
Dysregulation of Immune Cell Subpopulations in Atypical Hemolytic Uremic Syndrome

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Results
2.1. The Demography of Studied Cases
2.2. The Immunological Landscape of Immune Cells from aHUS, aHUS Family, and Healthy
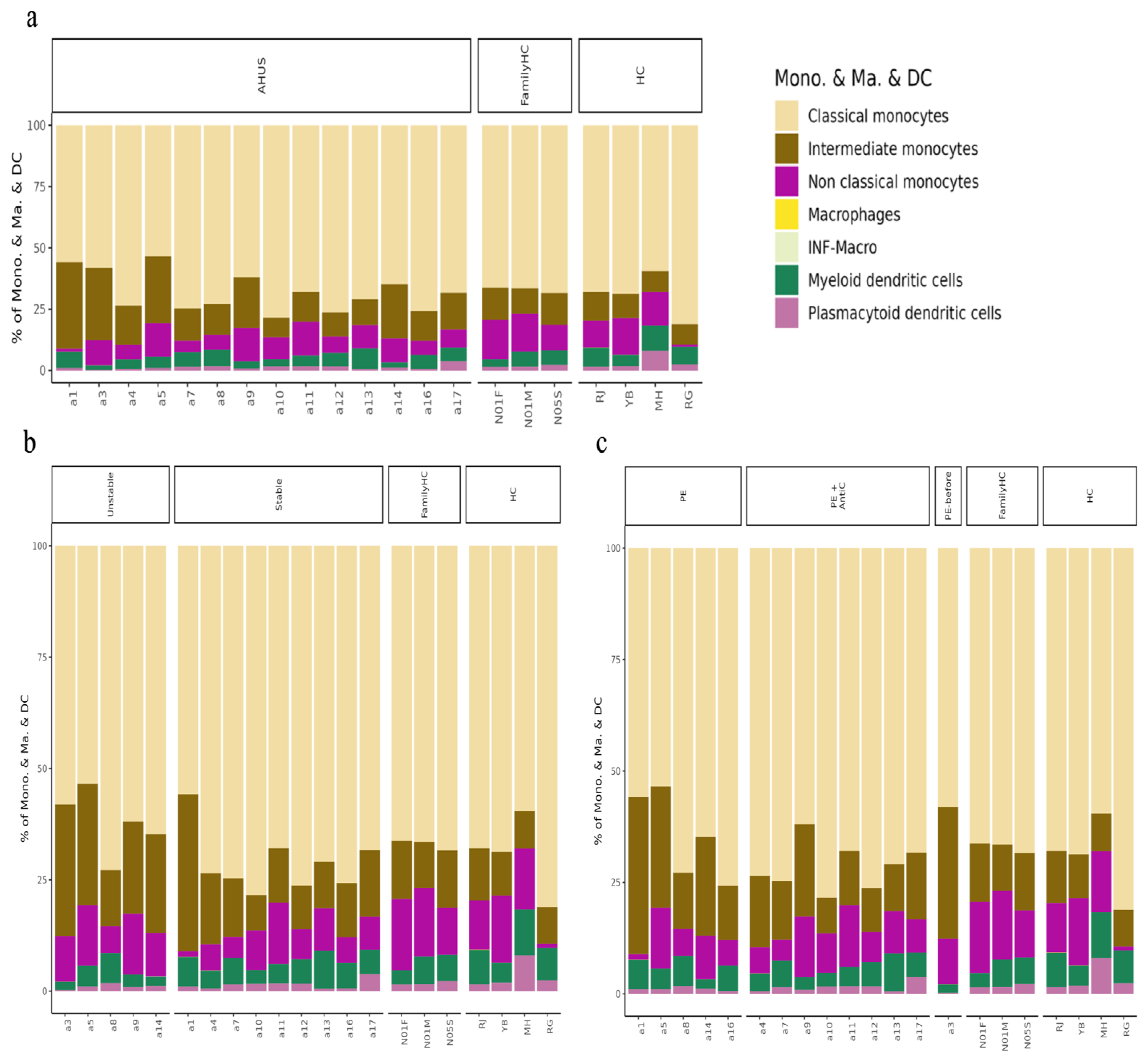
2.3. Cell Populations in PBMCs
2.3.1. Comparing aHUS Patients, aHUS Family, and Healthy Controls
2.3.2. Comparing Stable and Unstable aHUS Patients, aHUS Family, and Healthy Controls
2.3.3. Comparing Different Treatment in aHUS Patients, aHUS Family, and Healthy Controls
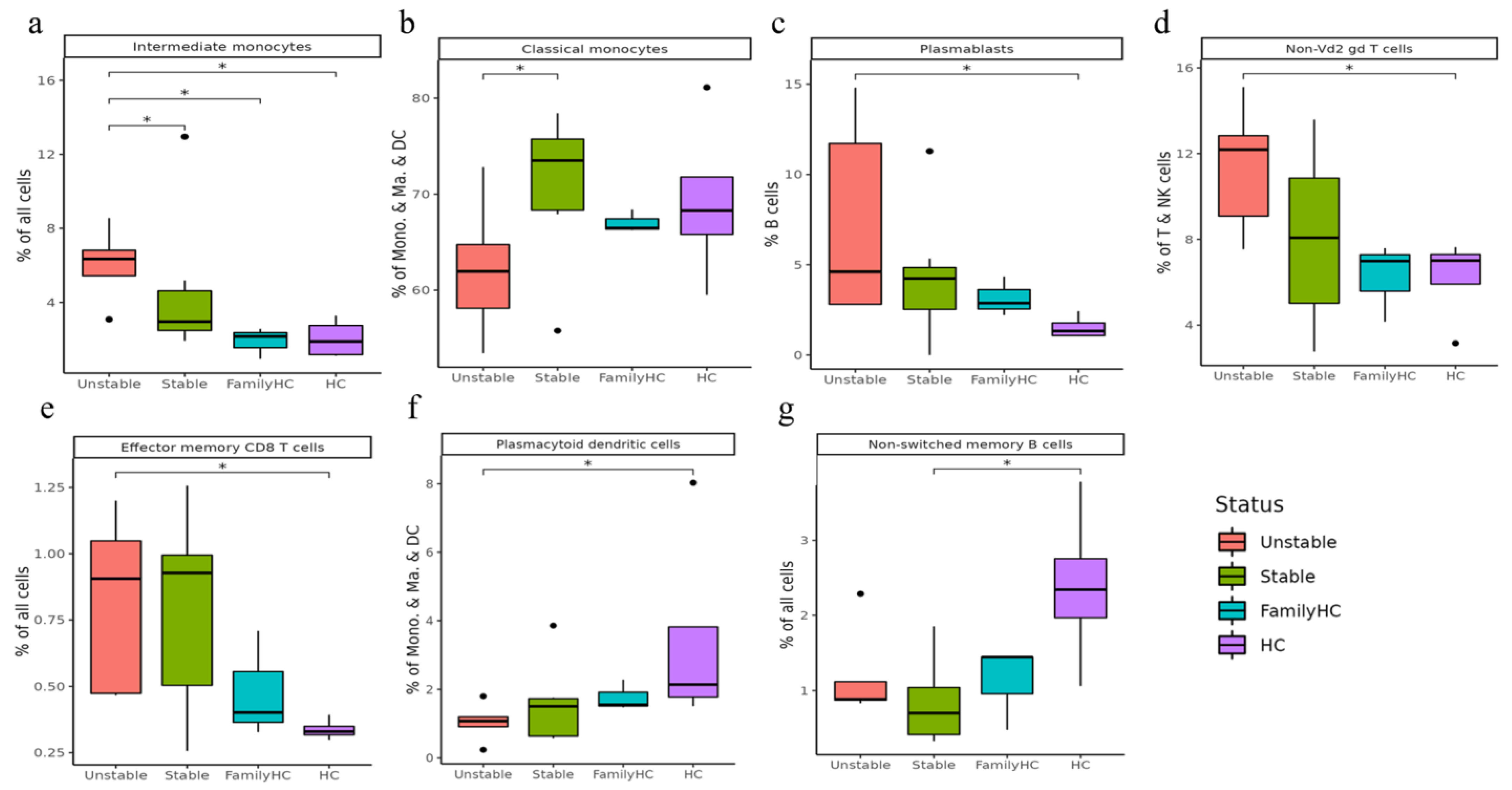
2.4. Cell Subclusters in PBMCs
2.4.1. Comparing aHUS Patients, aHUS Family, and Healthy Controls
2.4.2. Comparing Stable and Unstable aHUS Patients, aHUS Family, and Healthy Controls
2.5. Trajectory Analysis for B-Cell, T-Cell, and Monocyte
2.5.1. Comparing aHUS Patients, aHUS Family, and Healthy Control
2.5.2. Comparing Stable and Unstable aHUS Patients, aHUS Family, and Healthy Controls
2.6. Immune Cell Interactions in Blood Samples from aHUS, aHUS Family, and Healthy Control
3. Discussion
4. Materials and Methods
4.1. Patient Recruitment
4.2. Single Cell RNA-Seq and Data Analysis
4.3. Single Cell RNA-Seq Data Integration and Clustering
4.4. Cell Type Annotations
4.5. Clustering Analysis
4.6. Pseudotime Estimation
4.7. Cell–Cell Communication Analysis
4.8. Statistical Analysis
4.9. Ethics Approval and Consent to Participate
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| aHUS | Atypical hemolytic uremic syndrome |
| TMA | Thrombotic microangiopathy |
| CH50 | 50% hemolytic complement activity |
| PBMCs | Peripheral blood mononuclear cells |
| scRNA-seq | Single-cell RNA sequencing |
| PCA | Principal component analysis |
| rPCA | Robust principal component analysis |
| UMAP | Uniform manifold approximation and projection |
| NK | Natural killer |
| Mono | Monocytes |
| Ma | Macrophages |
| DC | Dendritic cells |
| STRING | Search Tool for the Retrieval of Interacting Genes/Proteins |
References
- Tsai, H.-M. Atypical Hemolytic Uremic Syndrome: Beyond Hemolysis and Uremia. Am. J. Med. 2019, 132, 161−167. [Google Scholar] [CrossRef] [PubMed]
- Noris, M.; Remuzzi, G. Atypical hemolytic-uremic syndrome. N. Engl. J. Med. 2009, 361, 1676–1687. [Google Scholar] [CrossRef] [PubMed]
- Bu, F.; Maga, T.; Meyer, N.C.; Wang, K.; Thomas, C.P.; Nester, C.M.; Smith, R.J. Comprehensive genetic analysis of complement and coagulation genes in atypical hemolytic uremic syndrome. J. Am. Soc. Nephrol. 2014, 25, 55–64. [Google Scholar] [CrossRef]
- Afshar-Kharghan, V. Atypical hemolytic uremic syndrome. Hematol. Am. Soc. Hematol. Educ. Program 2016, 2016, 217–225. [Google Scholar] [CrossRef]
- Tseng, M.H.; Lin, S.H.; Tsai, J.D.; Wu, M.S.; Tsai, I.J.; Chen, Y.C.; Chang, M.C.; Chou, W.C.; Chiou, Y.H.; Huang, C.C. Atypical hemolytic uremic syndrome: Consensus of diagnosis and treatment in Taiwan. J. Formos. Med. Assoc. 2023, 122, 366–375. [Google Scholar] [CrossRef]
- Merle, N.S.; Noe, R.; Halbwachs-Mecarelli, L.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement System Part II: Role in Immunity. Front. Immunol. 2015, 6, 257. [Google Scholar] [CrossRef]
- Fremeaux-Bacchi, V.; Fakhouri, F.; Garnier, A.; Bienaimé, F.; Dragon-Durey, M.A.; Ngo, S.; Moulin, B.; Servais, A.; Provot, F.; Rostaing, L.; et al. Genetics and outcome of atypical hemolytic uremic syndrome: A nationwide French series comparing children and adults. Clin. J. Am. Soc. Nephrol. 2013, 8, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Ding, Q.; Dai, D.F.; Padhy, B.; Nayak, M.K.; Li, C.; Purvis, M.; Jin, H.; Shu, C.; Chauhan, A.K.; et al. Loss of diacylglycerol kinase ε causes thrombotic microangiopathy by impairing endothelial VEGFA signaling. JCI Insight 2021, 6, e146959. [Google Scholar] [CrossRef]
- Challis, R.C.; Ring, T.; Xu, Y.; Wong, E.K.; Flossmann, O.; Roberts, I.S.; Ahmed, S.; Wetherall, M.; Salkus, G.; Brocklebank, V.; et al. Thrombotic Microangiopathy in Inverted Formin 2-Mediated Renal Disease. J. Am. Soc. Nephrol. 2017, 28, 1084–1091. [Google Scholar] [CrossRef]
- Van Quekelberghe, C.; Latta, K.; Kunzmann, S.; Grohmann, M.; Hansen, M. Atypical hemolytic uremic syndrome induced by SARS-CoV2 infection in infants with EXOSC3 mutation. Pediatr. Nephrol. 2022, 37, 2781–2784. [Google Scholar] [CrossRef]
- Canpolat, N.; Liu, D.; Atayar, E.; Saygili, S.; Kara, N.S.; Westfall, T.A.; Ding, Q.; Brown, B.J.; Braun, T.A.; Slusarski, D.; et al. A splice site mutation in the TSEN2 causes a new syndrome with craniofacial and central nervous system malformations, and atypical hemolytic uremic syndrome. Clin. Genet. 2022, 101, 346–358. [Google Scholar] [CrossRef]
- Sartain, S.E.; Turner, N.A.; Moake, J.L. Brain microvascular endothelial cells exhibit lower activation of the alternative complement pathway than glomerular microvascular endothelial cells. J. Biol. Chem. 2018, 293, 7195–7208. [Google Scholar] [CrossRef]
- Thergaonkar, R.W.; Narang, A.; Gurjar, B.S.; Tiwari, P.; Puraswani, M.; Saini, H.; Sinha, A.; Varma, B.; Mukerji, M.; Hari, P.; et al. Targeted exome sequencing in anti-factor H antibody negative HUS reveals multiple variations. Clin. Exp. Nephrol. 2018, 22, 653–660. [Google Scholar] [CrossRef]
- Schramm, E.C.; Roumenina, L.T.; Rybkine, T.; Chauvet, S.; Vieira-Martins, P.; Hue, C.; Maga, T.; Valoti, E.; Wilson, V.; Jokiranta, S.; et al. Mapping interactions between complement C3 and regulators using mutations in atypical hemolytic uremic syndrome. Blood 2015, 125, 2359–2369. [Google Scholar] [CrossRef]
- Urban, A.; Kowalska, D.; Stasiłojć, G.; Kuźniewska, A.; Skrobińska, A.; Arjona, E.; Alonso, E.C.; Fenollosa Segarra, M.Á.; Jongerius, I.; Spaapen, R.; et al. Gain-of-Function Mutations R249C and S250C in Complement C2 Protein Increase C3 Deposition in the Presence of C-Reactive Protein. Front. Immunol. 2021, 12, 724361. [Google Scholar] [CrossRef]
- Cheng, C.; Chen, L.; Wen, S.; Lin, Z.; Jiang, X. Case Report: Denys-Drash Syndrome With WT1 Causative Variant Presenting as Atypical Hemolytic Uremic Syndrome. Front. Pediatr. 2020, 8, 605889. [Google Scholar] [CrossRef]
- Zawada, A.M.; Rogacev, K.S.; Schirmer, S.H.; Sester, M.; Böhm, M.; Fliser, D.; Heine, G.H. Monocyte heterogeneity in human cardiovascular disease. Immunobiology 2012, 217, 1273–1284. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.L.; Yeap, W.H.; Tai, J.J.; Ong, S.M.; Dang, T.M.; Wong, S.C. The three human monocyte subsets: Implications for health and disease. Immunol. Res. 2012, 53, 41–57. [Google Scholar] [CrossRef] [PubMed]
- Ziegler-Heitbrock, L.; Ancuta, P.; Crowe, S.; Dalod, M.; Grau, V.; Hart, D.N.; Leenen, P.J.; Liu, Y.J.; MacPherson, G.; Randolph, G.J.; et al. Nomenclature of monocytes and dendritic cells in blood. Blood 2010, 116, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Perez, R.K.; Gordon, M.G.; Subramaniam, M.; Kim, M.C.; Hartoularos, G.C.; Targ, S.; Sun, Y.; Ogorodnikov, A.; Bueno, R.; Lu, A.; et al. Single-cell RNA-seq reveals cell type-specific molecular and genetic associations to lupus. Science 2022, 376, eabf1970. [Google Scholar] [CrossRef]
- Nehar-Belaid, D.; Hong, S.; Marches, R.; Chen, G.; Bolisetty, M.; Baisch, J.; Walters, L.; Punaro, M.; Rossi, R.J.; Chung, C.H.; et al. Mapping systemic lupus erythematosus heterogeneity at the single-cell level. Nat Immunol. 2020, 21, 1094–1106. [Google Scholar] [CrossRef]
- Li, Y.; Ma, C.; Liao, S.; Qi, S.; Meng, S.; Cai, W.; Dai, W.; Cao, R.; Dong, X.; Krämer, B.K.; et al. Combined proteomics and single cell RNA-sequencing analysis to identify biomarkers of disease diagnosis and disease exacerbation for systemic lupus erythematosus. Front. Immunol. 2022, 13, 969509. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lee, T.Y. Revealing the Immune Heterogeneity between Systemic Lupus Erythematosus and Rheumatoid Arthritis Based on Multi-Omics Data Analysis. Int. J. Mol. Sci. 2022, 23, 5166. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Peng, Y.; Li, J.; Zhang, P.; Liu, Z.; Lu, H.; Peng, L.; Zhou, J.; Fei, Y.; Zeng, X.; et al. Single-Cell Sequencing of Immune Cell Heterogeneity in IgG4-Related Disease. Front. Immunol. 2022, 13, 904288. [Google Scholar] [CrossRef]
- Chang, J.G.; Tu, S.J.; Huang, C.M.; Chen, Y.C.; Chiang, H.S.; Lee, Y.T.; Yen, J.C.; Lin, C.L.; Chung, C.C.; Liu, T.C.; et al. Single-cell RNA sequencing of immune cells in patients with acute gout. Sci. Rep. 2022, 12, 22130. [Google Scholar] [CrossRef]
- Hao, Y.; Hao, S.; Andersen-Nissen, E.; Mauck, W.M., III.; Zheng, S.; Butler, A.; Lee, M.J.; Wilk, A.J.; Darby, C.; Zager, M.; et al. Integrated analysis of multimodal single-cell data. Cell 2021, 184, 3573–3587. [Google Scholar] [CrossRef] [PubMed]
- Wolock, S.L.; Lopez, R.; Klein, A.M. Scrublet: Computational Identification of Cell Doublets in Single-Cell Transcriptomic Data. Cell Syst. 2019, 8, 281–291. [Google Scholar] [CrossRef]
- Wolf, F.A.; Angerer, P.; Theis, F.J. SCANPY: Large-scale single-cell gene expression data analysis. Genome Biol. 2018, 19, 15. [Google Scholar] [CrossRef]
- Aran, D.; Looney, A.; Liu, L.; Wu, E.; Fong, V.; Hsu, A.; Chak, S.; Naikawadi, R.P.; Wolters, P.J.; Abate, A.R.; et al. Reference-based analysis of lung single-cell sequencing reveals a transitional profibrotic macrophage. Nat. Immunol. 2019, 20, 163–172. [Google Scholar] [CrossRef]
- Cao, J.; Spielmann, M.; Qiu, X.; Huang, X.; Ibrahim, D.M.; Hill, A.J.; Zhang, F.; Mundlos, S.; Christiansen, L.; Steemers, F.J.; et al. The single-cell transcriptional landscape of mammalian organogenesis. Nature 2019, 566, 496–502. [Google Scholar] [CrossRef]
- Traag, V.A.; Waltman, L.; van Eck, N.J. From Louvain to Leiden: Guaranteeing well-connected communities. Sci. Rep. 2019, 9, 5233. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Guerrero-Juarez, C.F.; Zhang, L.; Chang, I.; Ramos, R.; Kuan, C.H.; Myung, P.; Plikus, M.V.; Nie, Q. Inference and analysis of cell-cell communication using CellChat. Nat. Commun. 2021, 12, 1088. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [PubMed]
- Fay, M.P.; Malinovsky, Y. Confidence intervals of the Mann-Whitney parameter that are compatible with the Wilcoxon-Mann-Whitney test. Stat. Med. 2018, 37, 3991–4006. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case | Age | Gender | TMA Involvement Organ | Treatment | Disease Activity |
|---|---|---|---|---|---|
| a1 | 39 | M | Kidney, brain, lung, heart | PE | Stable |
| a3/a5 | 81 | M | Kidney, heart | a3 before 1st PE a5 after PE | Unstable |
| a4 | 66 | M | Kidney, brain, heart | PE + anti C5 | Stable |
| a7 | 36 | F | Kidney, heart, pancreas, eye | PE + anti C5 | Stable |
| a8 | 33 | F | Kidney, brain, lung, heart | PE | Unstable |
| a9 | 62 | F | Kidney, brain, heart | PE + anti C5 | Unstable |
| a10 | 39 | M | Kidney, brain, lung, heart, eye | PE + anti C5 | Stable |
| a11 | 30 | F | Kidney, brain, lung, heart, eye, bowel | PE + anti C5 | Stable |
| a12 | 42 | F | Kidney, brain, lung, heart, pancreas, liver, eye, skin | PE + anti C5 | Stable |
| a13 | 53 | M | Kidney, brain, heart | PE + anti C5 | Stable |
| a14 | 70 | M | Kidney, brain, heart | PE | Unstable |
| a16 | 62 | F | Kidney, heart | PE | Stable |
| a17 | 49 | F | Kidney, brain, lung, heart, pancreas, liver, eye | PE + anti C5 | Stable |
| Significantly Increased Immune Cell Subclusters in aHUS Patients Compared to Healthy Controls with Correlated Gene Expression Increasing | |||
| Cell Subclusters | p value | Higher expression levels of gene | |
| Classical monocyte subclusters 6 | p < 0.01 | RPS27 | |
| Classical monocyte subclusters 7 | p < 0.05 | IFI27 | |
| Central memory CD8 T-cells subcluster 3 | p < 0.05 | CXCR4 | |
| Non-Vd2 gd T-cells subcluster 4 | p < 0.05 | SYNE2 | |
| Th1 cells subcluster 3 | p < 0.05 | MT-CYB | |
| Th17 cells subcluster 4 | p < 0.05 | MT-ATP6 | |
| Significantly Increased Immune Cell Subclusters in Healthy Controls Compared to aHUS Patients with Correlated Gene Expression Increasing | |||
| Cell Subclusters | p value | Higher expression levels of gene | |
| Central memory CD8 T-cells subcluster 1 | p < 0.05 | EIF3E | |
| Th1 cells subcluster 0 | p < 0.05 | RPS27 | |
| Non classical monocytes subcluster 5 | p < 0.01 | LYPD2 | |
| Terminal effector CD4 T-cells subcluster 3 | p < 0.01 | KLRD1 | |
| Th17 cells subcluster 3 | p < 0.05 | ACTG1, CD52 and LGALS1 | |
| Significantly Increased Immune Cell Subclusters in Unstable aHUS Patients Compared to Stable aHUS Patients with Correlated Gene Expression Increasing | |||
|---|---|---|---|
| Cell Subclusters | p value | Higher expression levels of gene | |
| Classical monocyte subclusters 4 | p < 0.05 | NEAT1, MT-ATP6 and MT-CYB | |
| Central memory CD8-T cells subcluster 2 | p < 0.05 | VIM | |
| Non-Vd2 gd T-cells subcluster 1 | p < 0.05 | ACTG1 | |
| Terminal effector CD8—cells subcluster 3 | p < 0.05 | RPL13 | |
| Terminal effector CD8 T-cells subcluster 5 | p < 0.01 | KLRB1 | |
| Significantly Increased Immune Cell Subclusters in stable aHUS Patients Compared to unstable aHUS Patients with Correlated Gene Expression Increasing | |||
| Cell Subclusters | p value | Higher expression levels of gene | |
| Central memory CD8 T-cells subcluster 1 | p < 0.05 | RPL23 | |
| Non-Vd2 gd T-cell subcluster 0 | p < 0.05 | GZMH | |
| Th1 cells subcluster 0 | p < 0.05 | RPS27, RPS4X | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, I.-R.; Huang, C.-C.; Tu, S.-J.; Wang, G.-J.; Lai, P.-C.; Lee, Y.-T.; Yen, J.-C.; Chang, Y.-S.; Chang, J.-G. Dysregulation of Immune Cell Subpopulations in Atypical Hemolytic Uremic Syndrome. Int. J. Mol. Sci. 2023, 24, 10007. https://doi.org/10.3390/ijms241210007
Chen I-R, Huang C-C, Tu S-J, Wang G-J, Lai P-C, Lee Y-T, Yen J-C, Chang Y-S, Chang J-G. Dysregulation of Immune Cell Subpopulations in Atypical Hemolytic Uremic Syndrome. International Journal of Molecular Sciences. 2023; 24(12):10007. https://doi.org/10.3390/ijms241210007
Chicago/Turabian StyleChen, I-Ru, Chiu-Ching Huang, Siang-Jyun Tu, Guei-Jane Wang, Ping-Chin Lai, Ya-Ting Lee, Ju-Chen Yen, Ya-Sian Chang, and Jan-Gowth Chang. 2023. "Dysregulation of Immune Cell Subpopulations in Atypical Hemolytic Uremic Syndrome" International Journal of Molecular Sciences 24, no. 12: 10007. https://doi.org/10.3390/ijms241210007
APA StyleChen, I.-R., Huang, C.-C., Tu, S.-J., Wang, G.-J., Lai, P.-C., Lee, Y.-T., Yen, J.-C., Chang, Y.-S., & Chang, J.-G. (2023). Dysregulation of Immune Cell Subpopulations in Atypical Hemolytic Uremic Syndrome. International Journal of Molecular Sciences, 24(12), 10007. https://doi.org/10.3390/ijms241210007