müller, D.J.; Fotiadis, D. Synthetic Biology: Bottom-Up Assembly of Molecular Systems. Chem. Rev. 2022, 122, 16294–16328. [Google Scholar] [CrossRef]
- Eto, S.; Matsumura, R.; Shimane, Y.; Fujimi, M.; Berhanu, S.; Kasama, T.; Kuruma, Y. Phospholipid synthesis inside phospholipid membrane vesicles. Commun. Biol. 2022, 5, 1016. [Google Scholar] [CrossRef]
- Gibson, D.G.; Glass, J.I.; Lartigue, C.; Noskov, V.N.; Chuang, R.Y.; Algire, M.A.; Benders, G.A.; Montague, M.G.; Ma, L.; Moodie, M.M.; et al. Creation of a bacterial cell controlled by a chemically synthesized genome. Science 2010, 329, 52–56. [Google Scholar] [CrossRef]
- Hutchison, C.A.; Chuang, R.Y.; Noskov, V.N.; Assad-Garcia, N.; Deerinck, T.J.; Ellisman, M.H.; Gill, J.; Kannan, K.; Karas, B.J.; Ma, L.; et al. Design and synthesis of a minimal bacterial genome. Science 2016, 351, aad6253. [Google Scholar] [CrossRef]
- Rosenblueth, A.; Wiener, N.; Bigelow, J. Behavior, Purpose and Teleology. Philos. Sci. 1943, 10, 18–24. [Google Scholar] [CrossRef]
- MacKay, D.M. In search of basic symbols. In Proceedings of the 8th Conference on Cybernetics, New York, NY, USA, 15–16 March 1951; von Foerster, H., Ed.; Josiah Macy Jr. Foundation: New York, NY, USA, 1951; pp. 181–221. [Google Scholar]
- Damiano, L.; Stano, P. Synthetic Biology and Artificial Intelligence. Grounding a cross-disciplinary approach to the synthetic exploration of (embodied) cognition. Complex Syst. 2018, 27, 199–228. [Google Scholar] [CrossRef]
- Damiano, L.; Stano, P. Explorative Synthetic Biology in AI. Criteria of relevance and a taxonomy for synthetic models of living and cognitive processes. Artif. Life 2023, 29, 367–387. [Google Scholar] [CrossRef] [PubMed]
- Ashby, W.R. An Introduction to Cybernetics; Chapman & Hall Ltd.: London, UK, 1956. [Google Scholar]
- Ashby, W.R. Design for a Brain, 2nd ed.; John Wiley & Sons Inc.: New York, NY, USA, 1960. [Google Scholar]
- Turing, A.M.I. Computing Machinery and Intelligence. Mind 1950, LIX, 433–460. [Google Scholar] [CrossRef]
- Cronin, L.; Krasnogor, N.; Davis, B.G.; Alexander, C.; Robertson, N.; Steinke, J.H.G.; Schroeder, S.L.M.; Khlobystov, A.N.; Cooper, G.; Gardner, P.M.; et al. The imitation game–a computational chemical approach to recognizing life. Nat. Biotechnol. 2006, 24, 1203–1206. [Google Scholar] [CrossRef] [PubMed]
- Damiano, L.; Stano, P. On the "Life-Likeness" of Synthetic Cells. Front. Bioeng. Biotechnol. 2020, 8, 953. [Google Scholar] [CrossRef] [PubMed]
- Pattee, H.H. Cell Psychology: An Evolutionary Approach to the Symbol-Matter Problem. In LAWS, LANGUAGE and LIFE: Howard Pattee’s Classic Papers on the Physics of Symbols with Contemporary Commentary; Pattee, H.H., Rączaszek-Leonardi, J., Eds.; Biosemiotics, Springer: Dordrecht, The Netherlands, 2012; pp. 165–179. [Google Scholar] [CrossRef]
- Damiano, L.; Stano, P. A Wetware Embodied AI? Towards an Autopoietic Organizational Approach Grounded in Synthetic Biology. Front. Bioeng. Biotechnol. 2021, 9, 873. [Google Scholar] [CrossRef]
- Stano, P.; Damiano, L. Synthetic cell research: Is technical progress leaving theoretical and epistemological investigations one step behind? Front. Robot. AI 2023, 10, 1143196. [Google Scholar] [CrossRef]
- Di Paolo, E.A. Organismically inspired robotics: Homeostatic adaptation and teleology beyond the closed sensorimotor loop. In Dynamical Systems Approach to Embodiment and Sociality; Murase, K., Asakura, T., Eds.; Advanced Knowledge International: Adelaide, Australia, 2003; pp. 19–42. [Google Scholar]
- Froese, T.; Ziemke, T. Enactive artificial intelligence: Investigating the systemic organization of life and mind. Artif. Intell. 2009, 173, 466–500. [Google Scholar] [CrossRef]
- Krinsky, N.; Kaduri, M.; Zinger, A.; Shainsky-Roitman, J.; Goldfeder, M.; Benhar, I.; Hershkovitz, D.; Schroeder, A. Synthetic Cells Synthesize Therapeutic Proteins inside Tumors. Adv. Healthc. Mater. 2018, 7, e1701163. [Google Scholar] [CrossRef] [PubMed]
- Diltemiz, S.E.; Tavafoghi, M.; Barros, N.R.d.; Kanada, M.; Heinämäki, J.; Contag, C.; Seidlits, S.K.; Ashammakhi, N. Use of artificial cells as drug carriers. Mater. Chem. Front. 2021, 5, 6672–6692. [Google Scholar] [CrossRef]
- Sato, W.; Zajkowski, T.; Moser, F.; Adamala, K.P. Synthetic cells in biomedical applications. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2022, 14, e1761. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Levin, R.; Landau, S.; Kaduri, M.; Adir, O.; Ianovici, I.; Krinsky, N.; Doppelt-Flikshtain, O.; Shklover, J.; Shainsky-Roitman, J.; et al. Implanted synthetic cells trigger tissue angiogenesis through de novo production of recombinant growth factors. Proc. Natl. Acad. Sci. USA 2022, 119, e2207525119. [Google Scholar] [CrossRef]
- Boyd, M.A.; Thavarajah, W.; Lucks, J.B.; Kamat, N.P. Robust and tunable performance of a cell-free biosensor encapsulated in lipid vesicles. Sci. Adv. 2023, 9, eadd6605. [Google Scholar] [CrossRef]
- Akyildiz, I.F.; Pierobon, M.; Balasubramaniam, S.; Koucheryavy, Y. The internet of Bio-Nano things. IEEE Commun. Mag. 2015, 53, 32–40. [Google Scholar] [CrossRef]
- Kusku, M.; Unluturk, B.D. Internet of Bio-Nano Things: A Review of Applications, Enabling Technologies and Key Challenges. ITU J. Future Evol. Technol. 2021, 2, 1–24. [Google Scholar]
- Nakano, T.; Eckford, A.W.; Haraguchi, T. Molecular Communication; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Nakano, T. Molecular Communication: A 10 Year Retrospective. IEEE Trans. Mol. Biol. Multi-Scale Commun. 2017, 3, 71–78. [Google Scholar] [CrossRef]
- Nauta, D. The Meaning of Information; Approaches to Semiotics [AS}, Mouton (De Grutyer): The Hague, The Netherlands, 1972. [Google Scholar]
- Gentili, P.L. Small steps towards the development of chemical artificial intelligent systems. RSC Adv. 2013, 3, 25523–25549. [Google Scholar] [CrossRef]
- Bray, D. Protein molecules as computational elements in living cells. Nature 1995, 376, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Bray, D. Wetware. A Computer in Every Living Cells; Yale University Press: New Haven, CT, USA; London, UK, 2009. [Google Scholar]
- Hellingwerf, K.J.; Postma, P.W.; Tommassen, J.; Westerhoff, H.V. Signal transduction in bacteria: Phospho-neural network(s) in Escherichia coli? FEMS Microbiol. Rev. 1995, 16, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Moorman, A.; Samaniego, C.C.; Maley, C.; Weiss, R. A Dynamical Biomolecular Neural Network. In Proceedings of the 2019 IEEE 58th Conference on Decision and Control (CDC), Nice, France, 11–13 December 2019; pp. 1797–1802. [Google Scholar] [CrossRef]
- van der Linden, A.J.; Pieters, P.A.; Bartelds, M.W.; Nathalia, B.L.; Yin, P.; Huck, W.T.S.; Kim, J.; de Greef, T.F.A. DNA Input Classification by a Riboregulator-Based Cell-Free Perceptron. ACS Synth. Biol. 2022, 11, 1510–1520. [Google Scholar] [CrossRef] [PubMed]
- Lakin, M.R.; Stefanovic, D. Supervised Learning in Adaptive DNA Strand Displacement Networks. ACS Synth. Biol. 2016, 5, 885–897. [Google Scholar] [CrossRef] [PubMed]
- Gentili, P.L.; Stano, P. Chemical Neural Networks Inside Synthetic Cells? A Proposal for Their Realization and Modeling. Front. Bioeng. Biotechnol. 2022, 10, 927110. [Google Scholar] [CrossRef]
- Stano, P. Chemical Neural Networks and Synthetic Cell Biotechnology: Preludes to Chemical AI. In Proceedings of the Computational Intelligence Methods for Bioinformatics and Biostatistics; Lecture Notes in Computer Science. Chicco, D., Facchiano, A., Tavazzi, E., Longato, E., Vettoretti, M., Bernasconi, A., Avesani, S., Cazzaniga, P., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 1–12. [Google Scholar] [CrossRef]
- Stano, P. Sketching How Synthetic Cells Can Function as a Platform to Investigate Chemical AI and Information Theories in the Wetware Domain. In Proceedings of the Fourth International Conference on Communication, Computing and Electronics Systems, Coimbatore, India, 15–16 September 2022; Lecture Notes in Electrical Engineering. Bindhu, V., Tavares, J.M.R.S., Vuppalapati, C., Eds.; Springer Nature: Singapore, 2023; pp. 571–584. [Google Scholar] [CrossRef]
- Stano, P. Two Possible AI-Related Paths for Bottom-Up Synthetic Cell Research. In Proceedings of the Artificial Life and Evolutionary Computation, Winterthur, Switzerland, 15–17 September 2021; Communications in Computer and Information Science. Schneider, J.J., Weyland, M.S., Flumini, D., Füchslin, R.M., Eds.; Springer Nature Switzerland: Cham, Switzerland, 2022; pp. 97–100. [Google Scholar] [CrossRef]
- Stano, P.; Gentili, P.L.; Rampioni, G.; Roli, A.; Damiano, L. En Route for Implanting a Minimal Chemical Perceptron into Artificial Cells. In Proceedings of the ALIFE 2022: The 2022 Conference on Artificial Life, Online Conference, 18–22 July 2022; Holler, S., Löffler, R., Bartlett, S., Eds.; MIT Press: Cambridge, MA, USA, 2022; pp. 465–467. [Google Scholar] [CrossRef]
- Braccini, M.; Collinson, E.; Roli, A.; Fellermann, H.; Stano, P. Recurrent neural networks in synthetic cells: A route to autonomous molecular agents? Front. Bioeng. Biotechnol. 2023, 11, 1210334. [Google Scholar] [CrossRef]
- Peruzzi, J.A.; Galvez, N.R.; Kamat, N.P. Engineering transmembrane signal transduction in synthetic membranes using two-component systems. Proc. Natl. Acad. Sci. USA 2023, 120, e2218610120. [Google Scholar] [CrossRef]
- Roli, A.; Kauffman, S.A. Emergence of Organisms. Entropy 2020, 22, 1163. [Google Scholar] [CrossRef]
- Bertschinger, N.; Olbrich, E.; Ay, N.; Jost, J. Autonomy: An information theoretic perspective. Biosystems 2008, 91, 331–345. [Google Scholar] [CrossRef]
- Varela, F.J.; Thompson, E.T.; Rosch, E. The Embodied Mind: Cognitive Science and Human Experience, Revised Edition; The MIT Press: Cambridge, MA, USA, 1992. [Google Scholar]
- Gibson, J.J. The Senses Considered as Perceptual Systems; Houghton Mifflin: Boston, MA, USA, 1966. [Google Scholar]
- von Uexküll, J. Theoretical Biology; Harcourt, Brace & Co.: New York, NY, USA, 1926. [Google Scholar]
- MacKay, D.M. Towards an information-flow model of cerebral organization. Adv. Sci. 1956, 12, 392–395. [Google Scholar]
- Dayan, P.; Hinton, G.E.; Neal, R.M.; Zemel, R.S. The Helmholtz machine. Neural Comput. 1995, 7, 889–904. [Google Scholar] [CrossRef] [PubMed]
- Gentili, P.L. Establishing a New Link between Fuzzy Logic, Neuroscience, and Quantum Mechanics through Bayesian Probability: Perspectives in Artificial Intelligence and Unconventional Computing. Molecules 2021, 26, 5987. [Google Scholar] [CrossRef] [PubMed]
- Valdés-Zorrilla, A.; Letelier, J.C.; Soto-Andrade, J. Metaphorising and bayesian inference according to biology of cognition and enaction. ResearchGate, 2023; preprint id 368923680. [Google Scholar]
- Friston, K.; Kilner, J.; Harrison, L. A free energy principle for the brain. J. Physiol. Paris 2006, 100, 70–87. [Google Scholar] [CrossRef] [PubMed]
- Pezzulo, G.; Parr, T.; Cisek, P.; Clark, A.; Friston, K. Generating Meaning: Active Inference and Generative AI. PsyArXiv 2023. [Google Scholar] [CrossRef]
- Guidoni, A. Verso il Robot Sapiens, 1st ed.; Collana Scienza, Edizioni Controluce: Monte Compatri, Italy, 2018. [Google Scholar]
- MacKay, D.M. Information, Mechanism and Meaning; MIT Press: Cambridge, MA, USA, 1969. [Google Scholar]
- Atlan, H. Sources of Information in Biological Systems. IFAC Proc. Vol. 1977, 10, 177–184. [Google Scholar] [CrossRef]
- Atlan, H. Self Creation of Meaning. Phys. Scr. 1987, 36, 563–576. [Google Scholar] [CrossRef]
- Fields, C.; Levin, M. How Do Living Systems Create Meaning? Philosophies 2020, 5, 36. [Google Scholar] [CrossRef]
- Kiverstein, J.; Kirchhoff, M.D.; Froese, T. The Problem of Meaning: The Free Energy Principle and Artificial Agency. Front. Neurorobot. 2022, 16, 844773. [Google Scholar] [CrossRef]
- Bedau, M.A.; McCaskill, J.S.; Packard, N.H.; Rasmussen, S. Living technology: Exploiting life’s principles in technology. Artif. Life 2010, 16, 89–97. [Google Scholar] [CrossRef]
- Amos, M.; Dittrich, P.; McCaskill, J.; Rasmussen, S. Biological and Chemical Information Technologies. Procedia Comput. Sci. 2011, 7, 56–60. [Google Scholar] [CrossRef]
- Altamura, E.; Carrara, P.; D’Angelo, F.; Mavelli, F.; Stano, P. Extrinsic stochastic factors (solute partition) in gene expression inside lipid vesicles and lipid-stabilized water-in-oil droplets: A review. Synth. Biol. 2018, 3, ysy011. [Google Scholar] [CrossRef] [PubMed]
- Egan, M.; Kuscu, M.; Barros, M.T.; Booth, M.; Llopis-Lorente, A.; Magarini, M.; Martins, D.P.; Schäfer, M.; Stano, P. Toward Interdisciplinary Synergies in Molecular Communications: Perspectives from Synthetic Biology, Nanotechnology, Communications Engineering and Philosophy of Science. Life 2023, 13, 208. [Google Scholar] [CrossRef] [PubMed]
- Bitbol, M.; Luisi, P.L. Autopoiesis with or without cognition: Defining life at its edge. J. R. Soc. Interface 2004, 1, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Longo, G.; Montévil, M. Perspectives on Organisms: Biological Time, Symmetries, and Singularities; Lecture Notes in Morphogenesis; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Roli, A.; Jaeger, J.; Kauffman, S.A. How Organisms Come to Know the World: Fundamental Limits on Artificial General Intelligence. Front. Ecol. Evol. 2022, 9, 806283. [Google Scholar] [CrossRef]
- Roli, A.; Kauffman, S.A. The hiatus between organism and machine evolution: Contrasting mixed microbial communities with robots. Biosystems 2022, 222, 104775. [Google Scholar] [CrossRef]
- Magarini, M.; Stano, P. Synthetic cells engaged in molecular communication: An opportunity for modelling Shannon- and semantic-information in the chemical domain. Front. Commun. Netw. 2021, 2, 724597. [Google Scholar] [CrossRef]
- Hayles, N.K. How We Became Posthuman. Virtual Bodies in Cybernetics, Literature, and Informatics; The University of Chicago Press: Chicago, IL, USA; London, UK, 1999. [Google Scholar]
- Newman, J. Some Observations on the Semantics of “Information”. Inf. Syst. Front. 2001, 3, 155–167. [Google Scholar] [CrossRef]
- Logan, R.K. What is information?: Why is it relativistic and what is its relationship to materiality, meaning and organization. Information 2012, 3, 68–91. [Google Scholar] [CrossRef]
- Kline, R.R. The Cybernetics Moment. Or Why We Call Our Age the Information Age; New Studies in American Intellectual and Cultural History; Johns Hopkins University Press: Baltimore, MD, USA, 2015. [Google Scholar]
- Anta, J. Information, meaning and physics: The intellectual evolution of the English School of Information Theory during 1946-1956. Sci. Context 2021, 34, 357–373. [Google Scholar] [CrossRef]
- Carnap, R.; Bar-Hillel, Y. An outline of a theory of semantic information. Brit. J. Phil. Sci. 1953, 4, 147–157. [Google Scholar]
- Reza, F.M. An Introduction to Information Theory; McGraw-Hill Electrical and Electronic Engineering Series; Mc-Graw-Hill: New York, NY, USA, 1961. [Google Scholar]
- Brillouin, L. Science and Information Theory, 2nd ed.; Academic Press, Inc.: New York, NY, USA, 1962. [Google Scholar]
- Ruzzante, B.; Del Moro, L.; Magarini, M.; Stano, P. Synthetic Cells Extract Semantic Information from their Environment. IEEE Trans. Mol. Biol. Multi-Scale Commun. 2023, 9, 23–27. [Google Scholar] [CrossRef]
- Del Moro, L.; Ruzzante, B.; Magarini, M.; Gentili, P.L.; Rampioni, G.; Roli, A.; Damiano, L.; Stano, P. Chemical Neural Networks and Semantic Information Investigated Through Synthetic Cells. In Proceedings of the Artificial Life and Evolutionary Computation, WIVACE 2022, Gaeta, Italy, 14–16 September 2022; Communications in Computer and Information Science. De Stefano, C., Fontanella, F., Vanneschi, L., Eds.; Springer Nature Switzerland: Cham, Switzerland, 2023; Volume 1780, pp. 27–39. [Google Scholar] [CrossRef]
- Del Moro, L.; Magarini, M.; Stano, P. On the Evaluation of Observed Semantic Information in Synthetic Cells. In Proceedings of the Artificial Life and Evolutionary Computation, WIVACE 2023, Venice, Italy, 6–8 September 2023; Communications in Computer and Information Science. Serra, R., Villani, M., Cagnoni, S., Eds.; Springer Nature Switzerland: Cham, Switzerland, 2024. in press. [Google Scholar]
- Mavelli, F.; Marangoni, R.; Stano, P. A Simple Protein Synthesis Model for the PURE System Operation. Bull. Math. Biol. 2015, 77, 1185–1212. [Google Scholar] [CrossRef] [PubMed]
- Carrara, P.; Altamura, E.; D’Angelo, F.; Mavelli, F.; Stano, P. Measurement and Numerical Modeling of Cell-Free Protein Synthesis: Combinatorial Block-Variants of the PURE System. Data 2018, 3, 41. [Google Scholar] [CrossRef]
- Di Paolo, E.A. Autopoiesis, Adaptivity, Teleology, Agency. Phenom. Cogn. Sci. 2005, 4, 429–452. [Google Scholar] [CrossRef]
- Pask, G. An Approach to Cybernetics; Hutchinson & Co., Ltd.: London, UK, 1961. [Google Scholar]
- Endy, D. Foundations for engineering biology. Nature 2005, 438, 449–453. [Google Scholar] [CrossRef]
- Andrianantoandro, E.; Basu, S.; Karig, D.K.; Weiss, R. Synthetic biology: New engineering rules for an emerging discipline. Mol. Syst. Biol. 2006, 2, 2006.0028. [Google Scholar] [CrossRef]
- de Lorenzo, V.; Danchin, A. Synthetic biology: Discovering new worlds and new words. EMBO Rep. 2008, 9, 822–827. [Google Scholar] [CrossRef]
- Hanczyc, M.M. The Early History of Protocells – The search for the recipe of life. In Protocells: Bridging Nonliving and Living Matter; Rasmussen, S., Bedau, M.A., Chen, L., Deamer, D., Krakauer, D.C., Packard, N.H., Eds.; MIT Press: Cambridge MA, USA, 2009; pp. 3–18. [Google Scholar]
- Chang, T.M.S. Semipermeable microcapsules. Science 1964, 146, 524–525. [Google Scholar] [CrossRef]
- Chang, T.M.S. Artificial Cells; Charles C. Thomas: Springfield, IL, USA, 1972. [Google Scholar]
- Luisi, P.L.; Varela, F.J. Self-replicating micelles — A chemical version of a minimal autopoietic system. Orig. Life Evol. Biosph. 1989, 19, 633–643. [Google Scholar] [CrossRef]
- Bachmann, P.; Walde, P.; Luisi, P.; Lang, J. Self-replicating reverse micelles and chemical autopoiesis. J. Am. Chem. Soc. 1990, 112, 8200–8201. [Google Scholar] [CrossRef]
- Schmidli, P.K.; Schurtenberger, P.; Luisi, P.L. Liposome-mediated enzymatic synthesis of phosphatidylcholine as an approach to self-replicating liposomes. J. Am. Chem. Soc. 1991, 113, 8127–8130. [Google Scholar] [CrossRef]
- Walde, P.; Goto, A.; Monnard, P.; Wessicken, M.; Luisi, P. Oparin’s Reactions Revisited: Enzymic Synthesis of Poly(adenylic acid) in Micelles and Self-Reproducing Vesicles. J. Am. Chem. Soc. 1994, 116, 7541–7547. [Google Scholar] [CrossRef]
- Oberholzer, T.; Wick, R.; Luisi, P.L.; Biebricher, C.K. Enzymatic RNA replication in self-reproducing vesicles: An approach to a minimal cell. Biochem. Biophys. Res. Commun. 1995, 207, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Oberholzer, T.; Nierhaus, K.H.; Luisi, P.L. Protein expression in liposomes. Biochem. Biophys. Res. Commun. 1999, 261, 238–241. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, A.C.; Breaker, R.R.; Joyce, G.F.; Deamer, D.W. Production of RNA by a polymerase protein encapsulated within phospholipid vesicles. J. Mol. Evol. 1994, 39, 555–559. [Google Scholar] [CrossRef]
- Chen, I.A.; Salehi-Ashtiani, K.; Szostak, J.W. RNA catalysis in model protocell vesicles. J. Am. Chem. Soc. 2005, 127, 13213–13219. [Google Scholar] [CrossRef]
- Yu, W.; Sato, K.; Wakabayashi, M.; Nakaishi, T.; Ko-Mitamura, E.P.; Shima, Y.; Urabe, I.; Yomo, T. Synthesis of functional protein in liposome. J. Biosci. Bioeng. 2001, 92, 590–593. [Google Scholar] [CrossRef]
- Stano, P. Gene Expression Inside Liposomes: From Early Studies to Current Protocols. Chemistry 2019, 25, 7798–7814. [Google Scholar] [CrossRef]
- Szostak, J.W.; Bartel, D.P.; Luisi, P.L. Synthesizing life. Nature 2001, 409, 387–390. [Google Scholar] [CrossRef]
- Oberholzer, T.; Luisi, P.L. The use of liposomes for constructing cell models. J. Biol. Phys. 2002, 28, 733–744. [Google Scholar] [CrossRef]
- Nomura, S.; Tsumoto, K.; Hamada, T.; Akiyoshi, K.; Nakatani, Y.; Yoshikawa, K. Gene expression within cell-sized lipid vesicles. Chembiochem 2003, 4, 1172–1175. [Google Scholar] [CrossRef] [PubMed]
- Noireaux, V.; Libchaber, A. A vesicle bioreactor as a step toward an artificial cell assembly. Proc. Natl. Acad. Sci. USA 2004, 101, 17669–17674. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, K.; Sato, K.; Shima, Y.; Urabe, I.; Yomo, T. Expression of a cascading genetic network within liposomes. FEBS Lett. 2004, 576, 387–390. [Google Scholar] [CrossRef]
- Altamura, E.; Albanese, P.; Mavelli, F.; Stano, P. The Rise of the Nested Multicompartment Model in Synthetic Cell Research. Front. Mol. Biosci. 2021, 8, 850. [Google Scholar] [CrossRef] [PubMed]
- Di Iorio, D.; Wegner, S.V. Towards applications of synthetic cells in nanotechnology. Curr. Opin. Chem. Biol. 2022, 68, 102145. [Google Scholar] [CrossRef] [PubMed]
- Gaur, D.; Dubey, N.C.; Tripathi, B.P. Biocatalytic self-assembled synthetic vesicles and coacervates: From single compartment to artificial cells. Adv. Colloid Interface Sci. 2022, 299, 102566. [Google Scholar] [CrossRef]
- Guindani, C.; da Silva, L.C.; Cao, S.; Ivanov, T.; Landfester, K. Synthetic Cells: From Simple Bio-Inspired Modules to Sophisticated Integrated Systems. Angew. Chem. Int. Ed. Engl. 2022, 61, e202110855. [Google Scholar] [CrossRef]
- Herianto, S.; Chien, P.J.; Ho, J.A.A.; Tu, H.L. Liposome-based artificial cells: From gene expression to reconstitution of cellular functions and phenotypes. Biomater. Adv. 2022, 142, 213156. [Google Scholar] [CrossRef]
- Jiang, W.; Wu, Z.; Gao, Z.; Wan, M.; Zhou, M.; Mao, C.; Shen, J. Artificial Cells: Past, Present and Future. ACS Nano 2022, 16, 15705–15733. [Google Scholar] [CrossRef]
- Karoui, H.; Patwal, P.S.; Pavan Kumar, B.V.V.S.; Martin, N. Chemical Communication in Artificial Cells: Basic Concepts, Design and Challenges. Front. Mol. Biosci. 2022, 9, 880525. [Google Scholar] [CrossRef]
- Lu, Y.; Allegri, G.; Huskens, J. Vesicle-based artificial cells: Materials, construction methods and applications. Mater. Horiz. 2022, 9, 892–907. [Google Scholar] [CrossRef] [PubMed]
- Albanese, P.; Mavelli, F.; Altamura, E. Light energy transduction in liposome-based artificial cells. Front. Bioeng. Biotechnol. 2023, 11, 1161730. [Google Scholar] [CrossRef] [PubMed]
- Bailoni, E.; Partipilo, M.; Coenradij, J.; Grundel, D.A.J.; Slotboom, D.J.; Poolman, B. Minimal Out-of-Equilibrium Metabolism for Synthetic Cells: A Membrane Perspective. ACS Synth. Biol. 2023, 12, 922–946. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.Y.; Ariyama, H.; Huck, W.T.S.; Deng, N.N. Division in synthetic cells. Chem. Soc. Rev. 2023, 52, 3307–3325. [Google Scholar] [CrossRef] [PubMed]
- Fasciano, S.; Wang, S. Recent advances of droplet-based microfluidics for engineering artificial cells. SLAS Technol. 2023, in press. [CrossRef]
- Jahnke, K.; Göpfrich, K. Engineering DNA-based cytoskeletons for synthetic cells. Interface Focus 2023, 13, 20230028. [Google Scholar] [CrossRef]
- Lin, Z.; Beneyton, T.; Baret, J.C.; Martin, N. Coacervate Droplets for Synthetic Cells. Small Methods 2023, e2300496. [Google Scholar] [CrossRef]
- Lin, A.J.; Sihorwala, A.Z.; Belardi, B. Engineering Tissue-Scale Properties with Synthetic Cells: Forging One from Many. ACS Synth. Biol. 2023, 12, 1889–1907. [Google Scholar] [CrossRef]
- Partipilo, M.; Claassens, N.J.; Slotboom, D.J. A Hitchhiker’s Guide to Supplying Enzymatic Reducing Power into Synthetic Cells. ACS Synth. Biol. 2023, 12, 947–962. [Google Scholar] [CrossRef]
- Takinoue, M. DNA droplets for intelligent and dynamical artificial cells: From the viewpoint of computation and non-equilibrium systems. Interface Focus 2023, 13, 20230021. [Google Scholar] [CrossRef]
- van de Cauter, L.; van Buren, L.; Koenderink, G.H.; Ganzinger, K.A. Exploring Giant Unilamellar Vesicle Production for Artificial Cells - Current Challenges and Future Directions. Small Methods 2023, e2300416. [Google Scholar] [CrossRef]
- Yu, X.; Mukwaya, V.; Mann, S.; Dou, H. Signal Transduction in Artificial Cells. Small Methods 2023, e2300231. [Google Scholar] [CrossRef]
- Walde, P.; Cosentino, K.; Engel, H.; Stano, P. Giant vesicles: Preparations and applications. Chembiochem 2010, 11, 848–865. [Google Scholar] [CrossRef] [PubMed]
- Dimova, R.; Stano, P.; Marques, C.M.; Walde, P. Preparation methods for giant unilamellar vesicles. In The Giant Vesicle Book; Dimova, R., Marques, C.M., Eds.; Taylor & Francis Group: Boca Raton, FL, USA, 2020; pp. 3–20. [Google Scholar]
- Pautot, S.; Frisken, B.J.; Weitz, D.A. Production of unilamellar vesicles using an inverted emulsion. Langmuir 2003, 19, 2870–2879. [Google Scholar] [CrossRef]
- Fujii, S.; Matsuura, T.; Sunami, T.; Nishikawa, T.; Kazuta, Y.; Yomo, T. Liposome display for in vitro selection and evolution of membrane proteins. Nat. Protoc. 2014, 9, 1578–1591. [Google Scholar] [CrossRef] [PubMed]
- Gardner, P.M.; Winzer, K.; Davis, B.G. Sugar synthesis in a protocellular model leads to a cell signalling response in bacteria. Nat. Chem. 2009, 1, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Stano, P.; Rampioni, G.; Carrara, P.; Damiano, L.; Leoni, L.; Luisi, P.L. Semi-synthetic minimal cells as a tool for biochemical ICT. BioSystems 2012, 109, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.M.S. ARTIFICIAL CELL evolves into nanomedicine, biotherapeutics, blood substitutes, drug delivery, enzyme/gene therapy, cancer therapy, cell/stem cell therapy, nanoparticles, liposomes, bioencapsulation, replicating synthetic cells, cell encapsulation/scaffold, biosorbent/immunosorbent haemoperfusion/plasmapheresis, regenerative medicine, encapsulated microbe, nanobiotechnology, nanotechnology. Artif. Cells Nanomed. Biotechnol. 2019, 47, 997–1013. [Google Scholar] [CrossRef]
- LeDuc, P.R.; Wong, M.S.; Ferreira, P.M.; Groff, R.E.; Haslinger, K.; Koonce, M.P.; Lee, W.Y.; Love, J.C.; McCammon, J.A.; Monteiro-Riviere, N.A.; et al. Towards an in vivo biologically inspired nanofactory. Nat. Nanotechnol. 2007, 2, 3–7. [Google Scholar] [CrossRef]
- Ding, Y.; Contreras-Llano, L.E.; Morris, E.; Mao, M.; Tan, C. Minimizing Context Dependency of Gene Networks Using Artificial Cells. ACS Appl. Mater. Interfaces 2018, 10, 30137–30146. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, Y.; Li, M.; Xiong, L.; Zhang, Z.; Yang, X.; He, X.; Wang, K.; Liu, J.; Mann, S. Enzyme-mediated nitric oxide production in vasoactive erythrocyte membrane-enclosed coacervate protocells. Nat. Chem. 2020, 12, 1165–1173. [Google Scholar] [CrossRef] [PubMed]
- Toparlak, D.; Zasso, J.; Bridi, S.; Serra, M.D.; Macchi, P.; Conti, L.; Baudet, M.L.; Mansy, S.S. Artificial cells drive neural differentiation. Sci. Adv. 2020, 6, eabb4920. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stano, P. Chemical Systems for Wetware Artificial Life: Selected Perspectives in Synthetic Cell Research. Int. J. Mol. Sci. 2023, 24, 14138. https://doi.org/10.3390/ijms241814138
Stano P. Chemical Systems for Wetware Artificial Life: Selected Perspectives in Synthetic Cell Research. International Journal of Molecular Sciences. 2023; 24(18):14138. https://doi.org/10.3390/ijms241814138
Chicago/Turabian StyleStano, Pasquale. 2023. "Chemical Systems for Wetware Artificial Life: Selected Perspectives in Synthetic Cell Research" International Journal of Molecular Sciences 24, no. 18: 14138. https://doi.org/10.3390/ijms241814138
APA StyleStano, P. (2023). Chemical Systems for Wetware Artificial Life: Selected Perspectives in Synthetic Cell Research. International Journal of Molecular Sciences, 24(18), 14138. https://doi.org/10.3390/ijms241814138