
Effect of Betaine and Arginine on Interaction of αB-Crystallin with Glycogen Phosphorylase b

 , ,
, ,
Abstract
:1. Introduction
2. Results
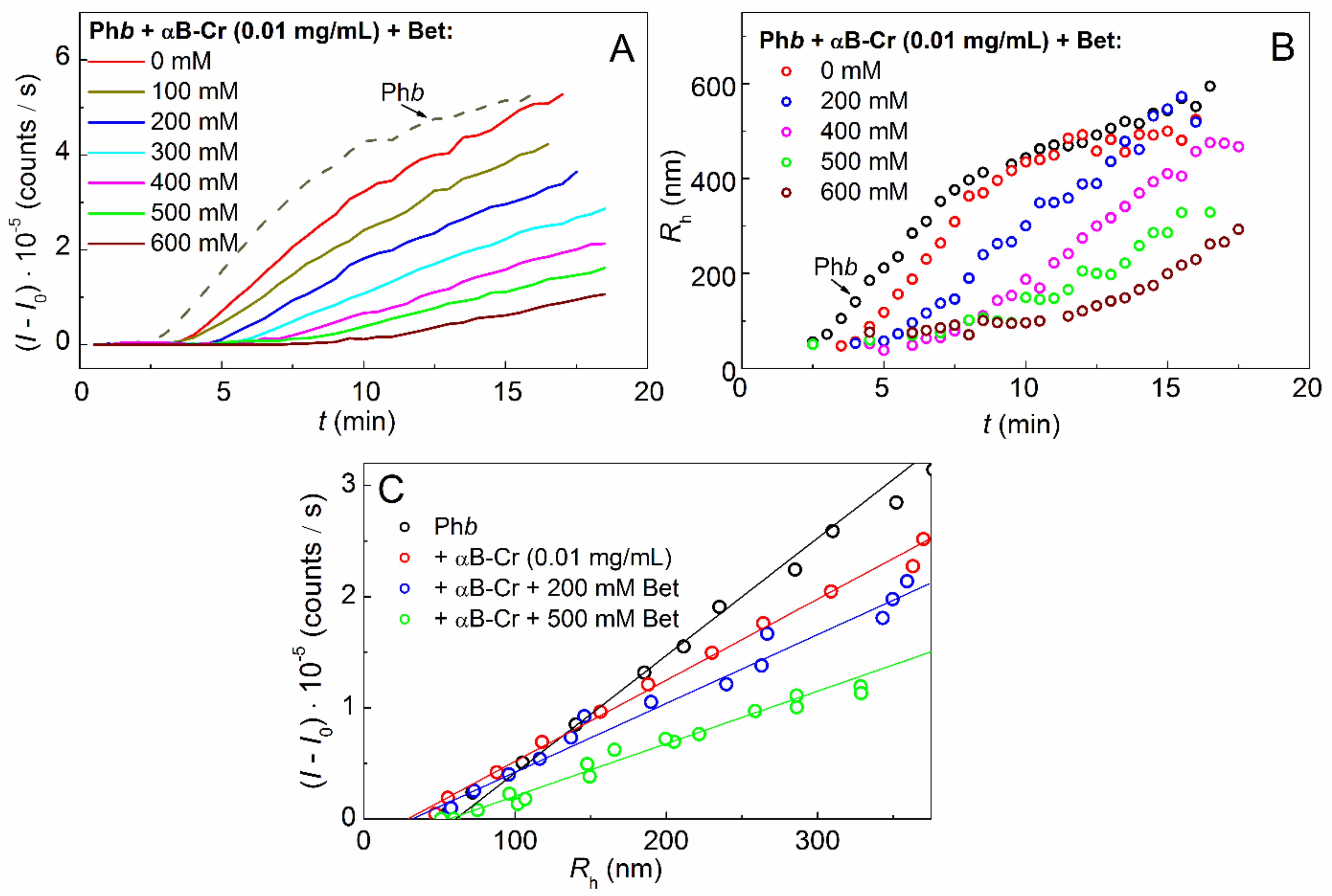
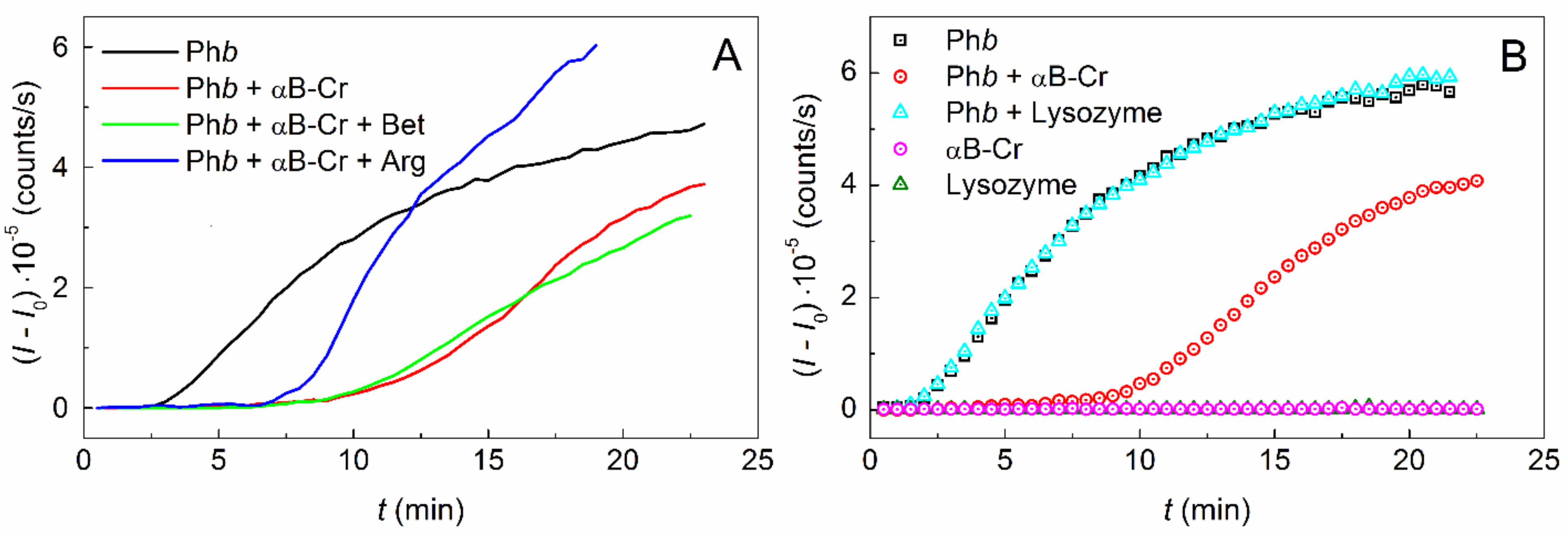
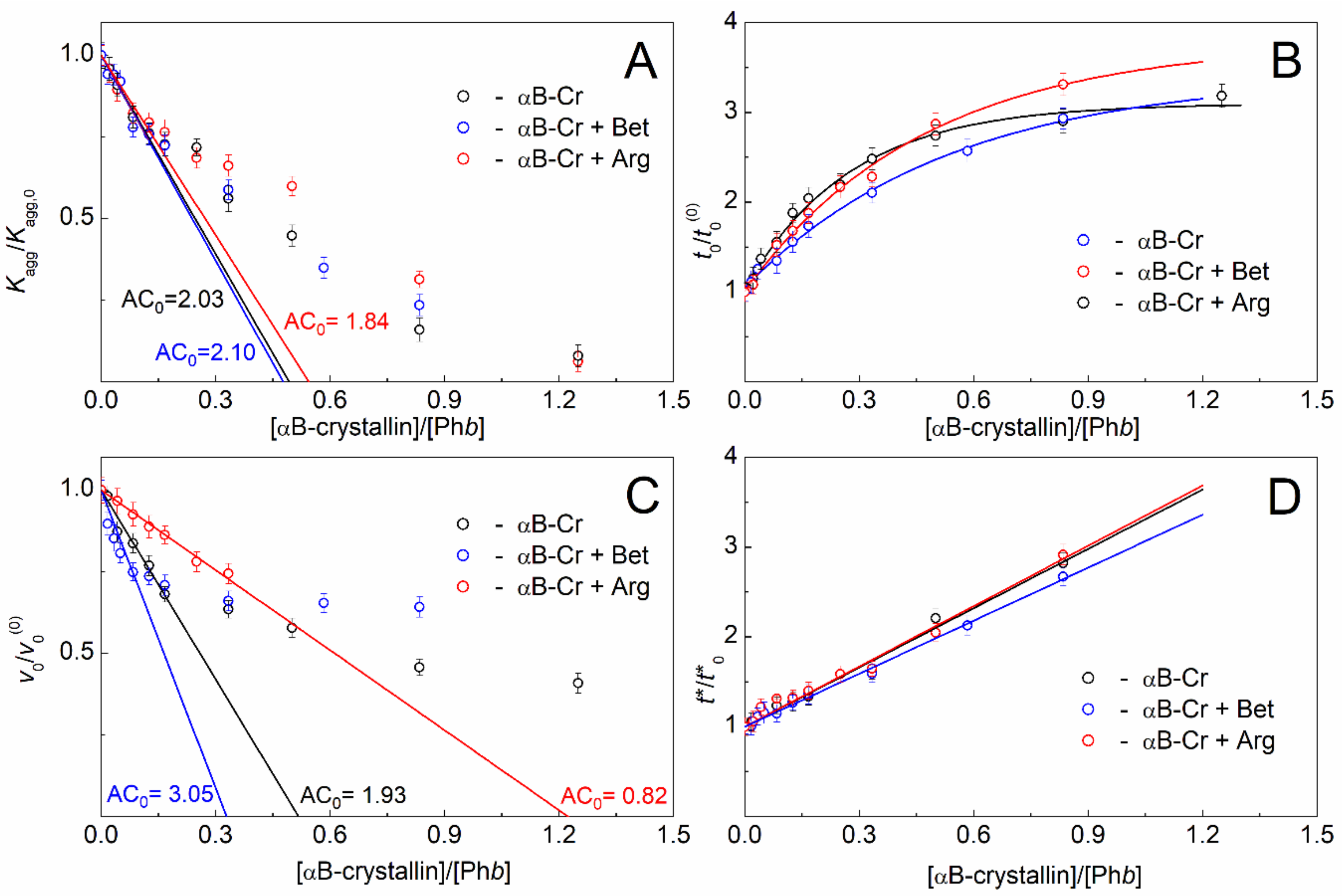
2.1. Effect of αB-Crystallin on Phb Aggregation in the Presence of Chemical Chaperones
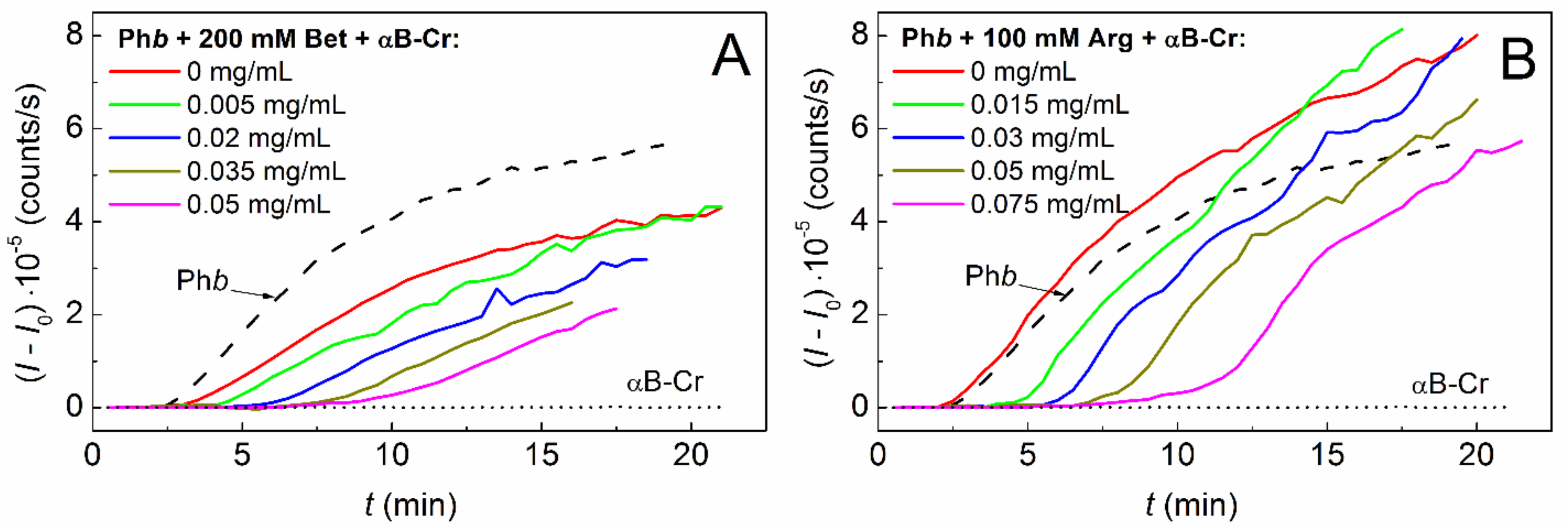
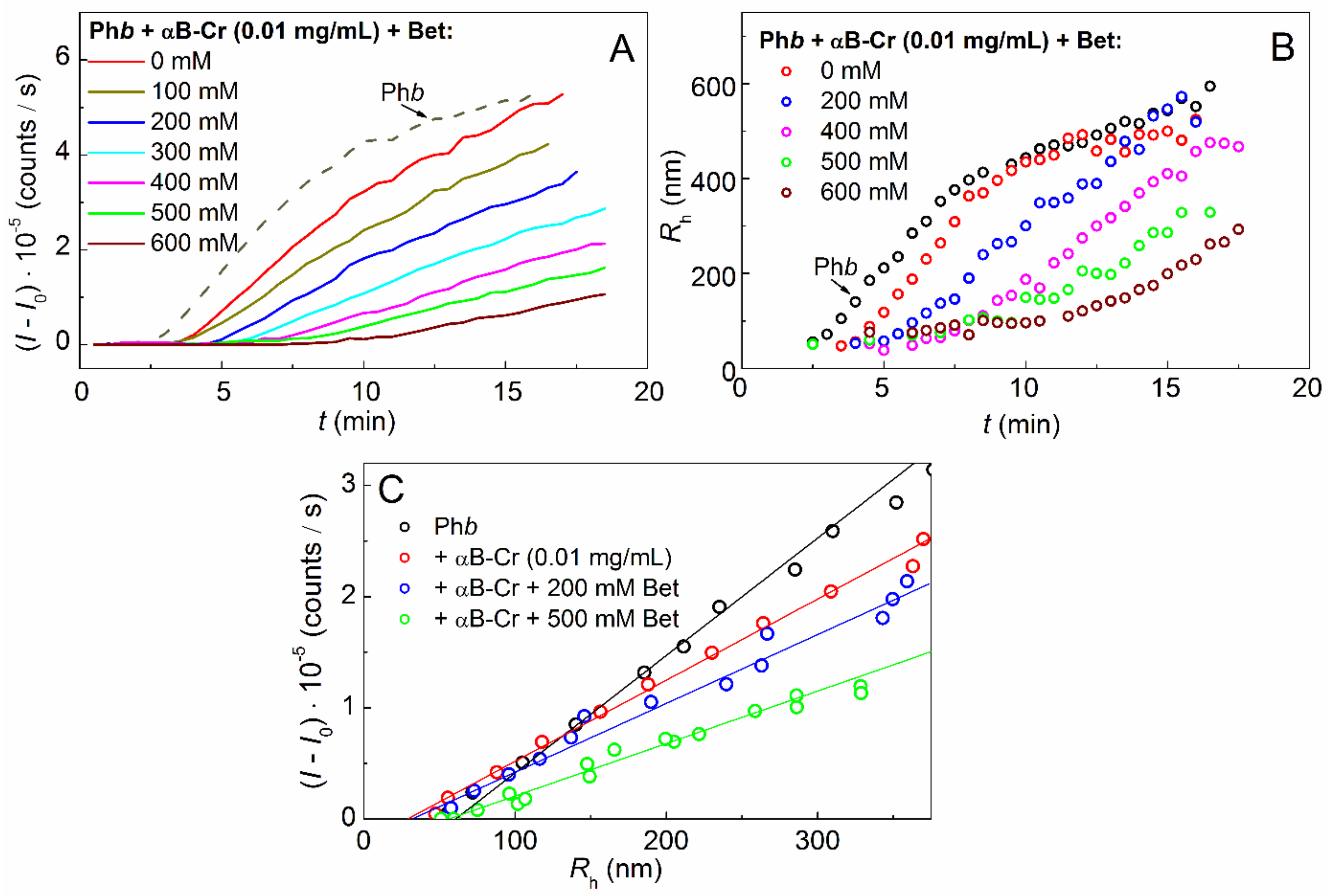
2.2. Effect of Chemical Chaperones on Phb Aggregation in the Presence of αB-Crystallin
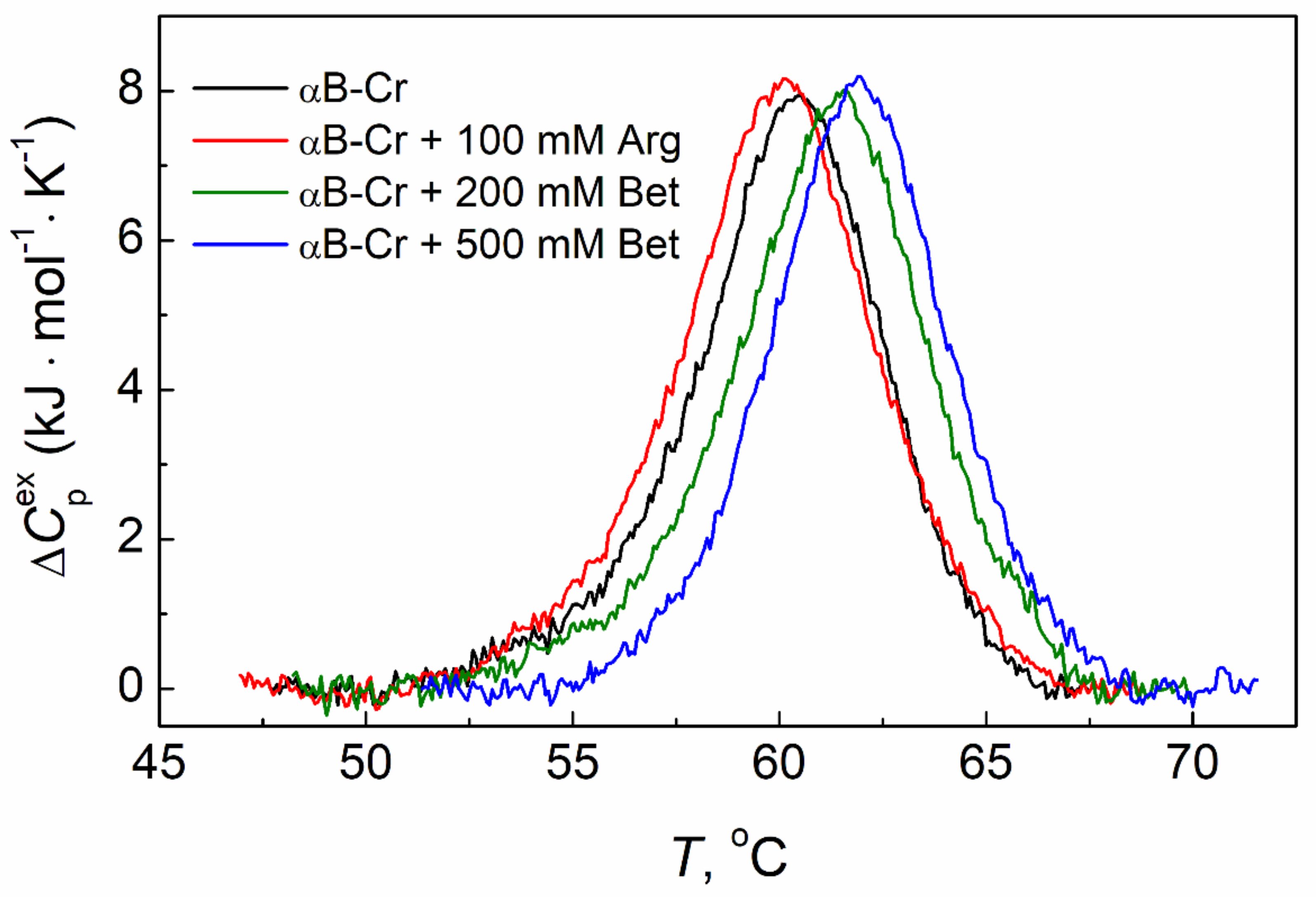
2.3. The Effect of Chemical Chaperones on Thermal Stability of αB-Crystallin
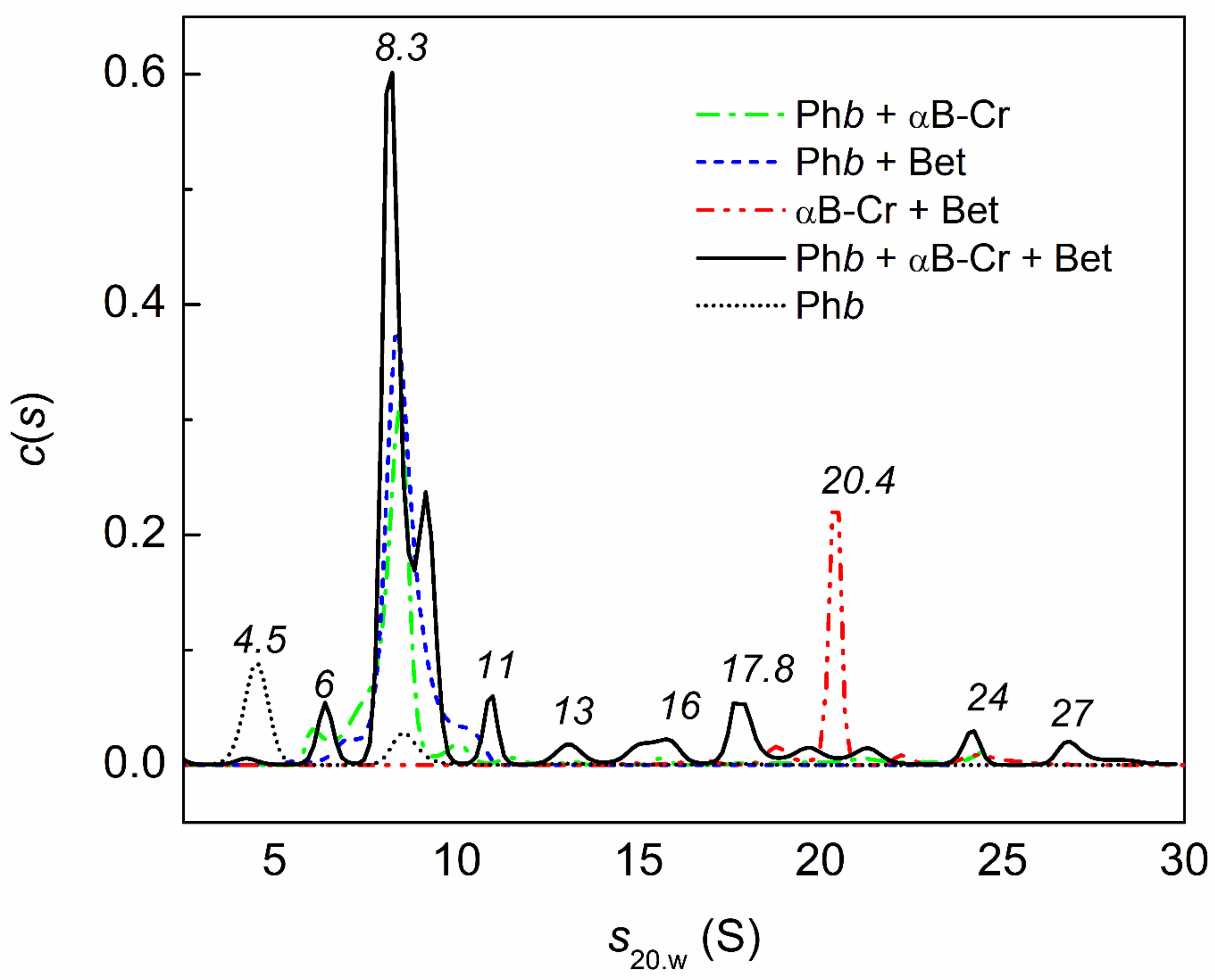
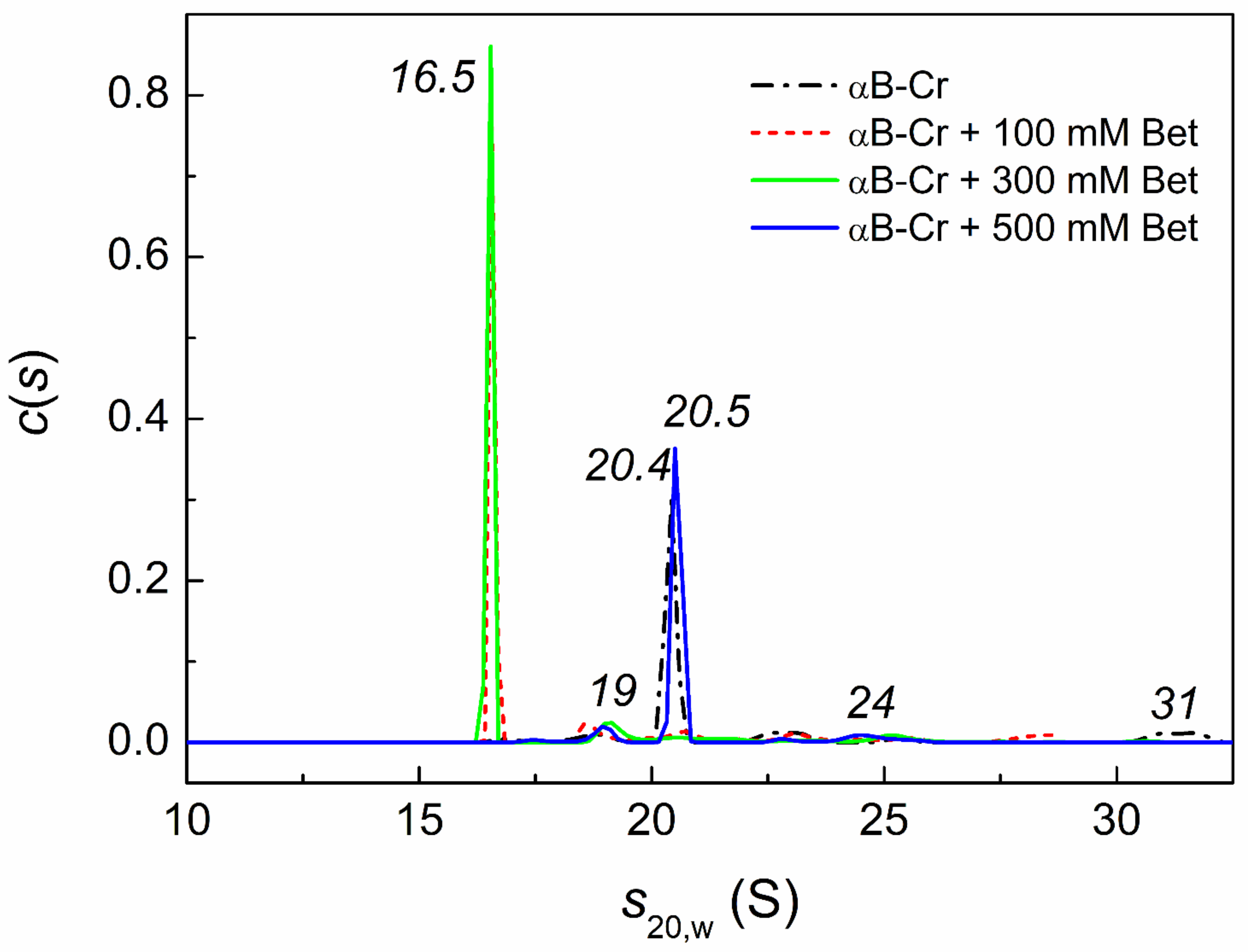
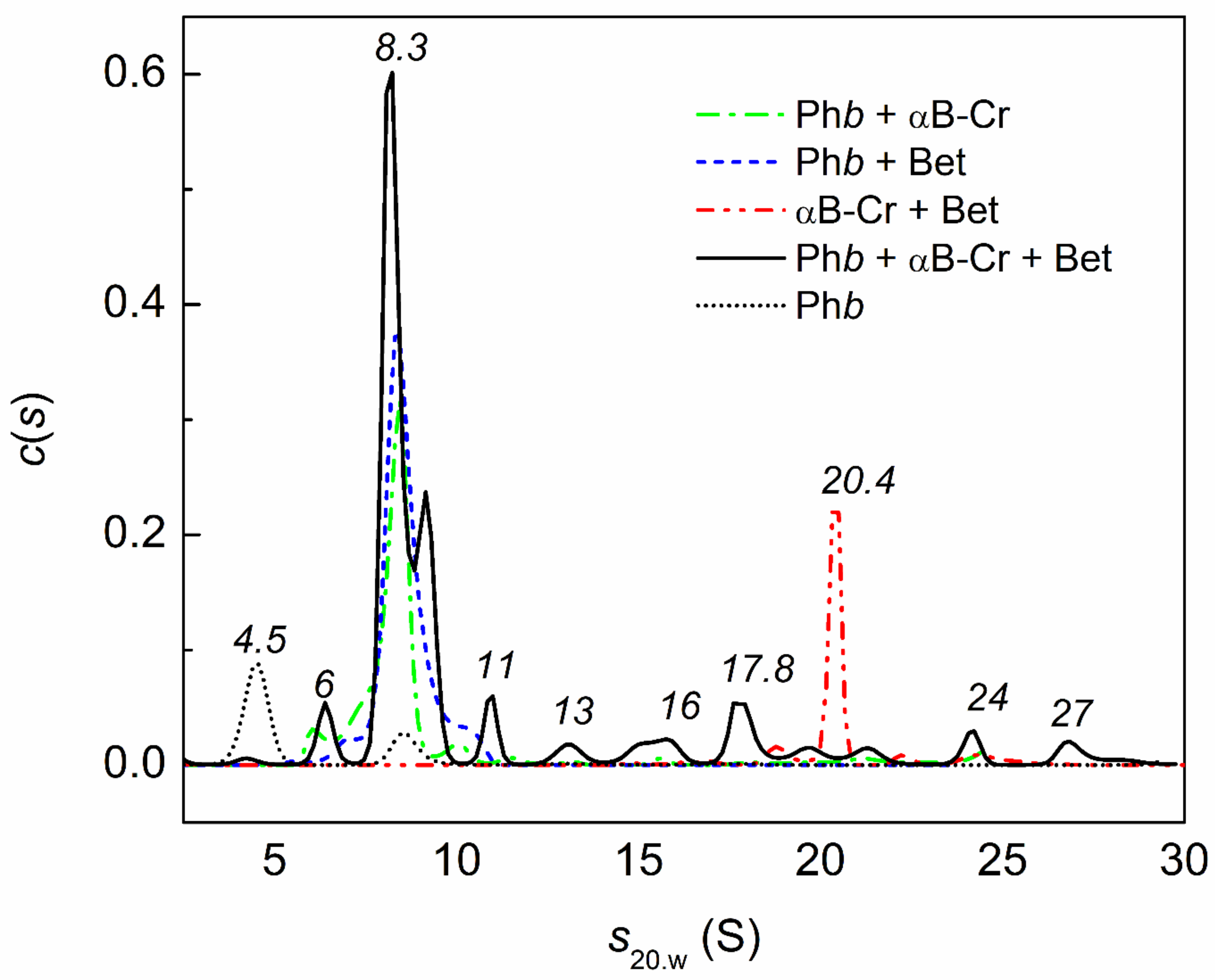
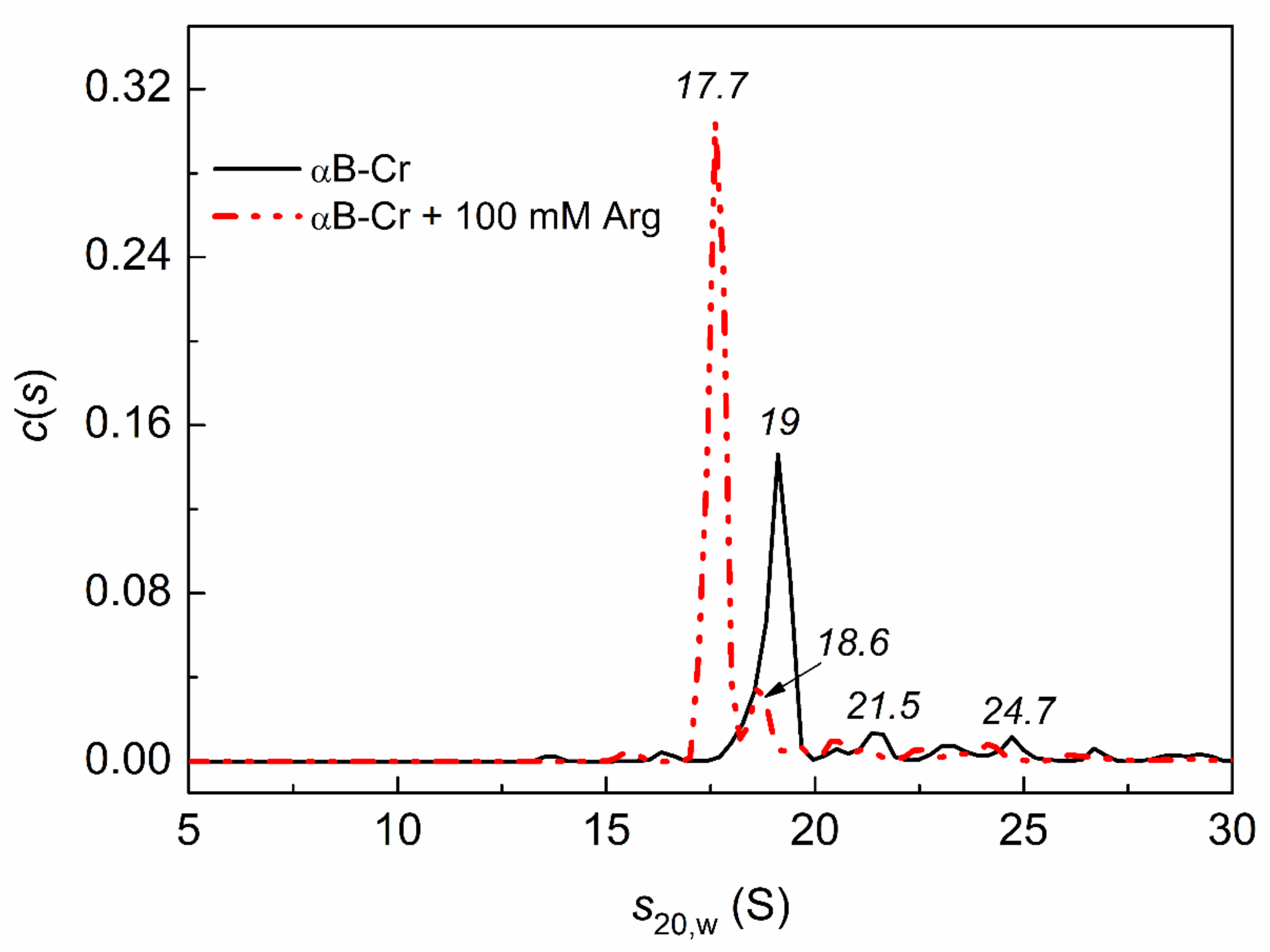
2.4. Analytical Ultracentrifugation of αB-Crystallin, Phb, and Their Mixture in the Presence of Chemical Chaperones under Heat Shock Conditions
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Isolation of αB-Crystallin (HspB5)
4.3. Dynamic Light Scattering (DLS) Study
4.4. Determination of the Adsorption Capacity of the Chaperone at Different Stages of Target Protein Aggregation
4.5. Differential Scanning Calorimetry (DSC) Studies
4.6. Analytical Ultracentrifugation (AUC)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Herce, H.D.; Deng, W.; Helma, J.; Leonhardt, H.; Cardoso, M.C. Visualization and targeted disruption of protein interactions in living cells. Nat. Commun. 2013, 4, 2660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nooren, I.M.A.; Thornton, J.M. Diversity of protein-protein interactions. EMBO J. 2003, 22, 3486–3492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gestwicki, J.E.; Shao, H. Inhibitors and chemical probes for molecular chaperone networks. J. Biol. Chem. 2019, 294, 2151–2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behnke, J.; Mann, M.J.; Scruggs, F.L.; Feige, M.J.; Hendershot, L.M. Members of the Hsp70 family recognize distinct types of sequences to execute ER quality control. Mol. Cell 2016, 63, 739–752. [Google Scholar] [CrossRef] [Green Version]
- Rosenzweig, R.; Nillegoda, N.B.; Mayer, M.P.; Bukau, B. The Hsp70 chaperone network. Nat. Mol. Cell Biol. 2019, 20, 665–680. [Google Scholar] [CrossRef]
- Schopf, F.H.; Biebl, M.M.; Buchner, J. The Hsp90 chaperone machinery. Nat. Rev. Mol. Cell 2017, 18, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Scheidta, T.; Carozzaa, J.A.; Kolbea, C.C.; Aprilea, F.A.; Tkachenko, O. The binding of the small heat-shock protein αB-crystallin to fibrils of α-synuclein is driven by entropic forces. Proc. Natl. Acad. Sci. USA 2021, 118, e2108790118. [Google Scholar] [CrossRef]
- Wang, L.; Xu, X.; Jiang, Z.; You, Q. Modulation of protein fate decision by small molecules: Targeting molecular chaperone machinery. Acta Pharm. Sin. B 2020, 10, 1904–1925. [Google Scholar] [CrossRef]
- Koldewey, P.; Horowitz, S.; Bardwell, J.C.A. Chaperone-client interactions: Non-specificity engenders multifunctionality. J. Biol. Chem. 2017, 292, 12010–12017. [Google Scholar] [CrossRef] [Green Version]
- Bukau, B.; Horwich, A.L. The Hsp70 and Hsp60 chaperone machines. Cell 1998, 92, 351–366. [Google Scholar] [CrossRef] [Green Version]
- Klaips, C.L.; Jayaraj, G.G.; Hartl, F.U. Pathways of cellular proteostasis in aging and disease. J. Cell Biol. 2018, 217, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.; Fox, S.G.; Cavallini, A.; Kerridge, C.; O’Neill, M.J.; Wolak, J.; Bose, S. Tau protein aggregates inhibit the protein-folding and vesicular trafficking arms of the cellular proteostasis network. J. Biol. Chem. 2019, 294, 7917–7930. [Google Scholar] [CrossRef] [PubMed]
- Haslbeck, M.; Vierling, E.A. First line of stress defense: Small heat shock proteins and their function in protein homeostasis. J. Mol. Biol. 2015, 427, 1537–1548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; MacRae, T.H. The small heat shock proteins and their role in human disease. FEBS J. 2005, 272, 2613–2627. [Google Scholar] [CrossRef] [PubMed]
- Laskowska, E.; Matuszewska, E.; Kuczynska-Wisnik, D. Small heat-shock proteins and protein-misfolding diseases. Curr. Pharm. Biotechnol. 2010, 11, 146–157. [Google Scholar] [CrossRef]
- Mymrikov, E.V.; Seit-Nebi, A.S.; Gusev, N.B. Large potentials of small heat shock proteins. Physiol. Rev. 2011, 91, 1123–1159. [Google Scholar] [CrossRef] [Green Version]
- De Jong, W.W.; Caspers, G.-J.; Leunissen, J.A.M. Genealogy of the alpha-crystallin-small heat-shock protein superfamily. Int. J. Biol. Macromol. 1998, 22, 151–162. [Google Scholar] [CrossRef]
- Riedl, M.; Strauch, A.; Catici, D.A.M.; Haslbeck, M. Proteinaceous transformers: Structural and functional variability of human sHsps. Int. J. Mol. Sci. 2020, 21, 5448. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, C.; Li, Y.; Zhao, C.; Li, T.; Li, D.; Zhang, S.; Li, C. Mechanistic insights into the switch of αB-crystallin chaperone activity and self-multimerization. J. Biol. Chem. 2018, 293, 14880–14890. [Google Scholar] [CrossRef] [Green Version]
- Sprague-Piercy, M.A.; Rocha, M.A.; Kwok, A.O.; Martin, R.W. α-Crystallins in the Vertebrate Eye Lens: Complex Oligomers and Molecular Chaperones. Annu. Rev. Phys. Chem. 2021, 72, 143–163. [Google Scholar] [CrossRef]
- Inoue, R.; Takata, T.; Fujii, N.; Ishii, K.; Uchiyama, S.; Sato, N.; Oba, Y.; Wood, K.; Kato, K.; Fuji, N.; et al. New insight into the dynamical system of αB-crystallin oligomers. Sci. Rep. 2016, 6, 29208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, J.; Carver, J.A. The multifaceted nature of αB-crystallin. Cell Stress Chaperones 2020, 25, 639–654. [Google Scholar] [CrossRef] [PubMed]
- Chebotareva, N.A.; Eronina, T.B.; Sluchanko, N.N.; Kurganov, B.I. Effect of Ca2+ and Mg2+ ions on oligomeric state and chaperone-like activity of αB-crystallin in crowded media. Int. J. Biol. Macromol. 2015, 76, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Voziyan, P.A.; Fisher, M.T. Chaperonin-assisted folding of glutamine synthetase under nonpermissive conditions: Off-pathway aggregation propensity does not determine the co-chaperonin requirement. Protein Sci. 2000, 9, 2405–2412. [Google Scholar] [CrossRef] [Green Version]
- Diamant, S.; Eliahu, N.; Rosenthal, D.; Goloubinoff, P. Chemical chaperones regulate molecular chaperones in vitro and in cells under combined salt and heat stresses. J. Biol. Chem. 2001, 276, 39586–39591. [Google Scholar] [CrossRef] [Green Version]
- Diamant, S.; Rosenthal, D.; Azem, A.; Eliahu, N.; Ben-Zvi, A.P.; Goloubinoff, P. Dicarboxylic amino acids and glycine-betaine regulate chaperone-mediated protein disaggregation under stress. Mol. Microbiol. 2003, 49, 401–410. [Google Scholar] [CrossRef]
- Srinivas, V.; Raman, B.; Rao, K.S.; Ramakrishna, T.; Rao, C.M. Structural perturbation and enhancement of the chaperone-like activity of alpha-crystallin by arginine hydrochloride. Protein Sci. 2003, 12, 1262–1270. [Google Scholar] [CrossRef]
- Srinivas, V.; Raman, B.; Rao, K.S.; Ramakrishna, T.; Rao, C.M. Arginine hydrochloride enhances the dynamics of subunit assembly and the chaperone-like activity of alpha-crystallin. Mol. Vis. 2005, 11, 249–255. [Google Scholar] [PubMed]
- Ecroyd, H.; Carver, J.A. The effect of small molecules in modulating chaperone activity of alphaB-crystallin against ordered and disordered protein aggregation. FEBS J. 2008, 275, 935–947. [Google Scholar] [CrossRef]
- Mikhaylova, V.V.; Eronina, T.B.; Chebotareva, N.A.; Shubin, V.V.; Kalacheva, D.I.; Kurganov, B.I. Effect of Arginine on Chaperone-Like Activity of HspB6 and Monomeric 14-3-3ζ. Int. J. Mol. Sci. 2020, 21, 2039. [Google Scholar] [CrossRef] [Green Version]
- Eronina, T.B.; Chebotareva, N.A.; Sluchanko, N.N.; Mikhaylova, V.V.; Makeeva, V.F.; Roman, S.G.; Kleymenov, S.Y.; Kurganov, B.I. Dual effect of arginine on aggregation of phosphorylase kinase. Int. J. Biol. Macromol. 2014, 68, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Treweek, T.M.; Meehan, S.; Ecroyd, H.; Carver, J.A. Small heat-shock proteins: Important players in regulating cellular proteostasis. Cell Mol. Life Sci. 2015, 72, 429–451. [Google Scholar] [CrossRef] [PubMed]
- Eronina, T.B.; Mikhaylova, V.V.; Chebotareva, N.A.; Kurganov, B.I. Kinetic regime of thermal aggregation of holo- and apoglycogen phosphorylases b. Int. J. Biol. Macromol. 2016, 92, 1252–1257. [Google Scholar] [CrossRef] [PubMed]
- Chebotareva, N.A.; Roman, S.G.; Borzova, V.A.; Eronina, T.B.; Mikhaylova, V.V.; Kurganov, B.I. Chaperone-like activity of HspB5: The effects of quaternary structure dynamics and crowding. Int. J. Mol. Sci. 2020, 21, 4940. [Google Scholar] [CrossRef]
- Eronina, T.B.; Mikhaylova, V.V.; Chebotareva, N.A.; Shubin, V.V.; Kleymenov, S.Y.; Kurganov, B.I. Effect of arginine on stability and aggregation of muscle glycogen phosphorylase b. Int. J. Biol. Macromol. 2020, 165, 365–374. [Google Scholar] [CrossRef]
- Eronina, T.B.; Mikhaylova, V.V.; Chebotareva, N.A.; Kleymenov, S.Y.; Pivovarova, A.V.; Kurganov, B.I. Combined action of chemical chaperones on stability, aggregation and oligomeric state of muscle glycogen phosphorylase b. Int. J. Biol. Macromol. 2022, 203, 406–416. [Google Scholar] [CrossRef]
- Golub, N.; Meremyanin, A.; Markossian, K.; Eronina, T.; Chebotareva, N.; Asryants, R.; Muronets, V.; Kurganov, B. Evidence for the formation of start aggregates as an initial stage of protein aggregation. FEBS Lett. 2007, 581, 4223–4227. [Google Scholar] [CrossRef] [Green Version]
- Khanova, H.A.; Markossian, K.A.; Kurganov, B.I.; Samoilov, A.M.; Kleimenov, S.Y.; Levitsky, D.I.; Yudin, I.K.; Timofeeva, A.C.; Muranov, K.O.; Ostrovsky, M.A. Mechanism of chaperone-like activity. Suppression of thermal aggregation of βL-crystallin by α-crystallin. Biochemistry 2005, 44, 15480–15487. [Google Scholar] [CrossRef]
- Chebotareva, N.A.; Eronina, T.B.; Roman, S.G.; Mikhaylova, V.V.; Sluchanko, N.N.; Gusev, N.B.; Kurganov, B.I. Oligomeric state of αB-crystallin under crowded conditions. Biochem. Biophys. Res. Commun. 2019, 508, 1101–1105. [Google Scholar] [CrossRef]
- Grosas, A.B.; Rekas, A.; Mata, J.P.; Thorn, D.C.; Carver, J.A. The aggregation of αB-crystallin under crowding conditions is prevented by αA-crystallin: Implications for α-crystallin stability and lens transparency. J. Mol. Biol. 2020, 432, 5593–5613. [Google Scholar] [CrossRef]
- Roman, S.G.; Chebotareva, N.A.; Eronina, T.B.; Kleymenov, S.Y.; Makeeva, V.P.; Poliansky, N.B.; Muranov, K.O.; Kurganov, B.I. Does the crowded ceii-like environment reduce the chaperone-like activity of a-crystallin? Biochemistry 2011, 50, 10607–10623. [Google Scholar] [CrossRef] [PubMed]
- Chebotareva, N.A.; Makeeva, V.F.; Bazhina, S.G.; Eronina, T.B.; Gusev, N.B.; Kurganov, B.I. Interaction of Hsp27 with native phosphorylase kinase under crowding conditions. Macromol. Biosci. 2010, 10, 783–789. [Google Scholar] [CrossRef] [PubMed]
- Arosto, P.; Michaels, T.C.T.; Linse, S.; Mansson, C.; Emanuelsson, C.; Presto, J.; Johansson, J.; Vendruscolo, M.; Dobson, C.M.; Knowles, T.P.J. Kinetic analysis reveals the diversity of microscopic mechanisms through which molecular chaperones suppress amyloid formation. Nat. Commun. 2016, 7, 10948. [Google Scholar] [CrossRef] [Green Version]
- Chebotareva, N.A.; Filippov, D.O.; Kurganov, B.I. Effect of crowding on several stages of protein aggregation in test systems in the presence of α-crystallin. Int. J. Biol. Macromol. 2015, 80, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Bumagina, Z.M.; Gurvits, B.Y.; Artemova, N.V.; Muranov, K.O.; Yudin, I.K.; Kurganov, B.I. Mechanism of suppression of dithiothreitol-induced aggregation of bovine α-lactalbumin by α-crystallin. Biophys. Chem. 2010, 146, 108–117. [Google Scholar] [CrossRef]
- Chebotareva, N.A.; Eronina, T.B.; Mikhaylova, V.V.; Roman, S.G.; Tugaeva, K.V.; Kurganov, B.I. Effect of trehalose on oligomeric state and anti-aggregation activity of αB-crystallin. Biochemistry 2022, 87, 121–130. [Google Scholar] [CrossRef]
- Kumar, C.S.; Swamy, M.J. Modulation of chaperone-like and membranolytic activities of major horse seminal plasma protein HSP-1/2 by L-carnitine. J. Biosci. 2017, 42, 469–479. [Google Scholar] [CrossRef]
- McHaourab, H.S.; Dobson, E.K.; Koteiche, H.A. Mechanism of chaperone function in small heat shock proteins. Two-mode binding of the excited states of T4 lysozyme mutants by αA-crystallin. J. Biol. Chem. 2002, 277, 40557–40566. [Google Scholar] [CrossRef] [Green Version]
- Arhar, T.; Shkedi, A.; Nadel, C.M.; Gestwiski, J.E. The interactions of molecular chaperones with client proteins: Why are they so weak? J. Biol. Chem. 2021, 297, 101282. [Google Scholar] [CrossRef]
- Vabulas, R.M.; Raychaudhuri, S.; Hayer-Hartl, M.; Hartl, F.U. Protein folding in the cytoplasm and the heat shock response. Cold Spring Harb. Perspect. Biol. 2010, 2, a004390. [Google Scholar] [CrossRef]
- Nagaraj, R.H.; Oya-Ito, T.; Padayatti, P.S.; Kumar, R.; Mehta, S.; West, K.; Levison, B.; Sun, J.; Crabb, J.W.; Padival, A.K. Enhancement of chaperone function of alpha-crystallin by methylglyoxal modification. Biochemistry 2003, 42, 10746–10755. [Google Scholar] [CrossRef] [PubMed]
- Ecroyd, H.; Meehan, S.; Horwitz, J.; Aquilina, J.A.; Benesch, J.L.; Robinson, C.V.; Macphee, C.E.; Carver, J.A. Mimicking αB-crystallin phosphorylation affects its chaperone activity. Biochem. J. 2007, 401, 129–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freilich, R.; Arhar, T.; Abrams, J.L.; Gestwicki, J.E. Protein-protein interactions in the molecular chaperone network. Acc. Chem. Res. 2018, 51, 940–949. [Google Scholar] [CrossRef]
- Haslbeck, M.; Miess, A.; Stromer, T.; Walter, S.; Buchner, J. Disassembling protein aggregates in the yeast cytosol. The cooperation of Hsp26 with Ssa1 and Hsp104. J. Biol. Chem. 2005, 280, 23861–23868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.J.; Vierling, E. A small heat shock protein cooperates withheat shock protein 70 system to reactivate a heat-denatured protein. Plant Physiol. 2000, 122, 189–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eronina, T.B.; Chebotareva, N.A.; Bazhina, S.G.; Makeeva, V.F.; Kleymenov, S.Y.; Kurganov, B.I. Effect of proline on thermal inactivation, denaturation and aggregation of glycogen phosphorylase b from rabbit skeletal muscle. Biophys. Chem. 2009, 141, 66–74. [Google Scholar] [CrossRef]
- Mymrikov, E.V.; Bukach, O.V.; Seit-Nebi, A.S.; Gusev, N.B. The pivotal role of the beta 7 strand in the intersubunit contacts of different human small heat shock proteins. Cell Stress Chaperones 2010, 15, 365–377. [Google Scholar] [CrossRef] [Green Version]
- Eronina, T.B.; Mikhaylova, V.V.; Chebotareva, N.A.; Borzova, V.A.; Yudin, I.K.; Kurganov, B.I. Mechanism of aggregation of UV-irradiated glycogen phosphorylase b at a low temperature in the presence of crowders and trimethylamine N-oxide. Biophys. Chem. 2018, 232, 12–21. [Google Scholar] [CrossRef]
- Kurganov, B.I. Antiaggregation activity of chaperones and its quantification. Biochemistry 2013, 78, 1554–1566. [Google Scholar] [CrossRef]
- Kurganov, B.I. Quantification of anti-aggregation activity of chaperones. Int. J. Biol. Macromol. 2017, 100, 104–117. [Google Scholar] [CrossRef]
- Brown, P.H.; Schuck, P. Macromolecular size-and-shape distributions by sedimentation velocity analytical ultracentrifugation. Biophys. J. 2006, 90, 4651–4661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikhaylova, V.V.; Eronina, T.B.; Chebotareva, N.A.; Kleymenov, S.Y.; Shubin, V.V.; Kurganov, B.I. A thermal after-effect of UV irradiation of muscle glycogen phosphorylase b. PLoS ONE 2017, 12, e0189125. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component with Variable Concentration | Nucleation Stage | Stage of Aggregate Growth |
|---|---|---|
| αB-Cr | AC0 = 2.03 ± 0.08 Phb monomer per 1 subunit of αB-Cr | AC0 = 1.93 ± 0.10 Phb monomer per 1 subunit of αB-Cr |
| αB-Cr in the presence of 200 mM Bet | AC0 = 2.10 ± 0.17 Phb monomer per 1 subunit of αB-Cr | AC0 = 3.05 ± 0.34 Phb monomer per 1 subunit of αB-Cr |
| αB-Cr in the presence of 100 mM Arg | AC0 = 1.84 ± 0.15 Phb monomer per 1 subunit of αB-Cr | AC0 = 0.82 ± 0.05 Phb monomer per 1 subunit of αB-Cr |
| Sample | Betaine (mM) | Rh,0 (nm) |
|---|---|---|
| Phb (0.3 mg/mL) | 0 | 62.7 ± 3 |
| Phb + αB-Cr | 0 | 30.8 ± 2 |
| 200 | 34.1 ± 2 | |
| 400 | 38.1 ± 2 | |
| 500 | 48.8 ± 2 | |
| 600 | 52.5 ± 2 |
| Sample | Tmax (°C) | ΔHcal (kJ·mol−1) |
|---|---|---|
| αB-Cr | 60.5 ± 0.1 | 44.2 ± 2.7 |
| αB-Cr + 100 mM Arg | 60.1 ± 0.1 | 48.9 ± 2.9 |
| αB-Cr + 200 mM Bet | 61.6 ± 0.1 | 46.4 ± 2.8 |
| αB-Cr + 500 mM Bet | 62.0 ± 0.1 | 44.4 ± 2.7 |
| Sample | γagg (%) |
|---|---|
| Phb (0.5 mg/mL) | 33 |
| Phb + αB-Cr + 500 mM Bet | 12 |
| Phb + 500 mM Bet | 0 |
| Phb + 100 mM Arg | 96 |
| Phb + αB-Cr + 100 mM Arg | 96 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eronina, T.B.; Mikhaylova, V.V.; Chebotareva, N.A.; Tugaeva, K.V.; Kurganov, B.I. Effect of Betaine and Arginine on Interaction of αB-Crystallin with Glycogen Phosphorylase b. Int. J. Mol. Sci. 2022, 23, 3816. https://doi.org/10.3390/ijms23073816
Eronina TB, Mikhaylova VV, Chebotareva NA, Tugaeva KV, Kurganov BI. Effect of Betaine and Arginine on Interaction of αB-Crystallin with Glycogen Phosphorylase b. International Journal of Molecular Sciences. 2022; 23(7):3816. https://doi.org/10.3390/ijms23073816
Chicago/Turabian StyleEronina, Tatiana B., Valeriya V. Mikhaylova, Natalia A. Chebotareva, Kristina V. Tugaeva, and Boris I. Kurganov. 2022. "Effect of Betaine and Arginine on Interaction of αB-Crystallin with Glycogen Phosphorylase b" International Journal of Molecular Sciences 23, no. 7: 3816. https://doi.org/10.3390/ijms23073816
APA StyleEronina, T. B., Mikhaylova, V. V., Chebotareva, N. A., Tugaeva, K. V., & Kurganov, B. I. (2022). Effect of Betaine and Arginine on Interaction of αB-Crystallin with Glycogen Phosphorylase b. International Journal of Molecular Sciences, 23(7), 3816. https://doi.org/10.3390/ijms23073816