
Spatial Distribution and Temporal Dynamics of Candidatus Liberibacter Asiaticus in Different Stages of Embryos, Nymphs and Adults of Diaphorina citri
{kind=link}
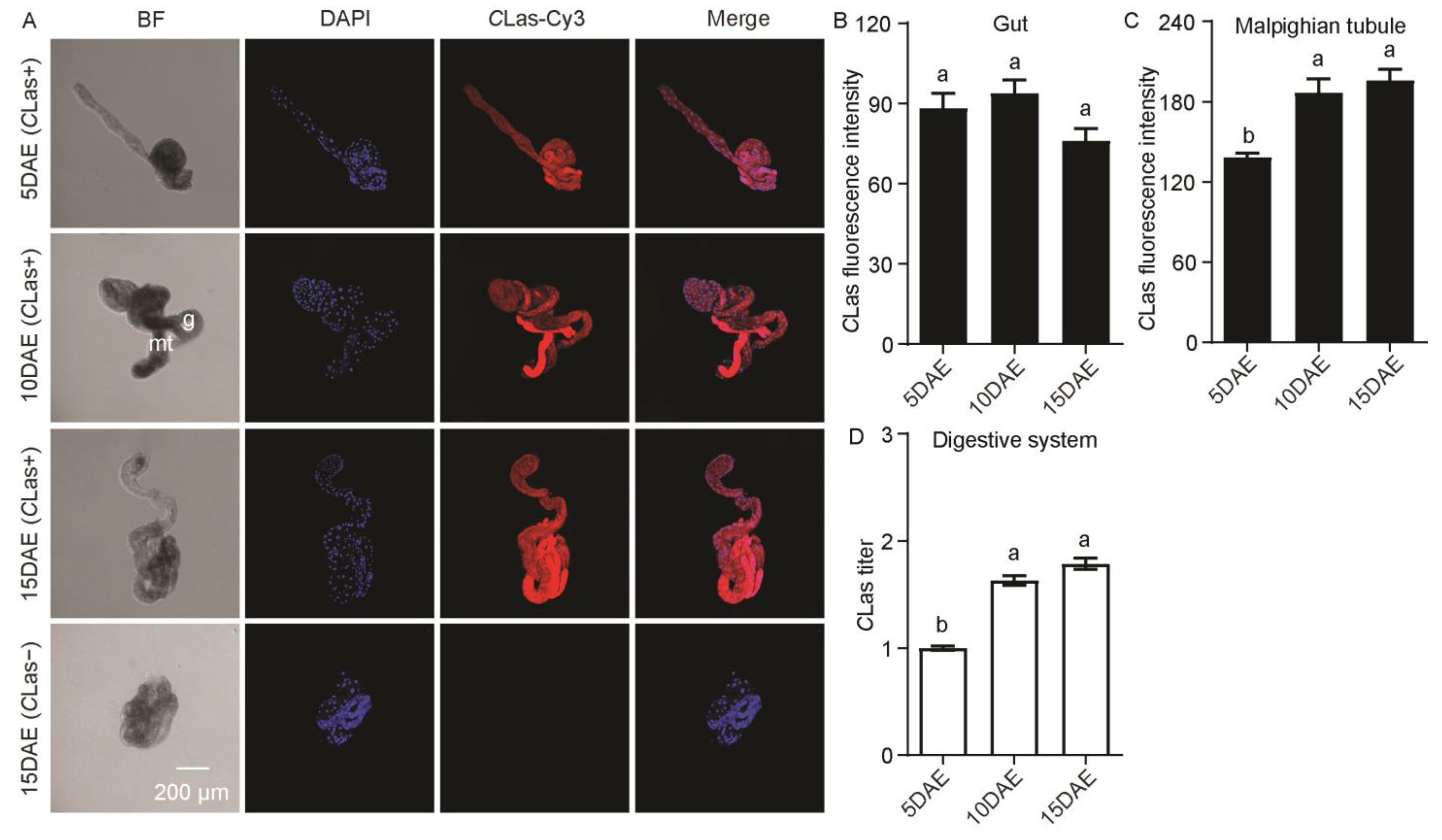{kind=link}
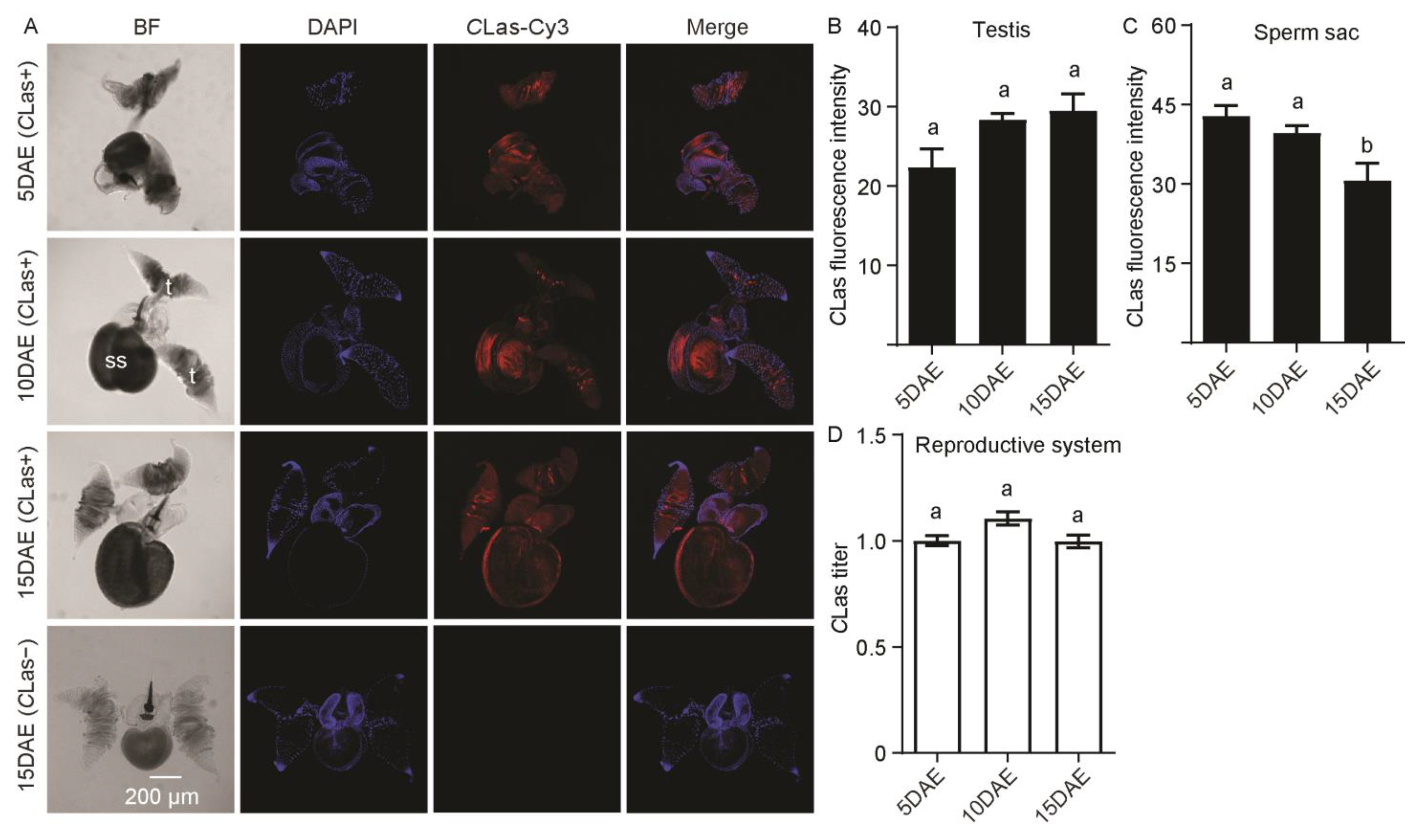{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Distribution and Relative Titers of CLas in CLas-Positive Adult Males
2.2. Distribution and Relative Titers of CLas in CLas-Positive Adult Females
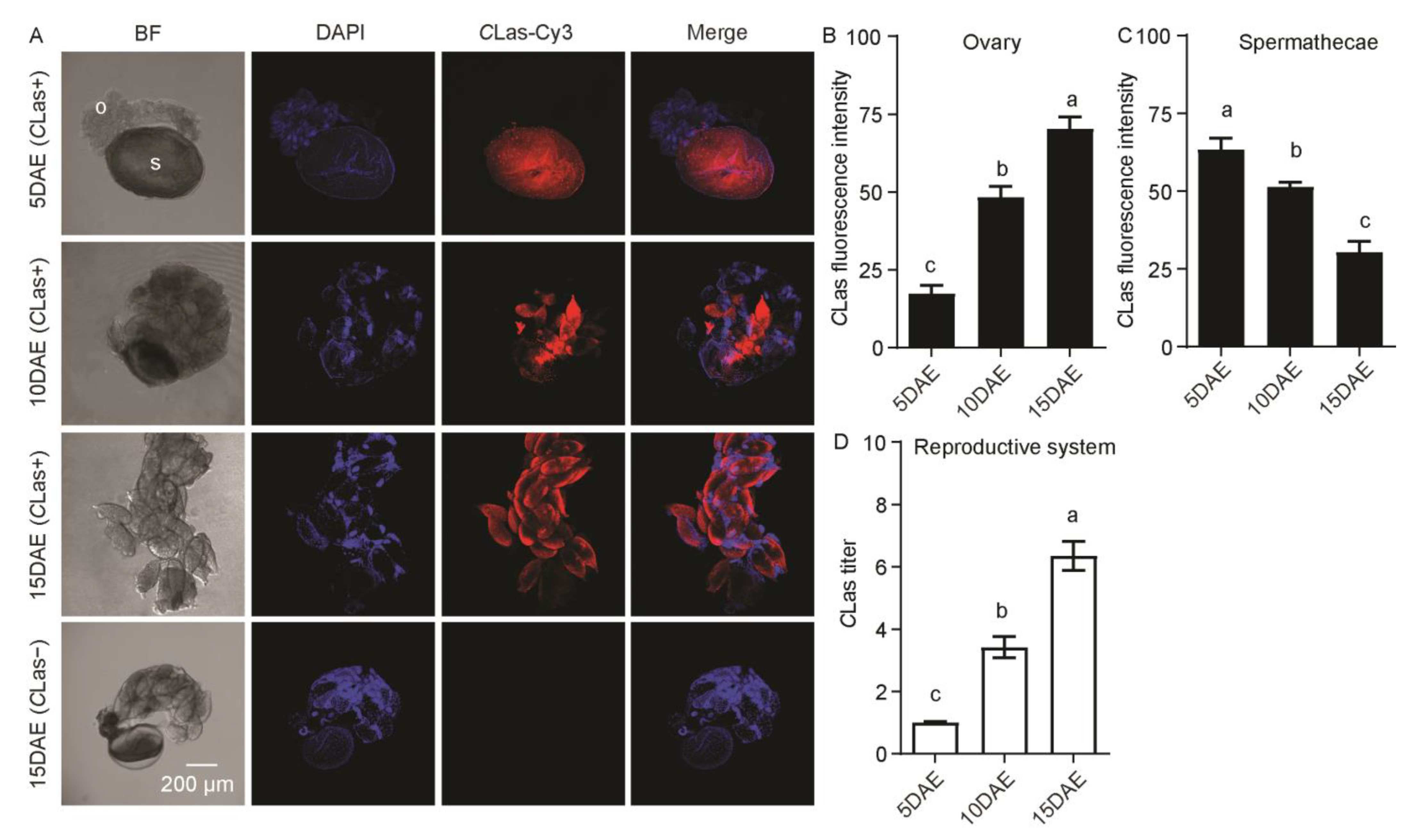
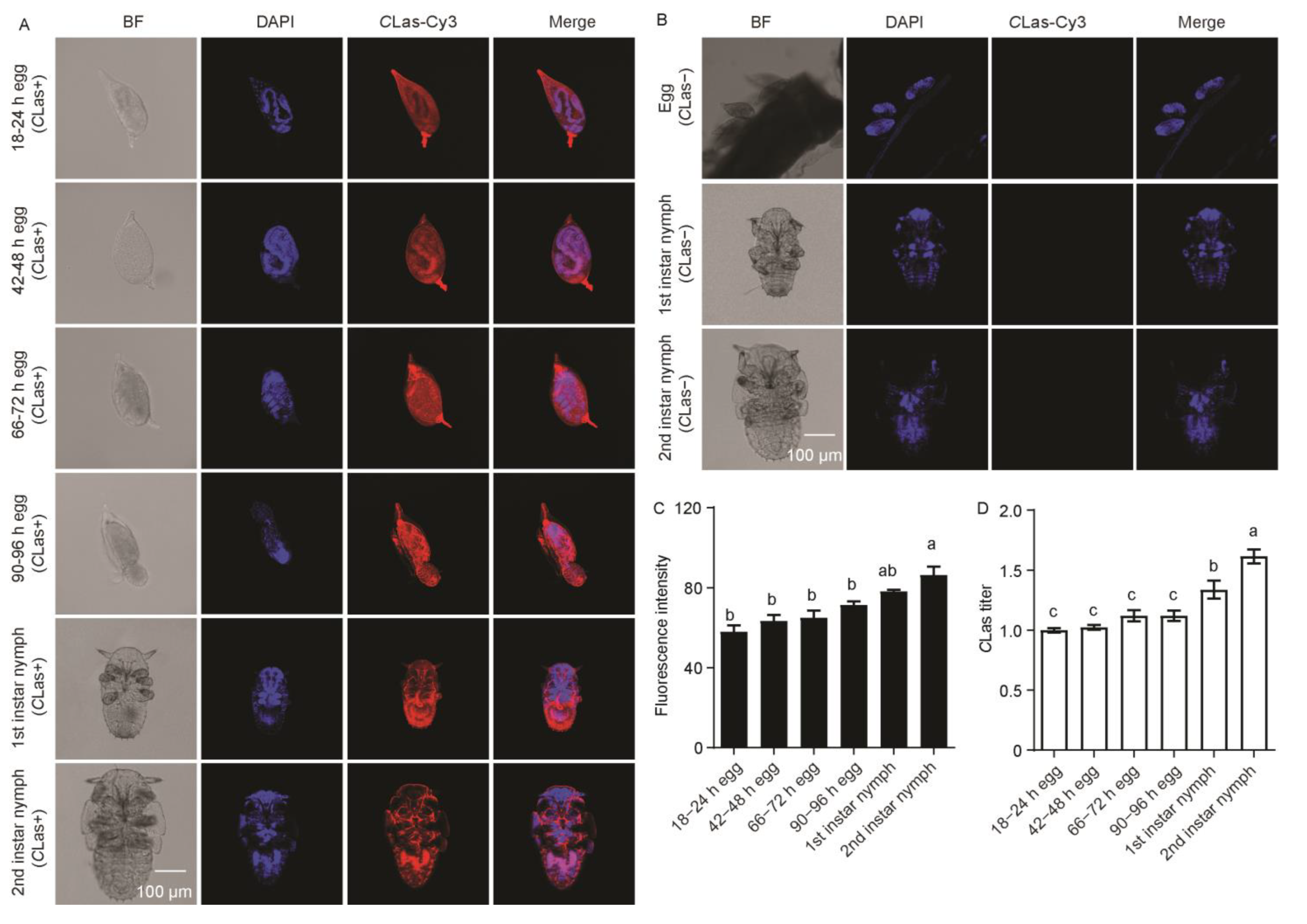
2.3. The Distribution and Dynamics of CLas in Embryos and Nymphs
3. Discussion
4. Materials and Methods
4.1. Host Plants and Insect Colonies
4.2. Tissue Dissection and Sample Collection
4.3. Preparation of Embryos and Subsequent Nymphs
4.4. Fluorescence In-Situ Hybridization (FISH)
4.5. qRT-PCR
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, N. The citrus Huanglongbing crisis and potential solutions. Mol. Plant Pathol. 2019, 12, 607–609. [Google Scholar] [CrossRef]
- Gottwald, T.; Poole, G.; McCollum, T.; Hall, D.; Hartung, J.; Bai, J.; Luo, W.; Posny, D.; Duan, Y.P.; Taylor, E.; et al. Canine olfactory detection of a vectored phytobacterial pathogen, Liberibacter asiaticus, and integration with disease control. Proc. Natl. Acad. Sci. USA 2020, 117, 3492–3501. [Google Scholar] [CrossRef]
- Capoor, S.P.; Rao, D.G.; Viswanath, S.M. Diaphorina citri Kuway. A vector of the greening disease of citrus in India. Indian J. Agric. Sci. 1967, 37, 572–576. [Google Scholar]
- Ammar, E.D.; Ramos, J.E.; Hall, D.G.; Dawson, W.O.; Shatters, R.G., Jr. Acquisition, replication and inoculation of Candidatus Liberibacter asiaticus following various acquisition periods on Huanglongbing-infected citrus by nymphs and adults of the Asian citrus psyllid. PLoS ONE 2016, 11, e0159594. [Google Scholar] [CrossRef]
- Hall, D.G.; Richardson, M.L.; Ammar, E.D.; Halbert, S.E. Asian citrus psyllid, Diaphorina citri, vector of citrus Huanglongbing disease. Entomol. Exp. Appl. 2013, 146, 207–223. [Google Scholar] [CrossRef]
- Inoue, H.; Ohnishi, J.; Ito, T.; Tomimura, K.; Ashihara, W. Enhanced proliferation and efficient transmission of Candidatus Liberibacter asiaticus by adult Diaphorina citri after acquisition feeding in the nymphal stage. Ann. Appl. Biol. 2010, 155, 29–36. [Google Scholar] [CrossRef]
- Ammar, E.D.; Shatters, R.G., Jr.; Lynch, C.; Hall, D.G. Detection and relative titer of Candidatus Liberibacter asiaticus in the salivary glands and alimentary canal of Diaphorina citri (Hemiptera: Psyllidae) vector of citrus huanglongbing disease. Ann. Entomol. Soc. Am. 2011, 104, 526–533. [Google Scholar] [CrossRef]
- Ammar, E.D.; Shatters, R.G., Jr.; Hall, D.G. Localization of Candidatus Liberibacter asiaticus, associated with citrus Huanglongbing disease, in its psyllid vector using fluorescence in situ hybridization. J. Phytopathol. 2011, 159, 726–734. [Google Scholar] [CrossRef]
- Ammar, E.D.; Achor, D.; Levy, A. Immuno-Ultrastructural localization and putative multiplication sites of Huanglongbing bacterium in Asian citrus psyllid Diaphorina citri. Insects 2019, 10, 422. [Google Scholar] [CrossRef]
- Hosseinzadeh, S.; Ramsey, J.; Mann, M.; Bennett, L.; Hunter, W.B.; Shams-Bakhsh, M.; Hall, D.G.; Heck, M. Color morphology of Diaphorina citri influences interactions with its bacterial endosymbionts and ‘Candidatus Liberibacter asiaticus’. PLoS ONE 2019, 14, e0216599. [Google Scholar] [CrossRef]
- Ghanim, M.; Achor, D.; Ghosh, S.; Kontsedalov, S.; Lebedev, G.; Levy, A. ‘Candidatus Liberibacter asiaticus’ accumulates inside endoplasmic reticulum associated vacuoles in the gut cells of Diaphorina citri. Sci. Rep. 2017, 7, 16945. [Google Scholar] [CrossRef]
- Moya, A.; Peretó, J.; Gil, R.; Latorre, A. Learning how to live together: Genomic insights into prokaryote-animal symbioses. Nat. Rev. Genet. 2008, 9, 218–229. [Google Scholar] [CrossRef]
- Lequime, S.; Paul, R.E.; Lambrechts, L. Determinants of arbovirus vertical transmission in mosquitoes. PLoS Pathog. 2016, 12, e1005548. [Google Scholar] [CrossRef]
- Tesh, R.B.; Chaniotis, B.N.; Johnson, K.M. Vesicular stomatitis virus (Indiana serotype): Transovarial transmission by phlebotomine sandlies. Science 1972, 175, 1477–1479. [Google Scholar] [CrossRef]
- Wei, J.; He, Y.Z.; Guo, Q.; Guo, T.; Liu, Y.Q.; Zhou, X.P.; Liu, S.S.; Wang, X.W. Vector development and vitellogenin determine the transovarial transmission of begomoviruses. Proc. Natl. Acad. Sci. USA 2017, 114, 6746–6751. [Google Scholar] [CrossRef]
- Mims, C.A. Vertical transmission of viruses. Microbiol. Rev. 1981, 45, 267–286. [Google Scholar] [CrossRef]
- Pelz-Stelinski, K.S.; Brlansky, R.H.; Ebert, T.A.; Rogers, M.E. Transmission parameters for Candidatus liberibacter asiaticus by Asian citrus psyllid (Hemiptera: Psyllidae). J. Econ. Entomol. 2010, 103, 1531–1541. [Google Scholar] [CrossRef]
- Kelley, A.J.; Pelz-Stelinski, K.S. Maternal contribution of Candidatus Liberibacter asiaticus to Asian citrus psyllid (Hemiptera: Liviidae) nymphs through oviposition site inoculation and transovarial transmission. J. Econ. Entomol. 2019, 112, 2565–2568. [Google Scholar] [CrossRef]
- Mann, M.; Fattah-Hosseini, S.; Ammar, E.D.; Stange, R.; Warrick, E.; Sturgeon, K.; Shatters, R.; Heck, M. Diaphorina citri nymphs are resistant to morphological changes Induced by "Candidatus Liberibacter asiaticus" in midgut epithelial cells. Infect. Immun. 2018, 86, e00889-17. [Google Scholar] [CrossRef]
- Kruse, A.; Fattah-Hosseini, S.; Saha, S.; Johnson, R.; Warwick, E.; Sturgeon, K.; Mueller, L.; MacCoss, M.J.; Shatters, R.G., Jr.; Heck, M.C. Combining ‘omics and microscopy to visualize interactions between the Asian citrus psyllid vector and the Huanglongbing pathogen Candidatus Liberibacter asiaticus in the insect gut. PLoS ONE 2017, 12, e0179531. [Google Scholar] [CrossRef]
- Ukuda-Hosokawa, R.; Sadoyama, Y.; Kishaba, M.; Kuriwada, T.; Anbutsu, H.; Fukatsu, T. Infection density dynamics of the citrus greening bacterium “Candidatus Liberibacter asiaticus” in field populations of the psyllid Diaphorina citri and its relevance to the efficiency of pathogen transmission to citrus plants. Appl. Environ. Microbiol. 2015, 81, 3728–3736. [Google Scholar] [CrossRef]
- Ammar, E.D.; Hall, D.G.; Shatters, R.G., Jr. Ultrastructure of the salivary glands, alimentary canal and bacteria-like organisms in the Asian citrus psyllid, vector of citrus huanglongbing disease bacteria. J. Microsc. Ultrastruct. 2017, 5, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Cicero, J.M.; Brown, J.K.; Roberts, P.D.; Stansly, P.A. The digestive system of Diaphorina citri and Bactericera cockerelli (hemiptera: Psyllidae). Ann. Entomol. Soc. Am. 2009, 102, 650–665. [Google Scholar] [CrossRef]
- Mann, R.S.; Pelz-Stelinski, K.; Hermann, S.L.; Tiwari, S.; Stelinski, L.L. Sexual transmission of a plant pathogenic bacterium, Candidatus Liberibacter asiaticus, between conspecific insect vectors during mating. PLoS ONE. 2011, 6, e29197. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzadeh, S.; Shams-Bakhsh, M.; Mann, M.; Fattah-Hosseini, S.; Bagheri, A.; Mehrabadi, M.; Heck, M. Distribution and variation of bacterial endosymbiont and “Candidatus Liberibacter asiaticus” titer in the Huanglongbing insect vector, Diaphorina citri Kuwayama. Microb. Ecol. 2019, 78, 206–222. [Google Scholar] [CrossRef]
- Nachappa, P.; Levy, J.; Pierson, E.; Tamborindeguy, C. Diversity of endosymbionts in the potato psyllid, Bactericera cockerelli (Triozidae), vector of zebra chip disease of potato. Curr. Microbiol. 2011, 62, 1510–1520. [Google Scholar] [CrossRef]
- Pedro, S.A.; Abelman, S.; Tonnang, H.E. Predicting rift valley fever inter-epidemic activities and outbreak patterns: Insights from a stochastic host-vector model. PLoS Negl. Trop. Dis. 2016, 10, e0005167. [Google Scholar] [CrossRef]
- Huo, Y.; Liu, W.W.; Zhang, F.J.; Chen, X.Y.; Li, L.; Liu, Q.F.; Zhou, Y.J.; Wei, T.Y.; Fang, R.X.; Wang, X.F. Transovarial transmission of a plant virus is mediated by vitellogenin of its insect vector. PLoS Pathog. 2014, 10, e1003949. [Google Scholar] [CrossRef]
- Ammar, E.D.; Tsai, C.W.; Whitfield, A.E.; Redinbaugh, M.G.; Hogenhout, S.A. Cellular and molecular aspects of rhabdovirus interactions with insect and plant hosts. Annu. Rev. Entomol. 2009, 54, 447–468. [Google Scholar] [CrossRef]
- Falk, B.W.; Tsai, J.H. Biology and molecular biology of viruses in the genus tenuivirus. Annu. Rev. Phytopathol. 1998, 36, 139–163. [Google Scholar] [CrossRef]
- Alves, M.N.; Cifuentes-Arenas, J.C.; Raiol-Junior, L.L.; Ferro, J.A.; Peña, L. Early population dynamics of “Candidatus Liberibacter asiaticus” in susceptible and resistant genotypes after inoculation with infected Diaphorina citri feeding on young shoots. Front. Microbiol. 2021, 12, 683923. [Google Scholar] [CrossRef] [PubMed]
- Ammar, E.D.; Shatters, R.G.; Heck, M. Huanglongbing pathogens: Acquisition, transmission and vector interactions. In Asian Citrus Psyllid: Biology, Ecology and Management of the Huanglongbing Vector; Qureshi, J.A., Stansly, P.A., Eds.; CAB International: Wallingford, CT, USA, 2020; pp. 113–139. [Google Scholar]
- Da Graça, J.V.; Douhan, G.W.; Halbert, S.E.; Keremane, M.L.; Lee, R.F.; Vidalakis, G.; Zhao, H. Huanglongbing: An overview of a complex pathosystem ravaging the world’s citrus. J. Integr. Plant Biol. 2016, 58, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Hartung, J.S.; Levy, L. Quantitative real-time PCR for detection and identiÞcation of Candidatus Liberibacter species associated with citrus Huanglongbing. J. Microbiol. Methods 2006, 66, 104–105. [Google Scholar] [CrossRef] [PubMed]
- Dan, H.; Ikeda, N.; Fujikami, M.; Nakabachi, A. Behavior of bacteriome symbionts during transovarial transmission and development of the Asian citrus psyllid. PLoS ONE 2017, 12, e0189779. [Google Scholar] [CrossRef] [PubMed]
- Grafton-Cardwell, E.E.; Stelinski, L.L.; Stansly, P.A. Biology and management of Asian citrus psyllid, vector of the huanglongbing pathogens. Annu. Rev. Entomol. 2013, 58, 413–432. [Google Scholar] [CrossRef]
- Kliot, A.; Kontsedalov, S.; Lebedev, G.; Brumin, M.; Cathrin, P.B.; Marubayashi, J.M. Fluorescence in situ hybridizations (FISH) for the localization of viruses and endosymbiotic bacteria in plant and insect tissues. J. Vis. Exp. 2014, 84, e51030. [Google Scholar]
- Van Bogaert, N.; De Jonghe, K.; Van Damme, E.J.; Maes, M.; Smagghe, G. Quantitation and localization of pospiviroids in aphids. J. Virol. Methods. 2015, 211, 51–54. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nian, X.; Wu, S.; Li, J.; Luo, Y.; He, J.; Tan, S.; Wang, D.; Cen, Y.; He, Y. Spatial Distribution and Temporal Dynamics of Candidatus Liberibacter Asiaticus in Different Stages of Embryos, Nymphs and Adults of Diaphorina citri. Int. J. Mol. Sci. 2023, 24, 8997. https://doi.org/10.3390/ijms24108997
Nian X, Wu S, Li J, Luo Y, He J, Tan S, Wang D, Cen Y, He Y. Spatial Distribution and Temporal Dynamics of Candidatus Liberibacter Asiaticus in Different Stages of Embryos, Nymphs and Adults of Diaphorina citri. International Journal of Molecular Sciences. 2023; 24(10):8997. https://doi.org/10.3390/ijms24108997
Chicago/Turabian StyleNian, Xiaoge, Shujie Wu, Jiayun Li, Yaru Luo, Jielan He, Shijian Tan, Desen Wang, Yijing Cen, and Yurong He. 2023. "Spatial Distribution and Temporal Dynamics of Candidatus Liberibacter Asiaticus in Different Stages of Embryos, Nymphs and Adults of Diaphorina citri" International Journal of Molecular Sciences 24, no. 10: 8997. https://doi.org/10.3390/ijms24108997
APA StyleNian, X., Wu, S., Li, J., Luo, Y., He, J., Tan, S., Wang, D., Cen, Y., & He, Y. (2023). Spatial Distribution and Temporal Dynamics of Candidatus Liberibacter Asiaticus in Different Stages of Embryos, Nymphs and Adults of Diaphorina citri. International Journal of Molecular Sciences, 24(10), 8997. https://doi.org/10.3390/ijms24108997