
Reviewing the Regulators of COL1A1

 , ,
, ,  ,
,
Abstract
1. Introduction
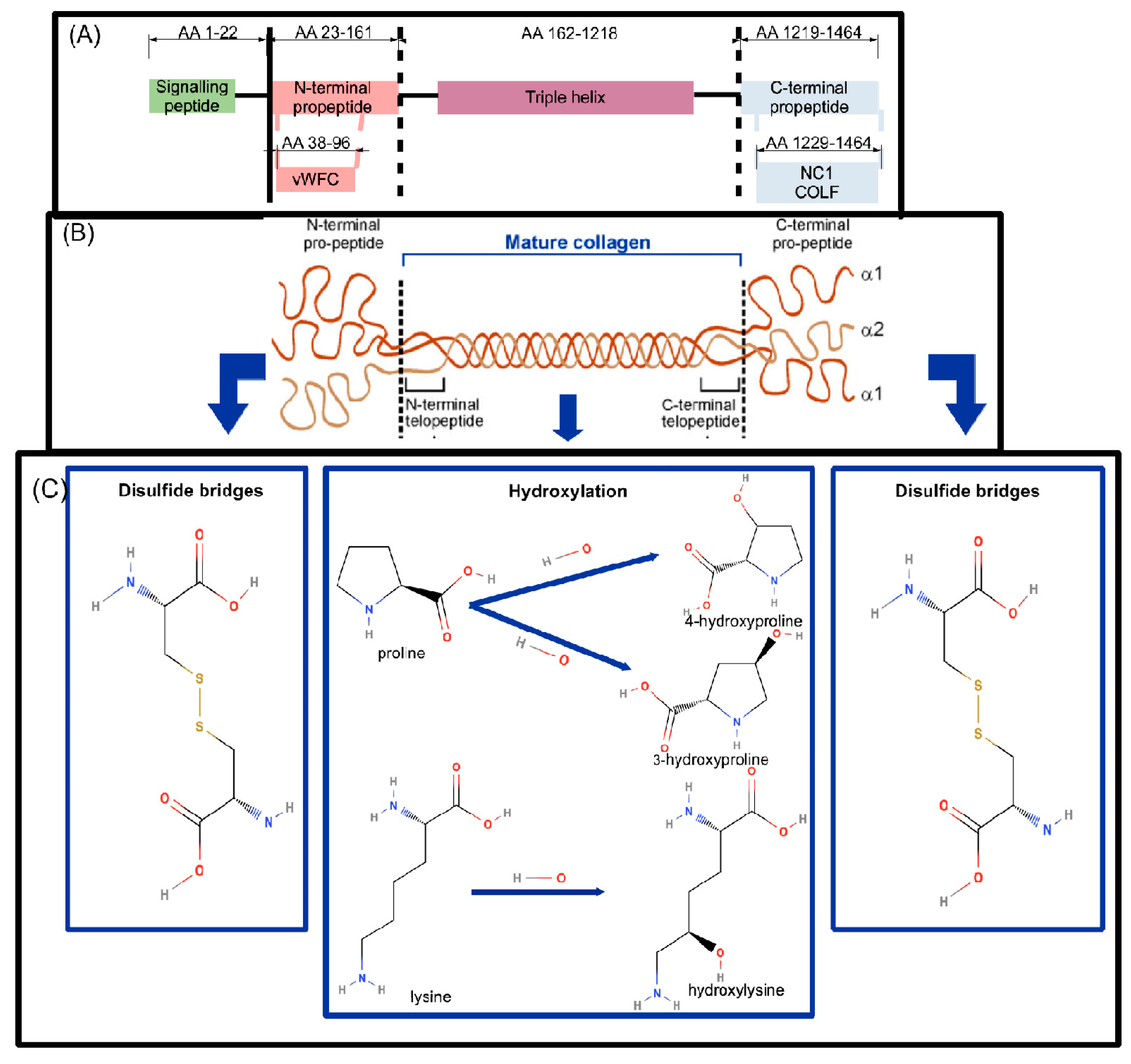
COL1A1 Synthesis and Basic Structure

2. Literature Search and Results
2.1. Literature Search
2.2. Regulators of COL1A1 and COLα1 (I)
2.2.1. COL1A1 Gene Epigenetic and Transcriptional Regulators
2.2.2. COL1A1 Regulation on the Post-Transcriptional Level
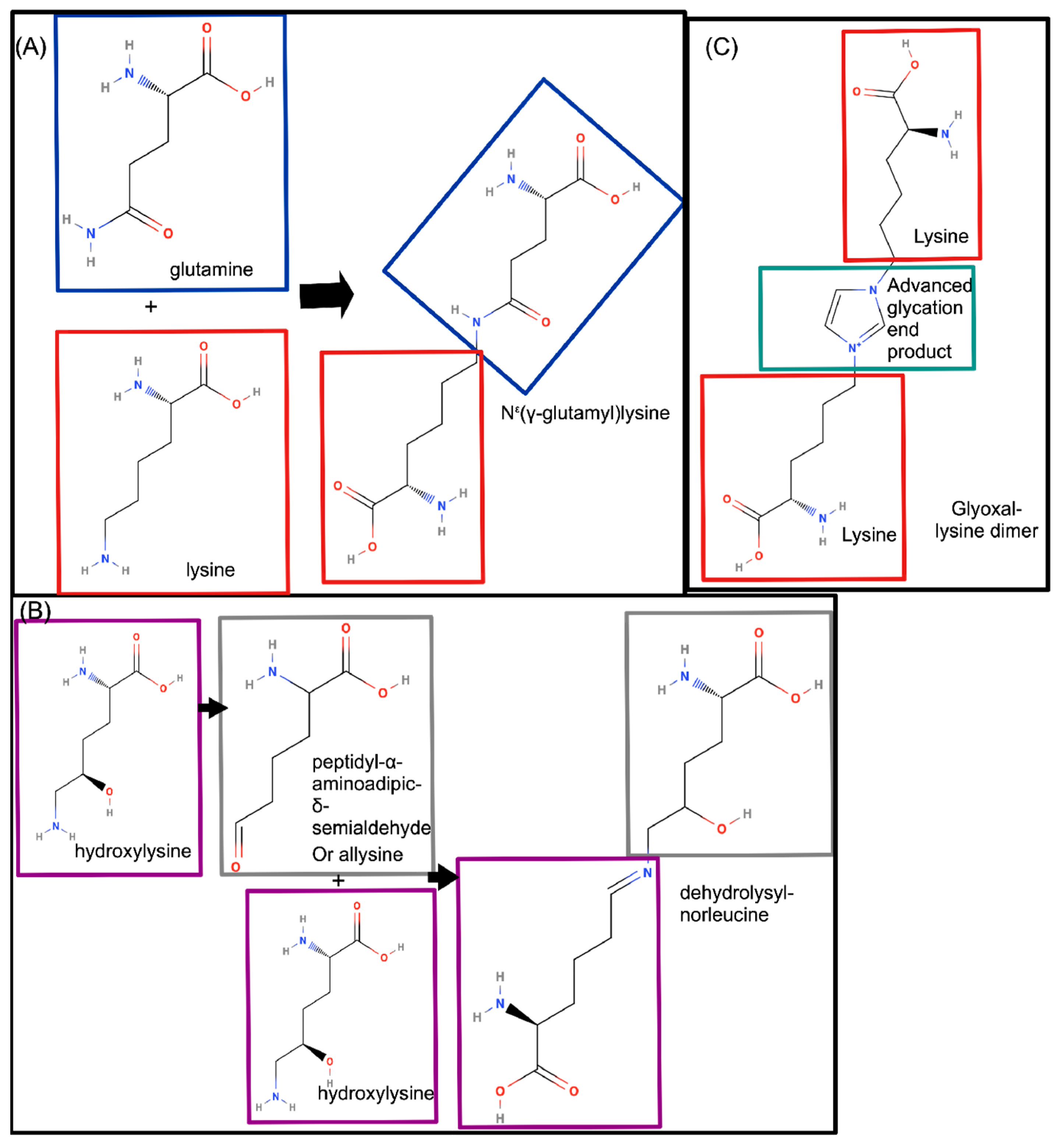
2.2.3. COLα1 (I) Regulation by Post-Translational Modifications and Cleavage
2.3. COLα1 (I)-Cell Receptor Interactions
3. Conclusions: Outlook and Therapeutic Implications
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nucleotides | ||
| One-Letter Code | Full Name | |
| C | Cytosine | |
| T | Thymidine | |
| Amino Acids | ||
| Three-letter code | One-letter code | Full name |
| Ala | A | Alanine |
| Cys | C | Cysteine |
| Gly | G | Glycine |
| Lys | K | Lysine |
| Pro | P | Proline |
| Ser | S | Serine |
| Thr | T | Threonine |
| Regulators | ||
| Abbreviation | In full | |
| ACTA2 | Actin Alpha 2 (gene, encoding αSMA) | |
| ADAMTS2 | A Disintegrin And Metalloproteinase with Thrombospondin motifS 2 | |
| AGE | Advanced Glycation End products | |
| AKT | Also known as Protein Kinase B (PKB) | |
| ANRIL | Antisense Non-coding RNA in the INK4 Locus | |
| AP2 | Activating enhancer binding Protein 2 | |
| BMP1 | Bone Morphogenic Protein 1 | |
| CCR2 | C-C chemokine Receptor 2 | |
| CD | Cluster of Differentiation | |
| COL1 | Type I collagen | |
| COL1A1 | Collagen type I alpha 1 chain (gene) | |
| COL1A2 | Collagen type I alpha 2 chain (gene) | |
| COL2 | Type II collagen | |
| COL3 | Type III collagen | |
| COL4A1 | Collagen type IV alpha 1 chain (gene) | |
| COL7 | Type VII collagen | |
| COLα1 (I) | Type I collagen, alpha 1 chain (protein, encoded by COL1A1) | |
| COLα1 (III) | Type III collagen, alpha 1 chain (protein, encoded by COL3A1) | |
| COLα2 (I) | Type I collagen, alpha 2 chain (protein, encoded by COL1A2) | |
| COX | Cyclo-Oxygenase 2 | |
| CRTAP | Cartilage-Associated Protein | |
| CTGF | Connective Tissue Growth Factor | |
| CXCR4 | C-X-C chemokine Receptor type 4 | |
| Cyr61 | Cysteine-rich angiogenic inducer 61 | |
| DDR | Discoidin Doman Receptor | |
| DNMT1 | DNA Methyl Transferase 1 | |
| DNMT3A | DNA Methyl Transferase 3A | |
| ERK | Extracellular signal Related Kinase | |
| FAK | Focal Adhesion Kinase | |
| GCN2 | General Control Nonderepressible 2 | |
| GRE | Glucocorticoid Response Element | |
| Grm78 | Glucose-Regulated Protein 78 | |
| HDAC1 | Histone Deacetylase 1 | |
| HIF1AN | Hypoxia Inducible Factor 1 subunit alpha inhibitor | |
| HIF1α | Hypoxia Inducible Factor 1 Subunit Alpha | |
| IFNγ | Interferon γ | |
| IgG4 | Immunoglobulin G4 | |
| IL-31 | Interleukin 31 | |
| JNK | c-Jun terminal kinase | |
| KDM1A | Lysine (K)-specific demethylase 1A or LSD1 | |
| KLF6 | Krüppel-like Factor 6 | |
| KLF11 | Krüppel-like Factor 11 | |
| LAIR | Leukocyte-Associated Immunoglobulin-like Receptor | |
| LOX | Lysyl Oxidase | |
| MAPK | Mitogen-Activated Protein Kinase | |
| MERTK | Mer Tyrosine Kinase | |
| miR | microRNA | |
| MMP-1 | Matrix MetalloProteinase 1 | |
| MMP-2 | Matrix MetalloProteinase 2 | |
| MMP-8 | Matrix MetalloProteinase 8 | |
| MMP-9 | Matrix MetalloProteinase 9 | |
| mTOR | Mammalian Target of Rapamycin | |
| Myo1c | Myosin1c | |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells | |
| NOTCH 2 | Neurogenic locus notch homolog protein 2 | |
| NR4A1 | Nuclear Receptor Subfamily 4 Group A Member 1 (also known as TR3, Nur77m, NGF-IB) | |
| OGR1 | Ovarian cancer G-protein coupled Receptor-1 | |
| P3H1 | Prolyl-3-hydroxylase | |
| PCP | Planar Cell Polarity | |
| PCPE-1 | Pro-collagen C-Proteinase Enhancer | |
| PD-1 | Programmed cell Death-1 | |
| PD-L1 | Programmed cell Death Ligand-1 | |
| PDGFC | Platelet Derived Growth Factor C | |
| PDI | Protein Disulfide Isomerase | |
| PFAR | Long non-coding RNA Protein Folding Activity of the Ribosome | |
| PI3K | PhosphoInositede 3 Kinase | |
| PKB | Protein Kinase B or Akt | |
| PKCε | Protein Kianse C ε | |
| PPIB | Peptidyl-prolyl cis-transisomerase cyclophilin B | |
| PTEN | Phosphatase and TENsin homolog | |
| Rac1-GTP | Guanosine Triphosphate (Rac1-GTP) | |
| RAGE | Receptor for Advanced Glycation End products | |
| RCOR1 | REST CORepressor-1 (also known as CoREST) | |
| RelA | Rel-associated protein or p65 | |
| REST | RE1 Silencing Transcription Factor | |
| RUNX2 | Runt-related transcription factor 2 | |
| SAM | S-adenosyl-L-Methionine | |
| Serpine1 | Serpin family E member 1 | |
| SIN3A | Swi-Indpendent transcription regulator family member A | |
| SLPI | Secretory Leukocyte Protease Inhibitor | |
| SMAD 2/3 | SMA and MAD related protein 2/3 | |
| SMAD 4 | SMA and MAD related protein 4 | |
| SMAD 6 | SMA and MAD related protein 6 | |
| SMAD 7 | SMA and MAD related protein 7 | |
| SP1 | Specific Protein 1 | |
| STAT3 | Signal Transducer and Activator of Transcription 3 | |
| TAZ | Tafazzin | |
| TENT5A | Terminal Nucleotidyltransferase 5A | |
| TGFβ | Tumour Growth Factor β | |
| TGFβR | TGFβ receptor | |
| uPARAP | Urokinase Plasminogen Activator Receptor-Associated Protein | |
| VCE | Viral Core Enhancer motif | |
| YAP | Yes-associated protein | |
| ZEB1 | Zinc finger E-box-Binding homeobox 1 | |
| αSMA | α-Smooth Muscle Actin (protein, encoded by ACTA2 gene) | |
| Other abbreviations | ||
| Abbreviation | In full | |
| AA | Amino Acid | |
| ARDS | Acute Respiratory Distress Syndrome | |
| bp | Base pairs | |
| CCl4 | Carbon tetrachloride | |
| circRNA | Circular RiboNucleic Acid | |
| DNA | DeoxyRibonucleic Acid | |
| ECM | Extracellular matrix | |
| FDA | US Food and Drug Administration | |
| H2O2 | Hydrogen Peroxide | |
| HSC | Hepatic Stellate Cell | |
| lncRNA | Long Non-Coding RNA | |
| MeSH | Medical Subject Headings | |
| MMPs | Matrix-metalloproteinases | |
| mRNA | messenger Ribonucleic Acid | |
| NAFLD | Non-Alcoholic Fatty Liver Disease | |
| NC1 | C-terminal non-collagenous domain; also denoted as COLF | |
| OI | Osteogenesis Imperfecta | |
| PBMC | Peripheral Blood Mononuclear cells | |
| PTMs | Post-Translational Modifications | |
| RNA | RiboNucleic Acid | |
| TIMP | Tissue Inhibitors of Matrix metalloProteinases | |
| vWFC | Von Willebrand factor type C domain | |
| Search Query | Number of Articles Retrieved | Number of Articles Included | Number of Reviews Excluded |
|---|---|---|---|
| (“col1a1”[All Fields]) AND (“signalling pathways”[All Fields]) | 15 | 1 | 2 |
| (“col1a1 protein human”[All Fields]) AND (“fibrosis”[All Fields]) | 34 | 9 | 2 |
| (“col1a1 protein human”[All Fields]) AND (“disease”[All Fields]) | 71 | 2 | 6 |
| (“col1a1”[All Fields]) AND (“protein function”[All Fields]) | 3 | 0 | 0 |
| (“col1a1”[All Fields]) AND (“protein structure”[All Fields]) | 33 | 2 | 2 |
| ((“col1a1”[All Fields]) AND (“post-translational protein processing”[All Fields])) AND (“fibrosis”[All Fields]) | 3 | 2 | 0 |
| (“col1a1”[All Fields]) AND (“post-translational protein processing”[All Fields]) | 25 | 1 | 2 |
| “col1a1 protein human”[All Fields] | 219 | 4 | 7 |
| (“collagen type i alpha 1”[All Fields]) AND (“signalling pathways”[All Fields]) | 5 | 0 | 1 |
| (“collagen type i alpha 1”[All Fields]) AND (“protein structure”[All Fields]) | 32 | 0 | 4 |
| (“collagen type i alpha 1”[All Fields]) AND (“post-translational protein processing”[All Fields]) AND (“fibrosis”[All Fields]) | 3 | 1 | 0 |
| (“collagen type i alpha 1”[All Fields]) AND (“post-translational protein processing”[All Fields]) | 20 | 1 | 0 |
| (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) | 290 * | 34 | 12 |
| ((“col1a1”[All Fields]) OR (“collagen type i alpha 1”[All Fields])) AND ((“protease”[All Fields]) OR (“proteolytic cleavage”[All Fields])) | 41 | 5 | 1 |
| Similar articles listed in PubMed for PMID: 9514209 (PMID 9514209: Collagen cross-links in mineralizing tissues: a review of their chemistry, function, and clinical relevance, Knott. L. and Bailey AJ., Bone, 1998) | 190 | 2 | 27 |
| Total | 984 | 64 | 66 |
| Title | DOI | Search Quote |
|---|---|---|
| Abnormal Downregulation of Caveolin-3 Mediates the Pro-Fibrotic Action of MicroRNA-22 in a Model of Myocardial Infarction | 10.1159/000487732 | (“col1a1”[All Fields]) AND (“protein structure”[All Fields]) |
| AGE-RAGE signal generates a specific NF-κB RelA “barcode” that directs collagen I expression | 10.1038/srep18822 | (“collagen type i alpha 1”[All Fields]) AND (“post-translational protein processing”[All Fields]) |
| circHIPK3 regulates lung fibroblast-to-myofibroblast transition by functioning as a competing endogenous RNA | 10.1038/s41419-019-1430-7 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Circular RNA hsa_circRNA_002178 silencing retards breast cancer progression via microRNA-328-3p-mediated inhibition of COL1A1 | 10.1111/jcmm.14875 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| COL1A1 promotes metastasis in colorectal cancer by regulating the WNT/PCP pathway | 10.3892/mmr.2018.8533 | “col1a1 protein human”[All Fields] |
| Collagen 1A1 (COL1A1) promotes metastasis of breast cancer and is a potential therapeutic target | “col1a1 protein human”[All Fields] | |
| Collagen 1a1 Expression by Airway Macrophages Increases In Fibrotic ILDs and Is Associated With FVC Decline and Increased Mortality | 10.3389/fimmu.2021.645548 | (“col1a1 protein human”[All Fields]) AND (“fibrosis”[All Fields]) |
| Collagen Characterization in a Model of Nonalcoholic Steatohepatitis with Fibrosis; A Call for Development of Targeted Therapeutics | 10.3390/molecules26113316 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Collagenolysis-dependent DDR1 signalling dictates pancreatic cancer outcome | 10.1038/s41586-022-05169-z | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Cytoplasmic polyadenylation by TENT5A is required for proper bone formation | 10.1016/j.celrep.2021.109015 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Deficiency of MicroRNA miR-1954 Promotes Cardiac Remodeling and Fibrosis | 10.1161/JAHA.119.012880 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Differential regulation of collagen, lysyl oxidase and MMP-2 in human periodontal ligament cells by low- and high-level mechanical stretching | 10.1111/jre.12028 | ((“col1a1”[All Fields]) OR (“collagen type i alpha 1”[All Fields])) AND ((“protease”[All Fields]) OR (“proteolytic cleavage”[All Fields])) |
| Distinct post-translational features of type I collagen are conserved in mouse and human periodontal ligament | 10.1111/jre.12475 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Downregulation of miR-141 deactivates hepatic stellate cells by targeting the PTEN/AKT/mTOR pathway | 10.3892/ijmm.2020.4578 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Dual inhibition of αvβ6 and αvβ1 reduces fibrogenesis in lung tissue explants from patients with IPF | 10.1186/s12931-021-01863-0 | (“col1a1 protein human”[All Fields]) AND (“fibrosis”[All Fields]) |
| Endo180 binds to the C-terminal region of type I collagen | 10.1074/jbc.M501155200 | (“col1a1”[All Fields]) AND (“protein structure”[All Fields]) |
| Epigenetic silencing of LncRNA ANRIL enhances liver fibrosis and HSC activation through activating AMPK pathway | 10.1111/jcmm.14987 | (“col1a1”[All Fields]) AND (“signalling pathways”[All Fields]) |
| FAM13A as potential therapeutic target in modulating TGF-β-induced airway tissue remodeling in COPD | 10.1152/ajplung.00477.2020 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| GCN2 Regulates ATF3-p38 MAPK Signaling Transduction in Pulmonary Veno-Occlusive Disease | 10.1177/10742484211015535 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Hepatocyte Stress Increases Expression of Yes-Associated Protein and Transcriptional Coactivator With PDZ-Binding Motif in Hepatocytes to Promote Parenchymal Inflammation and Fibrosis | 10.1002/hep.30928 | (“col1a1 protein human”[All Fields]) AND (“fibrosis”[All Fields]) |
| High collagen density augments mTOR-dependent cancer stem cells in ERα+ mammary carcinomas, and increases mTOR-independent lung metastases | 10.1016/j.canlet.2018.06.025 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Identification of COL1A1 associated with immune infiltration in brain lower grade glioma | 10.1371/journal.pone.0269533 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Immune modulating effects of additional supplementation of estradiol combined with testosterone in murine testosterone-deficient NAFLD model | 10.1152/ajpgi.00310.2019 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Insight into the Pathology of a COL1A1 Signal Peptide Heterozygous Mutation Leading to Severe Osteogenesis Imperfecta | 10.1007/s00223-017-0359-z | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Interaction of reactive astrocytes with type I collagen induces astrocytic scar formation through the integrin-N-cadherin pathway after spinal cord injury | 10.1038/nm.4354 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Intestinal Activation of pH-Sensing Receptor OGR1 [GPR68] Contributes to Fibrogenesis | 10.1093/ecco-jcc/jjy118 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Involvement of circHIPK3 in the pathogenesis of diabetic cardiomyopathy in mice | 10.1007/s00125-020-05353-8 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Isoliquiritigenin inhibits TGF-β1-induced fibrogenesis through activating autophagy via PI3K/AKT/mTOR pathway in MRC-5 cells | 10.1093/abbs/gmaa067 | (“col1a1 protein human”[All Fields]) AND (“fibrosis”[All Fields]) |
| LncRNA LINC00511 promotes COL1A1-mediated proliferation and metastasis by sponging miR-126-5p/miR-218-5p in lung adenocarcinoma | 10.1186/s12890-022-02070-3 | (“col1a1 protein human”[All Fields]) AND (“disease”[All Fields]) |
| LncRNA PFAR contributes to fibrogenesis in lung fibroblasts through competitively binding to miR-15a | 10.1042/BSR20190280 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| MERTK +/hi M2c Macrophages Induced by Baicalin Alleviate Non-Alcoholic Fatty Liver Disease | 10.3390/ijms221910604 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Microenvironment remodeled by tumor and stromal cells elevates fibroblast-derived COL1A1 and facilitates ovarian cancer metastasis | 10.1016/j.yexcr.2020.112153 | “col1a1 protein human”[All Fields] |
| MicroRNA expression signature and the therapeutic effect of the microRNA-21 antagomir in hypertrophic scarring | 10.3892/mmr.2017.6104 | (“col1a1 protein human”[All Fields]) AND (“fibrosis”[All Fields]) |
| MicroRNA-29b upregulation improves myocardial fibrosis and cardiac function in myocardial infarction rats through targeting SH2B3 | 10.26355/eurrev_201911_19581 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| MicroRNA-98 inhibits the cell proliferation of human hypertrophic scar fibroblasts via targeting Col1A1 | 10.1186/s40659-017-0127-6 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| MicroRNA-326 inhibits endometrial fibrosis by regulating TGF-β1/Smad3 pathway in intrauterine adhesions | 10.3892/mmr.2018.9187 | (“col1a1 protein human”[All Fields]) AND (“fibrosis”[All Fields]) |
| MiR-16-5p suppresses myofibroblast activation in systemic sclerosis by inhibiting NOTCH signaling | 10.18632/aging.202308 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| miR-185 and miR-29a are similarly expressed in the bronchoalveolar lavage cells in IPF and lung cancer but common targets DNMT1 and COL1A1 show disease specific patterns | 10.3892/mmr.2018.8778 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| MiR-29 mediates TGFβ1-induced extracellular matrix synthesis through activation of PI3K-AKT pathway in human lung fibroblasts | 10.1002/jcb.24474 | ((“col1a1”[All Fields]) AND (“post-translational protein processing”[All Fields])) AND (“fibrosis”[All Fields]) |
| MiR-29a Assists in Preventing the Activation of Human Stellate Cells and Promotes Recovery From Liver Fibrosis in Mice | 10.1038/mt.2016.127 | (“col1a1 protein human”[All Fields]) AND (“fibrosis”[All Fields]) |
| MiR-590-3p regulates proliferation, migration and collagen synthesis of cardiac fibroblast by targeting ZEB1 | 10.1111/jcmm.14704 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Mutations in FKBP10 cause recessive osteogenesis imperfecta and Bruck syndrome | 10.1002/jbmr.250 | (“col1a1”[All Fields]) AND (“post-translational protein processing”[All Fields]) |
| N-Homocysteinylation impairs collagen cross-linking in cystathionine β-synthase-deficient mice: a novel mechanism of connective tissue abnormalities | 10.1096/fj.201600539 | “col1a1 protein human”[All Fields] |
| NR4A1 is Involved in Fibrogenesis in Ovarian Endometriosis | 10.1159/000488838 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| PGE2 induces apoptosis of hepatic stellate cells and attenuates liver fibrosis in mice by downregulating miR-23a-5p and miR-28a-5p | 10.1016/j.bbadis.2017.11.001 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Progressive Fibrosis: A Progesterone- and KLF11-Mediated Sexually Dimorphic Female Response | 10.1210/en.2017-00171 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Proteasome blockade exerts an antifibrotic activity by coordinately down-regulating type I collagen and tissue inhibitor of metalloproteinase-1 and up-regulating metalloproteinase-1 production in human dermal fibroblasts | 10.1096/fj.05-4870fje | ((“col1a1”[All Fields]) AND (“post-translational protein processing”[All Fields])) AND (“fibrosis”[All Fields]) |
| Role and mechanism of microRNA-548c-3p/c-Myb in myocardial infarction fibrosis in rats | 10.26355/eurrev_201906_18081 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Role of Protease-Activated Receptor 2 in the development of lower urinary tract dysfunction | 10.1016/j.juro.2016.01.106 | ((“col1a1”[All Fields]) OR (“collagen type i alpha 1”[All Fields])) AND ((“protease”[All Fields]) OR (“proteolytic cleavage”[All Fields])) |
| RUNX2 promotes malignant progression in gastric cancer by regulating COL1A1 | 10.3233/CBM-200472 | (“col1a1 protein human”[All Fields]) AND (“disease”[All Fields]) |
| S-adenosyl-L-methionine inhibits collagen secretion in hepatic stellate cells via increased ubiquitination | 10.1111/j.1478-3231.2011.02512.x | (“collagen type i alpha 1”[All Fields]) AND (“post-translational protein processing”[All Fields]) AND (“fibrosis”[All Fields]) |
| Secretory leukocyte protease inhibitor gene deletion alters bleomycin-induced lung injury, but not development of pulmonary fibrosis | 10.1038/labinvest.2016.40 | ((“col1a1”[All Fields]) OR (“collagen type i alpha 1”[All Fields])) AND ((“protease”[All Fields]) OR (“proteolytic cleavage”[All Fields])) |
| Sennoside A prevents liver fibrosis by binding DNMT1 and suppressing DNMT1-mediated PTEN hypermethylation in HSC activation and proliferation | 10.1096/fj.202000494RR | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Substitution of murine type I collagen A1 3-hydroxylation site alters matrix structure but does not recapitulate osteogenesis imperfecta bone dysplasia | 10.1016/j.matbio.2020.02.003 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Suppression of pancreatic ductal adenocarcinoma growth and metastasis by fibrillar collagens produced selectively by tumor cells | 10.1038/s41467-021-22490-9 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Syndecan-4 Protects the Heart From the Profibrotic Effects of Thrombin-Cleaved Osteopontin | 10.1161/JAHA.119.013518 | (“col1a1 protein human”[All Fields]) AND (“fibrosis”[All Fields]) |
| TGF-β1 protein trap AVID200 beneficially affects hematopoiesis and bone marrow fibrosis in myelofibrosis | 10.1172/jci.insight.145651 | (“col1a1 protein human”[All Fields]) AND (“fibrosis”[All Fields]) |
| The let-7/Lin28 axis regulates activation of hepatic stellate cells in alcoholic liver injury | 10.1074/jbc.M116.773291 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| The miRNA-184 drives renal fibrosis by targeting HIF1AN in vitro and in vivo | 10.1007/s11255-018-2025-4 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| The prognostic potential of alpha-1 type I collagen expression in papillary thyroid cancer | 10.1016/j.bbrc.2019.04.119 | (“collagen type i alpha 1”[All Fields]) AND (“disease”[All Fields]) |
| Transcriptional regulation of matrix metalloproteinase-1 and collagen 1A2 explains the anti-fibrotic effect exerted by proteasome inhibition in human dermal fibroblasts | 10.1186/ar2991 | ((“col1a1”[All Fields]) OR (“collagen type i alpha 1”[All Fields])) AND ((“protease”[All Fields]) OR (“proteolytic cleavage”[All Fields])) |
| Umbilical cord/placenta-derived mesenchymal stem cells inhibit fibrogenic activation in human intestinal myofibroblasts via inhibition of myocardin-related transcription factor A | 10.1186/s13287-019-1385-8 | Similar articles listed in PubMed for PMID: 9514209 (PMID 9514209: Collagen cross-links in mineralizing tissues: a review of their chemistry, function, and clinical relevance, Knott. L. and Bailey AJ., Bone, 1998) |
| Zonal regulation of collagen-type proteins and posttranslational modifications in prostatic benign and cancer tissues by imaging mass spectrometry | 10.1002/pros.24031 | Similar articles listed in PubMed for PMID: 9514209 (PMID 9514209: Collagen cross-links in mineralizing tissues: a review of their chemistry, function, and clinical relevance, Knott. L. and Bailey AJ., Bone, 1998) |
| Main Regulator | Level of Regulation | Direct/ Indirect 1 | Effect | Reference | Disease | System | Downstream Pathways |
|---|---|---|---|---|---|---|---|
| DNMT1 | Epigenetic | I | Upregulation | [39,41] | Liver fibrosis (CCl4), idiopathic pulmonary fibrosis and lung cancer | Mouse model/rat hepatic stellate, human BAL cells (primary) | PTEN inhibited by DNMT1, leading to increased pAkt and pERK |
| DNMT3A | Epigenetic | I | Upregulation | [40] | Liver fibrosis (CCl4) | Rat model, human samples | DNMT3A inhibited by ANRIL |
| NF-κB | Transcriptional | D | Upregulation | [44] | Ageing | Murine endothelial fibroblasts | |
| Smad 2/3 2 | Transcriptional | D | Upregulation | [45,55,58,59,60,104] | Heart fibrosis, hepatitis C infection Myelofibrosis, pulmonary fibrosis | Mouse model, human HSC cell line Mononuclear cells, hepatocytes, mouse model (ARDS), mouse model (bleomycin) | JNK, Myo1c, integrin αvβ6/αvβ1: phosphorylate Smad2/3 Activate: MAPK, FAK, p38 kinase, PI3K/Akt/mTOR |
| NR4A1 | Transcriptional | D | Downregulation | [42] | Endometriosis | Human endometrial stromal cells | Corepressors Sp1, swi-independent-3 transcription regulator family member A (SIN3A), REST corepressor-1 (RCOR1 or CoREST), histone deacetylases (HDAC) and Lysine(K)-specific demethylase 1A (KDM1A or LSD1) Activated by TGFβ |
| SP1 | Transcriptional | D | Downregulation | [46,47] | Systemic sclerosis | TGFβ-challenged dermal fibroblasts | Activated by TGFβ |
| KLF6 | Transcriptional | D | Downregulation | [48] | Liver fibrosis | Mouse HSC, mouse model liver fibrosis | |
| KLF11 | Transcriptional | I | Upregulation (progesterone), Downregulation (estrogen, testosterone) | [68,69] | Liver fibrosis | Primary human peritoneal mesothelial cells / mouse model liver fibrosis | |
| c-Myb | Transcriptional | I | Upregulation | [49] | Cardiac fibrosis | Rat model | |
| NOTCH2 | Transcriptional | I | Upregulation | [50] | Systemic sclerosis | Blood samples, skin fibroblasts | Transcriptional activator complex with RBPJ |
| HIF1α | Transcriptional | I | Upregulation | [51] | Kidney fibrosis | Rat fibroblast cell line | HIF1α is inhibited by HIF1AN |
| YAP/TAZ | Transcriptional | I | Upregulation | [52] | Liver fibrosis | Mouse model | |
| ZEB1 | Transcriptional | I | Upregulation | [53] | Cardiac fibrosis (acute myocardial infarction-induced) | Human cardiac fibroblasts | Regulated by miR-590-3p |
| TENT5A | Post-transcriptional | D | Downregulation | [80] | Osteogenesis imperfecta | TENT5A knock-out mouse model | Prevention of polyadenylation reduces turnover time |
| LIN28B | Post-transcriptional | I | Downregulation | [81] | Alcohol induced liver fibrosis | LIN28B knock-out mouse model | |
| miR-98 | Post-transcriptional | D | Downregulation | [82] | Hypertrophic scar formation | Human hypertrophic scar fibroblasts | |
| miR-126-5p miR-218-5p | Post-transcriptional | D | Downregulation | [83] | Lung adenocarcinoma | Human lung adenocarcinoma cell lines | Downregulated by linc00511 |
| miR-328-3p | Post-transcriptional | D | Downregulation | [84] | Breast cancer | Primary human breast cancer cells | Downregulated by has-circRNa-002178 |
| miR-338-3p | Post-transcriptional | D | Downregulation | [85] | Pulmonary fibrosis | Mouse model, human lung fibroblast cell line, human embryonic kidney cell line | Downregulation by circHIPK3 |
| miR-29b-3p | Post-transcriptional | D | Downregulation | [86] | Diabetic cardiomyopathy | Mouse model | Downregulation by circHIPK3 (predicted) |
| miR-29b | Post-transcriptional | I | Downregulation | [87] | Cardiac fibrosis | Rat model | SH2B3 |
| miR-29a | Post-transcriptional | I | Downregulation | [88] | Liver fibrosis | Mouse models (2) | |
| miR-29a, miR-29b, miR-29c | Post-transcriptional | I | Downregulation | [89] | Pulmonary fibrosis | Pulmonary fibroblasts | |
| miR-21 | Post-transcriptional | I | Downregulation | [90] | Systemic sclerosis | Dermal fibroblasts | Target: PTEN (predicted) |
| miR-141 | Post-transcriptional | I | Downregulation | [91] | Chronic liver disease | Human hepatic stellate cell line | Target: PTEN |
| miR-23a miR-28a | Post-transcriptional | I | Downregulation | [93] | Liver fibrosis | Mouse model | Target: TGFβ Upregulation: PGE2 |
| miR-1954 | Post-transcriptional | I | Downregulation | [94] | Cardiac fibrosis | Mouse model | Thrombospondin-1 |
| PFAR | Post-transcriptional | I | Upregulation | [95] | Lung fibrosis | Mouse lung fibroblasts | Induces YAP |
| Lysyl oxidase | Post-translational | D | Upregulation | [4,96] | Bruck syndrome, mechanical stretching | Bone, periodontal osteoblastic cells | Associated with cross-linking |
| Prolyl hydroxylation complex | Post-translational | D | Upregulation | [14] | Osteogenesis imperfecta (OI) | Primary osteoblasts from OI mouse model | Associated with cross-linking |
| N-homocysteinylation | Post-translational | D | Downregulation | [98] | Cystathionine β-synthase deficiency | Mouse model of cystathionine β-synthase deficiency | Prevents cross-linking |
| Polyubiquitination | Post-translational | D | Downregulation | [99] | Liver fibrosis | Rat hepatic stellate cells treated with S-adenosyl-L-Methionine | proteasomal degradation |
| Gly22Arg mutation | Post-translational | D | Downregulation | [100] | Gly22Arg mutation in OI | Primary human fibroblasts | Prevents splicing |
| Incorrect splicing (N-terminal propeptide) by BMP1 | Post-translational | D | Upregulation | [101] | Human pancreatic adeno-carcinoma | Primary human pancreatic adenocarcinoma samples | Incorrect splicing (N-terminal propeptide) |
| RGDSLAYGLR osteopontin derived peptide | Post-translational | I | Upregulation | [18,67] | Heart fibrosis | Mouse model of pressure overloaded heart | Cleavage by thrombin. Protection from cleavage by Syndecan 4 |
| Secretory Leukocyte Protease Inhibitor | Post-translational | I | Downregulation | [102] | Pulmonary fibrosis | Mouse model | Knock-out of SLPI: protection against Col1a1 increase in pulmonary fibrosis |
| Protease-activated receptor 2 | Post-translational | I | Upregulation | [103] | Chronic prostatitis | Mouse model |
| Main Regulator | Direct/ Indirect 1 | Effect | Reference | Disease | System | Downstream Pathways |
|---|---|---|---|---|---|---|
| miR-326 | Direct | TGFβ mRNA inhibition | [92] | Endometriosis | Primary endometrial stromal cells | |
| MERTK | Indirect | TGFβ secretion | [61] | NAFLD | Mouse model | |
| PKCε | Indirect | TGFβ activation | [62] | Heart fibrosis | Rat model | PKCε is downregulated by caveolin3 |
| RGDSLAYGLR (osteopontin derived peptide) | Direct | TGFβ receptor activation | [18,67] | Heart fibrosis, idiopathic pulmonary fibrosis | Mouse model, alveolar macrophages (primary) | |
| JNK | Direct | Smad2/3 phosphorylation | [55] | myelofibrosis | Mononuclear cells | |
| integrin αvβ6/αvβ1 | Direct | Smad2/3 phosphorylation | [104] | Pulmonary fibrosis | Mouse model | |
| OGR1 | Indirect | Smad2/3 phosphorylation (reduced) | [63] | Pulmonary fibrosis/fibrosis in Crohn’s disease | Human pulmonary fibroblasts (primary), mouse model | |
| miR-23a and miR-28a | Indirect | Smad2/3 phosphorylation (reduced) | [93] | Liver fibrosis | Mouse model | |
| PI3K/Akt/mTOR | indirect | Downstream activated pathway | [60] | Pulmonary fibrosis | Human lung fibroblasts (primary) |
References
- Ricard-Blum, S. The Collagen Family. Cold Spring Harb. Perspect. Biol. 2011, 3, a004978. [Google Scholar] [CrossRef] [PubMed]
- Le Goff, C.; Somerville, R.P.; Kesteloot, F.; Powell, K.; Birk, D.E.; Colige, A.C.; Apte, S.S. Regulation of Procollagen Amino-Propeptide Processing during Mouse Embryogenesis by Specialization of Homologous ADAMTS Proteases: Insights on Collagen Biosynthesis and Dermatosparaxis. Development 2006, 133, 1587–1596. [Google Scholar] [CrossRef] [PubMed]
- Barnes, A.M.; Ashok, A.; Makareeva, E.N.; Brusel, M.; Cabral, W.A.; Weis, M.; Moali, C.; Bettler, E.; Eyre, D.R.; Cassella, J.P.; et al. COL1A1 C-Propeptide Mutations Cause ER Mislocalization of Procollagen and Impair C-Terminal Procollagen Processing. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 2210–2223. [Google Scholar] [CrossRef]
- Kelley, B.P.; Malfait, F.; Bonafe, L.; Baldridge, D.; Homan, E.; Symoens, S.; Willaert, A.; Elcioglu, N.; Van Maldergem, L.; Verellen-Dumoulin, C.; et al. Mutations in FKBP10 Cause Recessive Osteogenesis Imperfecta and Bruck Syndrome. J. Bone Min. Res. 2011, 26, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Pace, J.M.; Kuslich, C.D.; Willing, M.C.; Byers, P.H. Disruption of One Intra-Chain Disulphide Bond in the Carboxyl-Terminal Propeptide of the Proalpha1(I) Chain of Type I Procollagen Permits Slow Assembly and Secretion of Overmodified, but Stable Procollagen Trimers and Results in Mild Osteogenesis Imperfecta. J. Med. Genet. 2001, 38, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Fan, D.; Takawale, A.; Lee, J.; Kassiri, Z. Cardiac Fibroblasts, Fibrosis and Extracellular Matrix Remodeling in Heart Disease. Fibrogenesis Tissue Repair. 2012, 5, 15. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2023 Update. Nucleic Acids Res. 2023, 51, D1373–D1380. [Google Scholar] [CrossRef]
- PubChem Proline. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/145742 (accessed on 4 May 2023).
- PubChem Lysine. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/5962 (accessed on 4 May 2023).
- PubChem 4-Hydroxyproline. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/825 (accessed on 4 May 2023).
- PubChem 3-Hydroxyproline. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/150779 (accessed on 4 May 2023).
- PubChem 5-Hydroxylysine. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/3032849 (accessed on 4 May 2023).
- PubChem Cystine. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/67678 (accessed on 5 May 2023).
- Cabral, W.A.; Fratzl-Zelman, N.; Weis, M.; Perosky, J.E.; Alimasa, A.; Harris, R.; Kang, H.; Makareeva, E.; Barnes, A.M.; Roschger, P.; et al. Substitution of Murine Type I Collagen A1 3-Hydroxylation Site Alters Matrix Structure but Does Not Recapitulate Osteogenesis Imperfecta Bone Dysplasia. Matrix Biol. 2020, 90, 20–39. [Google Scholar] [CrossRef]
- Turecek, C.; Fratzl-Zelman, N.; Rumpler, M.; Buchinger, B.; Spitzer, S.; Zoehrer, R.; Durchschlag, E.; Klaushofer, K.; Paschalis, E.P.; Varga, F. Collagen Cross-Linking Influences Osteoblastic Differentiation. Calcif. Tissue Int. 2008, 82, 392–400. [Google Scholar] [CrossRef]
- DiChiara, A.S.; Taylor, R.J.; Wong, M.Y.; Doan, N.D.; Rosario, A.M.; Shoulders, M.D. Mapping and Exploring the Collagen-I Proteostasis Network. ACS Chem. Biol. 2016, 11, 1408–1421. [Google Scholar] [CrossRef]
- Giménez, A.; Duch, P.; Puig, M.; Gabasa, M.; Xaubet, A.; Alcaraz, J. Dysregulated Collagen Homeostasis by Matrix Stiffening and TGF-Β1 in Fibroblasts from Idiopathic Pulmonary Fibrosis Patients: Role of FAK/Akt. Int. J. Mol. Sci. 2017, 18, 2431. [Google Scholar] [CrossRef]
- Tsitoura, E.; Trachalaki, A.; Vasarmidi, E.; Mastrodemou, S.; Margaritopoulos, G.A.; Kokosi, M.; Fanidis, D.; Galaris, A.; Aidinis, V.; Renzoni, E.; et al. Collagen 1a1 Expression by Airway Macrophages Increases In Fibrotic ILDs and Is Associated With FVC Decline and Increased Mortality. Front. Immunol. 2021, 12, 645548. [Google Scholar] [CrossRef]
- Li, H.; Chang, H.M.; Shi, Z.; Leung, P.C.K. The P38 Signaling Pathway Mediates the TGF-Β1-Induced Increase in Type I Collagen Deposition in Human Granulosa Cells. FASEB J. 2020, 34, 15591–15604. [Google Scholar] [CrossRef] [PubMed]
- COL1A1—Collagen Alpha-1(I) Chain—Homo Sapiens (Human)|UniProtKB|UniProt. Available online: https://www.uniprot.org/uniprotkb/P02452/entry (accessed on 9 January 2023).
- Zhang, Z.; Wang, Y.; Zhang, J.; Zhong, J.; Yang, R. COL1A1 Promotes Metastasis in Colorectal Cancer by Regulating the WNT/PCP Pathway. Mol. Med. Rep. 2018, 17, 5037–5042. [Google Scholar] [CrossRef] [PubMed]
- Bornstein, P.; Walsh, V.; Tullis, J.; Stainbrook, E.; Bateman, J.F.; Hormuzdi, S.G. The Globular Domain of the Proalpha 1(I) N-Propeptide Is Not Required for Secretion, Processing by Procollagen N-Proteinase, or Fibrillogenesis of Type I Collagen in Mice. J. Biol. Chem. 2002, 277, 2605–2613. [Google Scholar] [CrossRef]
- Han, S.; Makareeva, E.; Kuznetsova, N.V.; DeRidder, A.M.; Sutter, M.B.; Losert, W.; Phillips, C.L.; Visse, R.; Nagase, H.; Leikin, S. Molecular Mechanism of Type I Collagen Homotrimer Resistance to Mammalian Collagenases. J. Biol. Chem. 2010, 285, 22276–22281. [Google Scholar] [CrossRef]
- Kuznetsova, N.V.; McBride, D.J.; Leikin, S. Changes in Thermal Stability and Microunfolding Pattern of Collagen Helix Resulting from the Loss of A2(I) Chain in Osteogenesis Imperfecta Murine. J. Mol. Biol. 2003, 331, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.-W.; Shefelbine, S.J.; Buehler, M.J. Structural and Mechanical Differences between Collagen Homo- and Heterotrimers: Relevance for the Molecular Origin of Brittle Bone Disease. Biophys. J. 2012, 102, 640–648. [Google Scholar] [CrossRef] [PubMed]
- Esposito, C.; Caputo, I. Mammalian Transglutaminases. FEBS J. 2005, 272, 615–631. [Google Scholar] [CrossRef] [PubMed]
- Avery, N.C.; Bailey, A.J. The Effects of the Maillard Reaction on the Physical Properties and Cell Interactions of Collagen. Pathol. Biol. 2006, 54, 387–395. [Google Scholar] [CrossRef]
- Feres-Filho, E.J.; Choi, Y.J.; Han, X.; Takala, T.E.; Trackman, P.C. Pre- and Post-Translational Regulation of Lysyl Oxidase by Transforming Growth Factor-Beta 1 in Osteoblastic MC3T3-E1 Cells. J. Biol. Chem. 1995, 270, 30797–30803. [Google Scholar] [CrossRef]
- Uzawa, K.; Grzesik, W.J.; Nishiura, T.; Kuznetsov, S.A.; Robey, P.G.; Brenner, D.A.; Yamauchi, M. Differential Expression of Human Lysyl Hydroxylase Genes, Lysine Hydroxylation, and Cross-Linking of Type I Collagen During Osteoblastic Differentiation In Vitro. J. Bone Miner. Res. 1999, 14, 1272–1280. [Google Scholar] [CrossRef] [PubMed]
- Hudson, D.M.; Garibov, M.; Dixon, D.R.; Popowics, T.; Eyre, D.R. Distinct Post-Translational Features of Type I Collagen Are Conserved in Mouse and Human Periodontal Ligament. J. Periodontal Res. 2017, 52, 1042–1049. [Google Scholar] [CrossRef] [PubMed]
- PubChem Allysine. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/160603 (accessed on 24 May 2023).
- PubChem Glutamine. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/5961 (accessed on 24 May 2023).
- PubChem Dehydrolysylnorleucine. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/193137 (accessed on 24 May 2023).
- PubChem Glyoxal-Lysine Dimer. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/46878529 (accessed on 24 May 2023).
- PubChem Epsilon-(Gamma-L-Glutamyl)-L-Lysine. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/7015684 (accessed on 24 May 2023).
- Craig, V.J.; Zhang, L.; Hagood, J.S.; Owen, C.A. Matrix Metalloproteinases as Therapeutic Targets for Idiopathic Pulmonary Fibrosis. Am. J. Respir. Cell. Mol. Biol. 2015, 53, 585–600. [Google Scholar] [CrossRef]
- Proteasix Search Rseults for Human COL1A1. Available online: http://proteasix.cs.man.ac.uk/GUI_substrate/P02452.txt (accessed on 9 January 2023).
- Vijver, S.V.; Singh, A.; Mommers-Elshof, E.T.A.M.; Meeldijk, J.; Copeland, R.; Boon, L.; Langermann, S.; Flies, D.; Meyaard, L.; Ramos, M.I.P. Collagen Fragments Produced in Cancer Mediate T Cell Suppression Through Leukocyte-Associated Immunoglobulin-Like Receptor 1. Front. Immunol. 2021, 12, 733561. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; He, C.; Zhao, H.; Jiang, W.; Xu, S.; Li, J.; Ma, T.; Huang, C. Sennoside A Prevents Liver Fibrosis by Binding DNMT1 and Suppressing DNMT1-mediated PTEN Hypermethylation in HSC Activation and Proliferation. FASEB J. 2020, 34, 14558–14571. [Google Scholar] [CrossRef]
- Yang, J.J.; Yang, Y.; Zhang, C.; Li, J. Epigenetic Silencing of LncRNA ANRIL Enhances Liver Fibrosis and HSC Activation through Activating AMPK Pathway. J. Cell. Mol. Med. 2020, 24, 2677–2687. [Google Scholar] [CrossRef]
- Bibaki, E.; Tsitoura, E.; Vasarmidi, E.; Margaritopoulos, G.; Trachalaki, A.; Koutoulaki, C.; Georgopoulou, T.; Spandidos, D.A.; Tzanakis, N.; Antoniou, K.M. MiR-185 and MiR-29a Are Similarly Expressed in the Bronchoalveolar Lavage Cells in IPF and Lung Cancer but Common Targets DNMT1 and COL1A1 Show Disease Specific Patterns. Mol. Med. Rep. 2018, 17, 7105–7112. [Google Scholar] [CrossRef]
- Zeng, X.; Yue, Z.; Gao, Y.; Jiang, G.; Zeng, F.; Shao, Y.; Huang, J.; Yin, M.; Li, Y. NR4A1 Is Involved in Fibrogenesis in Ovarian Endometriosis. Cell Physiol. Biochem. 2018, 46, 1078–1090. [Google Scholar] [CrossRef]
- Palumbo-Zerr, K.; Zerr, P.; Distler, A.; Fliehr, J.; Mancuso, R.; Huang, J.; Mielenz, D.; Tomcik, M.; Fürnrohr, B.G.; Scholtysek, C.; et al. Orphan Nuclear Receptor NR4A1 Regulates Transforming Growth Factor-β Signaling and Fibrosis. Nat. Med. 2015, 21, 150–158. [Google Scholar] [CrossRef]
- Peng, Y.; Kim, J.M.; Park, H.S.; Yang, A.; Islam, C.; Lakatta, E.G.; Lin, L. AGE-RAGE Signal Generates a Specific NF-ΚB RelA “Barcode” That Directs Collagen I Expression. Sci. Rep. 2016, 6, 18822. [Google Scholar] [CrossRef] [PubMed]
- Margadant, C.; Sonnenberg, A. Integrin–TGF-β Crosstalk in Fibrosis, Cancer and Wound Healing. EMBO Rep. 2010, 11, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Goffin, L.; Seguin-Estévez, Q.; Alvarez, M.; Reith, W.; Chizzolini, C. Transcriptional Regulation of Matrix Metalloproteinase-1 and Collagen 1A2 Explains the Anti-Fibrotic Effect Exerted by Proteasome Inhibition in Human Dermal Fibroblasts. Arthritis Res. Ther. 2010, 12, R73. [Google Scholar] [CrossRef] [PubMed]
- Fineschi, S.; Reith, W.; Guerne, P.A.; Dayer, J.M.; Chizzolini, C. Proteasome Blockade Exerts an Antifibrotic Activity by Coordinately Down-Regulating Type I Collagen and Tissue Inhibitor of Metalloproteinase-1 and up-Regulating Metalloproteinase-1 Production in Human Dermal Fibroblasts. FASEB J. 2006, 20, 562–564. [Google Scholar] [CrossRef]
- Ghiassi-Nejad, Z.; Hernandez-Gea, V.; Woodrell, C.; Lang, U.E.; Dumic, K.; Kwong, A.; Friedman, S.L. Reduced Hepatic Stellate Cell Expression of Kruppel-like Factor 6 Tumor Suppressor Isoforms Amplifies Fibrosis during Acute and Chronic Rodent Liver Injury. Hepatol. Baltim. Md. 2013, 57, 786–796. [Google Scholar] [CrossRef]
- Zhang, L.-X.; Zhang, S.-H.; Wang, C.-Q.; Bing, Q.; Zhao, Z.; Wang, J.; Zhang, L. Role and Mechanism of MicroRNA-548c-3p/c-Myb in Myocardial Infarction Fibrosis in Rats. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 4908–4916. [Google Scholar] [CrossRef]
- Yao, Q.; Xing, Y.; Wang, Z.; Liang, J.; Lin, Q.; Huang, M.; Chen, Y.; Lin, B.; Xu, X.; Chen, W. MiR-16-5p Suppresses Myofibroblast Activation in Systemic Sclerosis by Inhibiting NOTCH Signaling. Aging 2020, 13, 2640–2654. [Google Scholar] [CrossRef]
- Chen, B. The MiRNA-184 Drives Renal Fibrosis by Targeting HIF1AN in Vitro and in Vivo. Int. Urol. Nephrol. 2019, 51, 543–550. [Google Scholar] [CrossRef]
- Mooring, M.; Fowl, B.H.; Lum, S.Z.C.; Liu, Y.; Yao, K.; Softic, S.; Kirchner, R.; Bernstein, A.; Singhi, A.D.; Jay, D.G.; et al. Hepatocyte Stress Increases Expression of Yes-Associated Protein and Transcriptional Coactivator With PDZ-Binding Motif in Hepatocytes to Promote Parenchymal Inflammation and Fibrosis. Hepatology 2020, 71, 1813–1830. [Google Scholar] [CrossRef]
- Yuan, X.; Pan, J.; Wen, L.; Gong, B.; Li, J.; Gao, H.; Tan, W.; Liang, S.; Zhang, H.; Wang, X. MiR-590-3p Regulates Proliferation, Migration and Collagen Synthesis of Cardiac Fibroblast by Targeting ZEB1. J. Cell. Mol. Med. 2020, 24, 227–237. [Google Scholar] [CrossRef]
- Trelford, C.B.; Dagnino, L.; Di Guglielmo, G.M. Transforming Growth Factor-β in Tumour Development. Front. Mol. Biosci. 2022, 9, 991612. [Google Scholar] [CrossRef] [PubMed]
- Varricchio, L.; Iancu-Rubin, C.; Upadhyaya, B.; Zingariello, M.; Martelli, F.; Verachi, P.; Clementelli, C.; Denis, J.F.; Rahman, A.H.; Tremblay, G.; et al. TGF-Β1 Protein Trap AVID200 Beneficially Affects Hematopoiesis and Bone Marrow Fibrosis in Myelofibrosis. JCI Insight 2021, 6, e145651. [Google Scholar] [CrossRef] [PubMed]
- Tam, A.; Leclair, P.; Li, L.V.; Yang, C.X.; Li, X.; Witzigmann, D.; Kulkarni, J.A.; Hackett, T.L.; Dorscheid, D.R.; Singhera, G.K.; et al. FAM13A as Potential Therapeutic Target in Modulating TGF-β-Induced Airway Tissue Remodeling in COPD. Am. J. Physiol. Lung Cell Mol. Physiol. 2021, 321, L377–L391. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Liu, Y.; Kahn, M.; Ann, D.K.; Han, A.; Wang, H.; Nguyen, C.; Flodby, P.; Zhong, Q.; Krishnaveni, M.S.; et al. Interactions between β-Catenin and Transforming Growth Factor-β Signaling Pathways Mediate Epithelial-Mesenchymal Transition and Are Dependent on the Transcriptional Co-Activator CAMP-Response Element-Binding Protein (CREB)-Binding Protein (CBP). J. Biol. Chem. 2012, 287, 7026–7038. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhang, J.; Wei, D.; Chen, J.; Yang, J. GCN2 Regulates ATF3-P38 MAPK Signaling Transduction in Pulmonary Veno-Occlusive Disease. J. Cardiovasc. Pharmacol. Ther. 2021, 26, 677–689. [Google Scholar] [CrossRef]
- Nieto, N. Oxidative-Stress and IL-6 Mediate the Fibrogenic Effects of Rodent Kupffer Cells on Stellate Cells. Hepatology 2006, 44, 1487–1501. [Google Scholar] [CrossRef]
- He, J.; Peng, H.; Wang, M.; Liu, Y.; Guo, X.; Wang, B.; Dai, L.; Cheng, X.; Meng, Z.; Yuan, L.; et al. Isoliquiritigenin Inhibits TGF-Β1-Induced Fibrogenesis through Activating Autophagy via PI3K/AKT/MTOR Pathway in MRC-5 Cells. Acta Biochim. Biophys. Sin. Shanghai 2020, 52, 810–820. [Google Scholar] [CrossRef]
- Junior; Lai, Y.S.; Nguyen, H.T.; Salmanida, F.P.; Chang, K.T. MERTK +/Hi M2c Macrophages Induced by Baicalin Alleviate Non-Alcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2021, 22, 10604. [Google Scholar] [CrossRef]
- Zhang, L.; Yin, H.; Jiao, L.; Liu, T.; Gao, Y.; Shao, Y.; Zhang, Y.; Shan, H.; Yang, B. Abnormal Downregulation of Caveolin-3 Mediates the Pro-Fibrotic Action of MicroRNA-22 in a Model of Myocardial Infarction. Cell Physiol. Biochem. 2018, 45, 1641–1653. [Google Scholar] [CrossRef]
- Hutter, S.; van Haaften, W.T.; Hünerwadel, A.; Baebler, K.; Herfarth, N.; Raselli, T.; Mamie, C.; Misselwitz, B.; Rogler, G.; Weder, B.; et al. Intestinal Activation of PH-Sensing Receptor OGR1 [GPR68] Contributes to Fibrogenesis. J. Crohns Colitis 2018, 12, 1348–1358. [Google Scholar] [CrossRef]
- Rovati, L.; Kaneko, N.; Pedica, F.; Monno, A.; Maehara, T.; Perugino, C.; Lanzillotta, M.; Pecetta, S.; Stone, J.H.; Doglioni, C.; et al. Mer Tyrosine Kinase as a Possible Link between Resolution of Inflammation and Tissue Fibrosis in IgG4-Related Disease. Rheumatol. Oxf. Engl. 2021, 60, 4929–4941. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, K.; Koyanagi, T.; Berry, N.C.; Sun, L.; Mochly-Rosen, D. Pharmacological Inhibition of Epsilon-Protein Kinase C Attenuates Cardiac Fibrosis and Dysfunction in Hypertension-Induced Heart Failure. Hypertens. Dallas Tex. 1979 2008, 51, 1565–1569. [Google Scholar] [CrossRef]
- Nagel, D.J.; Rackow, A.R.; Ku, W.-Y.; Bell, T.J.; Sime, P.J.; Kottmann, R.M. Cell-Type-Specific Effects of the Ovarian Cancer G-Protein Coupled Receptor (OGR1) on Inflammation and Fibrosis; Potential Implications for Idiopathic Pulmonary Fibrosis. Cells 2022, 11, 2540. [Google Scholar] [CrossRef]
- Herum, K.M.; Romaine, A.; Wang, A.; Melleby, A.O.; Strand, M.E.; Pacheco, J.; Braathen, B.; Dunér, P.; Tønnessen, T.; Lunde, I.G.; et al. Syndecan-4 Protects the Heart From the Profibrotic Effects of Thrombin-Cleaved Osteopontin. J. Am. Heart Assoc. Cardiovasc. Cerebrovasc. Dis. 2020, 9, e013518. [Google Scholar] [CrossRef]
- Shenoy, C.C.; Khan, Z.; Zheng, Y.; Jones, T.L.; Khazaie, K.; Daftary, G.S. Progressive Fibrosis: A Progesterone- and KLF11-Mediated Sexually Dimorphic Female Response. Endocrinology 2017, 158, 3605–3619. [Google Scholar] [CrossRef] [PubMed]
- Okamura, T.; Hamaguchi, M.; Bamba, R.; Nakajima, H.; Yoshimura, Y.; Kimura, T.; Nishida, K.; Hashimoto, Y.; Fukuda, T.; Senmaru, T.; et al. Immune Modulating Effects of Additional Supplementation of Estradiol Combined with Testosterone in Murine Testosterone-Deficient NAFLD Model. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 318, G989–G999. [Google Scholar] [CrossRef] [PubMed]
- Duval, E.; Bouyoucef, M.; Leclercq, S.; Baugé, C.; Boumédiene, K. Hypoxia Inducible Factor 1 Alpha Down-Regulates Type i Collagen through Sp3 Transcription Factor in Human Chondrocytes. IUBMB Life 2016, 68, 756–763. [Google Scholar] [CrossRef]
- Zhang, Y.; Song, C.; Zhang, Y.; Wang, Y.; Feng, C.; Chen, J.; Wei, L.; Pan, Q.; Shang, D.; Zhu, Y.; et al. TcoFBase: A Comprehensive Database for Decoding the Regulatory Transcription Co-Factors in Human and Mouse. Nucleic Acids Res. 2022, 50, D391–D401. [Google Scholar] [CrossRef]
- Ding, H.; Xu, Y.; Jiang, N. Upregulation of MiR-101a Suppresses Chronic Renal Fibrosis by Regulating KDM3A via Blockade of the YAP-TGF-β-Smad Signaling Pathway. Mol. Ther. Nucleic Acids 2020, 19, 1276–1289. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, W.; Zhang, H.-M.; Xie, G.-Y.; Miao, Y.-R.; Xia, M.; Guo, A.-Y. HTFtarget: A Comprehensive Database for Regulations of Human Transcription Factors and Their Targets. Genom. Proteom. Bioinform. 2020, 18, 120–128. [Google Scholar] [CrossRef]
- Liska, O.; Bohár, B.; Hidas, A.; Korcsmáros, T.; Papp, B.; Fazekas, D.; Ari, E. TFLink: An Integrated Gateway to Access Transcription Factor–Target Gene Interactions for Multiple Species. Database 2022, 2022, baac083. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yang, Y.; Yu, H.; Wu, W.; Sun, Y.; Pan, Y.; Kong, L. Polydatin Inhibits ZEB1-Invoked Epithelial-Mesenchymal Transition in Fructose-Induced Liver Fibrosis. J. Cell Mol. Med. 2020, 24, 13208–13222. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, S.A.; Saitta, B. Alterations in the Regulation of Expression of the Al(I) Collagen Gene (COL1A1) in Systemic Sclerosis (Scleroderma). Springer Semin. Immunopathol. 2000, 21, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Beauchef, G.; Bigot, N.; Kypriotou, M.; Renard, E.; Porée, B.; Widom, R.; Dompmartin-Blanchere, A.; Oddos, T.; Maquart, F.-X.; Demoor, M.; et al. The P65 Subunit of NF-ΚB Inhibits COL1A1 Gene Transcription in Human Dermal and Scleroderma Fibroblasts through Its Recruitment on Promoter by Protein Interaction with Transcriptional Activators (c-Krox, Sp1, and Sp3). J. Biol. Chem. 2012, 287, 3462–3478. [Google Scholar] [CrossRef]
- Lu, S.-L.; Huang, C.-F.; Li, C.-L.; Lu, H.-K.; Chen, L.-S. Role of IL-6 and STAT3 Signaling in Dihydropyridine-Induced Gingival Overgrowth Fibroblasts. Oral. Dis. 2021, 27, 1796–1805. [Google Scholar] [CrossRef]
- Choi, D.; Kang, W.; Park, S.; Son, B.; Park, T. β-Ionone Attenuates Dexamethasone-Induced Suppression of Collagen and Hyaluronic Acid Synthesis in Human Dermal Fibroblasts. Biomolecules 2021, 11, 619. [Google Scholar] [CrossRef]
- Gewartowska, O.; Aranaz-Novaliches, G.; Krawczyk, P.S.; Mroczek, S.; Kusio-Kobiałka, M.; Tarkowski, B.; Spoutil, F.; Benada, O.; Kofroňová, O.; Szwedziak, P.; et al. Cytoplasmic Polyadenylation by TENT5A Is Required for Proper Bone Formation. Cell Rep. 2021, 35, 109015. [Google Scholar] [CrossRef]
- McDaniel, K.; Huang, L.; Sato, K.; Wu, N.; Annable, T.; Zhou, T.; Ramos-Lorenzo, S.; Wan, Y.; Huang, Q.; Francis, H.; et al. The Let-7/Lin28 Axis Regulates Activation of Hepatic Stellate Cells in Alcoholic Liver Injury. J. Biol. Chem. 2017, 292, 11336–11347. [Google Scholar] [CrossRef]
- Bi, S.; Chai, L.; Yuan, X.; Cao, C.; Li, S. MicroRNA-98 Inhibits the Cell Proliferation of Human Hypertrophic Scar Fibroblasts via Targeting Col1A1. Biol. Res. 2017, 50, 22. [Google Scholar] [CrossRef]
- Wang, Y.; Mei, X.; Song, W.; Wang, C.; Qiu, X. LncRNA LINC00511 Promotes COL1A1-Mediated Proliferation and Metastasis by Sponging MiR-126-5p/MiR-218-5p in Lung Adenocarcinoma. BMC Pulm. Med. 2022, 22, 272. [Google Scholar] [CrossRef]
- Liu, T.; Ye, P.; Ye, Y.; Lu, S.; Han, B. Circular RNA Hsa_circRNA_002178 Silencing Retards Breast Cancer Progression via MicroRNA-328-3p-mediated Inhibition of COL1A1. J. Cell Mol. Med. 2020, 24, 2189–2201. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lu, J.; Xie, H.; Wang, D.; Ni, H.; Zhu, Y.; Ren, L.; Meng, X.; Wang, R. CircHIPK3 Regulates Lung Fibroblast-to-Myofibroblast Transition by Functioning as a Competing Endogenous RNA. Cell Death Dis. 2019, 10, 182. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, S.; Xu, L.; Feng, Y.; Wu, X.; Zhang, M.; Yu, Z.; Zhou, X. Involvement of CircHIPK3 in the Pathogenesis of Diabetic Cardiomyopathy in Mice. Diabetologia 2021, 64, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jin, B.J.; Chen, Q.; Yan, B.J.; Liu, Z.L. MicroRNA-29b Upregulation Improves Myocardial Fibrosis and Cardiac Function in Myocardial Infarction Rats through Targeting SH2B3. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 10115–10122. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Itami, S.; Kuroda, M.; Yoshizato, K.; Kawada, N.; Murakami, Y. MiR-29a Assists in Preventing the Activation of Human Stellate Cells and Promotes Recovery From Liver Fibrosis in Mice. Mol. Ther. 2016, 24, 1848–1859. [Google Scholar] [CrossRef]
- Yang, T.; Liang, Y.; Lin, Q.; Liu, J.; Luo, F.; Li, X.; Zhou, H.; Zhuang, S.; Zhang, H. MiR-29 Mediates TGFβ1-Induced Extracellular Matrix Synthesis through Activation of PI3K-AKT Pathway in Human Lung Fibroblasts. J. Cell Biochem. 2013, 114, 1336–1342. [Google Scholar] [CrossRef]
- Guo, L.; Xu, K.; Yan, H.; Feng, H.; Wang, T.; Chai, L.; Xu, G. MicroRNA Expression Signature and the Therapeutic Effect of the MicroRNA-21 Antagomir in Hypertrophic Scarring. Mol. Med. Rep. 2017, 15, 1211–1221. [Google Scholar] [CrossRef]
- Liang, H.; Wang, X.; Si, C.; Duan, Y.; Chen, B.; Liang, H.; Yang, D. Downregulation of MiR-141 Deactivates Hepatic Stellate Cells by Targeting the PTEN/AKT/MTOR Pathway. Int. J. Mol. Med. 2020, 46, 406–414. [Google Scholar] [CrossRef]
- Ning, J.; Zhang, H.; Yang, H. MicroRNA-326 Inhibits Endometrial Fibrosis by Regulating TGF-β1/Smad3 Pathway in Intrauterine Adhesions. Mol. Med. Rep. 2018, 18, 2286–2292. [Google Scholar] [CrossRef]
- Brea, R.; Motiño, O.; Francés, D.; García-Monzón, C.; Vargas, J.; Fernández-Velasco, M.; Boscá, L.; Casado, M.; Martín-Sanz, P.; Agra, N. PGE2 Induces Apoptosis of Hepatic Stellate Cells and Attenuates Liver Fibrosis in Mice by Downregulating MiR-23a-5p and MiR-28a-5p. Biochim. Biophys. Acta BBA-Mol. Basis Dis. 2018, 1864, 325–337. [Google Scholar] [CrossRef]
- Chiasson, V.; Takano, A.P.C.; Guleria, R.S.; Gupta, S. Deficiency of MicroRNA MiR-1954 Promotes Cardiac Remodeling and Fibrosis. J. Am. Heart Assoc. 2019, 8, e012880. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Su, W.; Zhao, X.; Shan, T.; Jin, T.; Guo, Y.; Li, C.; Li, R.; Zhou, Y.; Shan, H.; et al. LncRNA PFAR Contributes to Fibrogenesis in Lung Fibroblasts through Competitively Binding to MiR-15a. Biosci. Rep. 2019, 39, BSR20190280. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-J.; Jeng, J.-H.; Chang, H.-H.; Huang, M.-Y.; Tsai, F.-F.; Jane Yao, C.-C. Differential Regulation of Collagen, Lysyl Oxidase and MMP-2 in Human Periodontal Ligament Cells by Low- and High-Level Mechanical Stretching. J. Periodontal Res. 2013, 48, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Forlino, A.; Cabral, W.A.; Barnes, A.M.; Marini, J.C. New Perspectives on Osteogenesis Imperfecta. Nat. Rev. Endocrinol. 2011, 7, 540–557. [Google Scholar] [CrossRef] [PubMed]
- Perla-Kajan, J.; Utyro, O.; Rusek, M.; Malinowska, A.; Sitkiewicz, E.; Jakubowski, H. N-Homocysteinylation Impairs Collagen Cross-Linking in Cystathionine β-Synthase-Deficient Mice: A Novel Mechanism of Connective Tissue Abnormalities. FASEB J. 2016, 30, 3810–3821. [Google Scholar] [CrossRef]
- Thompson, K.J.; Lakner, A.M.; Cross, B.W.; Tsukada, S.; Rippe, R.A.; McKillop, I.H.; Schrum, L.W. S-Adenosyl-L-Methionine Inhibits Collagen Secretion in Hepatic Stellate Cells via Increased Ubiquitination. Liver Int. 2011, 31, 891–901. [Google Scholar] [CrossRef]
- Lindert, U.; Gnoli, M.; Maioli, M.; Bedeschi, M.F.; Sangiorgi, L.; Rohrbach, M.; Giunta, C. Insight into the Pathology of a COL1A1 Signal Peptide Heterozygous Mutation Leading to Severe Osteogenesis Imperfecta. Calcif. Tissue Int. 2018, 102, 373–379. [Google Scholar] [CrossRef]
- Tian, C.; Huang, Y.; Clauser, K.R.; Rickelt, S.; Lau, A.N.; Carr, S.A.; Vander Heiden, M.G.; Hynes, R.O. Suppression of Pancreatic Ductal Adenocarcinoma Growth and Metastasis by Fibrillar Collagens Produced Selectively by Tumor Cells. Nat. Commun. 2021, 12, 2328. [Google Scholar] [CrossRef]
- Habgood, A.N.; Tatler, A.L.; Porte, J.; Wahl, S.M.; Laurent, G.J.; John, A.E.; Johnson, S.R.; Jenkins, G. Secretory Leukocyte Protease Inhibitor Gene Deletion Alters Bleomycin-Induced Lung Injury, but Not Development of Pulmonary Fibrosis. Lab. Investig. J. Technol. Methods Pathol. 2016, 96, 623–631. [Google Scholar] [CrossRef]
- Roman, K.; Murphy, S.F.; Done, J.D.; McKenna, K.E.; Schaeffer, A.J.; Thumbikat, P. Role of Protease-Activated Receptor 2 in the Development of Lower Urinary Tract Dysfunction. J. Urol. 2016, 196, 588–598. [Google Scholar] [CrossRef]
- Decaris, M.L.; Schaub, J.R.; Chen, C.; Cha, J.; Lee, G.G.; Rexhepaj, M.; Ho, S.S.; Rao, V.; Marlow, M.M.; Kotak, P.; et al. Dual Inhibition of Avβ6 and Avβ1 Reduces Fibrogenesis in Lung Tissue Explants from Patients with IPF. Respir. Res. 2021, 22, 265. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Kobayakawa, K.; Ohkawa, Y.; Kumamaru, H.; Yokota, K.; Saito, T.; Kijima, K.; Yoshizaki, S.; Harimaya, K.; Nakashima, Y.; et al. Interaction of Reactive Astrocytes with Type I Collagen Induces Astrocytic Scar Formation through the Integrin-N-Cadherin Pathway after Spinal Cord Injury. Nat. Med. 2017, 23, 818–828. [Google Scholar] [CrossRef] [PubMed]
- Angel, P.M.; Spruill, L.; Jefferson, M.; Bethard, J.R.; Ball, L.E.; Hughes-Halbert, C.; Drake, R.R. Zonal Regulation of Collagen-Type Proteins and Posttranslational Modifications in Prostatic Benign and Cancer Tissues by Imaging Mass Spectrometry. Prostate 2020, 80, 1071–1086. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, J.; Wang, C.; Xia, L.; Xu, J.; Xie, X.; Lu, W. Microenvironment Remodeled by Tumor and Stromal Cells Elevates Fibroblast-Derived COL1A1 and Facilitates Ovarian Cancer Metastasis. Exp. Cell Res. 2020, 394, 112153. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.K.; Nakamura, M.; Wienke, D.; Isacke, C.M.; Pozzi, A.; Liang, P. Endo180 Binds to the C-Terminal Region of Type I Collagen. J. Biol. Chem. 2005, 280, 22596–22605. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Yang, F.; Fu, R.; Trinh, B.; Sun, N.; Liu, J.; Kumar, A.; Baglieri, J.; Siruno, J.; Le, M.; et al. Collagenolysis-Dependent DDR1 Signalling Dictates Pancreatic Cancer Outcome. Nature 2022, 610, 366–372. [Google Scholar] [CrossRef]
- Horn, L.A.; Chariou, P.L.; Gameiro, S.R.; Qin, H.; Iida, M.; Fousek, K.; Meyer, T.J.; Cam, M.; Flies, D.; Langermann, S.; et al. Remodeling the Tumor Microenvironment via Blockade of LAIR-1 and TGF-β Signaling Enables PD-L1–Mediated Tumor Eradication. J. Clin. Invest. 2022, 132, e155148. [Google Scholar] [CrossRef]
- Kwon, Y.W.; Heo, S.C.; Lee, T.W.; Park, G.T.; Yoon, J.W.; Jang, I.H.; Kim, S.-C.; Ko, H.-C.; Ryu, Y.; Kang, H.; et al. N-Acetylated Proline-Glycine-Proline Accelerates Cutaneous Wound Healing and Neovascularization by Human Endothelial Progenitor Cells. Sci. Rep. 2017, 7, 43057. [Google Scholar] [CrossRef]
- Weathington, N.M.; van Houwelingen, A.H.; Noerager, B.D.; Jackson, P.L.; Kraneveld, A.D.; Galin, F.S.; Folkerts, G.; Nijkamp, F.P.; Blalock, J.E. A Novel Peptide CXCR Ligand Derived from Extracellular Matrix Degradation during Airway Inflammation. Nat. Med. 2006, 12, 317–323. [Google Scholar] [CrossRef]
- Feng, C.; Zhang, Y.; Yang, M.; Huang, B.; Zhou, Y. Collagen-Derived N-Acetylated Proline-Glycine-Proline in Intervertebral Discs Modulates CXCR1/2 Expression and Activation in Cartilage Endplate Stem Cells to Induce Migration and Differentiation Toward a Pro-Inflammatory Phenotype. Stem Cells Dayt. Ohio 2015, 33, 3558–3568. [Google Scholar] [CrossRef]
- Di Lullo, G.A.; Sweeney, S.M.; Korkko, J.; Ala-Kokko, L.; San Antonio, J.D. Mapping the Ligand-Binding Sites and Disease-Associated Mutations on the Most Abundant Protein in the Human, Type I Collagen. J. Biol. Chem. 2002, 277, 4223–4231. [Google Scholar] [CrossRef] [PubMed]
- Iwahashi, H.; Kawashima, Y.; Masaki, H. Decreased Levels of Endocytic Collagen Receptor Endo180 in Dermal Fibroblasts Lead to Decreased Production of Type I Collagen and Increased Expression of Matrix Metalloproteinase-1. Photodermatol. Photoimmunol. Photomed. 2022, 38, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Yeung, D.A.; Shanker, N.; Sohail, A.; Weiss, B.A.; Wang, C.; Wellmerling, J.; Das, S.; Ganju, R.K.; Miller, J.L.C.; Herr, A.B.; et al. Clustering, Spatial Distribution, and Phosphorylation of Discoidin Domain Receptors 1 and 2 in Response to Soluble Collagen I. J. Mol. Biol. 2019, 431, 368–390. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, G.; Mihai, C.; Iscru, D.F. Interaction of Discoidin Domain Receptor 1 with Collagen Type 1. J. Mol. Biol. 2007, 367, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Narayanan, S.; Peruzzi, G.; Apara, A.; Natarajan, K.; Margulies, D.H.; Coligan, J.E.; Borrego, F. A Single Residue, Arginine 65, Is Critical for the Functional Interaction of Leukocyte-Associated Inhibitory Receptor-1 with Collagens. J. Immunol. Baltim. Md. 1950 2009, 182, 5446–5452. [Google Scholar] [CrossRef]
- Ren, J.; Da, J.; Hu, N. Identification of COL1A1 Associated with Immune Infiltration in Brain Lower Grade Glioma. PLoS ONE 2022, 17, e0269533. [Google Scholar] [CrossRef]
- Shea, M.P.; O’Leary, K.A.; Wegner, K.A.; Vezina, C.M.; Schuler, L.A. High Collagen Density Augments MTOR-Dependent Cancer Stem Cells in ERα+ Mammary Carcinomas, and Increases MTOR-Independent Lung Metastases. Cancer Lett. 2018, 433, 1–9. [Google Scholar] [CrossRef]
- Liu, J.; Shen, J.X.; Wu, H.T.; Li, X.L.; Wen, X.F.; Du, C.W.; Zhang, G.J. Collagen 1A1 (COL1A1) Promotes Metastasis of Breast Cancer and Is a Potential Therapeutic Target. Discov. Med. 2018, 25, 211–223. [Google Scholar]
- Huang, C.; Yang, X.; Han, L.; Fan, Z.; Liu, B.; Zhang, C.; Lu, T. The Prognostic Potential of Alpha-1 Type I Collagen Expression in Papillary Thyroid Cancer. Biochem. Biophys. Res. Commun. 2019, 515, 125–132. [Google Scholar] [CrossRef]
- Li, Y.; Sun, R.; Zhao, X.; Sun, B. RUNX2 Promotes Malignant Progression in Gastric Cancer by Regulating COL1A1. Cancer Biomark. 2021, 31, 227–238. [Google Scholar] [CrossRef]
- Zhao, X.; Kwan, J.Y.Y.; Yip, K.; Liu, P.P.; Liu, F.-F. Targeting Metabolic Dysregulation for Fibrosis Therapy. Nat. Rev. Drug. Discov. 2020, 19, 57–75. [Google Scholar] [CrossRef] [PubMed]
- Lee, B. Cross-Linked Collagen Peptides as a Urinary Biomarker of OI Pathobiology; Clinicaltrials.gov: Houston, TX, USA, 2023. [Google Scholar]
- Search of: Col1A1—List Results—ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/results?cond=&term=col1A1&cntry=&state=&city=&dist=&Search=Search (accessed on 8 April 2023).




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Devos, H.; Zoidakis, J.; Roubelakis, M.G.; Latosinska, A.; Vlahou, A. Reviewing the Regulators of COL1A1. Int. J. Mol. Sci. 2023, 24, 10004. https://doi.org/10.3390/ijms241210004
Devos H, Zoidakis J, Roubelakis MG, Latosinska A, Vlahou A. Reviewing the Regulators of COL1A1. International Journal of Molecular Sciences. 2023; 24(12):10004. https://doi.org/10.3390/ijms241210004
Chicago/Turabian StyleDevos, Hanne, Jerome Zoidakis, Maria G. Roubelakis, Agnieszka Latosinska, and Antonia Vlahou. 2023. "Reviewing the Regulators of COL1A1" International Journal of Molecular Sciences 24, no. 12: 10004. https://doi.org/10.3390/ijms241210004
APA StyleDevos, H., Zoidakis, J., Roubelakis, M. G., Latosinska, A., & Vlahou, A. (2023). Reviewing the Regulators of COL1A1. International Journal of Molecular Sciences, 24(12), 10004. https://doi.org/10.3390/ijms241210004