
Effects of Zinc on the Right Cardiovascular Circuit in Long-Term Hypobaric Hypoxia in Wistar Rats
 ,
,  ,
,  ,
, 
Abstract
1. Introduction
2. Results
2.1. Body Weight and Hemoglobin
2.2. Plasma and Lung Zinc Levels
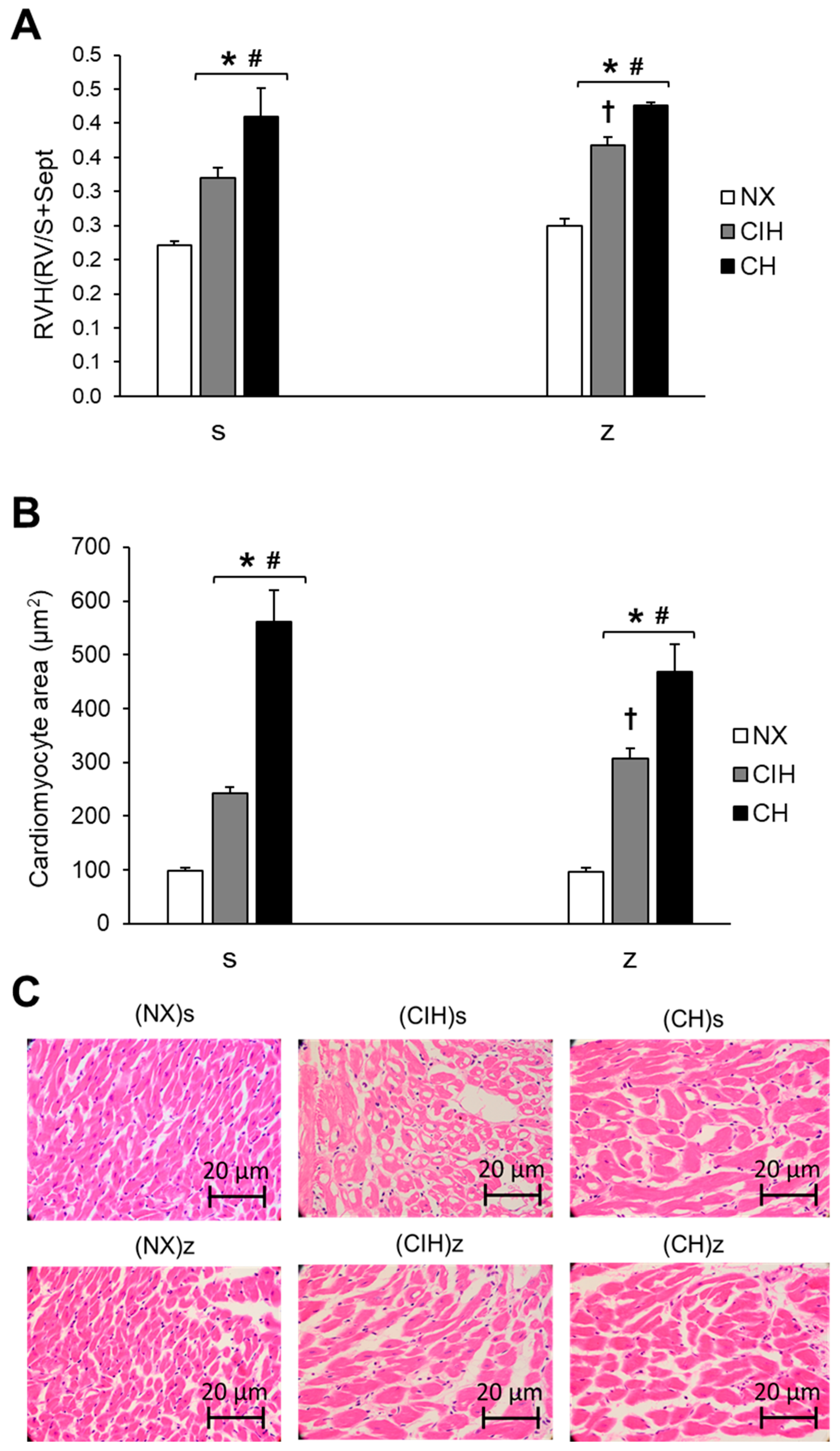
2.3. Right Ventricular Hypertrophy
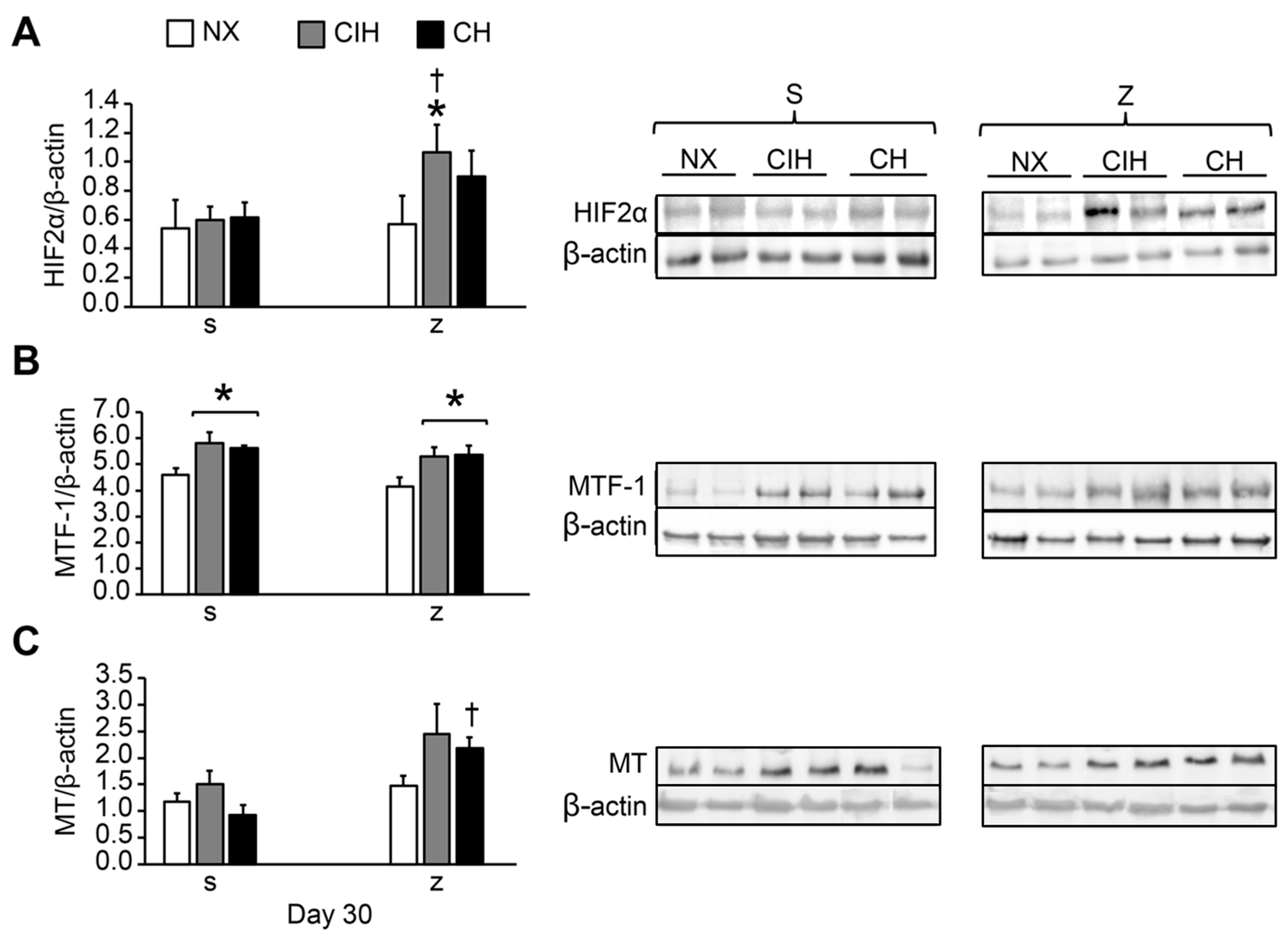
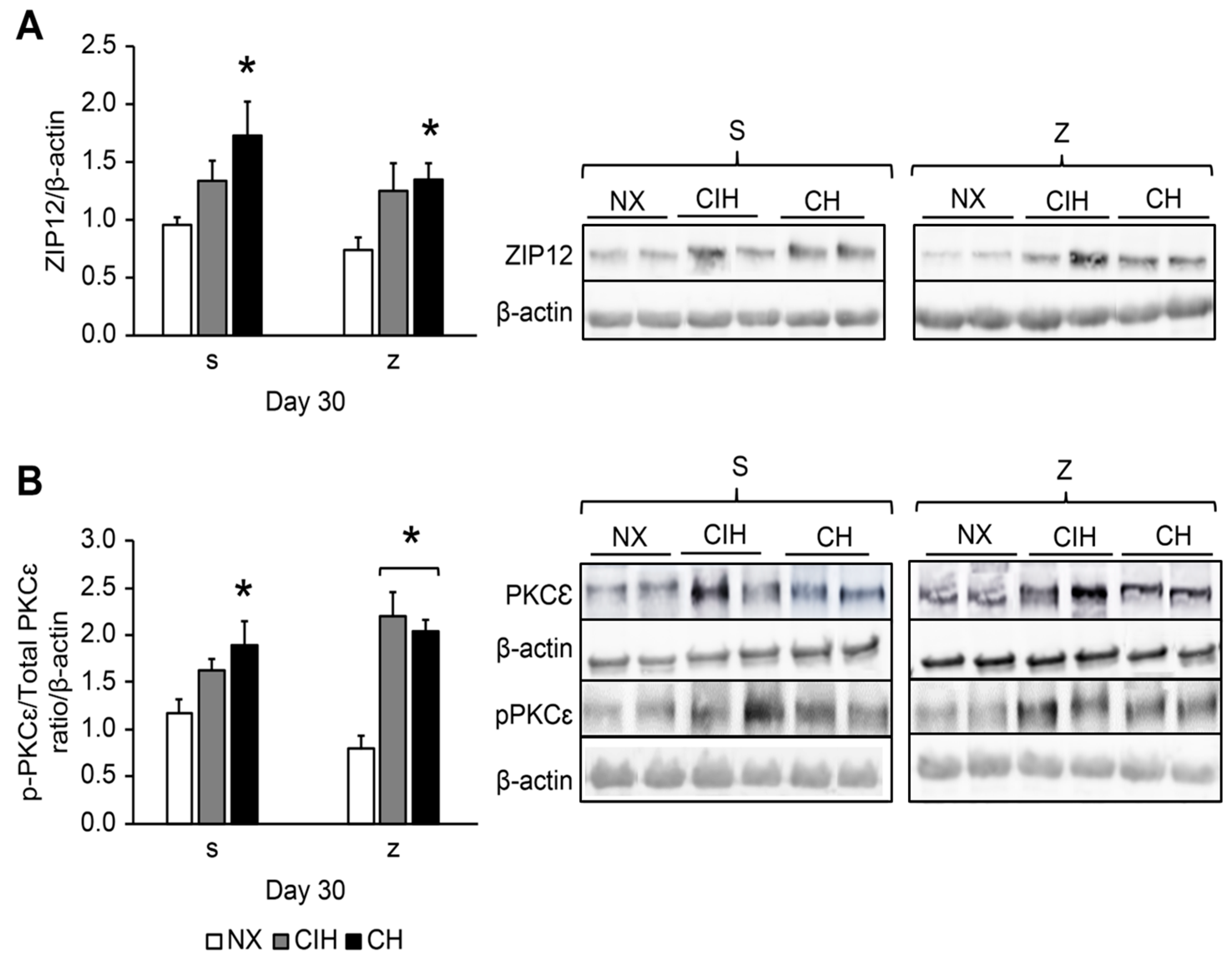
2.4. HIF2α, MTF-1, MT, ZIP12, and PKCε Protein Expression Levels
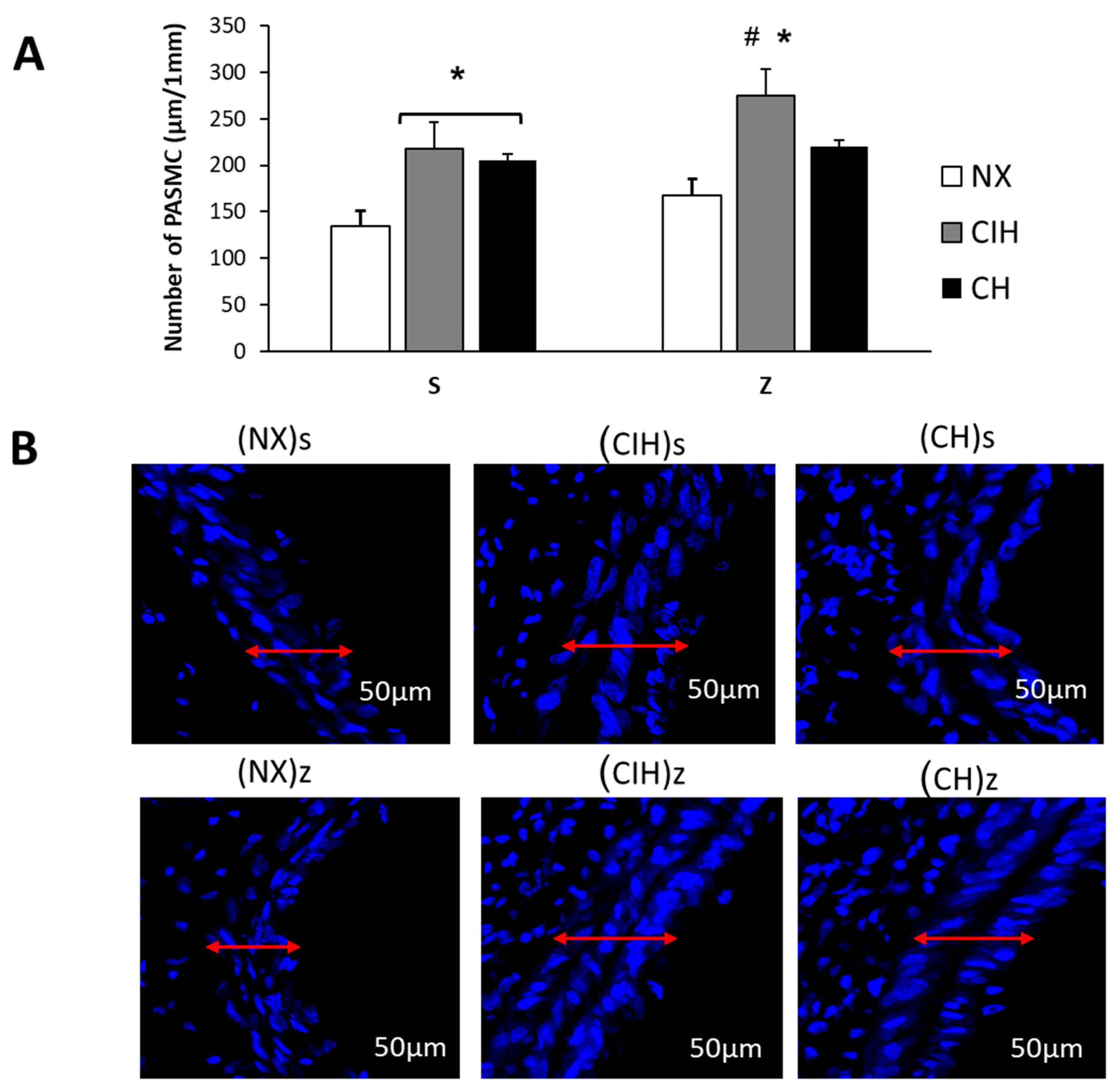
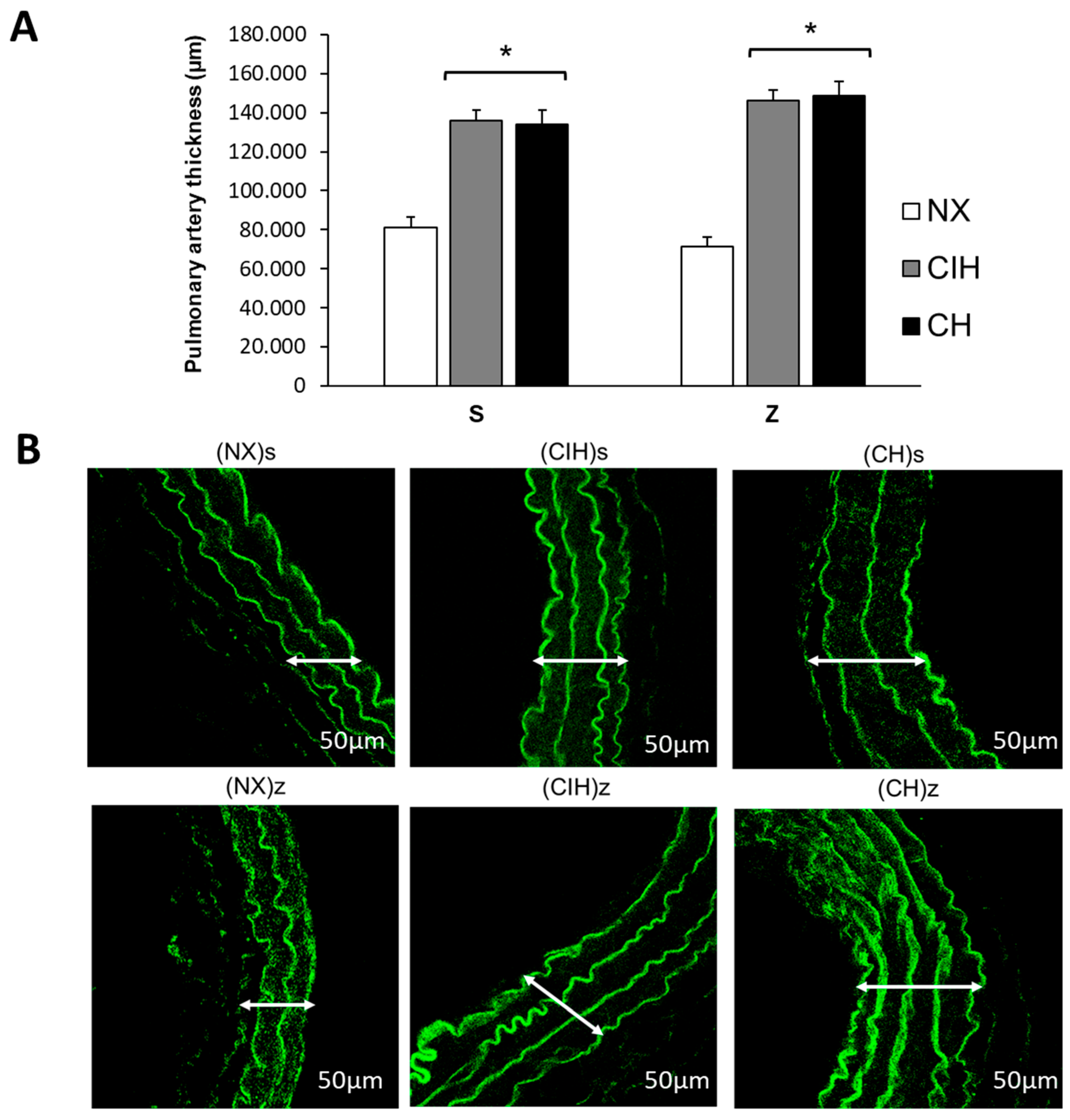
2.5. Pulmonary Artery Remodeling
2.6. Lipid Peroxidation
3. Discussion
Future Perspectives
4. Materials and Methods
4.1. Animal Model
4.2. Body Weight and Hemoglobin
4.3. Zinc Analysis
4.4. Right Ventricular Hypertrophy and Cardiac Histology
4.5. Western Blot Analysis
4.6. Confocal Microscopy
4.7. Lung Lipid Peroxidation
4.8. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AHH | acute hypobaric hypoxia |
| CH | chronic hypobaric hypoxia |
| CIH | chronic intermittent hypobaric hypoxia |
| HAPH | high-altitude pulmonary hypertension |
| Hb | hemoglobin |
| HIF | hypoxia-inducible factor |
| HIF1 | hypoxia-inducible factor 1 alpha |
| HIF2 | hypoxia-inducible factor 2 alpha |
| HRE | hypoxia regulatory element |
| HPV | hypoxic pulmonary vasoconstriction |
| MDA | malondialdehyde |
| MT | metallothionein |
| MTF-1 | metal-responsive transcription factor |
| PASMCs | pulmonary arterial smooth muscle cells |
| PKCε | protein kinase C epsilon |
| RVH | right ventricular hypertrophy |
| ROS | reactive oxygen species |
| Zn | zinc |
| ZIP12 | zinc-regulated 12, iron-regulated transporter-like protein |
References
- Shaw, D.M.; Cabre, G.; Gant, N. Hypoxic hypoxia and brain function in military aviation: Basic physiology and applied perspectives. Front. Physiol. 2021, 12, 665821. [Google Scholar] [CrossRef]
- Penaloza, D.; Arias-Stella, J. The heart and pulmonary circulation at high altitudes: Healthy highlanders and chronic mountain sickness. Circulation 2007, 115, 1132–1146. [Google Scholar] [CrossRef]
- Farias, J.G.; Jimenez, D.; Osorio, J.; Zepeda, A.B.; Figueroa, C.A.; Pulgar, V.M. Acclimatization to chronic intermittent hypoxia in mine workers: A challenge to mountain medicine in Chile. Biol. Res. 2013, 46, 59–67. [Google Scholar] [CrossRef]
- Brito, J.; Siques, P.; Pena, E. Long-term chronic intermittent hypoxia: A particular form of chronic high-altitude pulmonary hypertension. Pulm. Circ. 2020, 10, 5–12. [Google Scholar] [CrossRef]
- Tremblay, J.C.; Ainslie, P.N. Global and country-level estimates of human population at high altitude. Proc. Natl. Acad. Sci. USA 2021, 118, e2102463118. [Google Scholar] [CrossRef]
- Richalet, J.P.; Donoso, M.V.; Jiménez, D.; Antezana, A.M.; Hudson, C.; Cortès, G.; Osorio, J.; Leòn, A. Chilean miners commuting from sea level to 4500 m: A prospective study. High Alt. Med. Biol. 2002, 3, 159–166. [Google Scholar] [CrossRef]
- Beall, C.M. Two routes to functional adaptation: Tibetan and Andean high-altitude natives. Proc. Natl. Acad. Sci. USA 2007, 104 (Suppl. S1), 8655–8660. [Google Scholar] [CrossRef]
- Von Euler, U.S.; Liljestrand, G. Observations on the pulmonary arterial blood pressure in the cat. Acta Physiol. Scand. 1946, 12, 301–320. [Google Scholar] [CrossRef]
- Moudgil, R.; Michelakis, E.D.; Archer, S.L. Hypoxic pulmonary vasoconstriction. J. Appl. Physiol. 2005, 98, 390–403. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.Q.; Jing, Z.C. High-altitude pulmonary hypertension. Eur. Respir. Rev. 2009, 18, 13–17. [Google Scholar] [CrossRef]
- Zhao, L.; Oliver, E.; Maratou, K.; Atanur, S.S.; Dubois, O.D.; Cotroneo, E.; Chen, C.N.; Wang, L.; Arce, C.; Chabosseau, P.L.; et al. The zinc transporter ZIP12 regulates the pulmonary vascular response to chronic hypoxia. Nature 2015, 524, 356–360. [Google Scholar] [CrossRef] [PubMed]
- Cruz, J.B.F.; Soares, H.F. Uma revisão sobre o zinco. Ens. Ciên. Ciên. Biol. Agrar. Saúde 2011, 15, 207–222. [Google Scholar]
- Cruz, K.J.; de Oliveira, A.R.; Marreiro Ddo, N. Antioxidant role of zinc in diabetes mellitus. World J. Diabetes 2015, 6, 333–337. [Google Scholar] [CrossRef]
- Lima, V.B.; Fde, A.S.; Bezerra, D.L.; Neto, J.M.M.; Ddo, N.M. Parameters of glycemic control and their relationship with zinc concentrations in blood and with superoxide dismutase enzyme activity in type 2 diabetes patients. Arq. Bras. Endocrinol. Metabol. 2011, 55, 701–707. [Google Scholar] [CrossRef]
- Vázquez-Lorente, H.; Molina-López, J.; Herrera-Quintana, L.; Gamarra-Morales, Y.; Quintero-Osso, B.; López-González, B.; Planells, E. Erythrocyte Zn concentration and antioxidant response after supplementation with Zn in a postmenopausal population. A double-blind randomized trial. Exp. Gerontol. 2022, 162, 111766. [Google Scholar] [CrossRef]
- Mohammadi, H.; Talebi, S.; Ghavami, A.; Rafiei, M.; Sharifi, S.; Faghihimani, Z.; Ranjbar, G.; Miraghajani, M.; Askari, G. Effects of zinc supplementation on inflammatory biomarkers and oxidative stress in adults: A systematic review and meta-analysis of randomized controlled trials. J. Trace Elem. Med. Biol. 2021, 68, 126857. [Google Scholar] [CrossRef]
- Mohammadifard, N.; Humphries, K.H.; Gotay, C.; Mena-Sánchez, G.; Salas-Salvadó, J.; Esmaillzadeh, A.; Ignaszewski, A.; Sarrafzadegan, N. Trace minerals intake: Risks and benefits for cardiovascular health. Crit. Rev. Food Sci. Nutr. 2019, 59, 1334–1346. [Google Scholar] [CrossRef]
- Maret, W. Zinc coordination environments in proteins as redox sensors and signal transducers. Antioxid. Redox Signal. 2006, 8, 1419–1441. [Google Scholar] [CrossRef]
- Maret, W. The redox biology of redox-inert zinc ions. Free Radic. Biol. Med. 2019, 134, 311–326. [Google Scholar] [CrossRef]
- Pena, E.; Siques, P.; Brito, J.; Arribas, S.M.; Böger, R.; Hannemann, J.; León-Velarde, F.; González, M.C.; López, M.R.; de Pablo, Á.L.L. Nox2 Upregulation and p38α MAPK Activation in Right Ventricular Hypertrophy of Rats Exposed to Long-Term Chronic Intermittent Hypobaric Hypoxia. Int. J. Mol. Sci. 2020, 21, 8576. [Google Scholar] [CrossRef]
- Bernal, P.J.; Leelavanichkul, K.; Bauer, E.; Cao, R.; Wilson, A.; Wasserloos, K.J.; Watkins, S.C.; Pitt, B.R.; St Croix, C.M. Nitric-oxide-mediated zinc release contributes to hypoxic regulation of pulmonary vascular tone. Circ. Res. 2008, 102, 1575–1583. [Google Scholar] [CrossRef]
- Littler, C.M.; Wehling, C.A.; Wick, M.J.; Fagan, K.A.; Cool, C.D.; Messing, R.O.; Dempsey, E.C. Divergent contractile and structural responses of the murine PKC-epsilon null pulmonary circulation to chronic hypoxia. Am. J. Physiol. Lung Cell Mol. Physiol. 2005, 289, L1083–L1093. [Google Scholar] [CrossRef]
- Bernal, P.J.; Bauer, E.M.; Cao, R.; Maniar, S.; Mosher, M.; Chen, J.; Wang, Q.J.; Glorioso, J.C.; Pitt, B.R.; Watkins, S.C.; et al. A role for zinc in regulating hypoxia-induced contractile events in pulmonary endothelium. Am. J. Physiol. Lung Cell Mol. Physiol. 2011, 300, L874–L886. [Google Scholar] [CrossRef]
- Littler, C.M.; Morris, K.G., Jr.; Fagan, K.A.; McMurtry, I.F.; Messing, R.O.; Dempsey, E.C. Protein kinase C-epsilon-null mice have decreased hypoxic pulmonary vasoconstriction. Am. J. Physiol. Heart Circ. Physiol. 2003, 284, H1321–H1331. [Google Scholar] [CrossRef] [PubMed]
- Arriaza, K.; Cuevas, C.; Pena, E.; Siques, P.; Brito, J. Impact of zinc on oxidative signaling pathways in the development of pulmonary vasoconstriction induced by hypobaric hypoxia. Int. J. Mol. Sci. 2022, 23, 6974. [Google Scholar] [CrossRef] [PubMed]
- Pena, E.; Brito, J.; El Alam, S.; Siques, P. Oxidative stress, kinase activity and inflammatory implications in right ventricular hypertrophy and heart failure under hypobaric hypoxia. Int. J. Mol. Sci. 2020, 21, 6421. [Google Scholar] [CrossRef]
- Bailey, D.M.; Taudorf, S.; Berg, R.M.; Lundby, C.; McEneny, J.; Young, I.S.; Evans, K.A.; James, P.E.; Shore, A.; Hullin, D.A.; et al. Increased cerebral output of free radicals during hypoxia: Implications for acute mountain sickness? Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 297, R1283–R1292. [Google Scholar] [CrossRef]
- Faoro, V.; Fink, B.; Taudorf, S.; Dehnert, C.; Berger, M.M.; Swenson, E.R.; Bailey, D.M.; Bärtsch, P.; Mairbäurl, H. Acute in vitro hypoxia and high-altitude (4,559 m) exposure decreases leukocyte oxygen consumption. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R32–R39. [Google Scholar] [CrossRef]
- Jefferson, J.A.; Simoni, J.; Escudero, E.; Hurtado, M.E.; Swenson, E.R.; Wesson, D.E.; Schreiner, G.F.; Schoene, R.B.; Johnson, R.J.; Hurtado, A. Increased oxidative stress following acute and chronic high altitude exposure. High Alt. Med. Biol. 2004, 5, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.J.; Sato, B.G.; Dalton, T.P.; Laderoute, K.R. The metal-responsive transcription factor-1 contributes to HIF-1 activation during hypoxic stress. Biochem. Biophys. Res. Commun. 2005, 337, 860–867. [Google Scholar] [CrossRef]
- Murphy, B.J.; Kimura, T.; Sato, B.G.; Shi, Y.; Andrews, G.K. Metallothionein induction by hypoxia involves cooperative interactions between metal-responsive transcription factor-1 and hypoxia-inducible transcription factor-1alpha. Mol. Cancer Res. 2008, 6, 483–490. [Google Scholar] [CrossRef]
- Dubé, A.; Harrisson, J.F.; Saint-Gelais, G.; Séguin, C. Hypoxia acts through multiple signaling pathways to induce metallothionein transactivation by the metal-responsive transcription factor-1 (MTF-1). Biochem. Cell Biol. 2011, 89, 562–577. [Google Scholar] [CrossRef] [PubMed]
- Simler, N.; Grosfeld, A.; Peinnequin, A.; Guerre-Millo, M.; Bigard, A.X. Leptin receptor-deficient obese Zucker rats reduce their food intake in response to hypobaric hypoxia. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E591–E597. [Google Scholar] [CrossRef] [PubMed]
- Flores, K.; Siques, P.; Brito, J.; Ordenes, S.; Arriaza, K.; Pena, E.; León-Velarde, F.; López, R.; de Pablo, Á.L.L.; Arribas, S. Lower body weight in rats under hypobaric hypoxia exposure would lead to reduced right ventricular hypertrophy and increased AMPK activation. Front. Physiol. 2020, 11, 342. [Google Scholar] [CrossRef]
- Gunga, H.C.; Röcker, L.; Behn, C.; Hildebrandt, W.; Koralewski, E.; Rich, I.; Schobersberger, W.; Kirsch, K. Shift working in the Chilean Andes (>3,600 m) and its influence on erythropoietin and the low-pressure system. J. Appl. Physiol. 1996, 81, 846–852. [Google Scholar] [CrossRef] [PubMed]
- Heinicke, K.; Prommer, N.; Cajigal, J.; Viola, T.; Behn, C.; Schmidt, W. Long-term exposure to intermittent hypoxia results in increased hemoglobin mass, reduced plasma volume, and elevated erythropoietin plasma levels in man. Eur. J. Appl. Physiol. 2003, 88, 535–543. [Google Scholar] [CrossRef]
- Akunov, A.; Sydykov, A.; Toktash, T.; Doolotova, A.; Sarybaev, A. Hemoglobin changes after long-term intermittent work at high altitude. Front. Physiol. 2018, 9, 1552. [Google Scholar] [CrossRef]
- Siques, P.; Brito, J.; Flores, K.; Ordenes, S.; Arriaza, K.; Pena, E.; León-Velarde, F.; de Pablo, Á.L.L.; Gonzalez, M.C.; Arribas, S. Long-term chronic intermittent hypobaric hypoxia induces glucose transporter (GLUT4) translocation through AMP-activated protein kinase (AMPK) in the soleus muscle in lean rats. Front. Physiol. 2018, 9, 799. [Google Scholar] [CrossRef]
- Chen, Y.H.; Shiu, J.R.; Ho, C.L.; Jeng, S.S. Zinc as a signal to stimulate red blood cell formation in fish. Int. J. Mol. Sci. 2017, 18, 138. [Google Scholar] [CrossRef]
- Chen, Y.H.; Feng, H.L.; Jeng, S.S. Zinc supplementation stimulates red blood cell formation in rats. Int. J. Mol. Sci. 2018, 19, 2824. [Google Scholar] [CrossRef]
- Baranauskas, M.N.; Powell, J.; Fly, A.D.; Martin, B.J.; Mickleborough, T.D.; Paris, H.L.; Chapman, R.F. Influence of zinc on the acute changes in erythropoietin and proinflammatory cytokines with hypoxia. High Alt. Med. Biol. 2021, 22, 148–156. [Google Scholar] [CrossRef]
- World Health Organization. Environment Health Criteria 221: Zinc. Environmental Health Criteria; WHO: Geneva, Switzerland, 2000.
- Domingo, J.L.; Llobet, J.M.; Paternain, J.L.; Corbella, J. Acute zinc intoxication: Comparison of the antidotal efficacy of several chelating agents. Vet. Hum. Toxicol. 1988, 30, 224–228. [Google Scholar]
- Rawal, S.B.; Singh, M.V.; Tyagi, A.K.; Roy, J.; Dimri, G.P.; Selvamurthy, W. Effect of time exposure to high altitude on zinc and copper concentrations in human plasma. Aviat. Space Environ. Med. 1999, 70, 1161–1165. [Google Scholar]
- Vats, P.; Singh, S.N.; Kumria, M.M.; Ranganathan, S.; Arora, M.P.; Jain, C.L.; Sridharan, K. Effect of hypoxia on the circulating levels of essential mineral elements in rats. J. Environ. Biol. 2001, 22, 277–282. [Google Scholar]
- Moser, P.B.; Borel, J.; Majerus, T.; Anderson, R.A. Serum zinc and urinary zinc excretion of trauma patients. Nutr. Res. 1985, 5, 253–261. [Google Scholar] [CrossRef]
- Jackson, M.J. Physiology of zinc: General aspects. In Zinc in Human Biology; Mills, C., Ed.; Springer: London, UK, 1989; pp. 1–14. [Google Scholar]
- Giugliano, R.; Millward, D.J. Growth and zinc homeostasis in the severely Zn-deficient rat. Br. J. Nutr. 1984, 52, 545–560. [Google Scholar] [CrossRef]
- Hernández-Camacho, J.D.; Vicente-García, C.; Parsons, D.S.; Navas-Enamorado, I. Zinc at the crossroads of exercise and proteostasis. Redox Biol. 2020, 35, 101529. [Google Scholar] [CrossRef]
- Kiilerich, S.; Christensen, M.S.; Naestoft, J.; Christiansen, C. Determination of zinc in serum and urine by atomic absorption spectrophotometry; relationship between serum levels of zinc and proteins in 104 normal subjects. Clin. Chim. Acta 1980, 105, 231–239. [Google Scholar] [CrossRef]
- Hurley, L.S.; Gordon, P.; Keen, C.L.; Merkhofer, L. Circadian variation in rat plasma zinc and rapid effect of dietary zinc deficiency. Proc. Soc. Exp. Biol. Med. 1982, 170, 48–52. [Google Scholar] [CrossRef]
- King, J.C. Zinc: An essential but elusive nutrient. Am. J. Clin. Nutr. 2011, 94, 679S–684S. [Google Scholar] [CrossRef]
- Monge, C.C.; León-Velarde, F. El reto fisiológico de vivir en los andes. In Travaux del Institut Francais d’Etudes Andines (IFEA); IFEA/Universidad Peruana Cayetano Heredia: Lima, Peru, 2003; Volume 177, p. 435. [Google Scholar]
- Siqués, P.; Brito, J.; León-Velarde, F.; Barrios, L.; Cruz, J.J.; López, V.; Herruzo, R. Time course of cardiovascular and hematological responses in rats exposed to chronic intermittent hypobaric hypoxia (4600 m). High Alt. Med. Biol. 2006, 7, 72–80. [Google Scholar] [CrossRef]
- Tham, Y.K.; Bernardo, B.C.; Ooi, J.Y.; Weeks, K.L.; McMullen, J.R. Pathophysiology of cardiac hypertrophy and heart failure: Signaling pathways and novel therapeutic targets. Arch. Toxicol. 2015, 89, 1401–1438. [Google Scholar] [CrossRef]
- Noordegraaf, A.V.; Westerhof, B.E.; Westerhof, N. The relationship between the right ventricle and its load in pulmonary hypertension. J. Am. Coll. Cardiol. 2017, 69, 236–243. [Google Scholar] [CrossRef]
- Siques, P.; de Pablo, A.L.L.; Brito, J.; Arribas, S.M.; Flores, K.; Arriaza, K.; Naveas, N.; González, M.C.; Hoorntje, A.; León-Velarde, F.; et al. Nitric oxide and superoxide anion balance in rats exposed to chronic and long term intermittent hypoxia. Biomed. Res. Int. 2014, 2014, 610474. [Google Scholar] [CrossRef]
- Powell, F.L.; Garcia, N. Physiological effects of intermittent hypoxia. High Alt. Med. Biol. 2000, 1, 125–136. [Google Scholar] [CrossRef]
- Uchida, T.; Rossignol, F.; Matthay, M.A.; Mounier, R.; Couette, S.; Clottes, E.; Clerici, C. Prolonged hypoxia differentially regulates hypoxia-inducible factor (HIF)-1alpha and HIF-2alpha expression in lung epithelial cells: Implication of natural antisense HIF-1alpha. J. Biol. Chem. 2004, 279, 14871–14878. [Google Scholar] [CrossRef]
- Nilsson, H.; Jögi, A.; Beckman, S.; Harris, A.L.; Poellinger, L.; Påhlman, S. HIF-2alpha expression in human fetal paraganglia and neuroblastoma: Relation to sympathetic differentiation, glucose deficiency, and hypoxia. Exp. Cell Res. 2005, 303, 447–456. [Google Scholar] [CrossRef]
- Holmquist-Mengelbier, L.; Fredlund, E.; Löfstedt, T.; Noguera, R.; Navarro, S.; Nilsson, H.; Pietras, A.; Vallon-Christersson, J.; Borg, A.; Gradin, K.; et al. Recruitment of HIF-1alpha and HIF-2alpha to common target genes is differentially regulated in neuroblastoma: HIF-2alpha promotes an aggressive phenotype. Cancer Cell 2006, 10, 413–423. [Google Scholar] [CrossRef]
- Koh, M.Y.; Lemos Jr, R.; Liu, X.; Powis, G. The hypoxia-associated factor switches cells from HIF-1α- to HIF-2α-dependent signaling promoting stem cell characteristics, aggressive tumor growth and invasion. Cancer Res. 2011, 71, 4015–4027. [Google Scholar] [CrossRef]
- Lin, Q.; Cong, X.; Yun, Z. Differential hypoxic regulation of hypoxia-inducible factors 1alpha and 2alpha. Mol. Cancer Res. 2011, 9, 757–765. [Google Scholar] [CrossRef]
- Dai, Z.; Li, M.; Wharton, J.; Zhu, M.M.; Zhao, Y.Y. Prolyl-4 hydroxylase 2 (PHD2) deficiency in endothelial cells and hematopoietic cells induces obliterative vascular remodeling and severe pulmonary arterial hypertension in mice and humans through hypoxia-inducible factor-2α. Circulation 2016, 133, 2447–2458. [Google Scholar] [CrossRef]
- Bartoszewski, R.; Moszyńska, A.; Serocki, M.; Cabaj, A.; Polten, A.; Ochocka, R.; Dell’Italia, L.; Bartoszewska, S.; Króliczewski, J.; Dąbrowski, M.; et al. Primary endothelial cell-specific regulation of hypoxia-inducible factor (HIF)-1 and HIF-2 and their target gene expression profiles during hypoxia. FASEB J. 2019, 33, 7929–7941. [Google Scholar] [CrossRef]
- Hu, C.J.; Poth, J.M.; Zhang, H.; Flockton, A.; Laux, A.; Kumar, S.; McKeon, B.; Mouradian, G.; Li, M.; Riddle, S.; et al. Suppression of HIF2 signalling attenuates the initiation of hypoxia-induced pulmonary hypertension. Eur. Respir. J. 2019, 54, 1900378. [Google Scholar] [CrossRef]
- Nanduri, J.; Wang, N.; Yuan, G.; Khan, S.A.; Souvannakitti, D.; Peng, Y.J.; Kumar, G.K.; Garcia, J.A.; Prabhakar, N.R. Intermittent hypoxia degrades HIF-2alpha via calpains resulting in oxidative stress: Implications for recurrent apnea-induced morbidities. Proc. Natl. Acad. Sci. USA 2009, 106, 1199–1204. [Google Scholar] [CrossRef]
- Nakajima, E.; Hammond, K.B.; Hirata, M.; Shearer, T.R.; Azuma, M. Contribution of Calpain and Caspases to Cell Death in Cultured Monkey RPE Cells. Investig. Ophthalmol. Vis. Sci. 2017, 58, 5412–5420. [Google Scholar] [CrossRef]
- Pullamsetti, S.S.; Mamazhakypov, A.; Weissmann, N.; Seeger, W.; Savai, R. Hypoxia-inducible factor signaling in pulmonary hypertension. J. Clin. Investig. 2020, 130, 5638–5651. [Google Scholar] [CrossRef]
- Saydam, N.; Georgiev, O.; Nakano, M.Y.; Greber, U.F.; Schaffner, W. Nucleo-cytoplasmic trafficking of metal-regulatory transcription factor 1 is regulated by diverse stress signals. J. Biol. Chem. 2001, 276, 25487–25495. [Google Scholar] [CrossRef]
- Lee, M.; Won, Y.; Shin, Y.; Kim, J.H.; Chun, J.S. Reciprocal activation of hypoxia-inducible factor (HIF)-2alpha and the zinc-ZIP8-MTF1 axis amplifies catabolic signaling in osteoarthritis. Osteoarthr. Cartil. 2016, 24, 134–145. [Google Scholar] [CrossRef]
- Murphy, B.J. Regulation of malignant progression by the hypoxia-sensitive transcription factors HIF-1alpha and MTF-1. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2004, 139, 495–507. [Google Scholar] [CrossRef]
- Valsecchi, V.; Laudati, G.; Cuomo, O.; Sirabella, R.; Annunziato, L.; Pignataro, G. The hypoxia sensitive metal transcription factor MTF-1 activates NCX1 brain promoter and participates in remote postconditioning neuroprotection in stroke. Cell Death Dis. 2021, 12, 423. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Yin, X.; Wang, Y.; Tan, Y.; Cai, L.; Wang, B.; Cai, J.; Fu, Y. Intermittent hypoxia-induced renal antioxidants and oxidative damage in male mice: Hormetic dose response. Dose-Response 2012, 11, 385–400. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Wang, Y.; Tan, Y.; Cai, X.; Cai, L.; Cai, J.; Zheng, Y. Deletion of metallothionein exacerbates intermittent hypoxia-induced oxidative and inflammatory injury in aorta. Oxid. Med. Cell Longev. 2014, 2014, 141053. [Google Scholar] [CrossRef] [PubMed]
- Schanz, M.; Schaaf, L.; Dippon, J.; Biegger, D.; Fritz, P.; Alscher, M.D.; Kimmel, M. Renal effects of metallothionein induction by zinc in vitro and in vivo. BMC Nephrol. 2017, 18, 91. [Google Scholar] [CrossRef]
- Colvin, R.A.; Holmes, W.R.; Fontaine, C.P.; Maret, W. Cytosolic zinc buffering and muffling: Their role in intracellular zinc homeostasis. Metallomics 2010, 2, 306–317. [Google Scholar] [CrossRef]
- Thompson, M.W. Regulation of zinc-dependent enzymes by metal carrier proteins. Biometals 2022, 35, 187–213. [Google Scholar] [CrossRef]
- Ye, C.; Lian, G.; Wang, T.; Chen, A.; Chen, W.; Gong, J.; Luo, L.; Wang, H.; Xie, L. The zinc transporter ZIP12 regulates monocrotaline-induced proliferation and migration of pulmonary arterial smooth muscle cells via the AKT/ERK signaling pathways. BMC Pulm. Med. 2022, 22, 111. [Google Scholar] [CrossRef]
- Adamson, I.Y.; Prieditis, H.; Hedgecock, C.; Vincent, R. Zinc is the toxic factor in the lung response to an atmospheric particulate sample. Toxicol. Appl. Pharmacol. 2000, 166, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Siques, P.; Pena, E.; Brito, J.; El Alam, S. Oxidative Stress, Kinase Activation, and Inflammatory Pathways Involved in Effects on Smooth Muscle Cells During Pulmonary Artery Hypertension Under Hypobaric Hypoxia Exposure. Front. Physiol. 2021, 12, 690341. [Google Scholar] [CrossRef] [PubMed]
- Brito, J.; Siques, P.; Arribas, S.M.; de Pablo, A.L.L.; González, M.C.; Naveas, N.; Arriaza, K.; Flores, K.; León-Velarde, F.; Pulido, R.; et al. Adventitial alterations are the main features in pulmonary artery remodeling due to long-term chronic intermittent hypobaric hypoxia in rats. Biomed. Res. Int. 2015, 2015, 169841. [Google Scholar] [CrossRef]
- Ball, M.K.; Waypa, G.B.; Mungai, P.T.; Nielsen, J.M.; Czech, L.; Dudley, V.J.; Beussink, L.; Dettman, R.W.; Berkelhamer, S.K.; Steinhorn, R.H.; et al. Regulation of hypoxia-induced pulmonary hypertension by vascular smooth muscle hypoxia-inducible factor-1α. Am. J. Respir. Crit. Care Med. 2014, 189, 314–324. [Google Scholar] [CrossRef]
- Slepchenko, K.G.; Lu, Q.; Li, Y.V. Zinc wave during the treatment of hypoxia is required for initial reactive oxygen species activation in mitochondria. Int. J. Physiol. Pathophysiol. Pharmacol. 2016, 8, 44–51. [Google Scholar]
- Gonzales, G.F.; Tapia, V.; Gasco, M.; Rubio, J.; Gonzales-Castañeda, C. High serum zinc and serum testosterone levels were associated with excessive erythrocytosis in men at high altitudes. Endocrine 2011, 40, 472–480. [Google Scholar] [CrossRef]
- Kay, J.M. Effect of intermittent normoxia on chronic hypoxic pulmonary hypertension, right ventricular hypertrophy, and polycythemia in rats. Am. Rev. Respir. Dis. 1980, 121, 993–1001. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VARIABLES | NX | CIH | CH |
|---|---|---|---|
| BODY WEIGHT (gr) | |||
| Day 0 saline zinc | 310.50 ± 10.31 | 303.13 ± 12.01 | 317.50 ± 7.21 |
| 305.63 ± 9.46 | 322.38 ± 8.66 | 323.29 ± 11.67 | |
| Day 30 saline zinc | 326.13 ± 7.95 & | 277.38 ± 13.59 *& | 295.38 ± 10.02 & |
| 328.00 ± 7.82 & | 280.88 ± 13.72 *& | 297.00 ± 11.60 & | |
| HEMOGLOBIN (mg/dL) | |||
| Day 0 saline zinc | 14.92 ± 0.39 | 15.37 ± 0.47 | 14.83 ± 0.25 |
| 15.44 ± 0.38 | 15.40 ± 0.24 | 14.42 ± 0.30 *# | |
| Day 30 saline zinc | 14.46 ± 0.32 | 21.05 ± 0.47 *& | 23.30 ± 0.28 *#& |
| 14.91 ± 0.21 | 20.45 ± 0.40 *& | 22.84 ± 0.52 *#& | |
| PLASMA ZINC (µg/mL) | |||
| Day 0 saline zinc | 2.14 ± 0.16 | 2.27 ± 0.18 | 2.20 ± 0.17 |
| 2.18 ± 0.19 | 2.12 ± 0.17 | 2.25 ± 0.15 | |
| Day 30 saline zinc | 2.03 ± 0.21 | 0.90 ± 0.85 *& | 0.74 ± 0.08 *& |
| 1.99 ± 0.09 | 1.44 ± 0.06 *& | 1.25 ± 0.14 *&† | |
| ZINC LUNG (mg/mL) | |||
| Day 30 saline zinc | 103.69 ± 3.95 | 93.83 ± 3.57 | 102.42 ± 3.76 |
| 111.95 ± 3.44 | 91.66 ± 1.71 * | 94.14 ± 2.24 * | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arriaza, K.; Brito, J.; Siques, P.; Flores, K.; Ordenes, S.; Aguayo, D.; López, M.d.R.; Arribas, S.M. Effects of Zinc on the Right Cardiovascular Circuit in Long-Term Hypobaric Hypoxia in Wistar Rats. Int. J. Mol. Sci. 2023, 24, 9567. https://doi.org/10.3390/ijms24119567
Arriaza K, Brito J, Siques P, Flores K, Ordenes S, Aguayo D, López MdR, Arribas SM. Effects of Zinc on the Right Cardiovascular Circuit in Long-Term Hypobaric Hypoxia in Wistar Rats. International Journal of Molecular Sciences. 2023; 24(11):9567. https://doi.org/10.3390/ijms24119567
Chicago/Turabian StyleArriaza, Karem, Julio Brito, Patricia Siques, Karen Flores, Stefany Ordenes, Daniel Aguayo, María del Rosario López, and Silvia M. Arribas. 2023. "Effects of Zinc on the Right Cardiovascular Circuit in Long-Term Hypobaric Hypoxia in Wistar Rats" International Journal of Molecular Sciences 24, no. 11: 9567. https://doi.org/10.3390/ijms24119567
APA StyleArriaza, K., Brito, J., Siques, P., Flores, K., Ordenes, S., Aguayo, D., López, M. d. R., & Arribas, S. M. (2023). Effects of Zinc on the Right Cardiovascular Circuit in Long-Term Hypobaric Hypoxia in Wistar Rats. International Journal of Molecular Sciences, 24(11), 9567. https://doi.org/10.3390/ijms24119567