
GDF15 Promotes the Osteogenic Cell Fate of Periodontal Ligament Fibroblasts, thus Affecting Their Mechanobiological Response
 and
and
Abstract
1. Introduction
2. Results
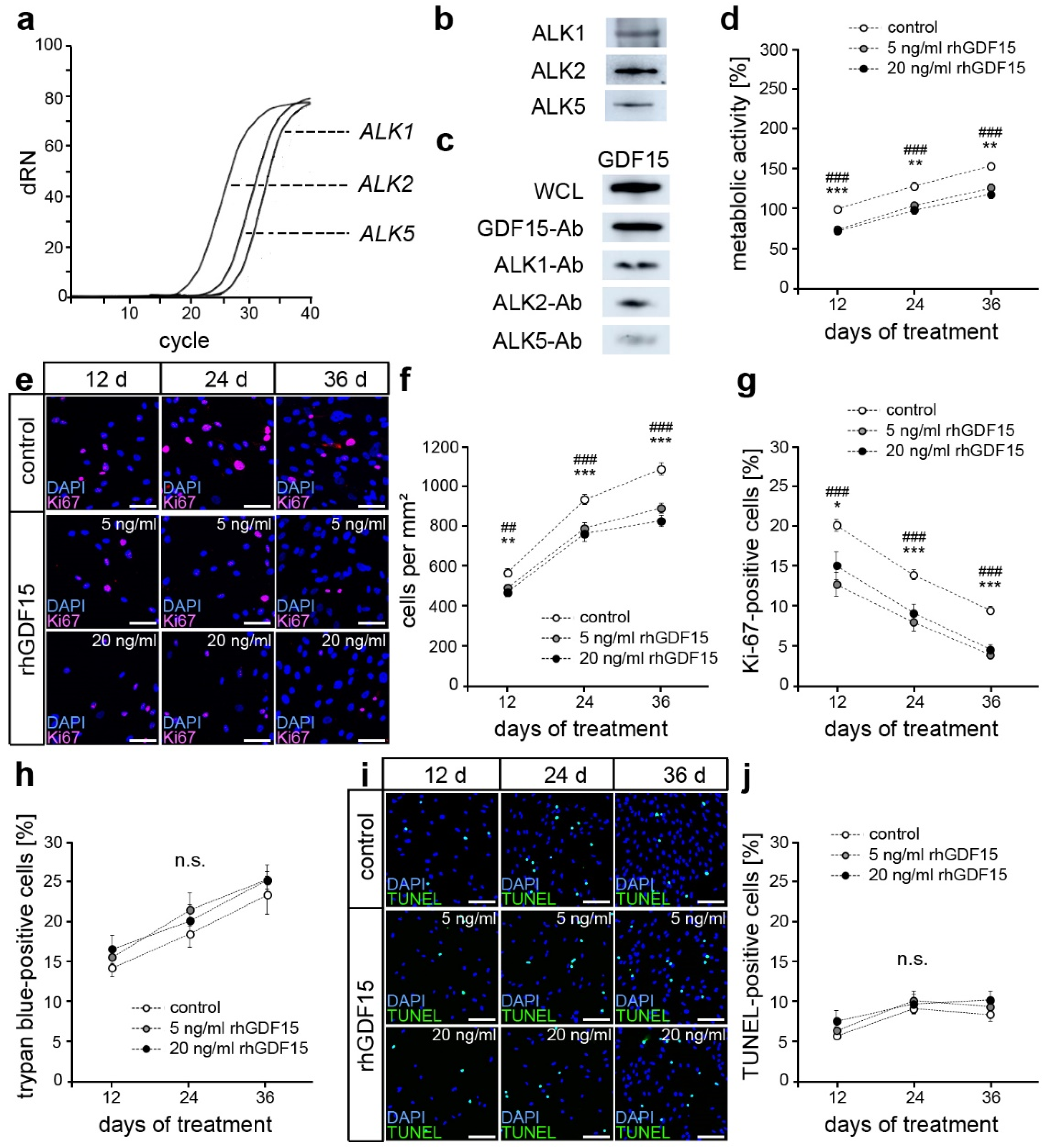
2.1. hPdLFs Express ALK1, ALK2, and ALK5
2.2. GDF15 Exposure Limits Cell Proliferation without Affecting the Survival of hPdLFs
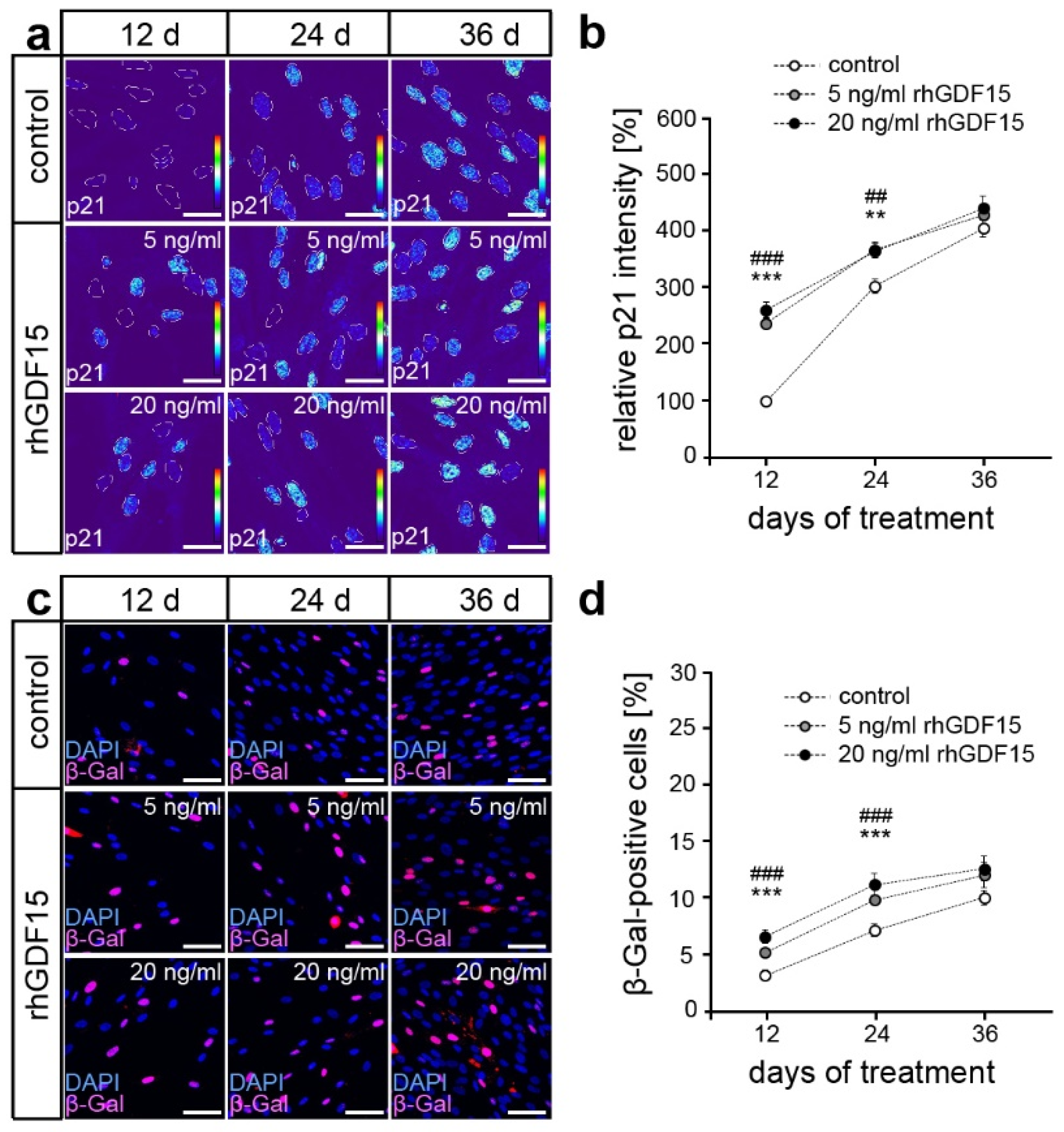
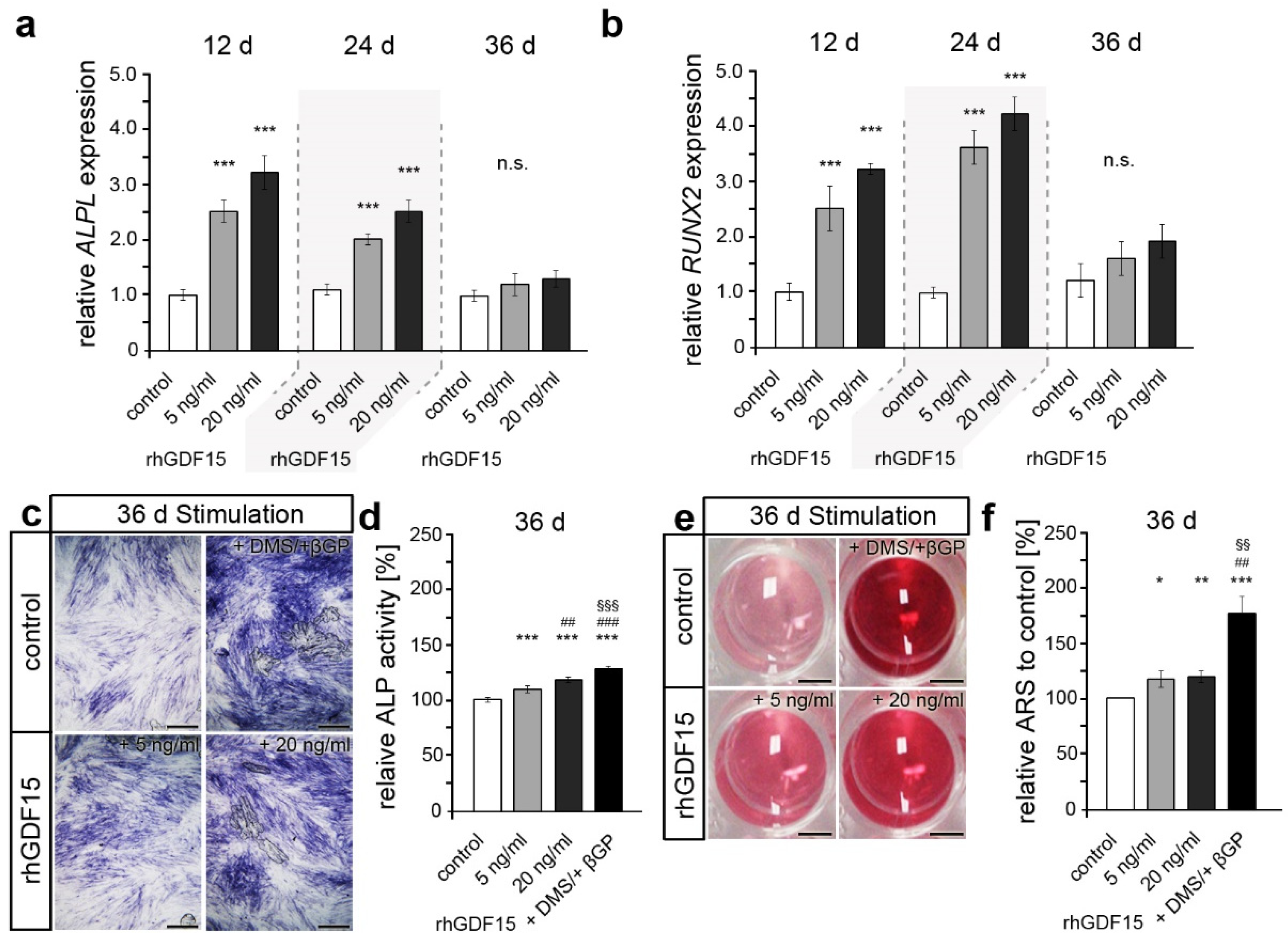
2.3. Long-Term Exposure to GDF15 Promotes Osteogenic Cell Fate of hPdLFs
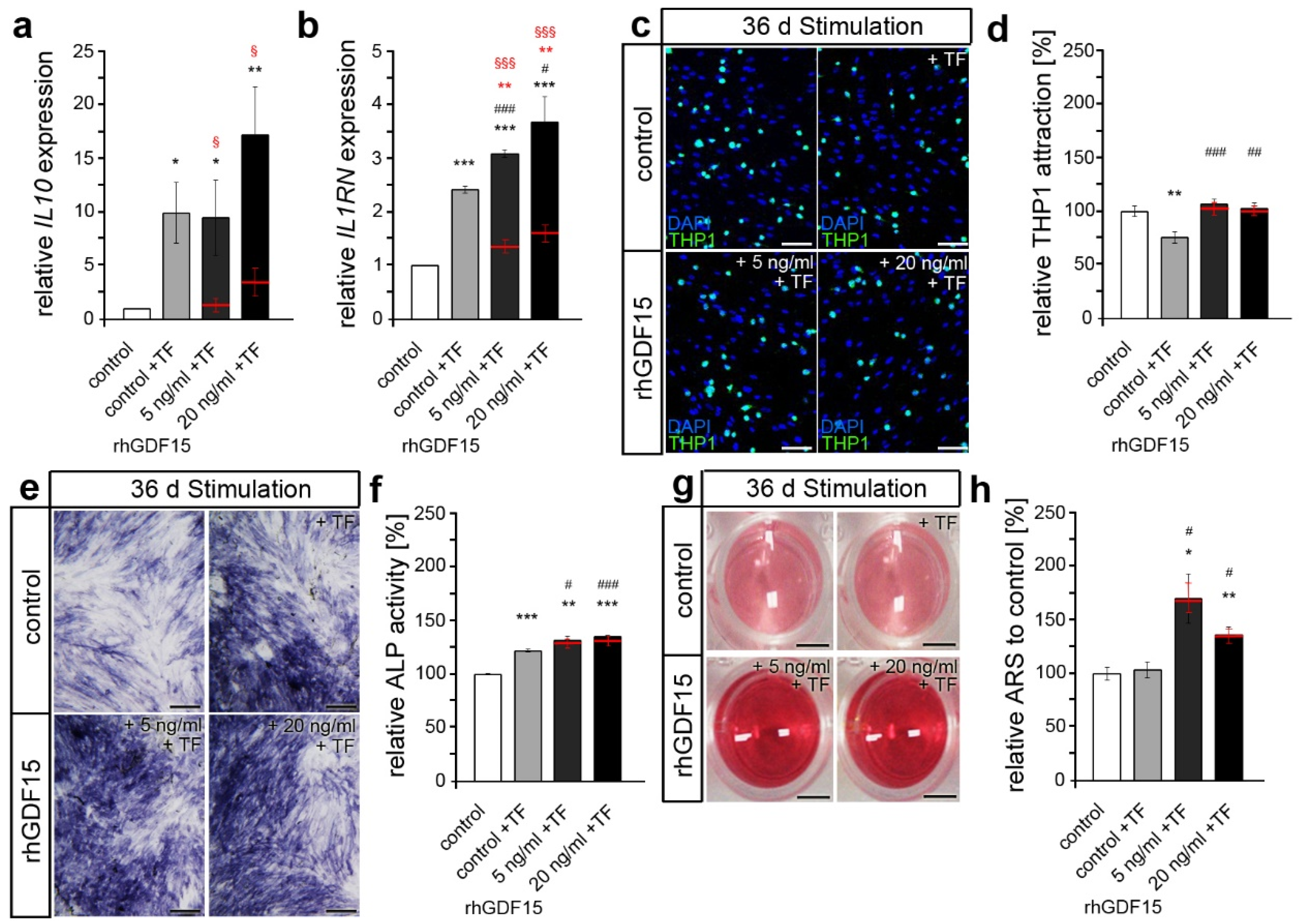
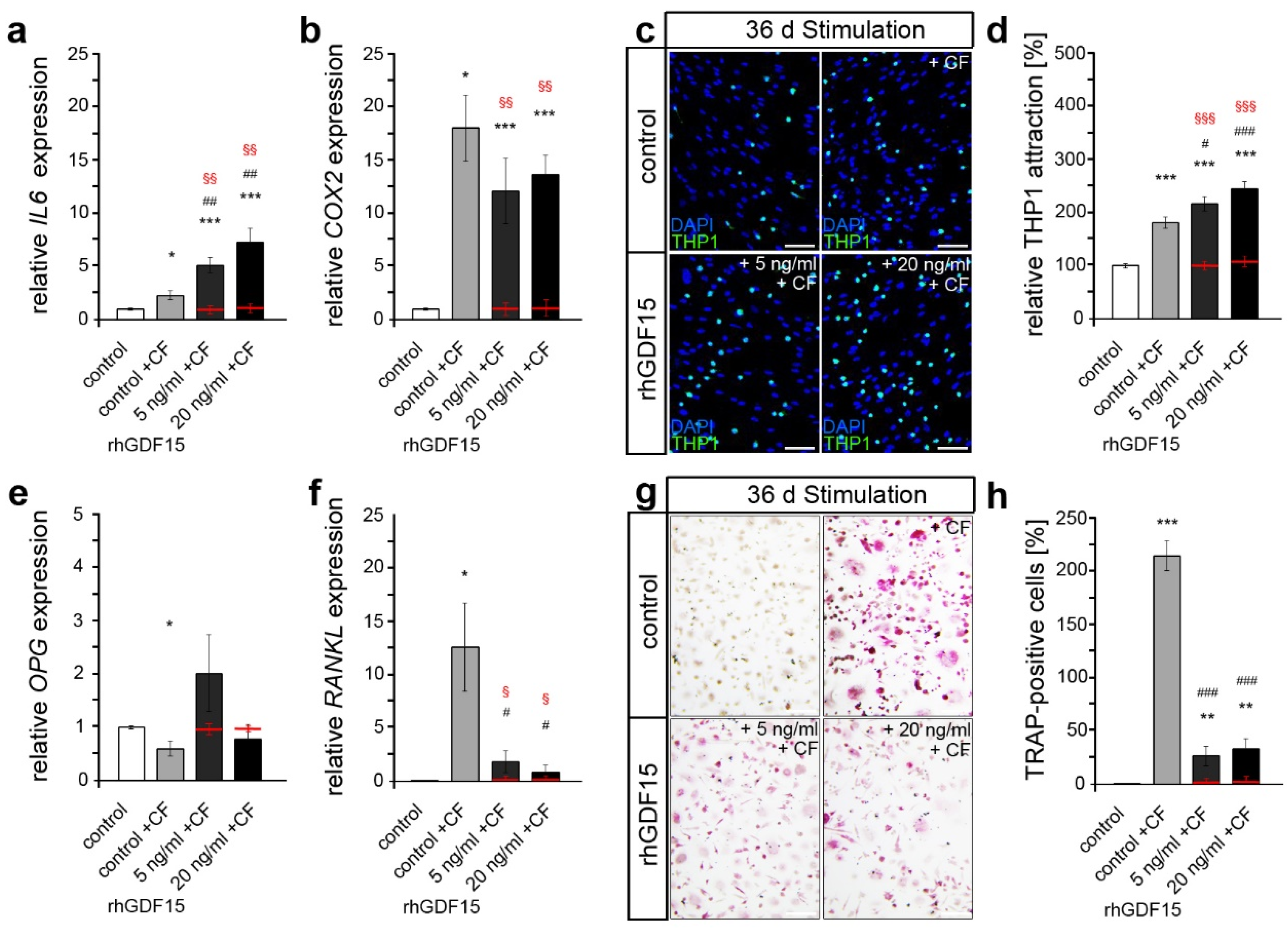
2.4. Prior Long-Term Exposure of hPdLFs to rhGDF15 Affects Their Mechanoreactivity
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Stimulation with Recombinant Human GDF15 Protein
4.3. Application of Mechanical Forces
4.4. RNA Expression Analysis
4.5. Co-Immunoprecipitation
4.6. Protein Preparation and Expression Analysis
4.7. Immunofluorescent Staining
4.8. MTT Assay
4.9. Cell Death Assays
4.10. Senescence Assay (β-Galactosidase Staining)
4.11. Alkaline Phosphatase Activity Assay
4.12. Alizarin Red Staining
4.13. THP1 Activation Assay
4.14. Osteoclast Activation Assay and TRAP Staining
4.15. Microscopy, Image Analysis, and Statistics
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Navabi, N.; Farnudi, H.; Rafiei, H.; Arashlow, M.T. Orthodontic treatment and the oral health-related quality of life of patients. J. Dent. 2012, 9, 247–254. [Google Scholar]
- Sa-Pinto, A.C.; Rego, T.M.; Marques, L.S.; Martins, C.C.; Ramos-Jorge, M.L.; Ramos-Jorge, J. Association between malocclusion and dental caries in adolescents: A systematic review and meta-analysis. Eur. Arch. Paediatr. Dent. 2018, 19, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Alsulaiman, A.A.; Kaye, E.; Jones, J.; Cabral, H.; Leone, C.; Will, L.; Garcia, R. Incisor malalignment and the risk of periodontal disease progression. Am. J. Orthod. Dentofac. Orthop. 2018, 153, 512–522. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Fukasawa, S. Is inflammation a friend or foe for orthodontic treatment?: Inflammation in orthodontically induced inflammatory root resorption and accelerating tooth movement. Int. J. Mol. Sci. 2021, 22, 2388. [Google Scholar] [CrossRef]
- Weltman, B.; Vig, K.W.; Fields, H.W.; Shanker, S.; Kaizar, E.E. Root resorption associated with orthodontic tooth movement: A systematic review. Am. J. Orthod. Dentofac. Orthop. 2010, 137, 462–476, discussion 412A. [Google Scholar] [CrossRef] [PubMed]
- Wishney, M. Potential risks of orthodontic therapy: A critical review and conceptual framework. Aust. Dent. J. 2017, 62 (Suppl. S1), 86–96. [Google Scholar] [CrossRef]
- Theodorou, C.I.; Kuijpers-Jagtman, A.M.; Bronkhorst, E.M.; Wagener, F. Optimal force magnitude for bodily orthodontic tooth movement with fixed appliances: A systematic review. Am. J. Orthod. Dentofac. Orthop. 2019, 156, 582–592. [Google Scholar] [CrossRef]
- Talic, N.F. Adverse effects of orthodontic treatment: A clinical perspective. Saudi Dent. J. 2011, 23, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Parcianello, R.G.; Amerio, E.; Giner Tarrida, L.; Nart, J.; Flores Mir, C.; Puigdollers Perez, A. Local hormones and growth factors to enhance orthodontic tooth movement: A systematic review of animal studies. Orthod. Craniofacial Res. 2022, 25, 281–303. [Google Scholar] [CrossRef]
- Andrade, I., Jr.; Taddei, S.R.A.; Souza, P.E.A. Inflammation and tooth movement: The role of cytokines, chemokines, and growth factors. Semin. Orthod. 2012, 18, 257–269. [Google Scholar] [CrossRef]
- Nanci, A.; Bosshardt, D.D. Structure of periodontal tissues in health and disease. Periodontol. 2000 2006, 40, 11–28. [Google Scholar] [CrossRef]
- Basdra, E.K.; Komposch, G. Osteoblast-like properties of human periodontal ligament cells: An in vitro analysis. Eur. J. Orthod. 1997, 19, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Somerman, M.J.; Young, M.F.; Foster, R.A.; Moehring, J.M.; Imm, G.; Sauk, J.J. Characteristics of human periodontal ligament cells in vitro. Arch. Oral. Biol. 1990, 35, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Arceo, N.; Sauk, J.J.; Moehring, J.; Foster, R.A.; Somerman, M.J. Human periodontal cells initiate mineral-like nodules in vitro. J. Periodontol. 1991, 62, 499–503. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, C.; Yang, Y. Effects of mechanical forces on osteogenesis and osteoclastogenesis in human periodontal ligament fibroblasts: A systematic review of in vitro studies. Bone Jt. Res. 2019, 8, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Garlet, T.P.; Coelho, U.; Silva, J.S.; Garlet, G.P. Cytokine expression pattern in compression and tension sides of the periodontal ligament during orthodontic tooth movement in humans. Eur. J. Oral. Sci. 2007, 115, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Howard, P.S.; Kucich, U.; Taliwal, R.; Korostoff, J.M. Mechanical forces alter extracellular matrix synthesis by human periodontal ligament fibroblasts. J. Periodontal Res. 1998, 33, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Nakashima, T.; Shinohara, M.; Negishi-Koga, T.; Komatsu, N.; Terashima, A.; Sawa, S.; Nitta, T.; Takayanagi, H. Osteoimmunology: The conceptual framework unifying the immune and skeletal systems. Physiol. Rev. 2017, 97, 1295–1349. [Google Scholar] [CrossRef]
- Tobeiha, M.; Moghadasian, M.H.; Amin, N.; Jafarnejad, S. Rankl/rank/opg pathway: A mechanism involved in exercise-induced bone remodeling. BioMed Res. Int. 2020, 2020, 6910312. [Google Scholar] [CrossRef]
- Ono, T.; Hayashi, M.; Sasaki, F.; Nakashima, T. Rankl biology: Bone metabolism, the immune system, and beyond. Inflamm. Regen. 2020, 40, 2. [Google Scholar] [CrossRef]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Tyrovola, J.B.; Spyropoulos, M.N.; Makou, M.; Perrea, D. Root resorption and the opg/rankl/rank system: A mini review. J. Oral. Sci. 2008, 50, 367–376. [Google Scholar] [CrossRef]
- Zhang, L.; Ding, Y.; Rao, G.Z.; Miao, D. Effects of il-10 and glucose on expression of opg and rankl in human periodontal ligament fibroblasts. Braz. J. Med. Biol. Res. 2016, 49, e4324. [Google Scholar] [CrossRef] [PubMed]
- Yan, T.; Xie, Y.; He, H.; Fan, W.; Huang, F. Role of nitric oxide in orthodontic tooth movement (review). Int. J. Mol. Med. 2021, 48, 168. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.; Zhao, Z.; Zhang, L.; Yu, Q.; Li, J.; Xu, Z.; Li, X. Up-regulated osteogenic transcription factors during early response of human periodontal ligament stem cells to cyclic tensile strain. Arch. Med. Sci. 2012, 8, 422–430. [Google Scholar] [CrossRef]
- Shen, T.; Qiu, L.; Chang, H.; Yang, Y.; Jian, C.; Xiong, J.; Zhou, J.; Dong, S. Cyclic tension promotes osteogenic differentiation in human periodontal ligament stem cells. Int. J. Clin. Exp. Pathol. 2014, 7, 7872–7880. [Google Scholar]
- Li, Y.; Jacox, L.A.; Little, S.H.; Ko, C.C. Orthodontic tooth movement: The biology and clinical implications. Kaohsiung J. Med. Sci. 2018, 34, 207–214. [Google Scholar] [CrossRef]
- Ullrich, N.; Schroder, A.; Jantsch, J.; Spanier, G.; Proff, P.; Kirschneck, C. The role of mechanotransduction versus hypoxia during simulated orthodontic compressive strain-an in vitro study of human periodontal ligament fibroblasts. Int. J. Oral. Sci. 2019, 11, 33. [Google Scholar] [CrossRef]
- Vansant, L.; Cadenas De Llano-Perula, M.; Verdonck, A.; Willems, G. Expression of biological mediators during orthodontic tooth movement: A systematic review. Arch. Oral. Biol. 2018, 95, 170–186. [Google Scholar] [CrossRef] [PubMed]
- Brooks, P.J.; Nilforoushan, D.; Manolson, M.F.; Simmons, C.A.; Gong, S.G. Molecular markers of early orthodontic tooth movement. Angle Orthod. 2009, 79, 1108–1113. [Google Scholar] [CrossRef] [PubMed]
- Bootcov, M.R.; Bauskin, A.R.; Valenzuela, S.M.; Moore, A.G.; Bansal, M.; He, X.Y.; Zhang, H.P.; Donnellan, M.; Mahler, S.; Pryor, K.; et al. Mic-1, a novel macrophage inhibitory cytokine, is a divergent member of the tgf-beta superfamily. Proc. Natl. Acad. Sci. USA 1997, 94, 11514–11519. [Google Scholar] [CrossRef]
- Assadi, A.; Zahabi, A.; Hart, R.A. Gdf15, an update of the physiological and pathological roles it plays: A review. Pflug. Arch. 2020, 472, 1535–1546. [Google Scholar] [CrossRef] [PubMed]
- Symmank, J.; Zimmermann, S.; Goldschmitt, J.; Schiegnitz, E.; Wolf, M.; Wehrbein, H.; Jacobs, C. Mechanically-induced gdf15 secretion by periodontal ligament fibroblasts regulates osteogenic transcription. Sci. Rep. 2019, 9, 11516. [Google Scholar] [CrossRef]
- Stemmler, A.; Symmank, J.; Steinmetz, J.; von Brandenstein, K.; Hennig, C.L.; Jacobs, C. Gdf15 supports the inflammatory response of pdl fibroblasts stimulated by p. Gingivalis lps and concurrent compression. Int. J. Mol. Sci. 2021, 22, 3608. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Day, E.A.; Townsend, L.K.; Djordjevic, D.; Jorgensen, S.B.; Steinberg, G.R. Gdf15: Emerging biology and therapeutic applications for obesity and cardiometabolic disease. Nat. Rev. Endocrinol. 2021, 17, 592–607. [Google Scholar] [CrossRef]
- Rochette, L.; Zeller, M.; Cottin, Y.; Vergely, C. Insights into mechanisms of gdf15 and receptor gfral: Therapeutic targets. Trends Endocrinol. Metab. 2020, 31, 939–951. [Google Scholar] [CrossRef] [PubMed]
- Mullican, S.E.; Lin-Schmidt, X.; Chin, C.N.; Chavez, J.A.; Furman, J.L.; Armstrong, A.A.; Beck, S.C.; South, V.J.; Dinh, T.Q.; Cash-Mason, T.D.; et al. Gfral is the receptor for gdf15 and the ligand promotes weight loss in mice and nonhuman primates. Nat. Med. 2017, 23, 1150–1157. [Google Scholar] [CrossRef]
- Wu, Q.; Jiang, D.; Matsuda, J.L.; Ternyak, K.; Zhang, B.; Chu, H.W. Cigarette smoke induces human airway epithelial senescence via growth differentiation factor 15 production. Am. J. Respir. Cell Mol. Biol. 2016, 55, 429–438. [Google Scholar] [CrossRef]
- Lin, W.; Zhang, W.W.; Lyu, N.; Cao, H.; Xu, W.D.; Zhang, Y.Q. Growth differentiation factor-15 produces analgesia by inhibiting tetrodotoxin-resistant nav1.8 sodium channel activity in rat primary sensory neurons. Neurosci. Bull. 2021, 37, 1289–1302. [Google Scholar] [CrossRef]
- Artz, A.; Butz, S.; Vestweber, D. Gdf-15 inhibits integrin activation and mouse neutrophil recruitment through the alk-5/tgf-betarii heterodimer. Blood 2016, 128, 529–541. [Google Scholar] [CrossRef]
- Min, K.W.; Liggett, J.L.; Silva, G.; Wu, W.W.; Wang, R.; Shen, R.F.; Eling, T.E.; Baek, S.J. Nag-1/gdf15 accumulates in the nucleus and modulates transcriptional regulation of the smad pathway. Oncogene 2016, 35, 377–388. [Google Scholar] [CrossRef]
- Lee, J.; Kim, I.; Yoo, E.; Baek, S.J. Competitive inhibition by nag-1/gdf-15 nls peptide enhances its anti-cancer activity. Biochem. Biophys. Res. Commun. 2019, 519, 29–34. [Google Scholar] [CrossRef]
- Wischhusen, J.; Melero, I.; Fridman, W.H. Growth/differentiation factor-15 (gdf-15): From biomarker to novel targetable immune checkpoint. Front. Immunol. 2020, 11, 951. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Huang, Y.; Lyu, Y.; Dai, W.; Tong, Y.; Li, Y. Gdf15 as a biomarker of ageing. Exp. Gerontol. 2021, 146, 111228. [Google Scholar] [CrossRef]
- Li, S.; Li, Q.; Zhu, Y.; Hu, W. Gdf15 induced by compressive force contributes to osteoclast differentiation in human periodontal ligament cells. Exp. Cell Res. 2020, 387, 111745. [Google Scholar] [CrossRef]
- Westhrin, M.; Moen, S.H.; Holien, T.; Mylin, A.K.; Heickendorff, L.; Olsen, O.E.; Sundan, A.; Turesson, I.; Gimsing, P.; Waage, A.; et al. Growth differentiation factor 15 (gdf15) promotes osteoclast differentiation and inhibits osteoblast differentiation and high serum gdf15 levels are associated with multiple myeloma bone disease. Haematologica 2015, 100, e511–e514. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, J.A.; Seshacharyulu, P.; Muniyan, S.; Pothuraju, R.; Khan, P.; Vengoji, R.; Chaudhary, S.; Maurya, S.K.; Lele, S.M.; Jain, M.; et al. Gdf15 promotes prostate cancer bone metastasis and colonization through osteoblastic ccl2 and rankl activation. Bone Res. 2022, 10, 6. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Walker, K.; Min, X.; Hale, C.; Tran, T.; Komorowski, R.; Yang, J.; Davda, J.; Nuanmanee, N.; Kemp, D.; et al. Long-acting mic-1/gdf15 molecules to treat obesity: Evidence from mice to monkeys. Sci. Transl. Med. 2017, 9, eaan8732. [Google Scholar] [CrossRef]
- Park, H.; Kim, C.H.; Jeong, J.H.; Park, M.; Kim, K.S. Gdf15 contributes to radiation-induced senescence through the ros-mediated p16 pathway in human endothelial cells. Oncotarget 2016, 7, 9634–9644. [Google Scholar] [CrossRef]
- Uchiyama, T.; Kawabata, H.; Miura, Y.; Yoshioka, S.; Iwasa, M.; Yao, H.; Sakamoto, S.; Fujimoto, M.; Haga, H.; Kadowaki, N.; et al. The role of growth differentiation factor 15 in the pathogenesis of primary myelofibrosis. Cancer Med. 2015, 4, 1558–1572. [Google Scholar] [CrossRef]
- Langenbach, F.; Handschel, J. Effects of dexamethasone, ascorbic acid and beta-glycerophosphate on the osteogenic differentiation of stem cells in vitro. Stem Cell Res. Ther. 2013, 4, 117. [Google Scholar] [CrossRef] [PubMed]
- Gry, M.; Rimini, R.; Stromberg, S.; Asplund, A.; Ponten, F.; Uhlen, M.; Nilsson, P. Correlations between rna and protein expression profiles in 23 human cell lines. BMC Genom. 2009, 10, 365. [Google Scholar] [CrossRef]
- Long, P.; Hu, J.; Piesco, N.; Buckley, M.; Agarwal, S. Low magnitude of tensile strain inhibits il-1beta-dependent induction of pro-inflammatory cytokines and induces synthesis of il-10 in human periodontal ligament cells in vitro. J. Dent. Res. 2001, 80, 1416–1420. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M. Rank/rankl/opg during orthodontic tooth movement. Orthod. Craniofacial Res. 2009, 12, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.H.; Si, Y.; Xu, G.; Chen, X.M.; Xiong, H.; Lai, L.; Zheng, Y.Q.; Zhang, Z.G. High-dose pma with rankl and mcsf induces thp1 cell differentiation into human functional osteoclasts in vitro. Mol. Med. Rep. 2017, 16, 8380–8384. [Google Scholar] [CrossRef]
- Asiry, M.A. Biological aspects of orthodontic tooth movement: A review of literature. Saudi J. Biol. Sci. 2018, 25, 1027–1032. [Google Scholar] [CrossRef]
- Sarkar, S.; Legere, S.; Haidl, I.; Marshall, J.; MacLeod, J.B.; Aguiar, C.; Lutchmedial, S.; Hassan, A.; Brunt, K.R.; Kienesberger, P.; et al. Serum gdf15, a promising biomarker in obese patients undergoing heart surgery. Front. Cardiovasc. Med. 2020, 7, 103. [Google Scholar] [CrossRef]
- Kim, Y.I.; Shin, H.W.; Chun, Y.S.; Park, J.W. Cst3 and gdf15 ameliorate renal fibrosis by inhibiting fibroblast growth and activation. Biochem. Biophys. Res. Commun. 2018, 500, 288–295. [Google Scholar] [CrossRef]
- Kim, Y.I.; Shin, H.W.; Chun, Y.S.; Cho, C.H.; Koh, J.; Chung, D.H.; Park, J.W. Epithelial cell-derived cytokines cst3 and gdf15 as potential therapeutics for pulmonary fibrosis. Cell Death Dis. 2018, 9, 506. [Google Scholar] [CrossRef]
- Guo, H.; Zhao, X.; Li, H.; Liu, K.; Jiang, H.; Zeng, X.; Chang, J.; Ma, C.; Fu, Z.; Lv, X.; et al. Gdf15 promotes cardiac fibrosis and proliferation of cardiac fibroblasts via the mapk/erk1/2 pathway after irradiation in rats. Radiat. Res. 2021, 196, 183–191. [Google Scholar] [CrossRef]
- Conte, M.; Giuliani, C.; Chiariello, A.; Iannuzzi, V.; Franceschi, C.; Salvioli, S. Gdf15, an emerging key player in human aging. Ageing Res. Rev. 2022, 75, 101569. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Ma, Y.M.; Zheng, P.S.; Zhang, P. Gdf15 promotes the proliferation of cervical cancer cells by phosphorylating akt1 and erk1/2 through the receptor erbb2. J. Exp. Clin. Cancer Res. 2018, 37, 80. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Chen, G.; Li, Y.P. Tgf-beta and bmp signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease. Bone Res. 2016, 4, 16009. [Google Scholar] [CrossRef]
- Quinn, J.M.; Itoh, K.; Udagawa, N.; Hausler, K.; Yasuda, H.; Shima, N.; Mizuno, A.; Higashio, K.; Takahashi, N.; Suda, T.; et al. Transforming growth factor beta affects osteoclast differentiation via direct and indirect actions. J. Bone Miner. Res. 2001, 16, 1787–1794. [Google Scholar] [CrossRef] [PubMed]
- Wakchoure, S.; Swain, T.M.; Hentunen, T.A.; Bauskin, A.R.; Brown, D.A.; Breit, S.N.; Vuopala, K.S.; Harris, K.W.; Selander, K.S. Expression of macrophage inhibitory cytokine-1 in prostate cancer bone metastases induces osteoclast activation and weight loss. Prostate 2009, 69, 652–661. [Google Scholar] [CrossRef] [PubMed]
- Tsai, V.W.W.; Husaini, Y.; Sainsbury, A.; Brown, D.A.; Breit, S.N. The mic-1/gdf15-gfral pathway in energy homeostasis: Implications for obesity, cachexia, and other associated diseases. Cell Metab. 2018, 28, 353–368. [Google Scholar] [CrossRef]
- Conte, M.; Ostan, R.; Fabbri, C.; Santoro, A.; Guidarelli, G.; Vitale, G.; Mari, D.; Sevini, F.; Capri, M.; Sandri, M.; et al. Human aging and longevity are characterized by high levels of mitokines. J. Gerontol. A Biol. Sci. Med. Sci. 2019, 74, 600–607. [Google Scholar] [CrossRef] [PubMed]
- Andersson-Hall, U.; Svedin, P.; Mallard, C.; Blennow, K.; Zetterberg, H.; Holmang, A. Growth differentiation factor 15 increases in both cerebrospinal fluid and serum during pregnancy. PLoS ONE 2021, 16, e0248980. [Google Scholar] [CrossRef]
- Klein, A.B.; Nicolaisen, T.S.; Ortenblad, N.; Gejl, K.D.; Jensen, R.; Fritzen, A.M.; Larsen, E.L.; Karstoft, K.; Poulsen, H.E.; Morville, T.; et al. Pharmacological but not physiological gdf15 suppresses feeding and the motivation to exercise. Nat. Commun. 2021, 12, 1041. [Google Scholar] [CrossRef]
- Kim, Y.; Noren Hooten, N.; Evans, M.K. Crp stimulates gdf15 expression in endothelial cells through p53. Mediat. Inflamm. 2018, 2018, 8278039. [Google Scholar] [CrossRef]
- Nishijima, Y.; Yamaguchi, M.; Kojima, T.; Aihara, N.; Nakajima, R.; Kasai, K. Levels of rankl and opg in gingival crevicular fluid during orthodontic tooth movement and effect of compression force on releases from periodontal ligament cells in vitro. Orthod. Craniofacial Res. 2006, 9, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Aihara, N.; Kojima, T.; Kasai, K. Rankl increase in compressed periodontal ligament cells from root resorption. J. Dent. Res. 2006, 85, 751–756. [Google Scholar] [CrossRef] [PubMed]
- Oshiro, T.; Shiotani, A.; Shibasaki, Y.; Sasaki, T. Osteoclast induction in periodontal tissue during experimental movement of incisors in osteoprotegerin-deficient mice. Anat. Rec. 2002, 266, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Bottner, M.; Laaff, M.; Schechinger, B.; Rappold, G.; Unsicker, K.; Suter-Crazzolara, C. Characterization of the rat, mouse, and human genes of growth/differentiation factor-15/macrophage inhibiting cytokine-1 (gdf-15/mic-1). Gene 1999, 237, 105–111. [Google Scholar] [CrossRef]
- Kempf, T.; Horn-Wichmann, R.; Brabant, G.; Peter, T.; Allhoff, T.; Klein, G.; Drexler, H.; Johnston, N.; Wallentin, L.; Wollert, K.C. Circulating concentrations of growth-differentiation factor 15 in apparently healthy elderly individuals and patients with chronic heart failure as assessed by a new immunoradiometric sandwich assay. Clin. Chem. 2007, 53, 284–291. [Google Scholar] [CrossRef]
- Benichou, O.; Coskun, T.; Gonciarz, M.D.; Garhyan, P.; Adams, A.C.; Du, Y.; Dunbar, J.D.; Martin, J.A.; Mather, K.J.; Pickard, R.T.; et al. Discovery, development, and clinical proof of mechanism of ly3463251, a long-acting gdf15 receptor agonist. Cell Metab. 2023, 35, 274–286.e210. [Google Scholar] [CrossRef]
- Kirschneck, C.; Batschkus, S.; Proff, P.; Kostler, J.; Spanier, G.; Schroder, A. Valid gene expression normalization by rt-qpcr in studies on hpdl fibroblasts with focus on orthodontic tooth movement and periodontitis. Sci. Rep. 2017, 7, 14751. [Google Scholar] [CrossRef]
- Symmank, J.; Appel, S.; Bastian, J.A.; Knaup, I.; Marciniak, J.; Hennig, C.L.; Doding, A.; Schulze-Spate, U.; Jacobs, C.; Wolf, M. Hyperlipidemic conditions impact force-induced inflammatory response of human periodontal ligament fibroblasts concomitantly challenged with p. Gingivalis-lps. Int. J. Mol. Sci. 2021, 22, 6069. [Google Scholar] [CrossRef]
- Nazet, U.; Schroder, A.; Spanier, G.; Wolf, M.; Proff, P.; Kirschneck, C. Simplified method for applying static isotropic tensile strain in cell culture experiments with identification of valid rt-qpcr reference genes for pdl fibroblasts. Eur. J. Orthod. 2020, 42, 359–370. [Google Scholar] [CrossRef]
- Schuldt, L.; Reimann, M.; von Brandenstein, K.; Steinmetz, J.; Doding, A.; Schulze-Spate, U.; Jacobs, C.; Symmank, J. Palmitate-triggered cox2/pge2-related hyperinflammation in dual-stressed pdl fibroblasts is mediated by repressive h3k27 trimethylation. Cells 2022, 11, 955. [Google Scholar] [CrossRef] [PubMed]
- Symmank, J.; Bayer, C.; Schmidt, C.; Hahn, A.; Pensold, D.; Zimmer-Bensch, G. Dnmt1 modulates interneuron morphology by regulating pak6 expression through crosstalk with histone modifications. Epigenetics 2018, 13, 536–556. [Google Scholar] [CrossRef] [PubMed]
- Symmank, J.; Chorus, M.; Appel, S.; Marciniak, J.; Knaup, I.; Bastian, A.; Hennig, C.L.; Doding, A.; Schulze-Spate, U.; Jacobs, C.; et al. Distinguish fatty acids impact survival, differentiation and cellular function of periodontal ligament fibroblasts. Sci. Rep. 2020, 10, 15706. [Google Scholar] [CrossRef] [PubMed]
- Schuldt, L.; von Brandenstein, K.; Jacobs, C.; Symmank, J. Oleic acid-related anti-inflammatory effects in force-stressed pdl fibroblasts are mediated by h3 lysine acetylation associated with altered il10 expression. Epigenetics 2022, 17, 1892–1904. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Symbol | NCBI Gene ID | Primer Sequence | Length |
|---|---|---|---|---|
| Activin A receptor like type 1 | ACVRL1 (alias ALK1) | 94 | fw: GTGGAGTGTGTGGGAAAAGG rev: CATGTCTGAGGCGATGAAGC | 180 bp |
| Activin A receptor type 1 | ACVR1 (alias ALK2) | 90 | fw: GCATTCCCAGAGCACCAATC rev: GGCCACTTCCCACAAAACAA | 166 bp |
| Alkaline Phosphatase | ALPL | 249 | fw: ACTGCAGACATTCTCAAA rev: GAGTGAGTGAGTGAGCA | 190 bp |
| Interleukin 6 | IL6 | 3569 | fw: CATCCTCGACGGCATCTCAG rev: TCACCAGGCAAGTCTCCTCA | 164 bp |
| Interleukin 10 | IL10 | 3586 | fw: AGCCATGAGTGAGTTTGACA rev: AGAGCCCCAGATCCGATTTT | 141 bp |
| Interleukin 1 receptor antagonist | IL1RN | 3557 | fw: GATGTGCCTGTCCTGTGTCA rev: ACTCAAAACTGGTGGTGGGG | 146 bp |
| Prostaglandin-endoperoxide synthase 2 | PTGS2 (alias COX2) | 4743 | fw: GATGATTGCCCGACTCCCTT rev: GGCCCTCGCTTATGATCTGT | 185 bp |
| Ribosomal protein L22 | RPL22 | 6146 | fw: TGATTGCACCCACCCTGTAG rev: GGTTCCCAGCTTTTCCGT TC | 98 bp |
| RUNX family transcription factor 2 | RUNX2 | 6146 | fw: CCCACGAATGCACTATCC rev: GGACATACCGAGGGACA | 120 bp |
| TATA-box binding protein | TBP | 6908 | fw: CGGCTGTTTAACTTCGCTTCC rev: TGGGTTATCTTCACACGCCAAG | 86 bp |
| TNF receptor superfamily member 11b | TNFRSF11B (alias OPG) | 4982 | fw: GAAGGGCGCTACCTTGA | 142 bp |
| rev: GCAAACTGTATTTCGCTC | ||||
| TNF Superfamily Member 11 | TNFSF11 (alias RANKL) | 8600 | fw: ATCACAGCACATCAGAGCAGA rev: TCACTTTATGGGAACCAGATGGG | 160 bp |
| Transforming growth factor beta receptor 1 | TGFBR1 (alias ALK5) | 7046 | fw: AAAACTTGCTCTGTCCACGG rev: TGCCAGTCCTAAGTCTGCAA | 157 bp |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lösch, L.; Stemmler, A.; Fischer, A.; Steinmetz, J.; Schuldt, L.; Hennig, C.-L.; Symmank, J.; Jacobs, C. GDF15 Promotes the Osteogenic Cell Fate of Periodontal Ligament Fibroblasts, thus Affecting Their Mechanobiological Response. Int. J. Mol. Sci. 2023, 24, 10011. https://doi.org/10.3390/ijms241210011
Lösch L, Stemmler A, Fischer A, Steinmetz J, Schuldt L, Hennig C-L, Symmank J, Jacobs C. GDF15 Promotes the Osteogenic Cell Fate of Periodontal Ligament Fibroblasts, thus Affecting Their Mechanobiological Response. International Journal of Molecular Sciences. 2023; 24(12):10011. https://doi.org/10.3390/ijms241210011
Chicago/Turabian StyleLösch, Lukas, Albert Stemmler, Adrian Fischer, Julia Steinmetz, Lisa Schuldt, Christoph-Ludwig Hennig, Judit Symmank, and Collin Jacobs. 2023. "GDF15 Promotes the Osteogenic Cell Fate of Periodontal Ligament Fibroblasts, thus Affecting Their Mechanobiological Response" International Journal of Molecular Sciences 24, no. 12: 10011. https://doi.org/10.3390/ijms241210011
APA StyleLösch, L., Stemmler, A., Fischer, A., Steinmetz, J., Schuldt, L., Hennig, C.-L., Symmank, J., & Jacobs, C. (2023). GDF15 Promotes the Osteogenic Cell Fate of Periodontal Ligament Fibroblasts, thus Affecting Their Mechanobiological Response. International Journal of Molecular Sciences, 24(12), 10011. https://doi.org/10.3390/ijms241210011