
Amphibian-Derived Natural Anticancer Peptides and Proteins: Mechanism of Action, Application Strategies, and Prospects

Abstract
:1. Introduction
2. Physicochemical Properties of Natural Anticancer Peptides and Proteins of Amphibian Origin
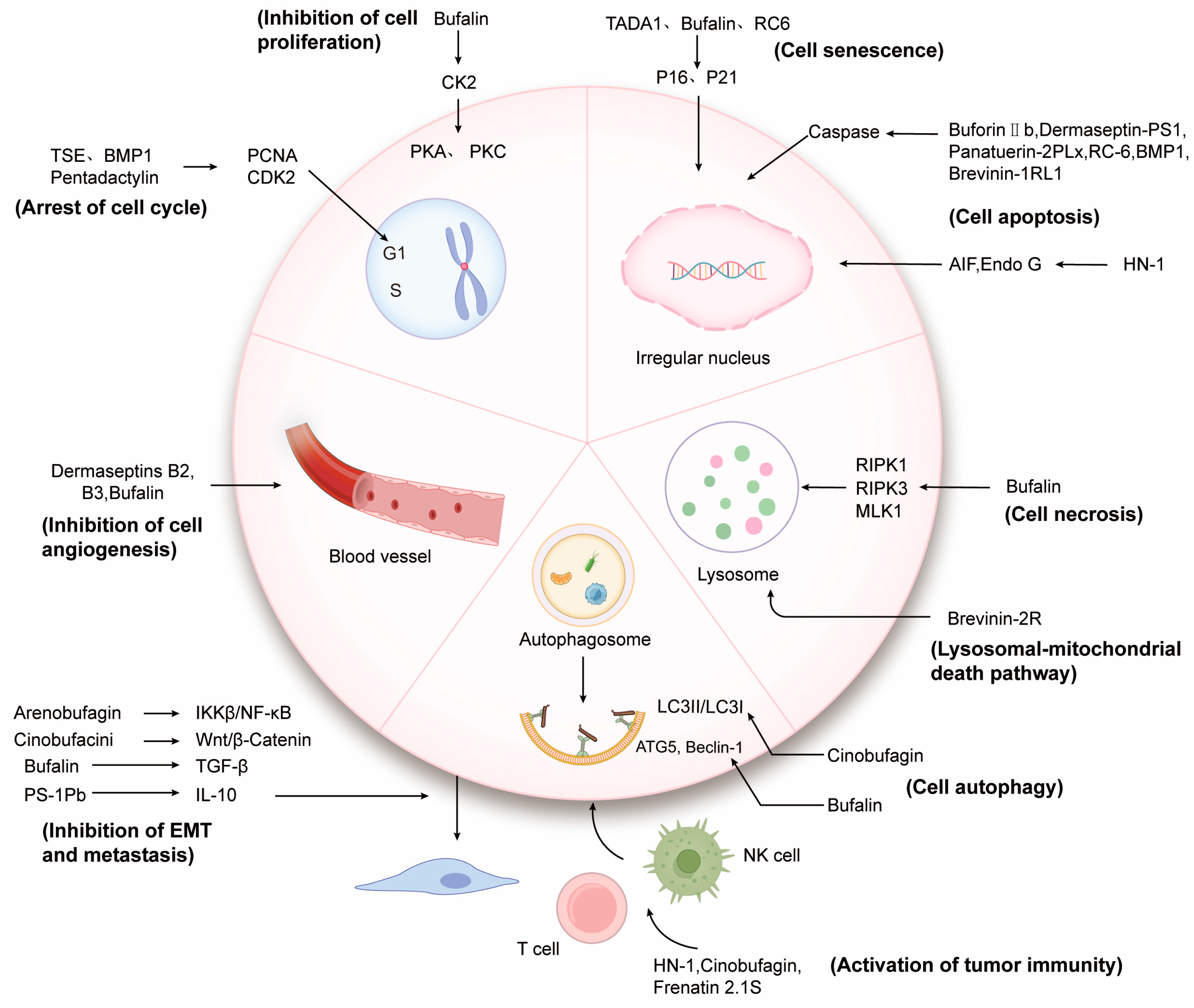
3. Action Mode and Mechanism of Anticancer Peptides and Proteins
3.1. Interaction Directly through the Cell Membrane
3.2. Acting on Tumor Growth
3.2.1. Regulation of Tumor Cell Cycle and Proliferation
3.2.2. Inducing Tumor Cells Senescence
3.2.3. Inducing Apoptosis of Tumor Cells
3.2.4. Inducing Tumor Cells Autophagy
3.2.5. Inducing Tumor Cells Necrosis
3.2.6. Inducing Other Forms of Tumor Cells Death
3.3. Inhibiting Tumor Angiogenesis
3.4. Inhibiting Epithelial Mesenchymal Transition and Metastasis of Tumor Cells
3.5. Activating Tumor Immunity
3.6. Other Possible Ways to Relate to Oxidative Stress
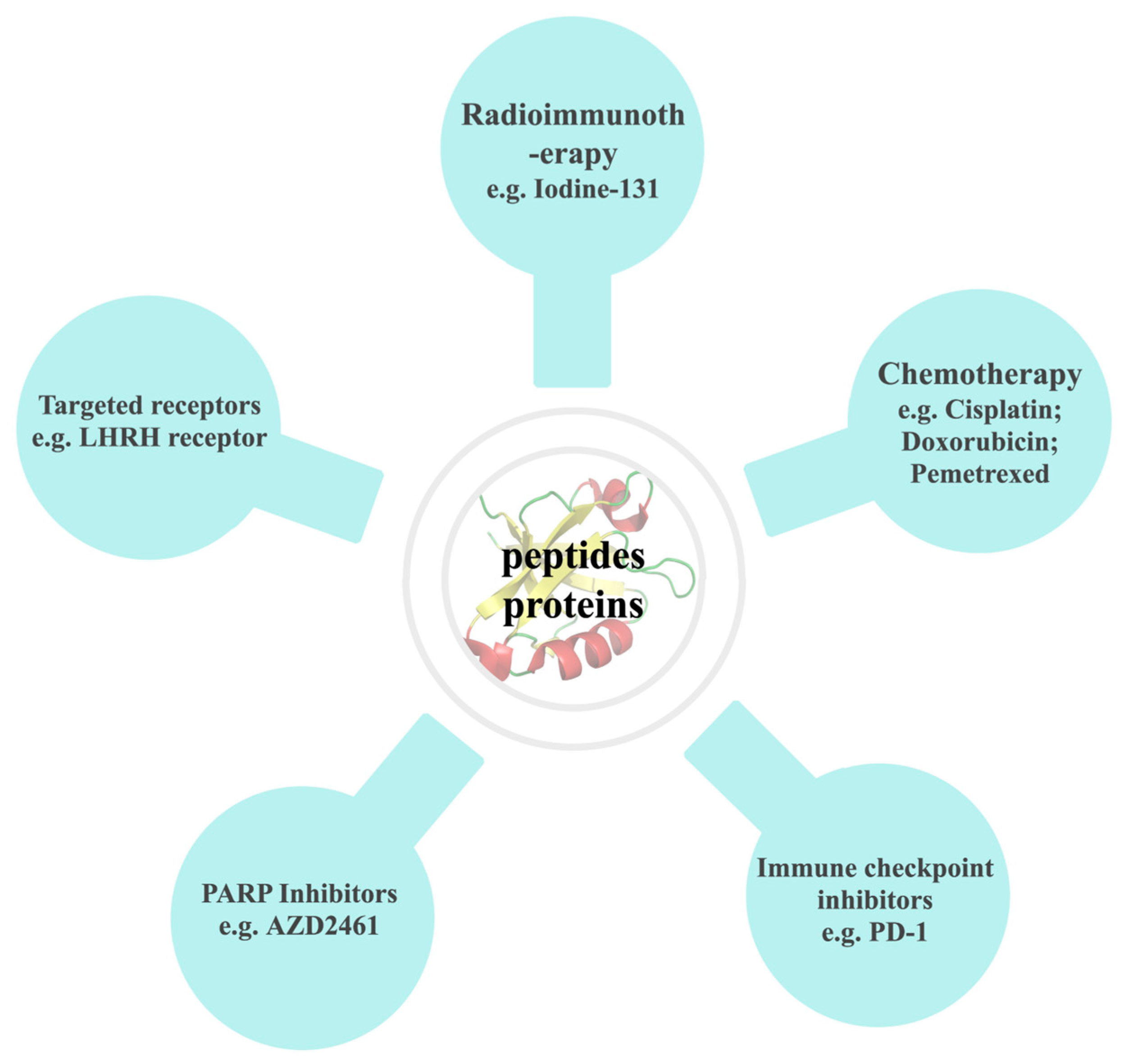
4. Application Strategies of Natural Anticancer Peptides and Proteins from Amphibians
4.1. Structural Modification
4.2. Combined with Other Drugs
4.2.1. Combined with Radioimmunotherapy
4.2.2. Combined with Chemotherapy
4.2.3. Combined with PD-1 Immune Checkpoint Inhibitors
4.2.4. Combined with PARP Inhibitors
4.2.5. Binding to Tumor Cell Targeted Receptors
5. Prospects for the Development of Natural Anticancer Peptides and Proteins from Amphibians
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Soerjomataram, I.; Bray, F. Planning for tomorrow: Global cancer incidence and the role of prevention 2020–2070. Nat. Rev. Clin. Oncol. 2021, 18, 663–672. [Google Scholar] [CrossRef]
- Miller, K.D.; Nogueira, L.; Mariotto, A.B.; Rowland, J.H.; Yabroff, K.R.; Alfano, C.M.; Jemal, A.; Kramer, J.L.; Siegel, R.L. Cancer treatment and survivorship statistics, 2019. CA A Cancer J. Clin. 2019, 69, 363–385. [Google Scholar] [CrossRef]
- Schirrmacher, V. From chemotherapy to biological therapy: A review of novel concepts to reduce the side effects of systemic cancer treatment (Review). Int. J. Oncol. 2019, 54, 407–419. [Google Scholar] [CrossRef]
- Kim, C.; Kim, B. Anti-Cancer Natural Products and Their Bioactive Compounds Inducing ER Stress-Mediated Apoptosis: A Review. Nutrients 2018, 10, 1021. [Google Scholar] [CrossRef]
- Bade, R.; Chan, H.F.; Reynisson, J. Characteristics of known drug space. Natural products, their derivatives and synthetic drugs. Eur. J. Med. Chem. 2010, 45, 5646–5652. [Google Scholar] [CrossRef]
- Conlon, J.M.; Woodhams, D.C.; Raza, H.; Coquet, L.; Leprince, J.; Jouenne, T.; Vaudry, H.; Rollins-Smith, L.A. Peptides with differential cytolytic activity from skin secretions of the lemur leaf frog Hylomantis lemur (Hylidae: Phyllomedusinae). Toxicon 2007, 50, 498–506. [Google Scholar] [CrossRef]
- Varga, J.F.A.; Bui-Marinos, M.P.; Katzenback, B.A. Frog Skin Innate Immune Defences: Sensing and Surviving Pathogens. Front. Immunol. 2018, 9, 3128. [Google Scholar] [CrossRef]
- Ni, G.; Chen, S.; Chen, M.; Wu, J.; Yang, B.; Yuan, J.; Walton, S.F.; Li, H.; Wei, M.Q.; Wang, Y.; et al. Host-Defense Peptides Caerin 1.1 and 1.9 Stimulate TNF-Alpha-Dependent Apoptotic Signals in Human Cervical Cancer HeLa Cells. Front. Cell Dev. Biol. 2020, 8, 676. [Google Scholar] [CrossRef]
- Bhattacharjee, P.; Giri, B.; Gomes, A. Apoptogenic activity and toxicity studies of a cytotoxic protein (BMP1) from the aqueous extract of common Indian toad (Bufo melanostictus Schneider) skin. Toxicon 2011, 57, 225–236. [Google Scholar] [CrossRef]
- Dong, Z.; Hu, H.; Yu, X.; Tan, L.; Ma, C.; Xi, X.; Li, L.; Wang, L.; Zhou, M.; Chen, T.; et al. Novel Frog Skin-Derived Peptide Dermaseptin-PP for Lung Cancer Treatment: In vitro/vivo Evaluation and Anti-tumor Mechanisms Study. Front. Chem. 2020, 8, 476. [Google Scholar] [CrossRef]
- Kariya, Y.; Tatsuta, T.; Sugawara, S.; Kariya, Y.; Nitta, K.; Hosono, M. RNase activity of sialic acid-binding lectin from bullfrog eggs drives antitumor effect via the activation of p38 MAPK to caspase-3/7 signaling pathway in human breast cancer cells. Int. J. Oncol. 2016, 49, 1334–1342. [Google Scholar] [CrossRef]
- Bobone, S.; Stella, L. Selectivity of Antimicrobial Peptides: A Complex Interplay of Multiple Equilibria. Adv. Exp. Med. Biol. 2019, 1117, 175–214. [Google Scholar]
- Conlon, J.M.; Mechkarska, M.; Lukic, M.L.; Flatt, P.R. Potential therapeutic applications of multifunctional host-defense peptides from frog skin as anti-cancer, anti-viral, immunomodulatory, and anti-diabetic agents. Peptides 2014, 57, 67–77. [Google Scholar] [CrossRef]
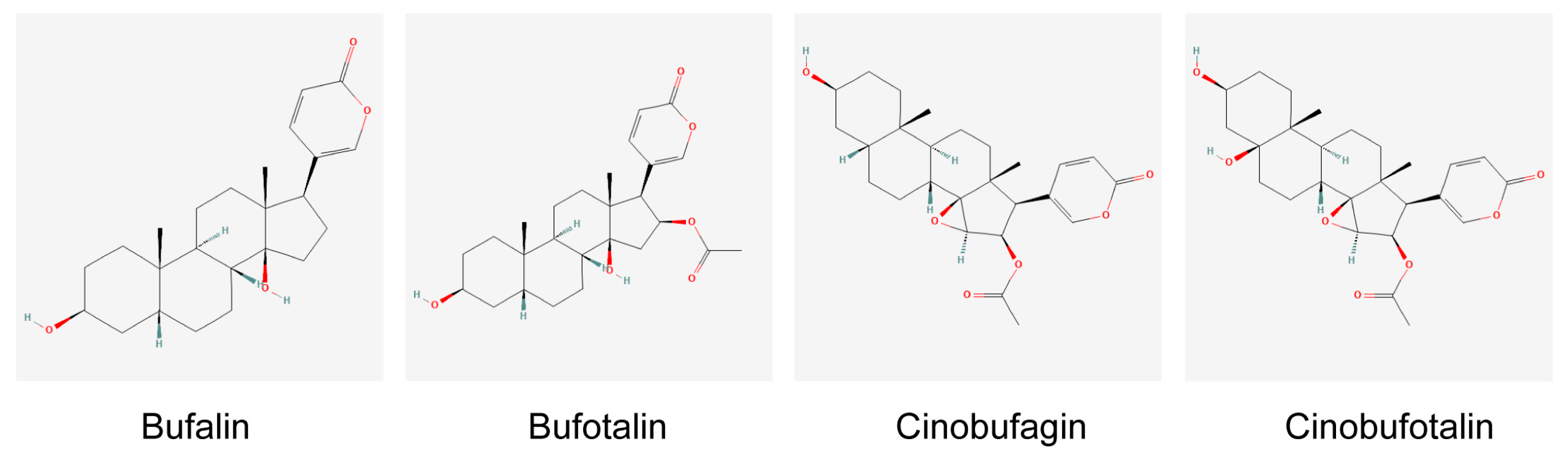
- Jia, J.; Li, J.; Zheng, Q.; Li, D. A research update on the antitumor effects of active components of Chinese medicine ChanSu. Front. Oncol. 2022, 12, 1014637. [Google Scholar] [CrossRef]
- Rozek, T.; Wegener, K.L.; Bowie, J.H.; Olver, I.N.; Carver, J.A.; Wallace, J.C.; Tyler, M.J. The antibiotic and anticancer active aurein peptides from the Australian Bell Frogs Litoria aurea and Litoria raniformis the solution structure of aurein 1.2. Eur. J. Biochem. 2000, 267, 5330–5341. [Google Scholar] [CrossRef]
- Lai, R.; Zheng, Y.T.; Shen, J.H.; Liu, G.J.; Liu, H.; Lee, W.H.; Tang, S.Z.; Zhang, Y. Antimicrobial peptides from skin secretions of Chinese red belly toad Bombina maxima. Peptides 2002, 23, 427–435. [Google Scholar] [CrossRef]
- Swithenbank, L.; Cox, P.; Harris, L.G.; Dudley, E.; Sinclair, K.; Lewis, P.; Cappiello, F.; Morgan, C. Temporin A and Bombinin H2 Antimicrobial Peptides Exhibit Selective Cytotoxicity to Lung Cancer Cells. Scientifica 2020, 2020, 3526286. [Google Scholar] [CrossRef]
- Mignogna, G.; Simmaco, M.; Kreil, G.; Barra, D. Antibacterial and haemolytic peptides containing D-alloisoleucine from the skin of Bombina variegata. EMBO J. 1993, 12, 4829–4832. [Google Scholar] [CrossRef]
- Simmaco, M.; Mignogna, G.; Canofeni, S.; Miele, R.; Mangoni, M.L.; Barra, D. Temporins, antimicrobial peptides from the European red frog Rana temporaria. Eur. J. Biochem. 1996, 242, 788–792. [Google Scholar] [CrossRef]
- Rinaldi, A.C.; Mangoni, M.L.; Rufo, A.; Luzi, C.; Barra, D.; Zhao, H.; Kinnunen, P.K.; Bozzi, A.; Di Giulio, A.; Simmaco, M. Temporin L: Antimicrobial, haemolytic and cytotoxic activities, and effects on membrane permeabilization in lipid vesicles. Biochem. J. 2002, 368 Pt 1, 91–100. [Google Scholar] [CrossRef]
- Conlon, J.M.; Al-Kharrge, R.; Ahmed, E.; Raza, H.; Galadari, S.; Condamine, E. Effect of aminoisobutyric acid (Aib) substitutions on the antimicrobial and cytolytic activities of the frog skin peptide, temporin-1DRa. Peptides 2007, 28, 2075–2080. [Google Scholar] [CrossRef]
- Sang, M.; Wu, Q.; Xi, X.; Ma, C.; Wang, L.; Zhou, M.; Burrows, J.F.; Chen, T. Identification and target-modifications of temporin-PE: A novel antimicrobial peptide in the defensive skin secretions of the edible frog, Pelophylax kl. esculentus. Biochem. Biophys. Res. Commun. 2018, 495, 2539–2546. [Google Scholar] [CrossRef]
- Mechkarska, M.; Attoub, S.; Sulaiman, S.; Pantic, J.; Lukic, M.L.; Conlon, J.M. Anti-cancer, immunoregulatory, and antimicrobial activities of the frog skin host-defense peptides pseudhymenochirin-1Pb and pseudhymenochirin-2Pa. Regul. Pept. 2014, 194–195, 69–76. [Google Scholar] [CrossRef]
- van Zoggel, H.; Hamma-Kourbali, Y.; Galanth, C.; Ladram, A.; Nicolas, P.; Courty, J.; Amiche, M.; Delbé, J. Antitumor and angiostatic peptides from frog skin secretions. Amino Acids 2012, 42, 385–395. [Google Scholar] [CrossRef]
- Charpentier, S.; Amiche, M.; Mester, J.; Vouille, V.; Le Caer, J.P.; Nicolas, P.; Delfour, A. Structure, synthesis, and molecular cloning of dermaseptins B, a family of skin peptide antibiotics. J. Biol. Chem. 1998, 273, 14690–14697. [Google Scholar] [CrossRef]
- Zhu, H.; Ding, X.; Li, W.; Lu, T.; Ma, C.; Xi, X.; Wang, L.; Zhou, M.; Burden, R.; Chen, T. Discovery of two skin-derived dermaseptins and design of a TAT-fusion analogue with broad-spectrum antimicrobial activity and low cytotoxicity on healthy cells. PeerJ 2018, 6, e5635. [Google Scholar] [CrossRef]
- Shi, D.; Hou, X.; Wang, L.; Gao, Y.; Wu, D.; Xi, X.; Zhou, M.; Kwok, H.F.; Duan, J.; Chen, T.; et al. Two Novel Dermaseptin-Like Antimicrobial Peptides with Anticancer Activities from the Skin Secretion of Pachymedusa dacnicolor. Toxins 2016, 8, 144. [Google Scholar] [CrossRef]
- Huang, L.; Chen, D.; Wang, L.; Lin, C.; Ma, C.; Xi, X.; Chen, T.; Shaw, C.; Zhou, M. Dermaseptin-PH: A Novel Peptide with Antimicrobial and Anticancer Activities from the Skin Secretion of the South American Orange-Legged Leaf Frog, Pithecopus (Phyllomedusa) hypochondrialis. Molecules 2017, 22, 1805. [Google Scholar] [CrossRef]
- Chen, D.; Zhou, X.; Chen, X.; Huang, L.; Xi, X.; Ma, C.; Zhou, M.; Wang, L.; Chen, T. Evaluating the Bioactivity of a Novel Antimicrobial and Anticancer Peptide, Dermaseptin-PS4(Der-PS4), from the Skin Secretion of Phyllomedusa sauvagii. Molecules 2019, 24, 2974. [Google Scholar] [CrossRef]
- Jiang, Y.; Wu, Y.; Wang, T.; Chen, X.; Zhou, M.; Ma, C.; Xi, X.; Zhang, Y.; Chen, T.; Shaw, C.; et al. Brevinin-1GHd: A novel Hylarana guentheri skin secretion-derived Brevinin-1 type peptide with antimicrobial and anticancer therapeutic potential. Biosci. Rep. 2020, 40, BSR20200019. [Google Scholar] [CrossRef]
- Pei, X.; Gong, Z.; Wu, Q.; Chen, X.; Wang, L.; Ma, C.; Xi, X.; Chen, T.; Shaw, C.; Zhou, M. Characterisation of a novel peptide, Brevinin-1H, from the skin secretion of Amolops hainanensis and rational design of several analogues. Chem. Biol. Drug Des. 2021, 97, 273–282. [Google Scholar] [CrossRef]
- Yao, A.; Ma, Y.; Chen, X.; Zhou, M.; Xi, X.; Ma, C.; Ren, S.; Chen, T.; Shaw, C.; Wang, L. Modification Strategy of D-leucine Residue Addition on a Novel Peptide from Odorrana schmackeri, with Enhanced Bioactivity and In Vivo Efficacy. Toxins 2021, 13, 611. [Google Scholar] [CrossRef]
- Peng, X.; Zhou, C.; Hou, X.; Liu, Y.; Wang, Z.; Peng, X.; Zhang, Z.; Wang, R.; Kong, D. Molecular characterization and bioactivity evaluation of two novel bombinin peptides from the skin secretion of Oriental fire-bellied toad, Bombina orientalis. Amino Acids 2018, 50, 241–253. [Google Scholar] [CrossRef]
- Zhou, C.; Wang, Z.; Peng, X.; Liu, Y.; Lin, Y.; Zhang, Z.; Qiu, Y.; Jin, M.; Wang, R.; Kong, D. Discovery of two bombinin peptides with antimicrobial and anticancer activities from the skin secretion of Oriental fire-bellied toad, Bombina orientalis. Chem. Biol. Drug Des. 2018, 91, 50–61. [Google Scholar] [CrossRef]
- Lyu, P.; Ge, L.; Ma, R.; Wei, R.; McCrudden, C.M.; Chen, T.; Shaw, C.; Kwok, H.F. Identification and pharmaceutical evaluation of novel frog skin-derived serine proteinase inhibitor peptide-PE-BBI (Pelophylax esculentus Bowman-Birk inhibitor) for the potential treatment of cancer. Sci. Rep. 2018, 8, 14502. [Google Scholar] [CrossRef]
- Liu, Y.; Du, Q.; Ma, C.; Xi, X.; Wang, L.; Zhou, M.; Burrows, J.F.; Chen, T.; Wang, H. Structure-activity relationship of an antimicrobial peptide, Phylloseptin-PHa: Balance of hydrophobicity and charge determines the selectivity of bioactivities. Drug Des. Dev. Ther. 2019, 13, 447–458. [Google Scholar] [CrossRef]
- Conlon, J.M.; Mechkarska, M.; Prajeep, M.; Arafat, K.; Zaric, M.; Lukic, M.L.; Attoub, S. Transformation of the naturally occurring frog skin peptide, alyteserin-2a into a potent, non-toxic anti-cancer agent. Amino Acids 2013, 44, 715–723. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, X.; Wu, Y.; Zhou, M.; Ma, C.; Xi, X.; Chen, T.; Walker, B.; Shaw, C.; Wang, L. A Bowman-Birk type chymotrypsin inhibitor peptide from the amphibian, Hylarana erythraea. Sci. Rep. 2018, 8, 5851. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, L.; Ma, C.; Zhang, Y.; Xi, X.; Wang, L.; Zhou, M.; Burrows, J.F.; Chen, T. A novel antimicrobial peptide, Ranatuerin-2PLx, showing therapeutic potential in inhibiting proliferation of cancer cells. Biosci. Rep. 2018, 38, BSR20180710. [Google Scholar] [CrossRef]
- Attoub, S.; Arafat, H.; Mechkarska, M.; Conlon, J.M. Anti-tumor activities of the host-defense peptide hymenochirin-1B. Regul. Pept. 2013, 187, 51–56. [Google Scholar] [CrossRef]
- Attoub, S.; Mechkarska, M.; Sonnevend, A.; Radosavljevic, G.; Jovanovic, I.; Lukic, M.L.; Conlon, J.M. Esculentin-2CHa: A host-defense peptide with differential cytotoxicity against bacteria, erythrocytes and tumor cells. Peptides 2013, 39, 95–102. [Google Scholar] [CrossRef]
- Santana, C.J.C.; Magalhães, A.C.M.; Dos Santos Júnior, A.C.M.; Ricart, C.A.O.; Lima, B.D.; Álvares, A.; Freitas, S.M.; Pires, O.R., Jr.; Fontes, W.; Castro, M.S. Figainin 1, a Novel Amphibian Skin Peptide with Antimicrobial and Antiproliferative Properties. Antibiotics 2020, 9, 625. [Google Scholar] [CrossRef]
- Abdelfatah, S.; Lu, X.; Schmeda-Hirschmann, G.; Efferth, T. Cytotoxicity and antimitotic activity of Rhinella schneideri and Rhinella marina venoms. J. Ethnopharmacol. 2019, 242, 112049. [Google Scholar] [CrossRef]
- Zhang, D.M.; Liu, J.S.; Deng, L.J.; Chen, M.F.; Yiu, A.; Cao, H.H.; Tian, H.Y.; Fung, K.P.; Kurihara, H.; Pan, J.X.; et al. Arenobufagin, a natural bufadienolide from toad venom, induces apoptosis and autophagy in human hepatocellular carcinoma cells through inhibition of PI3K/Akt/mTOR pathway. Carcinogenesis 2013, 34, 1331–1342. [Google Scholar] [CrossRef]
- Wang, W.; Wu, S.; Cen, Z.; Zhang, Y.; Chen, Y.; Huang, Y.; Cillo, A.R.; Prokopec, J.S.; Quarato, G.; Vignali, D.A.A.; et al. Mobilizing phospholipids on tumor plasma membrane implicates phosphatidylserine externalization blockade for cancer immunotherapy. Cell Rep. 2022, 41, 111582. [Google Scholar] [CrossRef]
- Ran, S.; Downes, A.; Thorpe, P.E. Increased exposure of anionic phospholipids on the surface of tumor blood vessels. Cancer Res. 2002, 62, 6132–6140. [Google Scholar]
- Wang, C.; Tian, L.L.; Li, S.; Li, H.B.; Zhou, Y.; Wang, H.; Yang, Q.Z.; Ma, L.J.; Shang, D.J. Rapid cytotoxicity of antimicrobial peptide tempoprin-1CEa in breast cancer cells through membrane destruction and intracellular calcium mechanism. PLoS ONE 2013, 8, e60462. [Google Scholar] [CrossRef]
- Wang, C.; Chen, Y.W.; Zhang, L.; Gong, X.G.; Zhou, Y.; Shang, D.J. Melanoma cell surface-expressed phosphatidylserine as a therapeutic target for cationic anticancer peptide, temporin-1CEa. J. Drug Target. 2016, 24, 548–556. [Google Scholar] [CrossRef]
- Dennison, S.R.; Harris, F.; Phoenix, D.A. Investigations into the potential anticancer activity of Maximin H5. Biochimie 2017, 137, 29–34. [Google Scholar] [CrossRef]
- Doyle, J.; Brinkworth, C.S.; Wegener, K.L.; Carver, J.A.; Llewellyn, L.E.; Olver, I.N.; Bowie, J.H.; Wabnitz, P.A.; Tyler, M.J. nNOS inhibition, antimicrobial and anticancer activity of the amphibian skin peptide, citropin 1.1 and synthetic modifications. The solution structure of a modified citropin 1.1. Eur. J. Biochem. 2003, 270, 1141–1153. [Google Scholar] [CrossRef]
- Lehmann, J.; Retz, M.; Sidhu, S.S.; Suttmann, H.; Sell, M.; Paulsen, F.; Harder, J.; Unteregger, G.; Stöckle, M. Antitumor activity of the antimicrobial peptide magainin II against bladder cancer cell lines. Eur. Urol. 2006, 50, 141–147. [Google Scholar] [CrossRef]
- Conlon, J.M.; Galadari, S.; Raza, H.; Condamine, E. Design of potent, non-toxic antimicrobial agents based upon the naturally occurring frog skin peptides, ascaphin-8 and peptide XT-7. Chem. Biol. Drug Des. 2008, 72, 58–64. [Google Scholar] [CrossRef]
- Li, M.; Xi, X.; Ma, C.; Chen, X.; Zhou, M.; Burrows, J.F.; Chen, T.; Wang, L. A Novel Dermaseptin Isolated from the Skin Secretion of Phyllomedusa tarsius and Its Cationicity-Enhanced Analogue Exhibiting Effective Antimicrobial and Anti-Proliferative Activities. Biomolecules 2019, 9, 628. [Google Scholar] [CrossRef]
- Long, Q.; Li, L.; Wang, H.; Li, M.; Wang, L.; Zhou, M.; Su, Q.; Chen, T.; Wu, Y. Novel peptide dermaseptin-PS1 exhibits anticancer activity via induction of intrinsic apoptosis signalling. J. Cell. Mol. Med. 2019, 23, 1300–1312. [Google Scholar] [CrossRef]
- Li, B.; Lyu, P.; Xie, S.; Qin, H.; Pu, W.; Xu, H.; Chen, T.; Shaw, C.; Ge, L.; Kwok, H.F. LFB: A Novel Antimicrobial Brevinin-Like Peptide from the Skin Secretion of the Fujian Large Headed Frog, Limnonectes fujianensi. Biomolecules 2019, 9, 242. [Google Scholar] [CrossRef]
- Wang, C.; Zhou, Y.; Li, S.; Li, H.; Tian, L.; Wang, H.; Shang, D. Anticancer mechanisms of temporin-1CEa, an amphipathic α-helical antimicrobial peptide, in Bcap-37 human breast cancer cells. Life Sci. 2013, 92, 1004–1014. [Google Scholar] [CrossRef]
- Giri, B.; Gomes, A.; Debnath, A.; Saha, A.; Biswas, A.K.; Dasgupta, S.C.; Gomes, A. Antiproliferative, cytotoxic and apoptogenic activity of Indian toad (Bufo melanostictus, Schneider) skin extract on U937 and K562 cells. Toxicon 2006, 48, 388–400. [Google Scholar] [CrossRef]
- Libério, M.S.; Joanitti, G.A.; Azevedo, R.B.; Cilli, E.M.; Zanotta, L.C.; Nascimento, A.C.; Sousa, M.V.; Pires Júnior, O.R.; Fontes, W.; Castro, M.S. Anti-proliferative and cytotoxic activity of pentadactylin isolated from Leptodactylus labyrinthicus on melanoma cells. Amino Acids 2011, 40, 51–59. [Google Scholar] [CrossRef]
- Gomes, A.; Giri, B.; Alam, A.; Mukherjee, S.; Bhattacharjee, P.; Gomes, A. Anticancer activity of a low immunogenic protein toxin (BMP1) from Indian toad (Bufo melanostictus, Schneider) skin extract. Toxicon 2011, 58, 85–92. [Google Scholar] [CrossRef]
- Collado, M.; Serrano, M. Senescence in tumours: Evidence from mice and humans. Nat. Rev. Cancer 2010, 10, 51–57. [Google Scholar] [CrossRef]
- Harrison, V.; Khan, S.F.; Damerell, V.; Bleloch, J.; ArulJothi, K.N.; Sinkala, M.; Lennard, K.; Mulder, N.; Calder, B.; Blackburn, J.; et al. Strongylopus grayii tadpole blastema extract exerts cytotoxic effects on embryonal rhabdomyosarcoma cells. Vitr. Cell. Dev. Biol. Anim. 2022, 58, 679–692. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, Y.; Melkus, M.W.; Yin, S.; Tang, S.N.; Jiang, P.; Pramanik, K.; Wu, W.; Kim, S.; Ye, M.; et al. Role of P53-Senescence Induction in Suppression of LNCaP Prostate Cancer Growth by Cardiotonic Compound Bufalin. Mol. Cancer Ther. 2018, 17, 2341–2352. [Google Scholar] [CrossRef]
- Ardelt, W.; Ardelt, B.; Darzynkiewicz, Z. Ribonucleases as potential modalities in anticancer therapy. Eur. J. Pharmacol. 2009, 625, 181–189. [Google Scholar] [CrossRef]
- Yiang, G.T.; Tsai, H.F.; Chen, J.R.; Chou, P.L.; Wu, T.K.; Liu, H.C.; Chang, W.J.; Liu, L.C.; Tseng, H.H.; Yu, Y.L. RC-6 ribonuclease induces caspase activation, cellular senescence and neuron-like morphology in NT2 embryonal carcinoma cells. Oncol. Rep. 2014, 31, 1738–1744. [Google Scholar] [CrossRef]
- Obeng, E. Apoptosis (programmed cell death) and its signals—A review. Braz. J. Biol. 2021, 81, 1133–1143. [Google Scholar] [CrossRef]
- Lee, H.S.; Park, C.B.; Kim, J.M.; Jang, S.A.; Park, I.Y.; Kim, M.S.; Cho, J.H.; Kim, S.C. Mechanism of anticancer activity of buforin IIb, a histone H2A-derived peptide. Cancer Lett. 2008, 271, 47–55. [Google Scholar] [CrossRef]
- Ju, X.; Fan, D.; Kong, L.; Yang, Q.; Zhu, Y.; Zhang, S.; Su, G.; Li, Y. Antimicrobial Peptide Brevinin-1RL1 from Frog Skin Secretion Induces Apoptosis and Necrosis of Tumor Cells. Molecules 2021, 26, 2059. [Google Scholar] [CrossRef]
- Giampazolias, E.; Tait, S.W.G. Caspase-independent cell death: An anti-cancer double whammy. Cell Cycle 2018, 17, 269–270. [Google Scholar] [CrossRef]
- Qiao, X.; Yang, H.; Gao, J.; Cai, S.; Shi, N.; Wang, M.; Wang, Y.; Yu, H. A small cytotoxic peptide from frog elicits potent antitumor immunity to prevent local tumor growth and metastases. Future Med. Chem. 2019, 11, 2505–2525. [Google Scholar] [CrossRef]
- Ichimiya, T.; Yamakawa, T.; Hirano, T.; Yokoyama, Y.; Hayashi, Y.; Hirayama, D.; Wagatsuma, K.; Itoi, T.; Nakase, H. Autophagy and Autophagy-Related Diseases: A Review. Int. J. Mol. Sci. 2020, 21, 8974. [Google Scholar] [CrossRef]
- Hamurcu, Z.; Delibaşı, N.; Nalbantoglu, U.; Sener, E.F.; Nurdinov, N.; Tascı, B.; Taheri, S.; Özkul, Y.; Donmez-Altuntas, H.; Canatan, H.; et al. FOXM1 plays a role in autophagy by transcriptionally regulating Beclin-1 and LC3 genes in human triple-negative breast cancer cells. J. Mol. Med. 2019, 97, 491–508. [Google Scholar] [CrossRef]
- Ma, K.; Zhang, C.; Huang, M.Y.; Li, W.Y.; Hu, G.Q. Cinobufagin induces autophagy-mediated cell death in human osteosarcoma U2OS cells through the ROS/JNK/p38 signaling pathway. Oncol. Rep. 2016, 36, 90–98. [Google Scholar] [CrossRef]
- Xia, H.; Green, D.R.; Zou, W. Autophagy in tumour immunity and therapy. Nat. Rev. Cancer 2021, 21, 281–297. [Google Scholar] [CrossRef]
- Xiong, X.; Lu, B.; Tian, Q.; Zhang, H.; Wu, M.; Guo, H.; Zhang, Q.; Li, X.; Zhou, T.; Wang, Y. Inhibition of autophagy enhances cinobufagin-induced apoptosis in gastric cancer. Oncol. Rep. 2019, 41, 492–500. [Google Scholar] [CrossRef]
- Sheng, X.; Zhu, P.; Qin, J.; Li, Q. The biological role of autophagy in regulating and controlling the proliferation of liver cancer cells induced by bufalin. Oncol. Rep. 2018, 39, 2931–2941. [Google Scholar] [CrossRef]
- Xie, C.M.; Chan, W.Y.; Yu, S.; Zhao, J.; Cheng, C.H. Bufalin induces autophagy-mediated cell death in human colon cancer cells through reactive oxygen species generation and JNK activation. Free. Radic. Biol. Med. 2011, 51, 1365–1375. [Google Scholar] [CrossRef]
- Shen, S.; Zhang, Y.; Wang, Z.; Liu, R.; Gong, X. Bufalin induces the interplay between apoptosis and autophagy in glioma cells through endoplasmic reticulum stress. Int. J. Biol. Sci. 2014, 10, 212–224. [Google Scholar] [CrossRef]
- Yiang, G.T.; Yu, Y.L.; Chou, P.L.; Tsai, H.F.; Chen, L.A.; Chen, Y.H.; Su, K.J.; Wang, J.J.; Bau, D.T.; Wei, C.W. The cytotoxic protein can induce autophagocytosis in addition to apoptosis in MCF-7 human breast cancer cells. In Vivo 2012, 26, 403–409. [Google Scholar]
- Ghavami, G.; Kiasari, R.E.; Pakzad, F.; Sardari, S. Effect of metformin alone and in combination with etoposide and epirubicin on proliferation, apoptosis, necrosis, and migration of B-CPAP and SW cells as thyroid cancer cell lines. Res. Pharm. Sci. 2023, 18, 185–201. [Google Scholar]
- Khan, I.; Yousif, A.; Chesnokov, M.; Hong, L.; Chefetz, I. A decade of cell death studies: Breathing new life into necroptosis. Pharmacol. Ther. 2021, 220, 107717. [Google Scholar] [CrossRef]
- Li, Y.; Gong, P.; Kong, C.; Tian, X. Bufalin engages in RIP1-dependent and ROS-dependent programmed necroptosis in breast cancer cells by targeting the RIP1/RIP3/PGAM5 pathway. Anti-Cancer Drugs 2019, 30, e0770. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.D.; Song, C.Y.; Kong, C.C.; Tian, X. Bufalin Induces Programmed Necroptosis in Triple-Negative Breast Cancer Drug-Resistant Cell Lines through RIP1/ROS-Mediated Pathway. Chin. J. Integr. Med. 2022, 28, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Tummers, B.; Green, D.R. Caspase-8: Regulating life and death. Immunol. Rev. 2017, 277, 76–89. [Google Scholar] [CrossRef] [PubMed]
- LingHu, H.R.; Luo, H.; Gang, L. Bufalin Induces Glioma Cell Death by Apoptosis or Necroptosis. OncoTargets Ther. 2020, 13, 4767–4778. [Google Scholar] [CrossRef]
- Lo, Y.L.; Fang, Y.H.; Chiu, Y.J.; Chang, C.Y.; Lee, C.H.; Liao, Z.X.; Wang, L.F. Light- and Redox-Responsive Block Copolymers of mPEG-SS-ONBMA as a Smart Drug Delivery Carrier for Cancer Therapy. Pharmaceutics 2022, 14, 2594. [Google Scholar] [CrossRef]
- Ghavami, S.; Asoodeh, A.; Klonisch, T.; Halayko, A.J.; Kadkhoda, K.; Kroczak, T.J.; Gibson, S.B.; Booy, E.P.; Naderi-Manesh, H.; Los, M. Brevinin-2R(1) semi-selectively kills cancer cells by a distinct mechanism, which involves the lysosomal-mitochondrial death pathway. J. Cell. Mol. Med. 2008, 12, 1005–1022. [Google Scholar] [CrossRef]
- Du, W.; Gu, M.; Hu, M.; Pinchi, P.; Chen, W.; Ryan, M.; Nold, T.; Bannaga, A.; Xu, H. Lysosomal Zn2+ release triggers rapid, mitochondria-mediated, non-apoptotic cell death in metastatic melanoma. Cell Rep. 2021, 37, 109848. [Google Scholar] [CrossRef]
- La Porta, S.; Roth, L.; Singhal, M.; Mogler, C.; Spegg, C.; Schieb, B.; Qu, X.; Adams, R.H.; Baldwin, H.S.; Savant, S.; et al. Endothelial Tie1-mediated angiogenesis and vascular abnormalization promote tumor progression and metastasis. J. Clin. Investig. 2018, 128, 834–845. [Google Scholar] [CrossRef]
- van Zoggel, H.; Carpentier, G.; Dos Santos, C.; Hamma-Kourbali, Y.; Courty, J.; Amiche, M.; Delbé, J. Antitumor and angiostatic activities of the antimicrobial peptide dermaseptin B2. PLoS ONE 2012, 7, e44351. [Google Scholar] [CrossRef]
- Yang, H.; Liu, Y.; Zhao, M.M.; Guo, Q.; Zheng, X.K.; Liu, D.; Zeng, K.W.; Tu, P.F. Therapeutic potential of targeting membrane-spanning proteoglycan SDC4 in hepatocellular carcinoma. Cell Death Dis. 2021, 12, 492. [Google Scholar] [CrossRef]
- Fang, K.; Zhan, Y.; Zhu, R.; Wang, Y.; Wu, C.; Sun, M.; Qiu, Y.; Yuan, Z.; Liang, X.; Yin, P.; et al. Bufalin suppresses tumour microenvironment-mediated angiogenesis by inhibiting the STAT3 signalling pathway. J. Transl. Med. 2021, 19, 383. [Google Scholar] [CrossRef] [PubMed]
- Pastushenko, I.; Blanpain, C. EMT Transition States during Tumor Progression and Metastasis. Trends Cell Biol. 2019, 29, 212–226. [Google Scholar] [CrossRef]
- Mirzaei, S.; Saghari, S.; Bassiri, F.; Raesi, R.; Zarrabi, A.; Hushmandi, K.; Sethi, G.; Tergaonkar, V. NF-κB as a regulator of cancer metastasis and therapy response: A focus on epithelial-mesenchymal transition. J. Cell. Physiol. 2022, 237, 2770–2795. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, Q.; Zou, G.; Gao, G.; Yue, Q. Arenobufagin, isolated from toad venom, inhibited epithelial-to-mesenchymal transition and suppressed migration and invasion of lung cancer cells via targeting IKKβ/NFκB signal cascade. J. Ethnopharmacol. 2020, 250, 112492. [Google Scholar] [CrossRef] [PubMed]
- Schatoff, E.M.; Goswami, S.; Zafra, M.P.; Foronda, M.; Shusterman, M.; Leach, B.I.; Katti, A.; Diaz, B.J.; Dow, L.E. Distinct Colorectal Cancer-Associated APC Mutations Dictate Response to Tankyrase Inhibition. Cancer Discov. 2019, 9, 1358–1371. [Google Scholar] [PubMed]
- Ghosh, N.; Hossain, U.; Mandal, A.; Sil, P.C. The Wnt signaling pathway: A potential therapeutic target against cancer. Ann. N. Y. Acad. Sci. 2019, 1443, 54–74. [Google Scholar] [CrossRef]
- Wang, J.; Cai, H.; Liu, Q.; Xia, Y.; Xing, L.; Zuo, Q.; Zhang, Y.; Chen, C.; Xu, K.; Yin, P.; et al. Cinobufacini Inhibits Colon Cancer Invasion and Metastasis via Suppressing Wnt/β-Catenin Signaling Pathway and EMT. Am. J. Chin. Med. 2020, 48, 703–718. [Google Scholar] [CrossRef]
- Bruno, A.; Bassani, B.; D’Urso, D.G.; Pitaku, I.; Cassinotti, E.; Pelosi, G.; Boni, L.; Dominioni, L.; Noonan, D.M.; Mortara, L.; et al. Angiogenin and the MMP9-TIMP2 axis are up-regulated in proangiogenic, decidual NK-like cells from patients with colorectal cancer. FASEB J. 2018, 32, 5365–5377. [Google Scholar] [CrossRef]
- Yin, J.H.; Zhu, X.Y.; Shi, W.D.; Liu, L.M. Huachansu injection inhibits metastasis of pancreatic cancer in mice model of human tumor xenograft. BMC Complement. Altern. Med. 2014, 14, 483. [Google Scholar] [CrossRef] [PubMed]
- Tauriello, D.V.F.; Palomo-Ponce, S.; Stork, D.; Berenguer-Llergo, A.; Badia-Ramentol, J.; Iglesias, M.; Sevillano, M.; Ibiza, S.; Cañellas, A.; Hernando-Momblona, X.; et al. TGFβ drives immune evasion in genetically reconstituted colon cancer metastasis. Nature 2018, 554, 538–543. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, C.; Xu, L.; Zang, K.; Ning, Z.; Jiang, F.; Chi, H.; Zhu, X.; Meng, Z. Bufalin suppresses hepatocellular carcinoma invasion and metastasis by targeting HIF-1α via the PI3K/AKT/mTOR pathway. Oncotarget 2016, 7, 20193–20208. [Google Scholar]
- Hong, S.H.; Kim, G.Y.; Chang, Y.C.; Moon, S.K.; Kim, W.J.; Choi, Y.H. Bufalin prevents the migration and invasion of T24 bladder carcinoma cells through the inactivation of matrix metalloproteinases and modulation of tight junctions. Int. J. Oncol. 2013, 42, 277–286. [Google Scholar] [PubMed]
- Gai, J.Q.; Sheng, X.; Qin, J.M.; Sun, K.; Zhao, W.; Ni, L. The effect and mechanism of bufalin on regulating hepatocellular carcinoma cell invasion and metastasis via Wnt/β-catenin signaling pathway. Int. J. Oncol. 2016, 48, 338–348. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.M.; Feng, L.X.; Liu, M.; Jin, W.H.; Luo, J.; Nie, A.Y.; Zhou, Y.; Li, Y.; Wu, W.Y.; Jiang, B.H.; et al. Possible target-related proteins and signal network of bufalin in A549 cells suggested by both iTRAQ-based and label-free proteomic analysis. Proteomics 2016, 16, 935–945. [Google Scholar] [CrossRef] [PubMed]
- Maimon, A.; Levi-Yahid, V.; Ben-Meir, K.; Halpern, A.; Talmi, Z.; Priya, S.; Mizraji, G.; Mistriel-Zerbib, S.; Berger, M.; Baniyash, M.; et al. Myeloid cell-derived PROS1 inhibits tumor metastasis by regulating inflammatory and immune responses via IL-10. J. Clin. Investig. 2021, 131, e126089. [Google Scholar] [CrossRef]
- Hinshaw, D.C.; Shevde, L.A. The Tumor Microenvironment Innately Modulates Cancer Progression. Cancer Res. 2019, 79, 4557–4566. [Google Scholar] [CrossRef]
- Wang, X.L.; Zhao, G.H.; Zhang, J.; Shi, Q.Y.; Guo, W.X.; Tian, X.L.; Qiu, J.Z.; Yin, L.Z.; Deng, X.M.; Song, Y. Immunomodulatory effects of cinobufagin isolated from Chan Su on activation and cytokines secretion of immunocyte in vitro. J. Asian Nat. Prod. Res. 2011, 13, 383–392. [Google Scholar] [CrossRef]
- Wang, J.Y.; Chen, L.; Zheng, Z.; Wang, Q.; Guo, J.; Xu, L. Cinobufocini inhibits NF-κB and COX-2 activation induced by TNF-α in lung adenocarcinoma cells. Oncol. Rep. 2012, 27, 1619–1624. [Google Scholar]
- Sivori, S.; Pende, D.; Quatrini, L.; Pietra, G.; Della Chiesa, M.; Vacca, P.; Tumino, N.; Moretta, F.; Mingari, M.C.; Locatelli, F.; et al. NK cells and ILCs in tumor immunotherapy. Mol. Asp. Med. 2021, 80, 100870. [Google Scholar] [CrossRef]
- Pantic, J.M.; Jovanovic, I.P.; Radosavljevic, G.D.; Gajovic, N.M.; Arsenijevic, N.N.; Conlon, J.M.; Lukic, M.L. The frog skin host-defense peptide frenatin 2.1S enhances recruitment, activation and tumoricidal capacity of NK cells. Peptides 2017, 93, 44–50. [Google Scholar] [CrossRef]
- Jiang, H.; Zuo, J.; Li, B.; Chen, R.; Luo, K.; Xiang, X.; Lu, S.; Huang, C.; Liu, L.; Tang, J.; et al. Drug-induced oxidative stress in cancer treatments: Angel or devil? Redox Biol. 2023, 63, 102754. [Google Scholar]
- Firczuk, M.; Bajor, M.; Graczyk-Jarzynka, A.; Fidyt, K.; Goral, A.; Zagozdzon, R. Harnessing altered oxidative metabolism in cancer by augmented prooxidant therapy. Cancer Lett. 2020, 471, 1–11. [Google Scholar] [PubMed]
- Li, Y.; Zhang, Y.; Wang, X.; Yang, Q.; Zhou, X.; Wu, J.; Yang, X.; Zhao, Y.; Lin, R.; Xie, Y.; et al. Bufalin induces mitochondrial dysfunction and promotes apoptosis of glioma cells by regulating Annexin A2 and DRP1 protein expression. Cancer Cell Int. 2021, 21, 424. [Google Scholar] [PubMed]
- Dong, Q.; Turdu, G.; Dongmulati, N.; Maimaitijang, A.; Aisa, H.A.; Yili, A. Bufadienolides from the Bufo viridis toad venom exert cytotoxic effects on cancer cells by inducing cell apoptosis and cell cycle arrest. Toxicol. Vitr. Int. J. Publ. Assoc. BIBRA 2023, 89, 105566. [Google Scholar]
- Lin, Y.; Jiang, Y.; Zhao, Z.; Lu, Y.; Xi, X.; Ma, C.; Chen, X.; Zhou, M.; Chen, T.; Shaw, C.; et al. Discovery of a Novel Antimicrobial Peptide, Temporin-PKE, from the Skin Secretion of Pelophylax kl. esculentus, and Evaluation of Its Structure-Activity Relationships. Biomolecules 2022, 12, 759. [Google Scholar]
- Qin, H.; Fang, H.; Chen, X.; Wang, L.; Ma, C.; Xi, X.; Chen, T.; Shaw, C.; Zhou, M. Exploration of the Structure-Function Relationships of a Novel Frog Skin Secretion-Derived Bioactive Peptide, t-DPH1, through Use of Rational Design, Cationicity Enhancement and In Vitro Studies. Antibiotics 2021, 10, 1529. [Google Scholar]
- Tan, Y.; Chen, X.; Ma, C.; Xi, X.; Wang, L.; Zhou, M.; Burrows, J.F.; Kwok, H.F.; Chen, T. Biological Activities of Cationicity-Enhanced and Hydrophobicity-Optimized Analogues of an Antimicrobial Peptide, Dermaseptin-PS3, from the Skin Secretion of Phyllomedusa sauvagii. Toxins 2018, 10, 320. [Google Scholar]
- André, S.; Raja, Z.; Humblot, V.; Piesse, C.; Foulon, T.; Sereno, D.; Oury, B.; Ladram, A. Functional Characterization of Temporin-SHe, a New Broad-Spectrum Antibacterial and Leishmanicidal Temporin-SH Paralog from the Sahara Frog (Pelophylax saharicus). Int. J. Mol. Sci. 2020, 21, 6713. [Google Scholar]
- Yang, Q.Z.; Wang, C.; Lang, L.; Zhou, Y.; Wang, H.; Shang, D.J. Design of potent, non-toxic anticancer peptides based on the structure of the antimicrobial peptide, temporin-1CEa. Arch. Pharmacal Res. 2013, 36, 1302–1310. [Google Scholar]
- Pandit-Taskar, N. Targeted Radioimmunotherapy and Theranostics with Alpha Emitters. J. Med. Imaging Radiat. Sci. 2019, 50 (Suppl. S1), S41–S44. [Google Scholar]
- Coerts, H.I.; de Keizer, B.; Marlowe, R.J.; Verburg, F.A. Recombinant or endogenous thyroid-stimulating hormone for radioactive iodine therapy in thyroid cancer: State of knowledge and current controversies. Eur. J. Endocrinol. 2023, 188, lvad006. [Google Scholar]
- Lin, R.; Ma, B.; Liu, N.; Zhang, L.; He, T.; Liu, X.; Chen, T.; Liu, W.; Liang, Y.; Wang, T.; et al. Targeted radioimmunotherapy with the iodine-131-labeled caerin 1.1 peptide for human anaplastic thyroid cancer in nude mice. Ann. Nucl. Med. 2021, 35, 811–822. [Google Scholar]
- Tan, X.; Liang, X.; Xi, J.; Guo, S.; Meng, M.; Chen, X.; Li, Y. Clinical efficacy and safety of Huachansu injection combination with platinum-based chemotherapy for advanced non-small cell lung cancer: A systematic review and meta-analysis of randomized controlled trials. Medicine 2021, 100, e27161. [Google Scholar]
- Xia, J.; Inagaki, Y.; Gao, J.; Qi, F.; Song, P.; Han, G.; Sawakami, T.; Gao, B.; Luo, C.; Kokudo, N.; et al. Combination of Cinobufacini and Doxorubicin Increases Apoptosis of Hepatocellular Carcinoma Cells through the Fas- and Mitochondria-Mediated Pathways. Am. J. Chin. Med. 2017, 45, 1537–1556. [Google Scholar]
- Sun, B.; Dong, Y.; Xu, J.; Wang, Z. Current status and progress in immunotherapy for malignant pleural mesothelioma. Chronic Dis. Transl. Med. 2022, 8, 91–99. [Google Scholar]
- Satoh, T.; Tatsuta, T.; Sugawara, S.; Hara, A.; Hosono, M. Synergistic anti-tumor effect of bullfrog sialic acid-binding lectin and pemetrexed in malignant mesothelioma. Oncotarget 2017, 8, 42466–42477. [Google Scholar]
- Tuyaerts, S.; Van Nuffel, A.M.T.; Naert, E.; Van Dam, P.A.; Vuylsteke, P.; De Caluwé, A.; Aspeslagh, S.; Dirix, P.; Lippens, L.; De Jaeghere, E.; et al. PRIMMO study protocol: A phase II study combining PD-1 blockade, radiation and immunomodulation to tackle cervical and uterine cancer. BMC Cancer 2019, 19, 506. [Google Scholar]
- Ni, G.; Yang, X.; Li, J.; Wu, X.; Liu, Y.; Li, H.; Chen, S.; Fogarty, C.E.; Frazer, I.H.; Chen, G.; et al. Intratumoral injection of caerin 1.1 and 1.9 peptides increases the efficacy of vaccinated TC-1 tumor-bearing mice with PD-1 blockade by modulating macrophage heterogeneity and the activation of CD8+ T cells in the tumor microenvironment. Clin. Transl. Immunol. 2021, 10, e1335. [Google Scholar]
- Wooten, J.; Mavingire, N.; Damar, K.; Perez, A.L.; Brantley, E. Triumphs and challenges in exploiting poly(ADP-ribose) polymerase inhibition to combat triple-negative breast cancer. J. Cell. Physiol. 2023, in press. [CrossRef]
- Raineri, A.; Prodomini, S.; Fasoli, S.; Gotte, G.; Menegazzi, M. Influence of onconase in the therapeutic potential of PARP inhibitors in A375 malignant melanoma cells. Biochem. Pharmacol. 2019, 167, 173–181. [Google Scholar]
- McNevin, C.S.; Baird, A.M.; McDermott, R.; Finn, S.P. Diagnostic Strategies for Treatment Selection in Advanced Prostate Cancer. Diagnostics 2021, 11, 345. [Google Scholar]
- Couty, M.; Dusaud, M.; Miro-Padovani, M.; Zhang, L.; Zadigue, P.; Zargarian, L.; Lequin, O.; de la Taille, A.; Delbe, J.; Hamma-Kourbali, Y.; et al. Antitumor Activity and Mechanism of Action of Hormonotoxin, an LHRH Analog Conjugated to Dermaseptin-B2, a Multifunctional Antimicrobial Peptide. Int. J. Mol. Sci. 2021, 22, 11303. [Google Scholar]
- Zhang, L.; Liu, G.; Xia, T.; Yang, X.; Sun, G.; Zhao, C.; Xu, C.; Zhang, H. Evolution of toll-like receptor gene family in amphibians. Int. J. Biol. Macromol. 2022, 208, 463–474. [Google Scholar]
- Torres-Dimas, E.; Cruz-Ramírez, A.; Bermúdez-Cruz, R.M. Cancer in Amphibia, a rare phenomenon? Cell Biol. Int. 2022, 46, 1992–1998. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Species | Amino Acid Numbers (aa) | Primary Structure | Cancers | References |
|---|---|---|---|---|---|
| Aurein 2.5 | Litoria aurea, Litoria raniformis | 16 | GLFDIVKKVVGAFGSL | Leukaemia, Lung, Colon, CNS, Melanoma, Ovarian, Renal, Prostate, Breast | [15] |
| Aurein 2.6 | Litoria raniformis | 16 | GLFDIAKKVIGVIGSL | ||
| Aurein 3.1 | Litoria aurea, Litoria raniformis | 17 | GLFDIVKKIAGHIAGSI | ||
| Aurein 3.2 | Litoria aurea, L itoria raniformis | 17 | GLFDIVKKIAGHIASSI | ||
| Aurein 3.3 | Litoria raniformis | 17 | GLFDIVKKIAGHIVSSI | ||
| Maximin-1 | Bombina maxima | 27 | GIGTKILGGVKTALKGALKELASTYAN | Leukaemia, Bladder | [16] |
| Maximin-3 | Bombina maxima | 27 | GIGGKILSGLKTALKGAAKELASTYLH | Leukaemia, Bladder | [16] |
| Maximin-4 | Bombina maxima | 27 | GIGGVLLSAGKAALKGLAKVLAEKYAN | Leukaemia | [16] |
| Maximin-5 | Bombina maxima | 27 | SIGAKILGGVKTFFKGALKELASTYLQ | Leukaemia | [16] |
| Bombinin H2 | Bombina variegata | 20 | IIGPVLGLVGSALGGLLKKI | NSCLC | [17,18] |
| Temporin A | Rana temporaria | 13 | FLPLIGRVLSGIL | NSCLC | [17,19] |
| Temporin L | Rana temporaria | 13 | FVQWFSKFLGRIL | Leukaemia, Lymphoma | [20] |
| Temporin-1DRa | Rana draytonii | 14 | HFLGTLVNLAKKIL | Liver | [21] |
| Temporin-PE | Pelophylax esculentus | 13 | FLPIVAKLLSGLL | Lung, Glioma, Prostate, Breast | [22] |
| Ps-1Pb | Pseudhymenochirus merlini | 29 | IKIPSFFRNILKKVGKEAVSLIAGALKQS | Lung, Breast, Colon | [23] |
| Ps-2Pa | Pseudhymenochirus merlini | 27 | GIFPIFAKLLGKVIKVASSLISKGRTE | Lung, Breast, Colon | [23] |
| Dermaseptin-B2 | Phyllomedusa bicolor | 33 | GLWSKIKEVGKEAAKAAAKAAGKAALGAVSEAV | Prostate, Pancreas, Melanoma | [24] |
| Dermaseptin-B3 | Phyllomedusa bicolor | 29 | ALWKNMLKGIGKLAGQAALGAVKTLVGAE | Prostate, Breast | [24] |
| Dermaseptin-B4 | Phyllomedusa bicolor | 28 | ALWKDILKNVGKAAGKAVLNTVTDMVNQ | Breast | [25] |
| DRS-DU-1 | Phyllomedusa duellmani | 28 | ALWKSLLKNVGKAAGKAALNAVTDMVNQ | Lung, Prostate | [26] |
| Dermaseptin-PD1 | Pachymedusa dacnicolor | 31 | GMWSKIKETAMAAAKEAAKAAGKTISDMIKQ | Glioma | [27] |
| Dermaseptin-PD2 | Pachymedusa dacnicolor | 33 | GMWSKIKNAGKAAAKAAAKAAGKAALDAVSEAI | Lung, Prostate, Glioma | [27] |
| Dermaseptin-PH | Phyllomedusa hypochondrialis | 23 | ALWKEVLKNAGKAALNEINNLVQ | Lung, Glioma, Prostate, Breast | [28] |
| Dermaseptin-PS4 | Phyllomedusa sauvagii | 28 | ALWKTLLKHVGKAAGKAALNAVTDMVNQ | Lung, Glioma, Prostate, Breast | [29] |
| Brevinin-1GHd | Hylarana guentheri | 24 | FLGALFKVASKLVPAAICSISKKC | Lung, Glioma, Prostate, Breast | [30] |
| Brevinin-1H | Amolops hainanensis | 21 | FALGAVTKVLPKLFCLITRKC | Lung, Prostate, Melanoma, Colon | [31] |
| Brevinin-1OS | Odorrana schmackeri | 23 | FPLIASLAGNVVPKIFCKITKRC | Lung, Glioma, Prostate, Breast, Colon | [32] |
| Bombinin-BO1 | Bombina orientalis | 25 | GIGSAILSAGKSIIKGLAKGLAEHF | Liver | [33] |
| Bombinin H-BO1 | Bombina orientalis | 17 | IIGPVLGLVGKALGGLL | Liver | [33] |
| BLP-7 | Bombina orientalis | 27 | GIGGALLSAGKSALKGLAKGLAEHFAN | Liver | [34] |
| Bombinin H-BO | Bombina orientalis | 17 | IIGPVLGLIGKALGGLL | Liver | [34] |
| PE-BBI | Pelophylax esculentus | 16 | GALKGCWTKSIPPKPC | Colon | [35] |
| Phylloseptin-PHa | Pithecopus hypochondrialis | 19 | FLSLIPAAISAVSALANHF | Colon, Breast | [36] |
| Alyteserin-2 | Alytes obstetricans | 16 | ILGKLLSTAAGLLSNL | Lung | [37] |
| HECI | Hylarana erythraea | 17 | TVLRGCWTFSFPPKPCI | Lung, Prostate, Breast | [38] |
| Ranatuerin-2PLx | Rhodopseudomonas palustris | 28 | GIMDTVKNAAKNLAGQLLDKLKCKITAC | Lung, Prostate, Breast, Glioma | [39] |
| Hymenochirin-1B | Hymenochirus boettgeri | 27 | KLSPETKDNLKKVLKGAIKGAIVAKMV | Lung, Breast, Colon, Liver | [40] |
| Esculentin-2CHa | Lithobates chiricahuensis | 37 | GFSSIFRGVAKFASKGLGKDLAKLGVDLVACKISKQC | Lung | [41] |
| Figainin 1 | Boana raniceps | 18 | FIGTLIPLALGALTKLFK | Melanoma, Breast, Cervical | [42] |
| Name | Species | Amino Acid Numbers (aa) | Primary Structure | Net Charges | Cancers | References |
|---|---|---|---|---|---|---|
| Ascaphin-8 | Ascaphus truei | 19 | GFKDLLKGAAKALVKTVLF | 3 | Liver | [52] |
| Aurein 1.2 | Litoria raniformis | 13 | GLFDIIKKIAESF | 0 | Leukaemia, Lung, Colon, CNS, Melanoma, Ovarian, Renal, Prostate, Breast | [15] |
| Citropin 1.1 | Litoria citropa | 16 | GLFDVIKKVASVIGGL | 1 | Leukaemia, Lung, Colon, CNS, Melanoma, Ovarian, Renal, Prostate, Breast | [50] |
| Dermaseptin-L1 | Hylomantis lemur | 32 | GLWSKIKEAAKAAGKAALNAVTGLVNQGDQPS | 2 | Liver | [6] |
| Dermaseptin-PT9 | Phyllomedusa tarsius | 25 | GLWSKIKDAAKTAGKAALGFVNEMV | 2 | Lung, Glioma, Prostate, Breast, Pancreas | [53] |
| Dermaseptin-PS1 | Paralabidochromis sauvagei | 31 | ALWKTMLKKLGTVALHAGKAALGAVADTISQ | 3.1 | Glioma | [54] |
| LFB | Limnonectes fujianensis | 33 | GLFSVVKGVLKGVGKNVSGSLLDQLKCKISGGC | 3.9 | Lung, Breast, Glioma, Colon | [55] |
| Magainin 2 | Bombina maxima | 23 | GIGKFLHSAKKFGKAFVGEIMNS | 3.1 | Bladder | [51] |
| Peptide XT-7 | Xenopus tropicalis | 18 | GLLGPLLKIAAKVGSNLL | 2 | Liver | [52] |
| Phylloseptin-L1 | Hylomantis lemur | 18 | LLGMIPLAISAISALSKL | 1 | Liver | [6] |
| Phylloseptin-PHa | Pithecopus hypochondrialis | 19 | FLSLIPAAISAVSALANHF | 0.1 | Lung, Glioma, Prostate, Breast, Colon | [36] |
| Temporin-1CEa | Rana chensinensis | 17 | FVDLKKIANIINSIFGK | 2 | Melanoma, Breast, Liver, Lung, Carcinoma, Colon | [47,48,56] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Q.; Wu, J.; Li, X.; Ye, Z.; Yang, H.; Mu, L. Amphibian-Derived Natural Anticancer Peptides and Proteins: Mechanism of Action, Application Strategies, and Prospects. Int. J. Mol. Sci. 2023, 24, 13985. https://doi.org/10.3390/ijms241813985
Chen Q, Wu J, Li X, Ye Z, Yang H, Mu L. Amphibian-Derived Natural Anticancer Peptides and Proteins: Mechanism of Action, Application Strategies, and Prospects. International Journal of Molecular Sciences. 2023; 24(18):13985. https://doi.org/10.3390/ijms241813985
Chicago/Turabian StyleChen, Qian, Jing Wu, Xiang Li, Ziyi Ye, Hailong Yang, and Lixian Mu. 2023. "Amphibian-Derived Natural Anticancer Peptides and Proteins: Mechanism of Action, Application Strategies, and Prospects" International Journal of Molecular Sciences 24, no. 18: 13985. https://doi.org/10.3390/ijms241813985
APA StyleChen, Q., Wu, J., Li, X., Ye, Z., Yang, H., & Mu, L. (2023). Amphibian-Derived Natural Anticancer Peptides and Proteins: Mechanism of Action, Application Strategies, and Prospects. International Journal of Molecular Sciences, 24(18), 13985. https://doi.org/10.3390/ijms241813985