
Investigation of Plasma-Derived Lipidome Profiles in Experimental Cerebral Malaria in a Mouse Model Study

 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Results
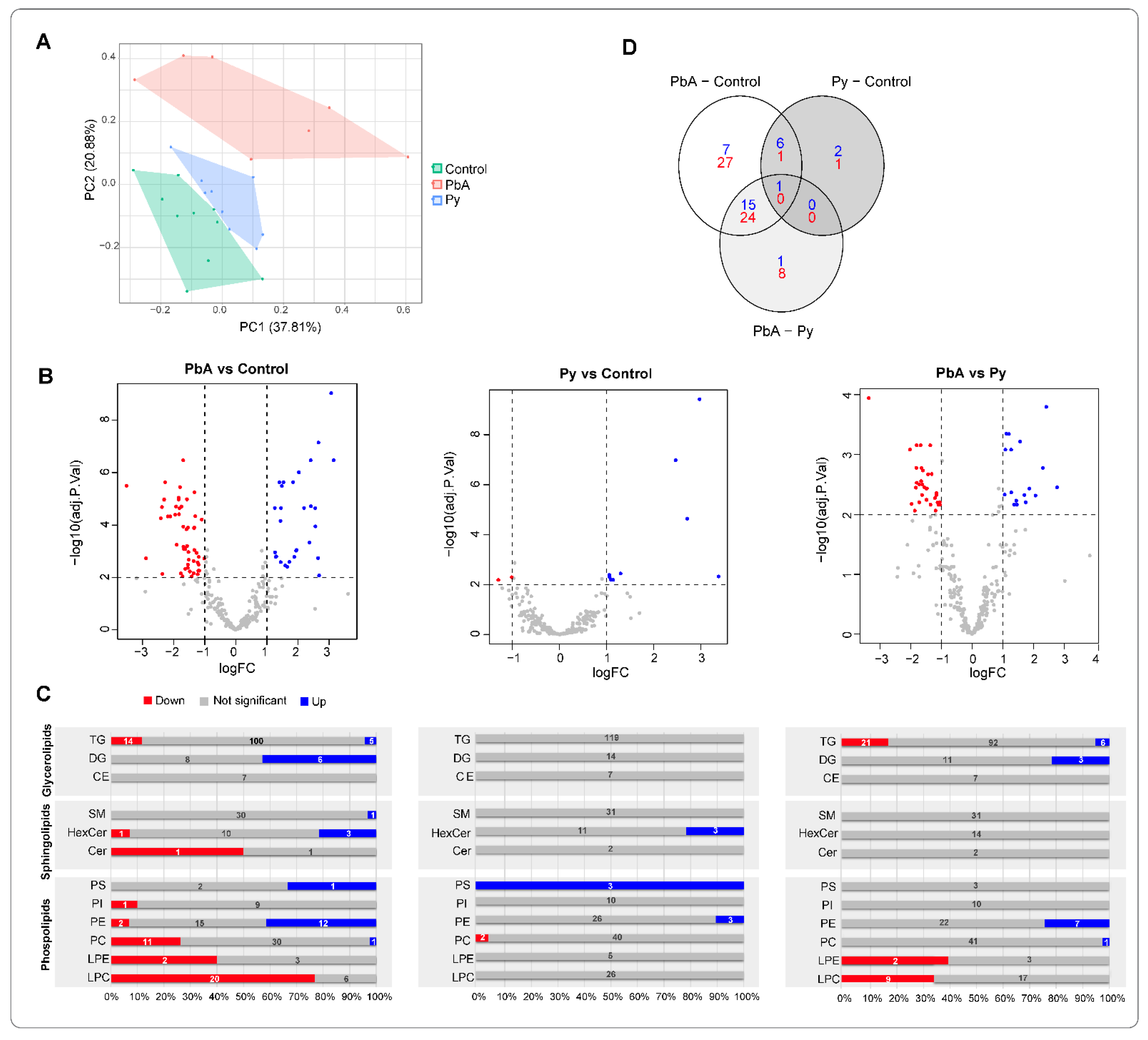
2.1. Qualitative and Quantitative Changes in EV/MV Produced in CM versus NCM
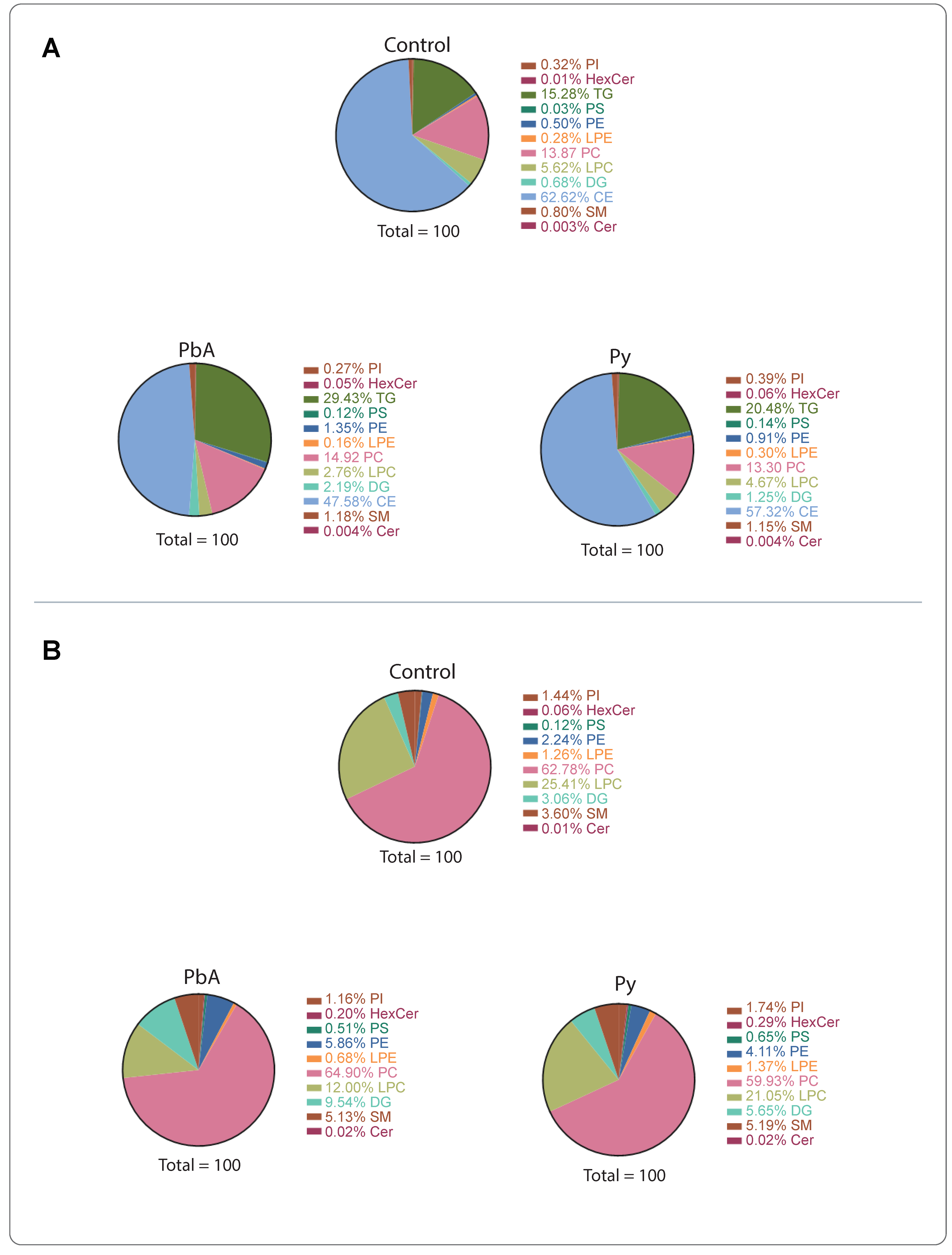
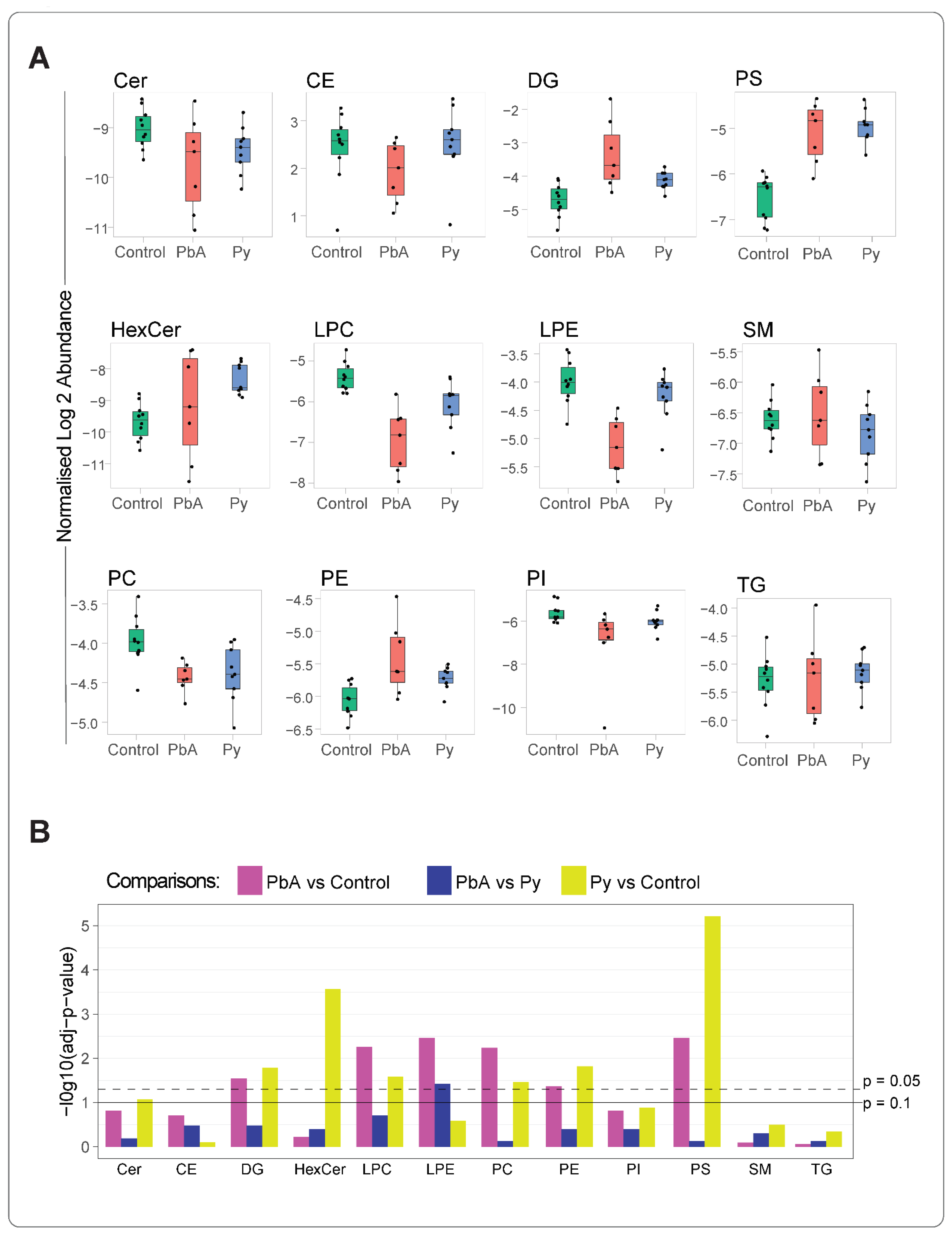
2.2. CM Caused Significant Alterations in MV-Derived Lipid Class Composition
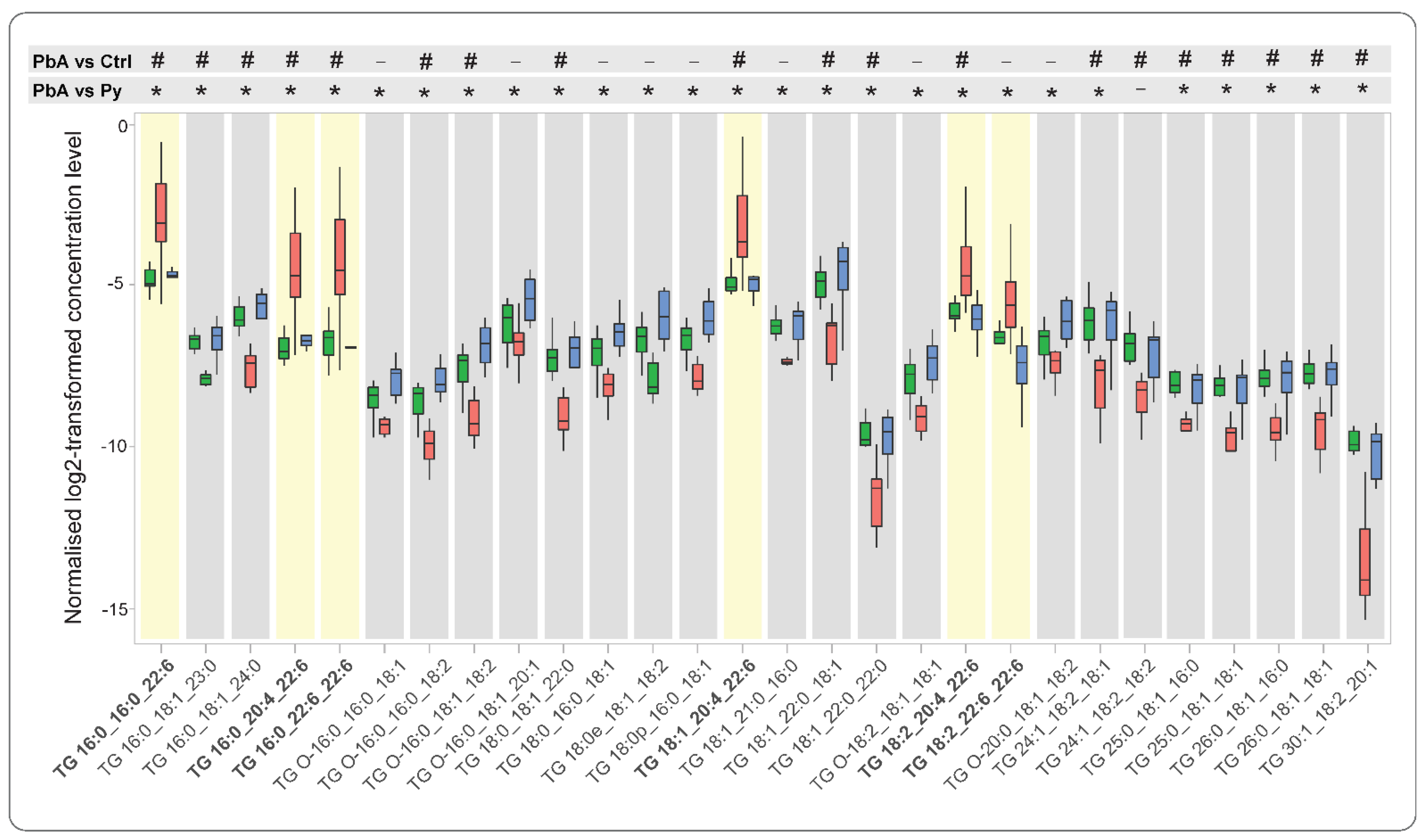
2.3. Quantitative Analysis of Lipid Species among Measured Lipid Classes
3. Discussion
4. Materials and Methods
4.1. Mice and Parasite Inoculation
4.2. Blood Sampling and MV Preparation
4.3. Lipid Extraction
4.4. Liquid Chromatography–Mass Spectrometry
4.5. Nomenclature
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Panda, C.; Mahapatra, R.K. An update on cerebral malaria for therapeutic intervention. Mol. Biol. Rep. 2022, 49, 10579–10591. [Google Scholar] [CrossRef] [PubMed]
- Bartoloni, A.; Zammarchi, L. Clinical aspects of uncomplicated and severe malaria. Mediterr. J. Hematol. Infect. Dis. 2012, 4, e2012026. [Google Scholar] [CrossRef] [PubMed]
- Luzolo, A.L.; Ngoyi, D.M. Cerebral malaria. Brain Res. Bull. 2019, 145, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Idro, R.; Jenkins, N.E.; Newton, C.R. Pathogenesis, clinical features, and neurological outcome of cerebral malaria. Lancet Neurol. 2005, 4, 827–840. [Google Scholar] [CrossRef] [PubMed]
- Idro, R.; Kakooza-Mwesige, A.; Balyejjussa, S.; Mirembe, G.; Mugasha, C.; Tugumisirize, J.; Byarugaba, J. Severe neurological sequelae and behaviour problems after cerebral malaria in Ugandan children. BMC Res. Notes 2010, 3, 104. [Google Scholar] [CrossRef]
- Idro, R.; Ndiritu, M.; Ogutu, B.; Mithwani, S.; Maitland, K.; Berkley, J.; Crawley, J.; Fegan, G.; Bauni, E.; Peshu, N.; et al. Burden, features, and outcome of neurological involvement in acute falciparum malaria in Kenyan children. JAMA 2007, 297, 2232–2240. [Google Scholar] [CrossRef]
- Hunt, N.H.; Grau, G.E. Cytokines: Accelerators and brakes in the pathogenesis of cerebral malaria. Trends Immunol. 2003, 24, 491–499. [Google Scholar] [CrossRef]
- Schofield, L.; Grau, G.E. Immunological processes in malaria pathogenesis. Nat. Rev. Immunol. 2005, 5, 722–735. [Google Scholar] [CrossRef]
- Hunt, N.H.; Golenser, J.; Chan-Ling, T.; Parekh, S.; Rae, C.; Potter, S.; Medana, I.M.; Miu, J.; Ball, H.J. Immunopathogenesis of cerebral malaria. Int. J. Parasitol. 2006, 36, 569–582. [Google Scholar] [CrossRef]
- van der Heyde, H.C.; Nolan, J.; Combes, V.; Gramaglia, I.; Grau, G.E. A unified hypothesis for the genesis of cerebral malaria: Sequestration, inflammation and hemostasis leading to microcirculatory dysfunction. Trends Parasitol. 2006, 22, 503–508. [Google Scholar] [CrossRef]
- Amante, F.H.; Haque, A.; Stanley, A.C.; Rivera Fde, L.; Randall, L.M.; Wilson, Y.A.; Yeo, G.; Pieper, C.; Crabb, B.S.; de Koning-Ward, T.F.; et al. Immune-mediated mechanisms of parasite tissue sequestration during experimental cerebral malaria. J. Immunol. 2010, 185, 3632–3642. [Google Scholar] [CrossRef] [PubMed]
- El-Assaad, F.; Wheway, J.; Hunt, N.H.; Grau, G.E.; Combes, V. Production, fate and pathogenicity of plasma microparticles in murine cerebral malaria. PLoS Pathog. 2014, 10, e1003839. [Google Scholar] [CrossRef] [PubMed]
- White, N.J.; Turner, G.D.; Medana, I.M.; Dondorp, A.M.; Day, N.P. The murine cerebral malaria phenomenon. Trends Parasitol. 2010, 26, 11–15. [Google Scholar] [CrossRef]
- Langhorne, J.; Buffet, P.; Galinski, M.; Good, M.; Harty, J.; Leroy, D.; Mota, M.M.; Pasini, E.; Renia, L.; Riley, E.; et al. The relevance of non-human primate and rodent malaria models for humans. Malar. J. 2011, 10, 23. [Google Scholar] [CrossRef] [PubMed]
- Riley, E.M.; Couper, K.N.; Helmby, H.; Hafalla, J.C.; de Souza, J.B.; Langhorne, J.; Jarra, W.B.; Zavala, F. Neuropathogenesis of human and murine malaria. Trends Parasitol. 2010, 26, 277–278. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, L.J. Murine cerebral malaria: How far from human cerebral malaria? Trends Parasitol. 2010, 26, 271–272. [Google Scholar] [CrossRef] [PubMed]
- Renia, L.; Gruner, A.C.; Snounou, G. Cerebral malaria: In praise of epistemes. Trends Parasitol. 2010, 26, 275–277. [Google Scholar] [CrossRef]
- Morrell, C.N.; Aggrey, A.A.; Chapman, L.M.; Modjeski, K.L. Emerging roles for platelets as immune and inflammatory cells. Blood 2014, 123, 2759–2767. [Google Scholar] [CrossRef]
- Nacer, A.; Movila, A.; Sohet, F.; Girgis, N.M.; Gundra, U.M.; Loke, P.; Daneman, R.; Frevert, U. Experimental cerebral malaria pathogenesis--hemodynamics at the blood brain barrier. PLoS Pathog. 2014, 10, e1004528. [Google Scholar] [CrossRef]
- Grau, G.E.; Fajardo, L.F.; Piguet, P.F.; Allet, B.; Lambert, P.H.; Vassalli, P. Tumor necrosis factor (cachectin) as an essential mediator in murine cerebral malaria. Science 1987, 237, 1210–1212. [Google Scholar] [CrossRef]
- Ghazanfari, N.; Mueller, S.N.; Heath, W.R. Cerebral Malaria in Mouse and Man. Front. Immunol. 2018, 9, 2016. [Google Scholar] [CrossRef] [PubMed]
- Cottrell, B.J.; Playfair, J.H.; De Souza, B.J. Cell-mediated immunity in mice vaccinated against malaria. Clin. Exp. Immunol. 1978, 34, 147–158. [Google Scholar] [PubMed]
- Skotland, T.; Hessvik, N.P.; Sandvig, K.; Llorente, A. Exosomal lipid composition and the role of ether lipids and phosphoinositides in exosome biology. J. Lipid Res. 2019, 60, 9–18. [Google Scholar] [CrossRef]
- Casares, D.; Escriba, P.V.; Rossello, C.A. Membrane Lipid Composition: Effect on Membrane and Organelle Structure, Function and Compartmentalization and Therapeutic Avenues. Int. J. Mol. Sci. 2019, 20, 2167. [Google Scholar] [CrossRef] [PubMed]
- Lazar, I.; Clement, E.; Attane, C.; Muller, C.; Nieto, L. A new role for extracellular vesicles: How small vesicles can feed tumors’ big appetite. J. Lipid Res. 2018, 59, 1793–1804. [Google Scholar] [CrossRef]
- Brancucci, N.M.B.; Gerdt, J.P.; Wang, C.; De Niz, M.; Philip, N.; Adapa, S.R.; Zhang, M.; Hitz, E.; Niederwieser, I.; Boltryk, S.D.; et al. Lysophosphatidylcholine Regulates Sexual Stage Differentiation in the Human Malaria Parasite Plasmodium falciparum. Cell 2017, 171, 1532–1544.e15. [Google Scholar] [CrossRef]
- Silva, A.R.; Moraes, B.P.T.; Goncalves-de-Albuquerque, C.F. Mediterranean Diet: Lipids, Inflammation, and Malaria Infection. Int. J. Mol. Sci. 2020, 21, 4489. [Google Scholar] [CrossRef]
- Colvin, H.N.; Joice Cordy, R. Insights into malaria pathogenesis gained from host metabolomics. PLoS Pathog. 2020, 16, e1008930. [Google Scholar] [CrossRef]
- Visser, B.J.; Wieten, R.W.; Nagel, I.M.; Grobusch, M.P. Serum lipids and lipoproteins in malaria—A systematic review and meta-analysis. Malar. J. 2013, 12, 442. [Google Scholar] [CrossRef]
- Combes, V.; El-Assaad, F.; Faille, D.; Jambou, R.; Hunt, N.H.; Grau, G.E. Microvesiculation and cell interactions at the brain-endothelial interface in cerebral malaria pathogenesis. Prog. Neurobiol. 2010, 91, 140–151. [Google Scholar] [CrossRef]
- Debs, S.; Cohen, A.; Hosseini-Beheshti, E.; Chimini, G.; Hunt, N.H.; Grau, G.E.R. Interplay of extracellular vesicles and other players in cerebral malaria pathogenesis. Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Stahl, P.D. Extracellular vesicles: A new communication paradigm? Nat. Rev. Mol. Cell. Biol. 2019, 20, 509–510. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, L.; Bali, J.; Barr, M.M.; Court, F.A.; Kramer-Albers, E.M.; Picou, F.; Raposo, G.; van der Vos, K.E.; van Niel, G.; Wang, J.; et al. Emerging roles of extracellular vesicles in the nervous system. J. Neurosci. 2014, 34, 15482–15489. [Google Scholar] [CrossRef]
- Hosseini-Beheshti, E.; Grau, G.E.R. Extracellular vesicles as mediators of immunopathology in infectious diseases. Immunol. Cell Biol. 2018, 96, 694–703. [Google Scholar] [CrossRef] [PubMed]
- Grau, G.E.R.; Hosseini-Beheshti, E. Extracellular Vesicles and Cerebral Malaria. Subcell Biochem. 2021, 97, 501–508. [Google Scholar] [CrossRef]
- Coumans, F.A.W.; Brisson, A.R.; Buzas, E.I.; Dignat-George, F.; Drees, E.E.E.; El-Andaloussi, S.; Emanueli, C.; Gasecka, A.; Hendrix, A.; Hill, A.F.; et al. Methodological Guidelines to Study Extracellular Vesicles. Circ. Res. 2017, 120, 1632–1648. [Google Scholar] [CrossRef]
- Dong, L.; Zieren, R.C.; Horie, K.; Kim, C.J.; Mallick, E.; Jing, Y.; Feng, M.; Kuczler, M.D.; Green, J.; Amend, S.R.; et al. Comprehensive evaluation of methods for small extracellular vesicles separation from human plasma, urine and cell culture medium. J. Extracell. Vesicles 2020, 10, e12044. [Google Scholar] [CrossRef]
- Akers, J.C.; Gonda, D.; Kim, R.; Carter, B.S.; Chen, C.C. Biogenesis of extracellular vesicles (EV): Exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J. Neurooncol. 2013, 113, 1–11. [Google Scholar] [CrossRef]
- Grau, G.E.; Tacchini-Cottier, F.; Vesin, C.; Milon, G.; Lou, J.N.; Piguet, P.F.; Juillard, P. TNF-induced microvascular pathology: Active role for platelets and importance of the LFA-1/ICAM-1 interaction. Eur. Cytokine Netw. 1993, 4, 415–419. [Google Scholar]
- Orikiiriza, J.; Surowiec, I.; Lindquist, E.; Bonde, M.; Magambo, J.; Muhinda, C.; Bergstrom, S.; Trygg, J.; Normark, J. Lipid response patterns in acute phase paediatric Plasmodium falciparum malaria. Metabolomics 2017, 13, 41. [Google Scholar] [CrossRef]
- Cordy, R.J.; Patrapuvich, R.; Lili, L.N.; Cabrera-Mora, M.; Chien, J.T.; Tharp, G.K.; Khadka, M.; Meyer, E.V.; Lapp, S.A.; Joyner, C.J.; et al. Distinct amino acid and lipid perturbations characterize acute versus chronic malaria. JCI Insight 2019, 4, e125156. [Google Scholar] [CrossRef] [PubMed]
- Combes, V.; Taylor, T.E.; Juhan-Vague, I.; Mege, J.L.; Mwenechanya, J.; Tembo, M.; Grau, G.E.; Molyneux, M.E. Circulating endothelial microparticles in malawian children with severe falciparum malaria complicated with coma. JAMA 2004, 291, 2542–2544. [Google Scholar] [CrossRef] [PubMed]
- Nantakomol, D.; Dondorp, A.M.; Krudsood, S.; Udomsangpetch, R.; Pattanapanyasat, K.; Combes, V.; Grau, G.E.; White, N.J.; Viriyavejakul, P.; Day, N.P.; et al. Circulating red cell-derived microparticles in human malaria. J. Infect. Dis. 2011, 203, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.; Zinger, A.; Tiberti, N.; Grau, G.E.R.; Combes, V. Differential plasma microvesicle and brain profiles of microRNA in experimental cerebral malaria. Malar. J. 2018, 17, 192. [Google Scholar] [CrossRef]
- Tiberti, N.; Latham, S.L.; Bush, S.; Cohen, A.; Opoka, R.O.; John, C.C.; Juillard, A.; Grau, G.E.; Combes, V. Exploring experimental cerebral malaria pathogenesis through the characterisation of host-derived plasma microparticle protein content. Sci. Rep. 2016, 6, 37871. [Google Scholar] [CrossRef]
- Zoia, M.; Yesodha Subramanian, B.; Eriksson, K.K.; Ravi, M.S.; Yaghmaei, S.; Fellay, I.; Scolari, B.; Walch, M.; Mantel, P.Y. Validation of Effective Extracellular Vesicles Isolation Methods Adapted to Field Studies in Malaria Endemic Regions. Front. Cell Dev. Biol. 2022, 10, 812244. [Google Scholar] [CrossRef]
- Gulati, S.; Ekland, E.H.; Ruggles, K.V.; Chan, R.B.; Jayabalasingham, B.; Zhou, B.; Mantel, P.Y.; Lee, M.C.; Spottiswoode, N.; Coburn-Flynn, O.; et al. Profiling the Essential Nature of Lipid Metabolism in Asexual Blood and Gametocyte Stages of Plasmodium falciparum. Cell Host Microbe 2015, 18, 371–381. [Google Scholar] [CrossRef]
- Martins, Y.C.; Smith, M.J.; Pelajo-Machado, M.; Werneck, G.L.; Lenzi, H.L.; Daniel-Ribeiro, C.T.; Carvalho, L.J. Characterization of cerebral malaria in the outbred Swiss Webster mouse infected by Plasmodium berghei ANKA. Int. J. Exp. Pathol. 2009, 90, 119–130. [Google Scholar] [CrossRef]
- Reis, P.A.; Comim, C.M.; Hermani, F.; Silva, B.; Barichello, T.; Portella, A.C.; Gomes, F.C.; Sab, I.M.; Frutuoso, V.S.; Oliveira, M.F.; et al. Cognitive dysfunction is sustained after rescue therapy in experimental cerebral malaria, and is reduced by additive antioxidant therapy. PLoS Pathog. 2010, 6, e1000963. [Google Scholar] [CrossRef]
- Pappa, V.; Seydel, K.; Gupta, S.; Feintuch, C.M.; Potchen, M.J.; Kampondeni, S.; Goldman-Yassen, A.; Veenstra, M.; Lopez, L.; Kim, R.S.; et al. Lipid metabolites of the phospholipase A2 pathway and inflammatory cytokines are associated with brain volume in paediatric cerebral malaria. Malar. J. 2015, 14, 513. [Google Scholar] [CrossRef]
- Hunt, N.H.; Too, L.K.; Khaw, L.T.; Guo, J.; Hee, L.; Mitchell, A.J.; Grau, G.E.; Ball, H.J. The kynurenine pathway and parasitic infections that affect CNS function. Neuropharmacology 2017, 112, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Hunt, N.H.; Ball, H.J.; Hansen, A.M.; Khaw, L.T.; Guo, J.; Bakmiwewa, S.; Mitchell, A.J.; Combes, V.; Grau, G.E. Cerebral malaria: Gamma-interferon redux. Front. Cell. Infect. Microbiol. 2014, 4, 113. [Google Scholar] [CrossRef] [PubMed]
- Sierro, F.; Grau, G.E.R. The ins and outs of cerebral malaria pathogenesis: Immunopathology, extracellular vesicles, immunometabolism, and trained immunity. Front. Immunol. 2019, 10, 830. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Zhu, W.; Chen, C.; Yan, B.; Zhu, L.; Chen, X.; Peng, C. The mechanisms of lysophosphatidylcholine in the development of diseases. Life Sci. 2020, 247, 117443. [Google Scholar] [CrossRef] [PubMed]
- Kabarowski, J.H. G2A and LPC: Regulatory functions in immunity. Prostaglandins Other Lipid Mediat. 2009, 89, 73–81. [Google Scholar] [CrossRef]
- Mantel, P.Y.; Hoang, A.N.; Goldowitz, I.; Potashnikova, D.; Hamza, B.; Vorobjev, I.; Ghiran, I.; Toner, M.; Irimia, D.; Ivanov, A.R.; et al. Malaria-Infected Erythrocyte-Derived Microvesicles Mediate Cellular Communication within the Parasite Population and with the Host Immune System. Cell Host Microbe 2013, 13, 521–534. [Google Scholar] [CrossRef]
- Scaccabarozzi, D.; Deroost, K.; Corbett, Y.; Lays, N.; Corsetto, P.; Sale, F.O.; Van den Steen, P.E.; Taramelli, D. Differential induction of malaria liver pathology in mice infected with Plasmodium chabaudi AS or Plasmodium berghei NK65. Malar. J. 2018, 17, 18. [Google Scholar] [CrossRef]
- Simopoulos, A.P. An Increase in the Omega-6/Omega-3 Fatty Acid Ratio Increases the Risk for Obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef]
- Fenton, J.I.; Hord, N.G.; Ghosh, S.; Gurzell, E.A. Immunomodulation by dietary long chain omega-3 fatty acids and the potential for adverse health outcomes. Prostaglandins Leukot. Essent. Fat. Acids 2013, 89, 379–390. [Google Scholar] [CrossRef]
- Skotland, T.; Sagini, K.; Sandvig, K.; Llorente, A. An emerging focus on lipids in extracellular vesicles. Adv. Drug Deliv. Rev. 2020, 159, 308–321. [Google Scholar] [CrossRef]
- Record, M.; Silvente-Poirot, S.; Poirot, M.; Wakelam, M.J.O. Extracellular vesicles: Lipids as key components of their biogenesis and functions. J. Lipid Res. 2018, 59, 1316–1324. [Google Scholar] [CrossRef] [PubMed]
- Vincke, L.H.; Bafort, J. Results of 2 years of observation of the cyclical transmission of Plasmodium berghei. Ann. Soc. Belges Med. Trop. Parasitol. Mycol. 1968, 48, 439–454. [Google Scholar] [PubMed]
- Playfair, J.H.; De Souza, J.B.; Cottrell, B.J. Protection of mice against malaria by a killed vaccine: Differences in effectiveness against P. yoelii and P. berghei. Immunology 1977, 33, 507–515. [Google Scholar]
- Grau, G.E.; Piguet, P.F.; Engers, H.D.; Louis, J.A.; Vassalli, P.; Lambert, P.H. L3T4+ T lymphocytes play a major role in the pathogenesis of murine cerebral malaria. J. Immunol. 1986, 137, 2348–2354. [Google Scholar] [PubMed]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef]
- Liebisch, G.; Fahy, E.; Aoki, J.; Dennis, E.A.; Durand, T.; Ejsing, C.S.; Fedorova, M.; Feussner, I.; Griffiths, W.J.; Kofeler, H.; et al. Update on LIPID MAPS classification, nomenclature, and shorthand notation for MS-derived lipid structures. J. Lipid Res. 2020, 61, 1539–1555. [Google Scholar] [CrossRef]
- Phipson, B.; Lee, S.; Majewski, I.J.; Alexander, W.S.; Smyth, G.K. Robust Hyperparameter Estimation Protects against Hypervariable Genes and Improves Power to Detect Differential Expression. Ann. Appl. Stat. 2016, 10, 946–963. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate—A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Sheldon, R.M. Copyright. In Introductory Statistics, 4th ed.; Ross, S.M., Ed.; Academic Press: Oxford, UK, 2017; p. iv. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
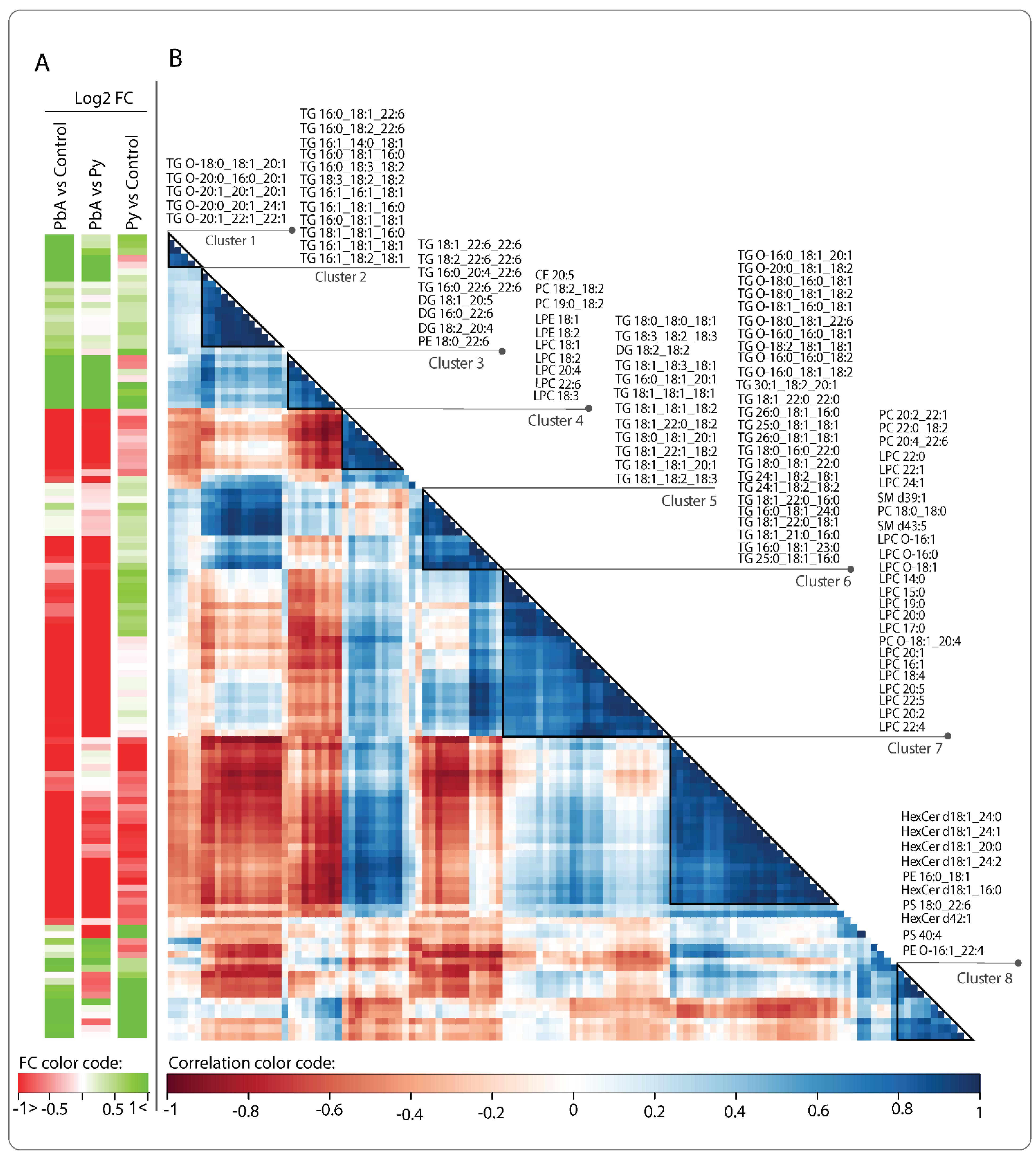{kind=link}
| Lipid Ion | Log FC | Adjusted p-Value | Lipid Ion | Log FC | Adjusted p-Value |
|---|---|---|---|---|---|
| Glycerolipids | Phospholipids | ||||
| DG 16:0_22:6 | 3.15 | 3.45 × 10−7 | LPC 18:1 | −1.58 | 0.000144 |
| DG 18:1_20:4 | 2.19 | 2.22 × 10−5 | LPC 18:2 | −2.11 | 4.64 × 10−5 |
| DG 18:1_20:5 | 2.37 | 0.000461 | LPC 18:4 | −2.28 | 2.35 × 10−6 |
| DG 18:1_22:6 | 1.72 | 0.002531 | LPC 19:0 | −1.75 | 1.94 × 10−5 |
| DG 18:2_20:4 | 2.41 | 1.90 × 10−5 | LPC 20:0 | −1.81 | 2.24 × 10−5 |
| DG 18:2_22:6 | 1.95 | 0.000892 | LPC 20:1 | −1.84 | 9.10 × 10−6 |
| TG 16:0_16:0_22:6 | 1.94 | 0.000949 | LPC 20:2 | −2.3 | 1.06 × 10−5 |
| TG 16:0_18:1_23:0 | −1.19 | 0.001628 | LPC 20:4 | −1.54 | 0.000123 |
| TG 16:0_18:1_24:0 | −1.54 | 0.003227 | LPC 20:5 | −2.37 | 2.04 × 10−5 |
| TG 16:0_20:4_22:6 | 2.56 | 0.000112 | LPC 22:1 | −1.23 | 0.007528 |
| TG 16:0_22:6_22:6 | 2.64 | 0.001803 | LPC 22:4 | −2.41 | 5.44 × 10−5 |
| TG O-16:0_16:0_18:2 | −1.2 | 0.005488 | LPC 22:5 | −2.18 | 4.64 × 10−5 |
| TG O-16:0_18:1_18:2 | −1.36 | 0.006879 | LPC 22:6 | −1.33 | 0.000892 |
| TG 18:0_18:1_22:0 | −1.62 | 0.004668 | LPE 18:1 | −1.79 | 3.58 × 10−6 |
| TG 18:1_20:4_22:6 | 1.88 | 0.001628 | LPE 18:2 | −1.92 | 1.94 × 10−5 |
| TG 18:1_22:0_18:1 | −1.71 | 0.009094 | PC 15:0_18:2 | −1.33 | 5.73 × 10−6 |
| TG 18:1_22:0_22:0 | −1.9 | 0.000559 | PC 16:0_20:5 | −1.01 | 0.00189 |
| TG 18:2_20:4_22:6 | 1.59 | 0.003492 | PC O-16:0_20:4 | −1.34 | 0.00013 |
| TG 24:1_18:2_18:1 | −1.77 | 0.006916 | PC O-16:0_22:4 | −1.23 | 0.002657 |
| TG 24:1_18:2_18:2 | −1.48 | 0.005911 | PC 16:1_20:4 | −1.3 | 4.54 × 10−5 |
| TG 25:0_18:1_16:0 | −1.26 | 0.001152 | PC 17:0_18:2 | −1.1 | 6.07 × 10−5 |
| TG 25:0_18:1_18:1 | −1.63 | 0.000827 | PC 18:0_22:6 | 1.28 | 0.001628 |
| TG 26:0_18:1_16:0 | −1.52 | 0.001061 | PC O-18:1_20:4 | −1.57 | 1.04 × 10−5 |
| TG 26:0_18:1_18:1 | −1.67 | 0.000815 | PC 18:2_18:2 | −1.95 | 3.99 × 10−5 |
| TG 30:1_18:2_20:1 | −3.51 | 3.19 × 10−6 | PC 19:0_18:2 | −1.69 | 3.45 × 10−7 |
| Sphingolipids | PC 20:4_22:6 | −1.17 | 0.0055 | ||
| SM 36:0;O2 | 1.65 | 0.003903 | PC 22:0_18:2 | −1.57 | 0.000624 |
| HexCer 18:1;O2/16:0 | 3.06 | 9.36 × 10−10 | PE 16:0_18:1 | 2.03 | 9.88 × 10−7 |
| HexCer 41:1;O2 | −2.36 | 0.007303 | PE 16:0_18:2 | 1.4 | 2.35 × 10−6 |
| HexCer 42:1;O2 | 2.67 | 0.00836 | PE 16:0_20:4 | 1.53 | 2.35 × 10−6 |
| PE 16:0_20:5 | 1.44 | 0.002587 | |||
| Phospholipids | PE 18:0_20:5 | 1.44 | 6.91 × 10−5 | ||
| LPC 14:0 | −1.69 | 0.000114 | PE 18:0_22:6 | 2.66 | 7.25 × 10−8 |
| LPC 15:0 | −1.83 | 3.83 × 10−5 | PE O-18:1_18:2 | −1.36 | 0.002351 |
| LPC O-16:0 | −1.19 | 0.003254 | PE 18:1_18:1 | 1.3 | 0.001572 |
| LPC O-16:1 | −1.54 | 0.004291 | PE 18:1_20:4 | 1.25 | 2.22 × 10−5 |
| LPC 16:1 | −1.84 | 1.04 × 10−5 | PE 18:1_22:6 | 1.48 | 3.26 × 10−6 |
| LPC 17:0 | −1.34 | 0.000123 | PI O-34:3 | −2.89 | 0.001838 |
| LPC O-18:1 | −1.68 | 0.000668 | PS 18:0_22:6 | 2.4 | 3.45 × 10−7 |
| Lipid Ion | Log FC | Adjusted p-Value |
|---|---|---|
| Sphingolipids | ||
| HexCer 18:1:O2/16:0 | 2.97 | 3.76 × 10−10 |
| HexCer 42:1;O2 | 3.38 | 0.004698 |
| Phospholipids | ||
| PC 18:2_22:6 | −1.01 | 0.005038 |
| PC 20:4_22:6 | −1.3 | 0.006469 |
| PE 16:0_18:1 | 1.06 | 0.005038 |
| PE O-16:1_22:4 | 1.3 | 0.003583 |
| PE 18:0_22:6 | 1.09 | 0.006469 |
| PS 18:0_20:3 | 1.14 | 0.006469 |
| PS 18:0_22:6 | 2.46 | 1.05 × 10−7 |
| PS 40:4 | 1.06 | 0.004093 |
| Lipid Ion | Log FC | Adjusted p-Value | Lipid Ion | Log FC | Adjusted p-Value |
|---|---|---|---|---|---|
| Glycerolipids | Phospholipids | ||||
| DG 16:0_22:6 | 2.41 | 0.00016 | LPC 16:1 | −1.16 | 0.00438 |
| DG 18:1_20:4 | 1.37 | 0.006795 | LPC 18:1 | −1.21 | 0.005347 |
| DG 18:2_20:4 | 1.46 | 0.006804 | LPC 18:2 | −1.62 | 0.003162 |
| TG 16:0_16:0_22:6 | 1.86 | 0.003682 | LPC 18:4 | −1.43 | 0.002132 |
| TG 16:0_18:1_23:0 | −1.16 | 0.004729 | LPC 20:1 | −1.05 | 0.006804 |
| TG 16:0_18:1_24:0 | −1.63 | 0.004729 | LPC 20:2 | −1.5 | 0.003682 |
| TG 16:0_20:4_22:6 | 2.3 | 0.001677 | LPC 20:4 | −1.1 | 0.006804 |
| TG 16:0_22:6_22:6 | 2.77 | 0.003539 | LPC 22:4 | −1.83 | 0.003564 |
| TG O-16:0_16:0_18:1 | −1.32 | 0.002115 | LPC 22:5 | −1.82 | 0.001677 |
| TG O-16:0_16:0_18:2 | −1.8 | 0.000701 | LPE 18:1 | −1.36 | 0.000701 |
| TG O-16:0_18:1_18:2 | −2.02 | 0.000832 | LPE 18:2 | −1.68 | 0.000701 |
| TG O-16:0_18:1_20:1 | −1.35 | 0.006804 | PC 18:0_22:6 | 1.27 | 0.004289 |
| TG 18:0_18:1_22:0 | −1.72 | 0.006201 | PE 16:0_18:2 | 1.11 | 0.000449 |
| TG O-16:0_16:0_18:1 | −1.6 | 0.001852 | PE 16:0_20:4 | 1.08 | 0.000832 |
| TG O-16:0_18:1_18:2 | −1.72 | 0.003162 | PE 16:0_20:5 | 1.44 | 0.005871 |
| TG O-18:1_16:0_18:1 | −1.65 | 0.001677 | PE 16:0_22:6 | 1.27 | 0.000832 |
| TG 18:1_20:4_22:6 | 1.74 | 0.006201 | PE 18:0_20:5 | 1.06 | 0.004658 |
| TG 18:1_21:0_16:0 | −1.05 | 0.006201 | PE 18:0_22:6 | 1.56 | 0.000608 |
| TG 18:1_22:0_18:1 | −1.97 | 0.006618 | PE 18:1_22:6 | 1.2 | 0.000455 |
| TG 18:1_22:0_22:0 | −1.8 | 0.002948 | |||
| TG O-18:2_18:1_18:1 | −1.48 | 0.005676 | |||
| TG 18:2_20:4_22:6 | 1.7 | 0.004729 | |||
| TG 18:2_22:6_22:6 | 2.06 | 0.004839 | |||
| TG O-20:0_18:1_18:2 | −1.19 | 0.008578 | |||
| TG 24:1_18:2_18:1 | −1.87 | 0.008578 | |||
| TG 25:0_18:1_16:0 | −1.12 | 0.006236 | |||
| TG 25:0_18:1_18:1 | −1.56 | 0.003502 | |||
| TG 26:0_18:1_16:0 | −1.47 | 0.003682 | |||
| TG 26:0_18:1_18:1 | −1.66 | 0.002776 | |||
| TG 30:1_18:2_20:1 | −3.36 | 0.000114 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batarseh, A.M.; Vafaee, F.; Hosseini-Beheshti, E.; Safarchi, A.; Chen, A.; Cohen, A.; Juillard, A.; Hunt, N.H.; Mariani, M.; Mitchell, T.; et al. Investigation of Plasma-Derived Lipidome Profiles in Experimental Cerebral Malaria in a Mouse Model Study. Int. J. Mol. Sci. 2023, 24, 501. https://doi.org/10.3390/ijms24010501
Batarseh AM, Vafaee F, Hosseini-Beheshti E, Safarchi A, Chen A, Cohen A, Juillard A, Hunt NH, Mariani M, Mitchell T, et al. Investigation of Plasma-Derived Lipidome Profiles in Experimental Cerebral Malaria in a Mouse Model Study. International Journal of Molecular Sciences. 2023; 24(1):501. https://doi.org/10.3390/ijms24010501
Chicago/Turabian StyleBatarseh, Amani M., Fatemeh Vafaee, Elham Hosseini-Beheshti, Azadeh Safarchi, Alex Chen, Amy Cohen, Annette Juillard, Nicholas Henry Hunt, Michael Mariani, Todd Mitchell, and et al. 2023. "Investigation of Plasma-Derived Lipidome Profiles in Experimental Cerebral Malaria in a Mouse Model Study" International Journal of Molecular Sciences 24, no. 1: 501. https://doi.org/10.3390/ijms24010501
APA StyleBatarseh, A. M., Vafaee, F., Hosseini-Beheshti, E., Safarchi, A., Chen, A., Cohen, A., Juillard, A., Hunt, N. H., Mariani, M., Mitchell, T., & Grau, G. E. R. (2023). Investigation of Plasma-Derived Lipidome Profiles in Experimental Cerebral Malaria in a Mouse Model Study. International Journal of Molecular Sciences, 24(1), 501. https://doi.org/10.3390/ijms24010501