

Rice OsMRG702 and Its Partner OsMRGBP Control Flowering Time through H4 Acetylation
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
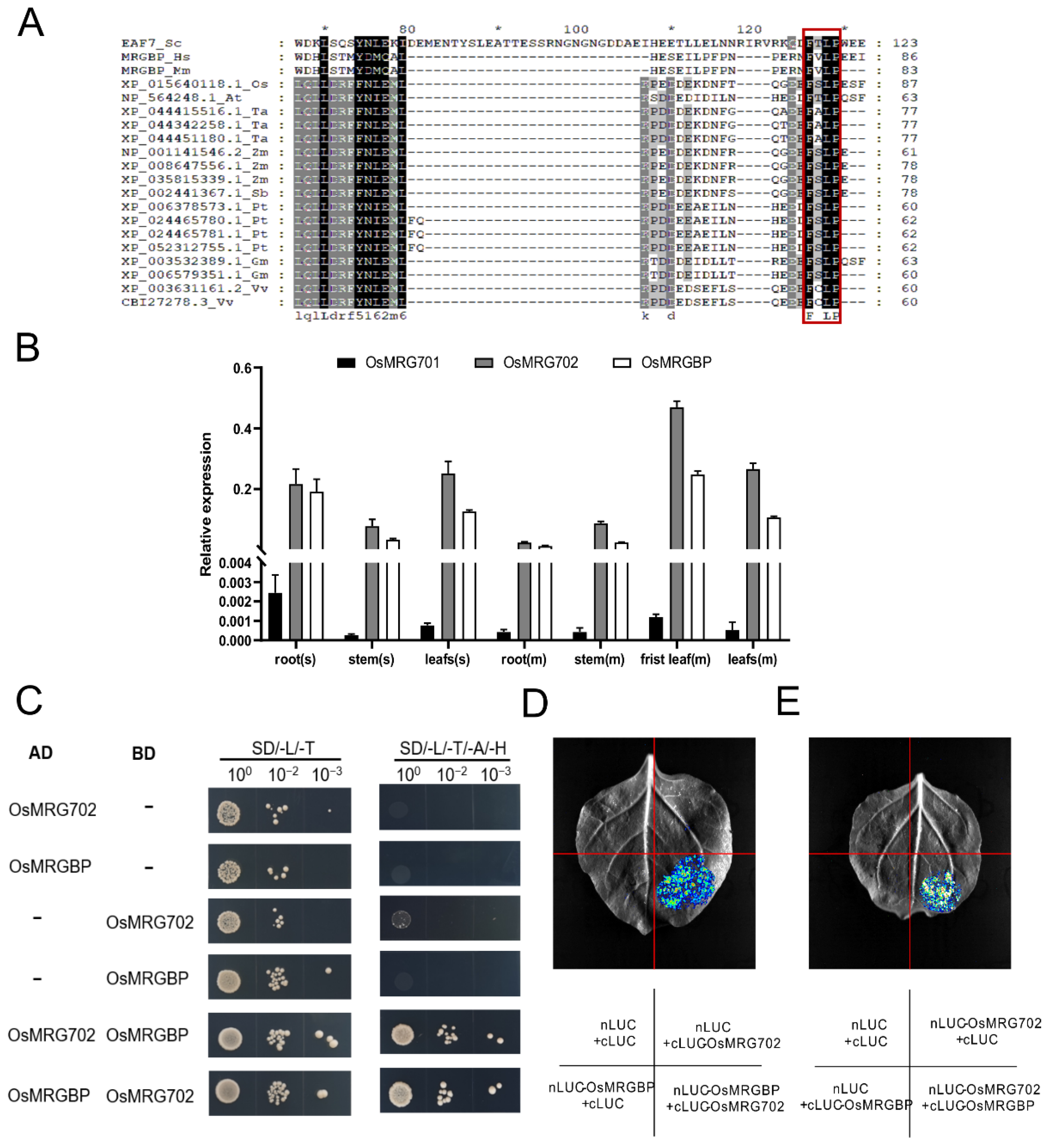
2.1. OsMRGBP Directly Interacts with OsMRG702
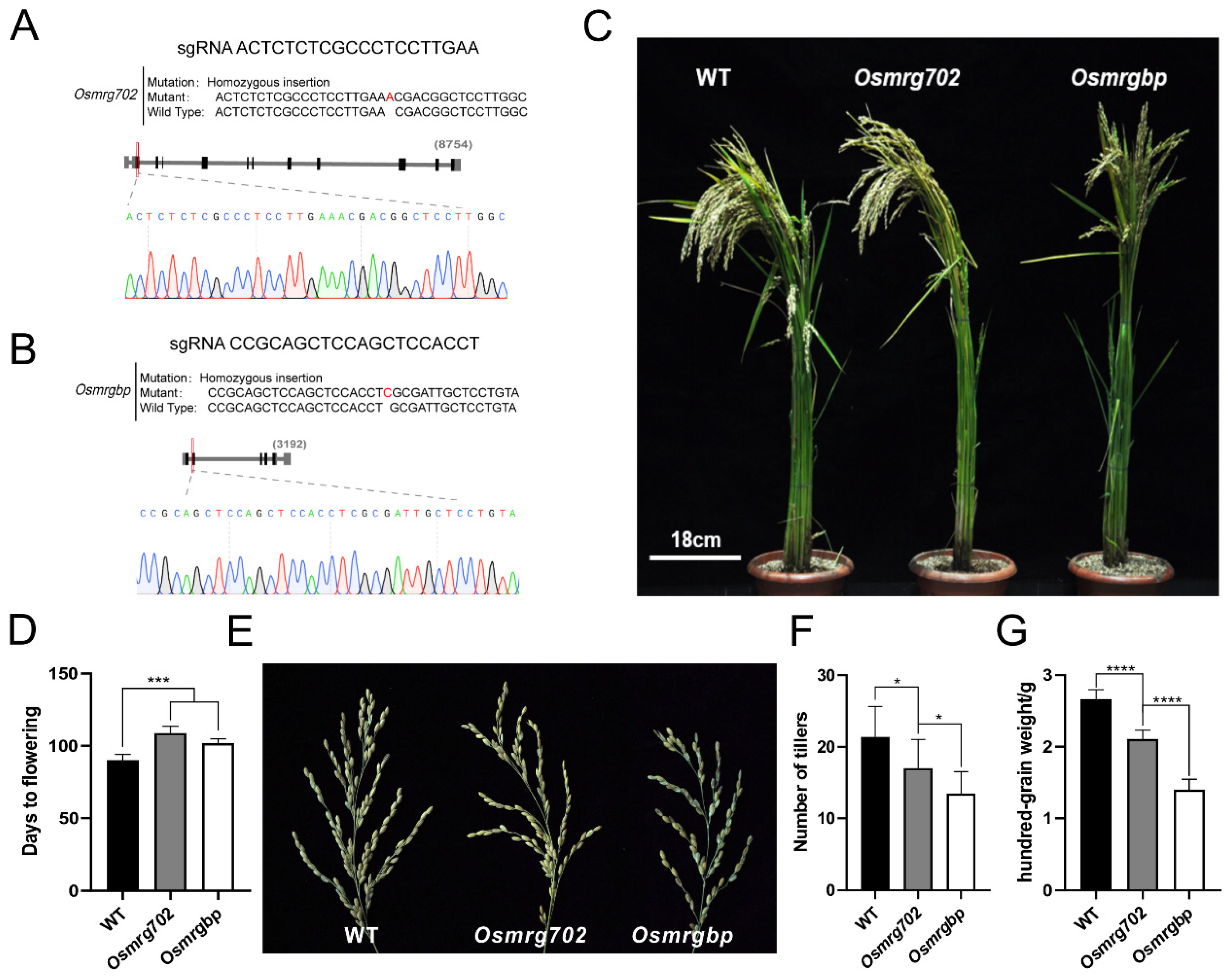
2.2. Flowering Time Is Delayed in Osmrg702 and Osmrgbp Mutants
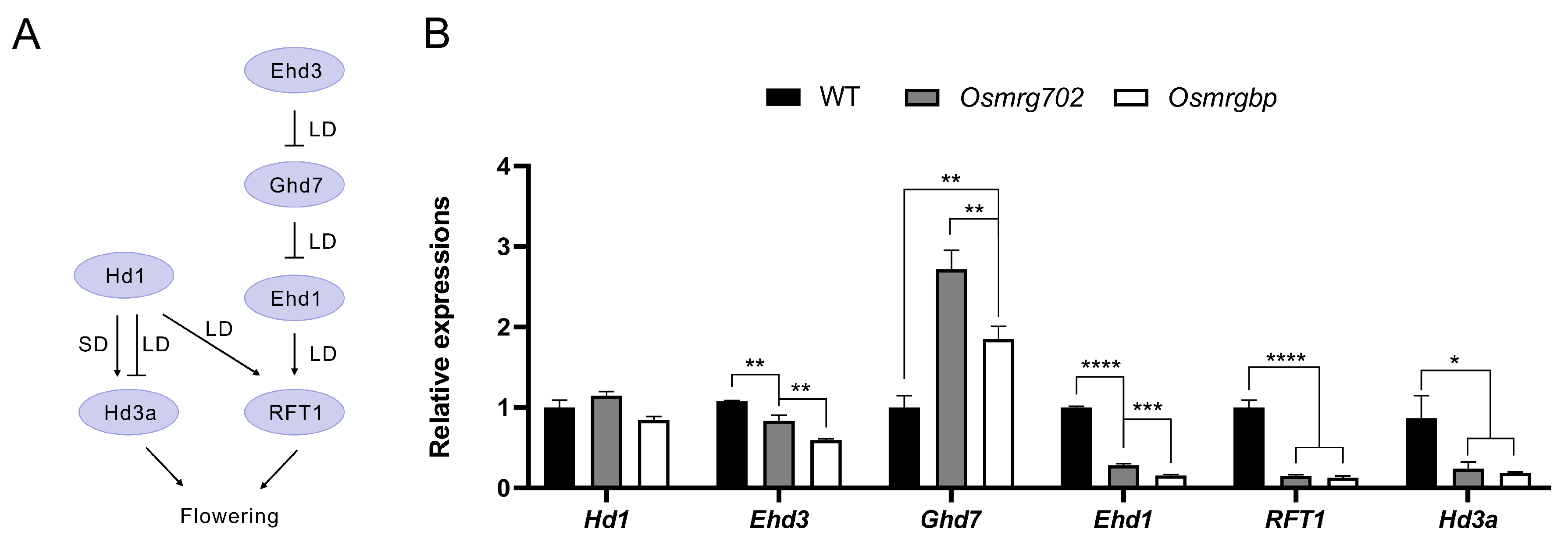
2.3. Multiple Flowering Genes Are Altered in Expression Levels in Osmrg702 and Osmrgbp Mutants
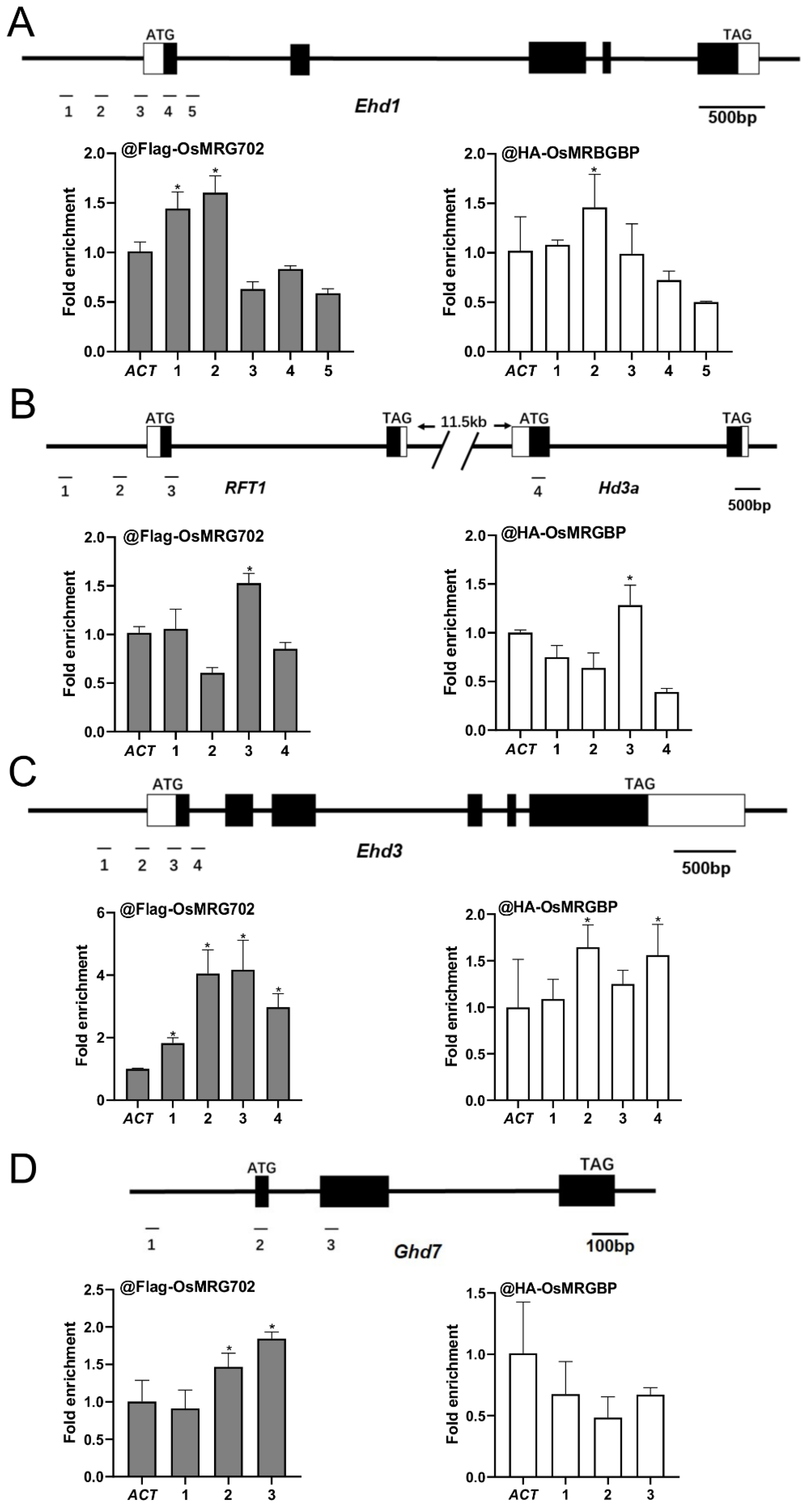
2.4. OsMRG702 and OsMRGBP Directly Bind to the Loci of Multiple Key Flowering Genes
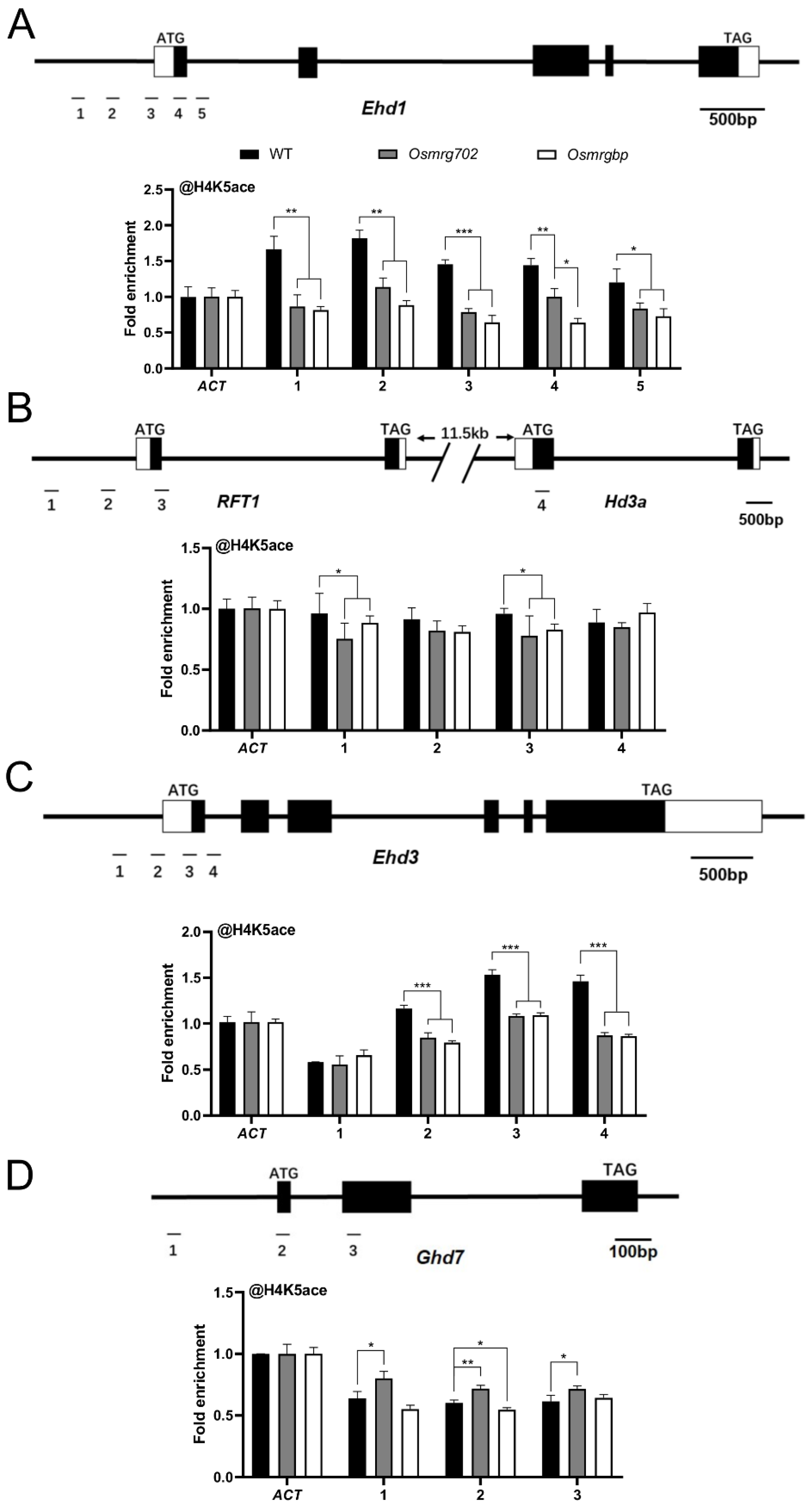
2.5. The H4K5ac Level of the Key Flowering Time Genes Were Decreased in Osmrg702 and Osmrgbp
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. CRISPR/Cas9 for Genome Editing
4.3. Plasmid Construction
4.4. Yeast Two-Hybrid (Y2H)
4.5. Luciferase Complementation Assay (LCA)
4.6. Quantitative RT-PCR
4.7. Chromatin Immunoprecipitation (ChIP)
4.8. Nuclear Extractions and Western Blot
4.9. Measurement of Agronomic Traits
4.10. Phylogenetic Analysis
4.11. Accession Numbers
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Silva, C.S.; Puranik, S.; Round, A.; Brennich, M.; Jourdain, A.; Parcy, F.; Hugouvieux, V.; Zubieta, C. Evolution of the Plant Reproduction Master Regulators LFY and the MADS Transcription Factors: The Role of Protein Structure in the Evolutionary Development of the Flower. Front. Plant Sci. 2015, 6, 1193. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; An, H.R.; Tong, C.G.; Jang, S. Flowering and flowering genes: From model plants to orchids. Hortic. Environ. Biotechnol. 2021, 62, 135–148. [Google Scholar] [CrossRef]
- Corbesier, L.; Vincent, C.; Jang, S.H.; Fornara, F.; Fan, Q.Z.; Searle, I.; Giakountis, A.; Farrona, S.; Gissot, L.; Turnbull, C.; et al. FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis. Science 2007, 316, 1030–1033. [Google Scholar] [CrossRef]
- Tiwari, S.B.; Shen, Y.; Chang, H.C.; Hou, Y.L.; Harris, A.; Ma, S.F.; McPartland, M.; Hymus, G.J.; Adam, L.; Marion, C.; et al. The flowering time regulator constans is recruited to the FLOWERING LOCUS T promoter via a unique cis-element. New Phytol. 2010, 187, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Kojima, S.; Takahashi, Y.; Kobayashi, Y.; Monna, L.; Sasaki, T.; Araki, T.; Yano, M. Hd3a, a rice ortholog of the Arabidopsis FT gene, promotes transition to flowering downstream of Hd1 under short-day conditions. Plant Cell Physiol. 2002, 43, 1096–1105. [Google Scholar] [CrossRef] [PubMed]
- Komiya, R.; Ikegami, A.; Tamaki, S.; Yokoi, S.; Shimamoto, K. Hd3a and RFT1 are essential for flowering in rice. Development 2008, 135, 767–774. [Google Scholar] [CrossRef]
- Hayama, R.; Yokoi, S.; Tamaki, S.; Yano, M.; Shimamoto, K. Adaptation of photoperiodic control pathways produces short-day flowering in rice. Nature 2003, 422, 719–722. [Google Scholar] [CrossRef]
- Komiya, R.; Yokoi, S.; Shimamoto, K. A gene network for long-day flowering activates RFT1 encoding a mobile flowering signal in rice. Development 2009, 136, 3443–3450. [Google Scholar] [CrossRef]
- Sun, C.H.; Chen, D.; Fang, J.; Wang, P.R.; Deng, X.J.; Chu, C.C. Understanding the genetic and epigenetic architecture in complex network of rice flowering pathways. Protein Cell 2014, 5, 889–898. [Google Scholar] [CrossRef]
- Matsubara, K.; Yamanouchi, U.; Nonoue, Y.; Sugimoto, K.; Wang, Z.X.; Minobe, Y.; Yano, M. Ehd3, encoding a plant homeodomain finger-containing protein, is a critical promoter of rice flowering. Plant J. 2011, 66, 603–612. [Google Scholar] [CrossRef]
- Choi, S.C.; Lee, S.; Kim, S.R.; Lee, Y.S.; Liu, C.Y.; Cao, X.F.; An, G. Trithorax Group Protein Oryza sativa Trithorax1 Controls Flowering Time in Rice via Interaction with Early heading date3. Plant Physiol. 2014, 164, 1326–1337. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.Y.; Xing, Y.Z.; Weng, X.Y.; Zhao, Y.; Tang, W.J.; Wang, L.; Zhou, H.J.; Yu, S.B.; Xu, C.G.; Li, X.H.; et al. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nat. Genet. 2008, 40, 761–767. [Google Scholar] [CrossRef]
- Weng, X.Y.; Wang, L.; Wang, J.; Hu, Y.; Du, H.; Xu, C.G.; Xing, Y.Z.; Li, X.H.; Xiao, J.H.; Zhang, Q.F. Grain Number, Plant Height, and Heading Date7 Is a Central Regulator of Growth, Development, and Stress Response. Plant Physiol. 2014, 164, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Doi, K.; Izawa, T.; Fuse, T.; Yamanouchi, U.; Kubo, T.; Shimatani, Z.; Yano, M.; Yoshimura, A. Ehd1, a B-type response regulator in rice, confers short-day promotion of flowering and controls FT-Iike gene expression independently of Hd1l. Genes Dev. 2004, 18, 926–936. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y. Modern epigenetics methods in biological research. Methods 2021, 187, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.; Choi, W.L.; Lim, J.Y.; Huh, J.H. Epigenome editing: Targeted manipulation of epigenetic modifications in plants. Genes Genom. 2022, 44, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Kouzarides, T. Chromatin modifications and their function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Shirvaliloo, M. The landscape of histone modifications in epigenomics since 2020. Epigenomics 2022, 14, 1465–1477. [Google Scholar] [CrossRef]
- Thorstensen, T.; Grini, P.E.; Aalen, R.B. SET domain proteins in plant development. Biochim. Biophys. Acta Gene Regul. Mech. 2011, 1809, 407–420. [Google Scholar] [CrossRef]
- Sun, C.H.; Fang, J.; Zhao, T.L.; Xu, B.; Zhang, F.T.; Liu, L.C.; Tang, J.Y.; Zhang, G.F.; Deng, X.J.; Chen, F.; et al. The Histone Methyltransferase SDG724 Mediates H3K36me2/3 Deposition at MADS50 and RFT1 and Promotes Flowering in Rice. Plant Cell 2012, 24, 3235–3247. [Google Scholar] [CrossRef] [PubMed]
- Sui, P.F.; Shi, J.L.; Gao, X.Y.; Shen, W.H.; Dong, A.W. H3K36 Methylation Is Involved in Promoting Rice Flowering. Mol. Plant. 2013, 6, 975–977. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Wei, G.; Shi, J.L.; Jin, J.; Shen, T.; Ni, T.; Shen, W.H.; Yu, Y.; Dong, A.W. SET DOMAIN GROUP 708, a histone H3 lysine 36-specific methyltransferase, controls flowering time in rice (Oryza sativa). New Phytol. 2016, 210, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Yun, M.; Wu, J.; Workman, J.L.; Li, B. Readers of histone modifications. Cell Res. 2011, 21, 564–578. [Google Scholar] [CrossRef] [PubMed]
- Devoucoux, M.; Roques, C.; Lachance, C.; Lashgari, A.; Joly-Beauparlant, C.; Jacquet, K.; Alerasool, N.; Prudente, A.; Taipale, M.; Droit, A.; et al. MRG Proteins Are Shared by Multiple Protein Complexes with Distinct Functions. Mol. Cell. Proteom. 2022, 21, 100253. [Google Scholar] [CrossRef] [PubMed]
- Sathianathan, A.; Ravichandran, P.; Lippi, J.M.; Cohen, L.; Messina, A.; Shaju, S.; Swede, M.J.; Ginsburg, D.S. The Eaf3/5/7 Subcomplex Stimulates NuA4 Interaction with Methylated Histone H3 Lys-36 and RNA Polymerase II. J. Biol. Chem. 2016, 291, 21195–21207. [Google Scholar] [CrossRef]
- Xu, Y.F.; Gan, E.S.; Zhou, J.; Wee, W.Y.; Zhang, X.Y.; Ito, T. Arabidopsis MRG domain proteins bridge two histone modifications to elevate expression of flowering genes. Nucleic Acids Res. 2014, 42, 10960–10974. [Google Scholar] [CrossRef]
- Bu, Z.Y.; Yu, Y.; Li, Z.P.; Liu, Y.C.; Jiang, W.; Huang, Y.; Dong, A.W. Regulation of Arabidopsis Flowering by the Histone Mark Readers MRG1/2 via Interaction with CONSTANS to Modulate FT Expression. PLoS Genet. 2014, 10, e1004617. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.H.; Li, Z.P.; Liu, Y.H.; An, Z.X.; Peng, M.L.; Shen, W.H.; Dong, A.W.; Yu, Y. MRG1/2 histone methylation readers and HD2C histone deacetylase associate in repression of the florigen gene FT to set a proper flowering time in response to day-length changes. New Phytol. 2020, 227, 1453–1466. [Google Scholar] [CrossRef]
- An, Z.X.; Yin, L.F.; Liu, Y.H.; Peng, M.L.; Shen, W.H.; Dong, A.W. The histone methylation readers MRG1/MRG2 and the histone chaperones NRP1/NRP2 associate in fine-tuning Arabidopsis flowering time. Plant J. 2020, 103, 1010–1024. [Google Scholar] [CrossRef]
- Jin, J.; Shi, J.L.; Liu, B.; Liu, Y.C.; Huang, Y.; Yu, Y.; Dong, A.W. MORF-RELATED GENE702, a Reader Protein of Trimethylated Histone H3 Lysine 4 and Histone H3 Lysine 36, Is Involved in Brassinosteroid-Regulated Growth and Flowering Time Control in Rice. Plant Physiol. 2015, 168, 1275–1285. [Google Scholar] [CrossRef]
- Liu, Y.C.; Wu, H.; Yu, Y.; Huang, Y. Structural studies on MRG701 chromodomain reveal a novel dimerization interface of MRG proteins in green plants. Protein Cell 2016, 7, 792–803. [Google Scholar] [CrossRef] [PubMed]
- Rossetto, D.; Cramet, M.; Wang, A.Y.; Steunou, A.L.; Lacoste, N.; Schulze, J.M.; Cote, V.; Monnet-Saksouk, J.; Piquet, S.; Nourani, A.; et al. Eaf5/7/3 form a functionally independent NuA4 submodule linked to RNA polymerase II-coupled nucleosome recycling. EMBO J. 2014, 33, 1397–1415. [Google Scholar] [CrossRef] [PubMed]
- Dalwadi, U.; Corrado, E.; Fleming, K.D.; Moeller, B.E.; Nam, S.E.; Burke, J.E.; Yip, C.K. Biochemical Characterization of the TINTIN Module of the NuA4 Complex Reveals Allosteric Regulation of Nucleosome Interaction. Mol. Cell. Biol. 2022, 42, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.Y.; Xia, P.; Wing, R.A.; Zhang, Q.F.; Luo, M.Z. Dynamic Intra-Japonica Subspecies Variation and Resource Application. Mol. Plant 2012, 5, 218–230. [Google Scholar] [CrossRef]
- Liu, B.; Liu, Y.H.; Wang, B.H.; Luo, Q.; Shi, J.L.; Gan, J.H.; Shen, W.H.; Yu, Y.; Dong, A.W. The transcription factor OsSUF4 interacts with SDG725 in promoting H3K36me3 establishment. Nat. Commun. 2019, 10, 2999. [Google Scholar] [CrossRef]
- Ul Ikram, A.; Zhang, F.; Xu, Z.T.; Li, E.Z.; Xue, G.; Wang, S.L.; Zhang, C.; Yang, Y.C.; Su, Y.H.; Ding, Y. Chromatin remodeling factors OsYAF9 and OsSWC4 interact to promote internode elongation in rice. Plant Physiol. 2022, 188, 2199–2214. [Google Scholar] [CrossRef] [PubMed]
- Tang, W. Additional virulence genes and sonication enhance Agrobacterium tumefaciens-mediated loblolly pine transformation. Plant Cell Rep. 2003, 21, 555–562. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, F.; Zhang, K.; Wen, J.; Yu, S.; Li, W.; Du, G.; Wu, C.; Zhu, K.; Xu, Y. Rice OsMRG702 and Its Partner OsMRGBP Control Flowering Time through H4 Acetylation. Int. J. Mol. Sci. 2023, 24, 9219. https://doi.org/10.3390/ijms24119219
Gong F, Zhang K, Wen J, Yu S, Li W, Du G, Wu C, Zhu K, Xu Y. Rice OsMRG702 and Its Partner OsMRGBP Control Flowering Time through H4 Acetylation. International Journal of Molecular Sciences. 2023; 24(11):9219. https://doi.org/10.3390/ijms24119219
Chicago/Turabian StyleGong, Feng, Kaixin Zhang, Jing Wen, Shenbo Yu, Wenjin Li, Gaofeng Du, Cui Wu, Kangjing Zhu, and Yifeng Xu. 2023. "Rice OsMRG702 and Its Partner OsMRGBP Control Flowering Time through H4 Acetylation" International Journal of Molecular Sciences 24, no. 11: 9219. https://doi.org/10.3390/ijms24119219
APA StyleGong, F., Zhang, K., Wen, J., Yu, S., Li, W., Du, G., Wu, C., Zhu, K., & Xu, Y. (2023). Rice OsMRG702 and Its Partner OsMRGBP Control Flowering Time through H4 Acetylation. International Journal of Molecular Sciences, 24(11), 9219. https://doi.org/10.3390/ijms24119219