Abstract
Early life stress (ELS) encompasses exposure to aversive experiences during early development, such as neglect or maltreatment. Animal and human studies indicate that ELS has maladaptive effects on brain development, leaving individuals more vulnerable to developing behavioral and neuropsychiatric disorders later in life. This result occurs in part to disruptions in Brain derived neurotrophic factor (Bdnf) gene regulation, which plays a vital role in early neural programming and brain health in adulthood. A potential treatment mechanism to reverse the effects of ELS on Bdnf expression is aerobic exercise due to its neuroprotective properties and positive impact on Bdnf expression. Aerobic exercise opens the door to exciting and novel potential treatment strategies because it is a behavioral intervention readily and freely available to the public. In this review, we discuss the current literature investigating the use of exercise interventions in animal models of ELS to reverse or mitigate ELS-induced changes in Bdnf expression. We also encourage future studies to investigate sensitive periods of exercise exposure, as well as sufficient duration of exposure, on epigenetic and behavioral outcomes to help lead to standardized practices in the exercise intervention field.
1. Introduction
Over 30 years ago, Dr. David Barker presented the Barker Hypothesis, which stated that the perinatal environment sets off a chain reaction of neural programming that determines cognitive function and emotional health in adulthood [1]. In line with this idea, the perinatal period is marked by a degree of neural plasticity that is highly sensitive to environmental influences and not seen during any later period of life [2,3,4]. During this developmental period, a cascade of neural processes work in tandem to program the brain [4]. Negative experiences during early development, such as stress, can alter the epigenetic regulation of neurotrophins [5,6,7,8] and thereby increase an individual’s susceptibility to later development of neuropsychiatric and neurodegenerative disorders [6,7,8,9]. Epigenetics refers to the addition of molecules, such as methyl groups, to DNA strands that alter the way that DNA can be read and used in the body. Hence, the consequences of these early disruptions can become embedded in the DNA and stay with a person through their entire lifespan.
Neurotrophins regulate brain development during infancy and adolescence, and in adulthood, they facilitate synaptic plasticity and neuronal survival [6,10,11,12]. Perturbations during development often dysregulate neurotrophin expression, leading to brain-region-specific maladaptive changes in expression [13,14,15,16]. In contrast, aerobic exercise typically upregulates neurotrophin expression, and is thought to be neuroprotective [17,18,19,20]. Most notably, exercise has a neuroprotective effect on the brain by preventing neuronal cell death [21] and facilitating adaptive cellular processes, including synaptogenesis (formation of synapses) [21] and neurogenesis [22]. For example, rodent studies show that aerobic exercise promotes neurogenesis in the hippocampus, a brain region where neurogenesis is abundant throughout adulthood [22,23]. This is important when considering that neurogenesis is reduced in the hippocampus of rodents exposed to developmental stress, making exercise a potential treatment mechanism for the maladaptive biological effects of stress [24,25,26,27]. Consistent with findings in animal subjects, imaging studies in humans indicate that child maltreatment leads to a decrease in hippocampal volume [28,29,30], as well as decreased hippocampal activation during threat detection [31] and memory tasks [32].
In this review we discuss the literature on rodent models of early life stress (ELS) and exercise interventions. We pay special attention to the effect of these experiences on neurotrophin regulation, with the capacity of exercise to correct biological processes and aberrant behavior associated with early stress.
2. Modeling Early Life Stress and Exercise in Rodents
In humans, ELS encompasses many different experiences. These range from physical and emotional abuse and neglect to living in a war-torn country, experiencing extreme poverty, or the death of a caregiver. In the laboratory, researchers recapitulate similar experiences in animal models to investigate the biological consequences of stress during early development. Two common models used are the limited bedding and nesting (LBN) model and the maternal separation (MS) model. In the LBN model, a rodent mother is not provided with sufficient nesting material to properly care for the pups. This causes stress in the dam and elicits more aversive behaviors from her towards the pups, such as actively avoiding and rough handling them, and less frequent or fragmented maternal behaviors such as arched back nursing and hovering over the pups [33,34]. This model is used to study the consequences of disrupted infant–caregiver interactions during early life. In the MS models, pups are separated from the dam for varying periods of time during early development [33,35] to approximate the experience of caregiver neglect.
Rodent models of aerobic exercise either use involuntary treadmill running or voluntary wheel running paradigms. Currently, the timing and duration of exercise exposure as a treatment intervention has not been standardized (see Figure 1). In voluntary models, rodents have continuous free access to running wheels, usually via a window cut out in the cage that allows the subjects to freely migrate to an attached wheel. Involuntary exercise models employ a treadmill apparatus that keeps the subject’s feet on the treadmill, forcing the subject to locomote for a set duration of time and intensity. Additional studies have employed both voluntary and involuntary aerobic exercise to compare the effects of the two running models. One study found that voluntary running wheel exercise decreased immobility time during a forced swim test of adolescent rats exposed to maternal separation; however, this antidepressant-like effect was lost in the involuntary exercised subjects [36]. Studies of this nature question the efficacy of forced exercise models as they likely upregulate the subject’s stress response, leading to a diminished neuroprotective effect. For example, Ke and colleagues [37] employed an animal model of stroke to measure the ability of aerobic exercise to recover motor behavior function and increase brain-derived neurotrophic factor (BDNF) protein levels in the hippocampus. Their results showed that voluntary, but not involuntary, exercise improved motor behavior overtime and increased BDNF in the hippocampus. In contrast, rats in the involuntary exercise group showed increased corticosterone (CORT) levels as well as a decrease in BDNF levels in the hippocampus compared to the control group [37]. These results are further supported by a 2016 study that found involuntary exercise to be maladaptive for stroke recovery in animal models. Svensson and colleagues [38] showed that, in a model of ischemic stroke, involuntary exercise increased anxiety-like behaviors on the open-field test (OFT), increased neuron loss in the right hippocampus, and increased fecal CORT levels following the OFT. They also noted a positive correlation between CORT levels and neuron loss [38]. In a 2014 study by Uysal and colleagues [39], voluntary exercise led to a decrease in basal CORT levels compared to both sedentary and involuntary exercised rats that was also accompanied by an increase in locomotion on the OFT in voluntarily exercised rats. Results from this study also showed that female rats exposed to voluntary exercise had increased BDNF protein levels in the prefrontal cortex (PFC) compared to sedentary rats. In male rats, BDNF levels increased in both exercise groups compared to the sedentary rats; however, there was a significantly greater increase in BDNF levels in the PFC of voluntarily exercised rats compared to the involuntary exercise group [39]. Taken together, these studies indicate that voluntary exercise models may be more advantageous when investigating the anxiolytic and neuroprotective effects of exercise, as involuntary exercise may exacerbate neural insults, including those caused by ELS, by upregulating CORT reactivity in the brain.
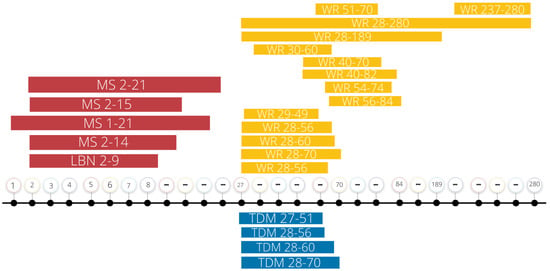
Figure 1.
Summary of current ELS and exercise intervention protocols. This summary figure represents the days, recorded in postnatal days, of early life stress exposure and exercise interventions recounted in this review. Note the variability in experimental parameters. MS, maternal separation; LBN, limited bedding and nesting; WR, wheel running; TDM, treadmill; “…”, breaks in the timeline.
In addition to the type of exercise, the duration of exercise and age of exposure may play a role in its effectiveness. For example, Greenwood and colleagues [40] showed that six, but not three, weeks of voluntary wheel running was sufficient to prevent learned helplessness behaviors when subjects were exposed to uncontrolled tail shocks later. This same research group also showed that chronic voluntary exercise exposure is more rewarding in rats compared to short-term exposure. Six weeks, but not two weeks, of voluntary exercise led to exercise-induced changes in gene expression and receptor activity in the mesolimbic dopamine pathway that were accompanied by preference for a chamber that was previously paired with wheel running exposure on a conditioned place preference task [41]. Six weeks of voluntary exercise is also sufficient to decrease habituation time to future stress (loud noise exposure) as measured by significantly reduced plasma CORT levels compared to sedentary rats [42]. A breakdown of the ELS models and exercise intervention methods used in the current literature is provided in Table 1.

Table 1.
Summary of Methods in Currently Published ELS and Exercise Intervention Studies.
3. Behavioral Outcomes in Stress and Exercise Models
In laboratory models, the effects of ELS combined with later exercise experience vary based on the specific experimental parameters (see Table 2). Though nuances are present in the literature, a frequent finding suggests that rats with a history of ELS show increased anxiety- [52,53,58,60,62] and depressive-like [6,53,54,56,57,58,62] phenotypes, and these phenotypes are ameliorated by exercise exposure [36,51,52,53,54,56,57,58]. These outcomes are mostly illustrated in male rats exposed to MS, with information on female rodent outcomes and sex differences, as well as rats exposed to other models of early stress, severely lacking. However, there is some evidence that stress and exercise differentially affect sexes. For example, James and colleagues [52] showed that exercise ameliorates anxiety-like behaviors in male rats exposed to MS but worsens these behaviors in females exposed to MS. An outcome such as this underscores the importance of studying sex differences, including the effect of hormones on behavior and susceptibility to stress. Indeed recent studies have shown that estrogen can significantly impact the effect of trauma on the brain and susceptibility to psychiatric disorders [63,64].
Table 2.
Summary of Behavioral Findings in Currently Published ELS-Exercise Intervention Studies.
The effects of exercise are not always consistent on ameliorating ELS-phenotypes, with increases in anxiety behavior [49,52] or no effects [36] sometimes observed. Rats may use the wheels to facilitate an escape behavior [44], which could have consequences for anxiety behavior. Another factor contributing to the inconsistency in exercise effects reflects the use of a voluntary WR treatment verses an involuntary treadmill (TM) exercise treatment. For example, Sadeghi and colleagues [36] reported that WR exposure decreased depressive-like behavior in rats but that TM exposure had no effect. Outside of affecting any ELS outcomes, exercise did bolster behavioral performance on cognitive and memory tasks in several studies [49,61], as is a common finding in the exercise literature [65].
4. Epigenetics and Neurotrophins
One way our experiences can get under the skin to affect genes, including the Bdnf gene, is through epigenetic mechanisms. Epigenetics refers to modifications to DNA that affect gene expression without making changes to the genetic sequence. One form of epigenetic regulation is called DNA methylation, wherein a methyl group is added to the cytosine at a CG site (cytosine-guanine dinucleotide) on the DNA [66,67]. CG sites are highly potent surrounding the promotor regions of most genes, making them a prime target for gene regulation. Increased methylation at promoter regions typically leads to decreases in gene expression because methyl groups recruit repressor proteins, interact with chromatic structure, and inhibit transcription factors from binding [66,68].
As researchers look to understand how early life stress can have long-term behavioral consequences and how exercise can reprogram the brain to have neurotherapeutic effects, focus often turns to neurotrophins, especially BDNF (see Figure 2 for our theoretical framework). Neurotrophins are a family of proteins which induce the development, survival, and function of neurons [6,69,70]. BDNF’s neurotrophic actions are vital for brain development and plasticity, and BDNF exhibits activity-regulated release in the central nervous system [71,72,73]. BDNF is a neurotrophin important for neural development, neural plasticity, learning, memory, and synaptic plasticity later in life, especially within the hippocampus [74,75]. Typically, increased methylation of the Bdnf gene is associated with decreased expression of its genetic material [68]. Methylation at any of Bdnf’s nine promotor regions can lead to decreased transcription of total Bdnf mRNA [76]. Stress during neonatal development has the capacity to alter Bdnf methylation for the long haul [5,77,78], which is important as decreased BDNF protein expression is found in patients with neurogenerative diseases and neuropsychiatric disorders [79,80,81,82]. These data highlight Bdnf as an important genetic locus for studies investigating epigenetic-behavioral interactions.
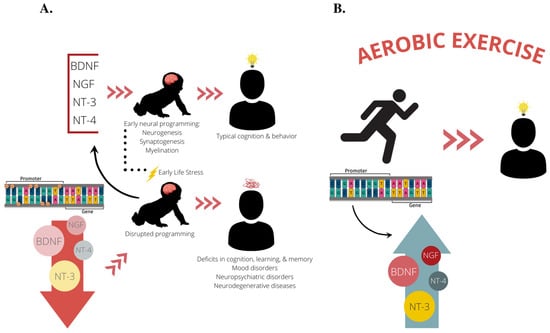
Figure 2.
Our proposed theoretical framework. (A) Based on previous studies, it is known that BDNF, NGF, NT-3, and NT-4 facilitate early neural programming of the brain by promoting neuronal survival and outgrowth during the perinatal period. Proper neural development during early life facilitates cognitive abilities and protects against neurobehavioral insults later in life, such as depression and anxiety disorders and neurodegenerative diseases. Experiencing early life stress can lead to dysregulation of these vital neurotrophins through epigenetic mechanisms such as DNA methylation, often leading a reduction in neurotrophin expression. This leads to disruptions in early neural programming that can stay with an individual throughout their lifespan and put them at an increased risk for developing deficits in cognition, learning, and memory, as well as neuropsychiatric disorders and neurodegenerative diseases. (B) We propose aerobic exercise as a treatment mechanism to normalize neurotrophin expression and bolster cognition and neural health by reversing epigenetic perturbations set forth in early life. BDNF, brain-derived neurotrophic factor; NGF, nerve growth factor; NT-3, neurotrophin-3; NT-4, neurotrophin-4.
5. Stress, Exercise and Neurotrophins
Many studies have shown that ELS reduces levels of both Bdnf mRNA and BDNF protein in multiple brain regions, including the prefrontal cortex and hippocampus [5,70,77,78,83,84,85]. Early life exposure to stress, especially within a caregiving environment, can result in a decrease in Bdnf gene expression through increased methylation of the Bdnf gene [5,86,87,88]. This impact of developmental stress extends to humans. For example, Bdnf DNA methylation correlates with the number of aversive childhood experiences in patients with bipolar disorder [89]. Further research in humans sheds light on the transgenerational effects of ELS on BDNF, in that babies born to mothers who experienced ELS show changes in Bdnf methylation and expression in blood cells obtained from the umbilical cord based on infant sex and degree of maternal fear [90].
While ELS generally decreases neurotrophin levels, aerobic exercise increases neurotrophin expression and is thought to be neuroprotective [17,91,92,93]. Exercise has positive impacts on neurotrophin expression, which directly impact neuronal survival and neurogenesis. Previous studies have identified exercise as a behavioral mechanism that specifically increases Bdnf expression [55,56,93] and decreases Bdnf methylation [94,95]. Several studies have also reported significant associations between exercise-induce BDNF upregulation and improved cognition [96] and depression symptoms [97].
The effect of exercise on neurotrophin expression in rodents exposed to ELS is understudied, with only 6 studies beginning to elucidate this relationship to date. Within these studies, exercise affected Bdnf/BDNF expression in a nonuniform and nuanced manner. Three studies showed that voluntary WR increased Bdnf mRNA in the hippocampus following 3hrs/daily MS during the first 2-3 weeks of life [48,55,59], and this increase was associated with rescued hippocampal neurogenesis in the dentate gyrus [48]. At the protein level, one study found that exercise increased BDNF expression in the striatum but not the ventral hippocampus compared to sedentary MS-exposed rats [56]. Given the complicated nature of Bdnf expression and gene regulation, Wearick-Silva and colleagues [61] investigated the exon-specific effects of exercise in the hippocampus. They reported that MS decreased Bdnf exon IV expression and increased exon IX expression, while exercise had an opposite effect on exon IX and increased Bdnf exon I expression. To shed light on the mechanism behind exercise-induced increases in Bdnf/BDNF expression, future studies should measure Bdnf exon-specific methylation in conjunction with expression to determine if specific genetic loci act in tandem to alter de novo expression.
Little is known regarding the effect of exercise on other neurotrophins in this ELS context. Marais and colleagues [56] produced the only current study investigating NT-3 and nerve growth factor (NGF) in this model, where they reported no significant effects on these neurotrophins in the ventral hippocampus and striatum. However, further investigation is warranted given that areas known to be highly impacted by stress and exercise, including the cerebellum, PFC, and dorsal hippocampus [98,99], have been overlooked.
6. Call to Action: Further Exploring ELS & Exercise
ELS alters long-term neurotrophin expression in the brain. These epigenetic changes contribute to an individual’s risk of numerous neurological, immune, and psychiatric disorders. We propose use of aerobic exercise as a treatment mechanism in future studies to understand the capacity of exercise to bolster the brain and body against ELS-induced disruption of biological processes. Support for our line of thinking comes from studies showing that exercise improves spatial memory, autoimmune and neurodegenerative disease symptomatology, muscle function, and gut health with concomitant changes in various neurotrophin expression levels.
Exercise has become a popular research area in neuroprotective research fields for its promising effects on brain health. A 2017 study following the natural aging of older adults (for ~10 years) showed that exercise is positively related to total cerebral and hippocampal volumes, and negatively related to developing Alzheimer’s Disease and dementia [100]. This suggests exercise is neural protective against neuronal and glial cell loss through the lifespan. It’s widely accepted that exercise has a positive effect on BDNF expression and epigenetic regulation [94,101], however, the data on other neurotrophins is much more nuanced and understudied [102]. Currently, only two studies have investigated the effect of exercise on NT-3 expression in humans [103,104]. A 2021 cutting-edge study reported that 12 weeks of high-intensity interval training increased serum levels of BDNF, NGF, NT-3, and NT-4 in elderly, obese, Chinese subjects [104]. Importantly, future research must critically examine exercise and participant parameters as these variables seem to be important when comparing participant outcomes. For example, in a 2018 clinical trial of adult obese males, high-intensity interval training (HIIT) had no effect on blood BDNF, NT-3, or NT-4 [103]. However, resistance training increased NT-3 and NT-4 levels, and combined exercise (resistance training plus HIIT) increased NT-3 and BDNF levels. In the combined exercise group, BDNF and NT-3 levels were positively correlated [103]. Taken together, these studies underscore the importance of continued research at the intersection of exercise science and neuroscience.
Further research indicates that aerobic exercise may be a valuable treatment mechanism for neurodevelopmental, autoimmune and psychological disorders. Aerobic exercise is a popular intervention strategy in studies investigating the molecular and cognitive effects of fetal alcohol spectrum disorders, with exercise exposure ameliorating Bdnf dysregulation [94], corpus callosum volume deficits [105], and executive functioning [106]. Recent work also demonstrates that aerobic exercise decreases the pathogenesis of multiple sclerosis [107], while increasing peripherally circulating BDNF and NGF [108]. Moderate physical exercise for 6 weeks is sufficient to reduce self-reported depression levels and increase peripheral BDNF and NGF levels in postmenopausal woman [109]. The therapeutic benefits of exercise on disease states underline a promising future for exercise intervention models.
Author Contributions
Conceptualization, T.S.C.; literature review; T.S.C., U.G., K.M.D., C.M.N.; writing—original draft preparation, T.S.C., K.M.D. and U.G.; writing—review and editing, T.S.C. and T.L.R.; figure design, T.S.C. and U.G.; supervision, T.S.C. and T.L.R.; funding acquisition, T.L.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by The Eunice Kennedy Shriver National Institute of Child Health and Human Development (NICHD); 1R01HD087509-01.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hoeijmakers, L.; Lucassen, P.J.; Korosi, A. The Interplay of Early-Life Stress, Nutrition, and Immune Activation Programs Adult Hippocampal Structure and Function. Front. Mol. Neurosci. 2015, 7, 103. [Google Scholar] [CrossRef] [PubMed]
- Andersen, S.L. Trajectories of Brain Development: Point of Vulnerability or Window of Opportunity? Neurosci. Biobehav. Rev. 2003, 27, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Burns, S.B.; Szyszkowicz, J.K.; Luheshi, G.N.; Lutz, P.E.; Turecki, G. Plasticity of the Epigenome during Early-Life Stress. Semin. Cell Dev. Biol. 2018, 77, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Milbocker, K.A.; Campbell, T.S.; Collins, N.; Kim, S.H.; Smith, I.F.; Roth, T.L.; Klintsova, A.Y. Glia-Driven Brain Circuit Refinement is Altered by Early-Life Adversity: Behavioral Outcomes. Front. Behav. Neurosci. 2021, 15. [Google Scholar] [CrossRef] [PubMed]
- Roth, T.L.; Lubin, F.D.; Funk, A.J.; Sweatt, J.D. Lasting Epigenetic Influence of Early-Life Adversity on the BDNF Gene. Biol. Psychiatry 2009, 65, 760–769. [Google Scholar] [CrossRef] [PubMed]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in Neuronal Development and Function. Annu. Rev. Neurosci. 2001, 24, 677. [Google Scholar] [CrossRef]
- Duman, R.S. Role of Neurotrophic Factors in the Etiology and Treatment of Mood Disorders. NeuroMolecular Med. 2004, 5, 11–25. [Google Scholar] [CrossRef]
- Mitchelmore, C.; Gede, L. Brain Derived Neurotrophic Factor: Epigenetic Regulation in Psychiatric Disorders. Brain Res. 2014, 1586, 162–172. [Google Scholar] [CrossRef]
- Woo, N.H.; Lu, B. Regulation of Cortical Interneurons by Neurotrophins: From Development to Cognitive Disorders. Neuroscientist 2006, 12, 43–56. [Google Scholar] [CrossRef]
- Tessarollo, L. Pleiotropic Functions of Neurotrophins in Development. Cytokine Growth Factor Rev. 1998, 9, 125–137. [Google Scholar] [CrossRef]
- Oliveira, S.L.B.; Pillat, M.M.; Cheffer, A.; Lameu, C.; Schwindt, T.T.; Ulrich, H. Functions of Neurotrophins and Growth Factors in Neurogenesis and Brain Repair. Cytom. Part A 2013, 83, 76–89. [Google Scholar] [CrossRef]
- Blum, R.; Konnerth, A. Neurotrophin-Mediated Rapid Signaling in the Central Nervous System: Mechanisms and Functions. Physiology 2005, 20, 70–78. [Google Scholar] [CrossRef]
- Bath, K.G.; Schilit, A.; Lee, F.S. Stress Effects on BDNF Expression: Effects of Age, Sex, and Form of Stress. Neuroscience 2013, 239, 149–156. [Google Scholar] [CrossRef]
- Miao, Z.; Wang, Y.; Sun, Z. The Relationships between Stress, Mental Disorders, and Epigenetic Regulation of BDNF. Int. J. Mol. Sci. 2020, 21, 1375. [Google Scholar] [CrossRef]
- Alleva, E.; Santucci, D. Psychosocial vs. “Physical” Stress Situations in Rodents and Humans: Role of Neurotrophins. Physiol. Behav. 2001, 73, 313–320. [Google Scholar] [CrossRef]
- Smith, M.A.; Makino, S.; Kvetnansky, R.; Post, R.M. Stress and Glucocorticoids Affect the Expression of Brain-Derived Neurotrophic Factor and Neurotrophin-3 MRNAs in the Hippocampus. J. Neurosci. 1995, 15, 1768–1777. [Google Scholar] [CrossRef]
- Afzalpour, M.E.; Chadorneshin, H.T.; Foadoddini, M.; Eivari, H.A. Comparing Interval and Continuous Exercise Training Regimens on Neurotrophic Factors in Rat Brain. Physiol. Behav. 2015, 147, 78–83. [Google Scholar] [CrossRef]
- Duman, R.S. Neurotrophic Factors and Regulation of Mood: Role of Exercise, Diet and Metabolism. Neurobiol. Aging 2005, 26, 88–93. [Google Scholar] [CrossRef]
- Baek, S.-S. Role of Exercise on the Brain. J. Exerc. Rehabil. 2016, 12, 380. [Google Scholar] [CrossRef]
- Szuhany, K.L.; Bugatti, M.; Otto, M.W. A Meta-Analytic Review of the Effects of Exercise on Brain-Derived Neurotrophic Factor. J. Psychiatr. Res. 2015, 60, 56. [Google Scholar] [CrossRef]
- Swain, R.A.; Berggren, K.L.; Kerr, A.L.; Patel, A.; Peplinski, C.; Sikorski, A.M. On Aerobic Exercise and Behavioral and Neural Plasticity. Brain Sci. 2012, 2, 709–744. [Google Scholar] [CrossRef]
- Grońska-Pęski, M.; Gonçalves, J.T.; Hébert, J.M. Enriched Environment Promotes Adult Hippocampal Neurogenesis through FGFRs. J. Neurosci. 2021, 41, 2899–2910. [Google Scholar] [CrossRef]
- Eriksson, P.S.; Perfilieva, E.; Björk-Eriksson, T.; Alborn, A.M.; Nordborg, C.; Peterson, D.A.; Gage, F.H. Neurogenesis in the Adult Human Hippocampus. Nat. Med. 1998, 4, 1313–1317. [Google Scholar] [CrossRef]
- Manni, L.; Micera, A.; Pistillo, L.; Aloe, L. Neonatal Handling in Eae-Susceptible Rats Alters NGF levels and Mast Cell Distribution in the Brain. Int. J. Dev. Neurosci. 1998, 16, 1–8. [Google Scholar] [CrossRef]
- Fabricius, K.; Wörtwein, G.; Pakkenberg, B. The Impact of Maternal Separation on Adult Mouse Behaviour and on the Total Neuron Number in the Mouse Hippocampus. Brain Struct. Funct. 2008, 212, 403–416. [Google Scholar] [CrossRef]
- Marais, L.; van Rensburg, S.J.; van Zyl, J.M.; Stein, D.J.; Daniels, W.M.U. Maternal Separation of Rat Pups Increases the Risk of Developing Depressive-like Behavior after Subsequent Chronic Stress by Altering Corticosterone and Neurotrophin Levels in the Hippocampus. Neurosci. Res. 2008, 61, 106–112. [Google Scholar] [CrossRef]
- Daniels, W.M.U.; Marais, L.; Stein, D.J.; Russell, V.A. Exercise Normalizes Altered Expression of Proteins in the Ventral Hippocampus of Rats Subjected to Maternal Separation. Exp. Physiol. 2012, 97, 239–247. [Google Scholar] [CrossRef]
- Riem, M.M.E.; Alink, L.R.A.; Out, D.; Van Ijzendoorn, M.H.; Bakermans-Kranenburg, M.J. Beating the Brain about Abuse: Empirical and Meta-Analytic Studies of the Association between Maltreatment and Hippocampal Volume across Childhood and Adolescence. Dev. Psychopathol. 2015, 27, 507–520. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, C.; Torres, H.; Harcourt, S. The Neurological and Neuropsychological Effects of Child Maltreatment. Aggress. Violent Behav. 2020, 54, 101408. [Google Scholar] [CrossRef]
- Hart, H.; Rubia, K.; Heinze, H.-J.; Holt, D.J. Neuroimaging of Child Abuse: A Critical Review. Front. Hum. Neurosci. 2012, 6, 52. [Google Scholar] [CrossRef]
- Lange, I.; Goossens, L.; Bakker, J.; Michielse, S.; Van Winkel, R.; Lissek, S.; Leibold, N.; Marcelis, M.; Wichers, M.; Van Os, J.; et al. Neurobehavioural Mechanisms of Threat Generalization Moderate the Link between Childhood Maltreatment and Psychopathology in Emerging Adulthood. J. Psychiatry Neurosci. 2019, 44, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Bremner, J.D.; Vythilingam, M.; Vermetten, E.; Southwick, S.M.; McGlashan, T.; Nazeer, A.; Khan, S.; Vaccarino, L.V.; Soufer, R.; Garg, P.K.; et al. MRI and PET Study of Deficits in Hippocampal Structure and Function in Women with Childhood Sexual Abuse and Posttraumatic Stress Disorder. Am. J. Psychiatry 2003, 160, 924–932. [Google Scholar] [CrossRef]
- Walker, C.-D.; Bath, K.G.; Joels, M.; Korosi, A.; Larauche, M.; Lucassen, P.J.; Morris, M.J.; Raineki, C.; Roth, T.L.; Sullivan, R.M.; et al. Chronic Early Life Stress Induced by Limited Bedding and Nesting (LBN) Material in Rodents: Critical Considerations of Methodology, Outcomes and Translational Potential. Stress 2017, 20, 421–448. [Google Scholar] [CrossRef] [PubMed]
- Ivy, A.S.; Brunson, K.L.; Sandman, C.; Baram, T.Z. Dysfunctional Nurturing Behavior in Rat Dams with Limited Access to Nesting Material: A Clinically Relevant Model for Early-Life Stress. Neuroscience 2008, 154, 1132–1142. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Levine, J.L.S.; Avila-Quintero, V.; Bloch, M.; Kaffman, A. Systematic Review and Meta-Analysis: Effects of Maternal Separation on Anxiety-like Behavior in Rodents. Transl. Psychiatry 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, M.; Peeri, M.; Hosseini, M.J. Adolescent Voluntary Exercise Attenuated Hippocampal Innate Immunity Responses and Depressive-like Behaviors Following Maternal Separation Stress in Male Rats. Physiol. Behav. 2016, 163, 177–183. [Google Scholar] [CrossRef]
- Ke, Z.; Yip, S.P.; Li, L.; Zheng, X.-X.; Tong, K.-Y. The Effects of Voluntary, Involuntary, and Forced Exercises on Brain-Derived Neurotrophic Factor and Motor Function Recovery: A Rat Brain Ischemia Model. PLoS ONE 2011, 6, e16643. [Google Scholar] [CrossRef]
- Svensson, M.; Rosvall, P.; Boza-Serrano, A.; Andersson, E.; Lexell, J.; Deierborg, T. Forced Treadmill Exercise Can Induce Stress and Increase Neuronal Damage in a Mouse Model of Global Cerebral Ischemia. Neurobiol. Stress 2016, 5, 8–18. [Google Scholar] [CrossRef]
- Uysal, N.; Kiray, M.; Sisman, A.; Camsari, U.; Gencoglu, C.; Baykara, B. Biotechnic & Histochemistry Effects of Voluntary and Involuntary Exercise on Cognitive Functions, and VEGF and BDNF Levels in Adolescent Rats. Biotech. Histochem. 2014, 90, 55–68. [Google Scholar] [CrossRef]
- Greenwood, B.N.; Foley, T.E.; Burhans, D.; Maier, S.F.; Fleshner, M. The Consequences of Uncontrollable Stress Are Sensitive to Duration of Prior Wheel Running. Brain Res. 2005, 1033, 164–178. [Google Scholar] [CrossRef]
- Greenwood, B.N.; Foley, T.E.; Le, T.V.; Strong, P.V.; Loughridge, A.B.; Day, H.E.W.; Fleshner, M. Long-Term Voluntary Wheel Running Is Rewarding and Produces Plasticity in the Mesolimbic Reward Pathway. Behav. Brain Res. 2011, 217, 354–362. [Google Scholar] [CrossRef]
- Sasse, S.K.; Greenwood, B.N.; Masini, C.V.; Nyhuis, T.J.; Fleshner, M.; Day, H.E.W.; Campeau, S.; Masini, C. Chronic Voluntary Wheel Running Facilitates Corticosterone Response Habituation to Repeated Audiogenic Stress Exposure in Male Rats: Original Research Report. Stress 2008, 11, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Abbink, M.R.; Naninck, E.F.G.; Lucassen, P.J.; Korosi, A. Early-life Stress Diminishes the Increase in Neurogenesis after Exercise in Adult Female Mice. Hippocampus 2017, 27, 839–844. [Google Scholar] [CrossRef]
- Botha, M.; Grace, L.; Bugarith, K.; Russell, V.A.; Kidd, M.; Seedat, S.; Hemmings, S.M. The Impact of Voluntary Exercise on Relative Telomere Length in a Rat Model of Developmental Stress. BMC Res. Notes 2012, 5, 1–7. [Google Scholar] [CrossRef]
- Dimatelis, J.J.; Hendricks, S.; Hsieh, J.; Vlok, N.M.; Bugarith, K.; Daniels, W.M.U.; Russell, V.A. Exercise Partly Reverses the Effect of Maternal Separation on Hippocampal Proteins in 6-hydroxydopamine-lesioned Rat Brain. Wiley Online Libr. 2013, 98, 233–244. [Google Scholar] [CrossRef]
- Eller, O.; Morris, E.; Thyfault, J.; Christianson, J.A. Early Life Stress Reduces Voluntary Exercise and Its Prevention of Diet-Induced Obesity and Metabolic Dysfunction in Mice. Physiol. Behav. 2020, 223, 113000. [Google Scholar]
- Eller, O.C.; Yang, X.; Fuentes, I.M.; Pierce, A.N.; Jones, B.M.; Brake, A.D.; Wang, R.; Dussor, G.; Christianson, J.A. Voluntary Wheel Running Partially Attenuates Early Life Stress-Induced Neuroimmune Measures in the Dura and Evoked Migraine-Like Behaviors in Female Mice. Front. Physiol. 2021, 12, 665732. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, I.M.; Jones, B.M.; Brake, A.D.; Pierce, A.N.; Eller, O.C.; Supple, R.M.; Wright, D.E.; Christianson, J.A. Voluntary Wheel Running Improves Outcomes in an Early Life Stress-Induced Model of Urologic Chronic Pelvic Pain Syndrome in Male Mice. Pain 2021, 162, 1681–1691. [Google Scholar] [CrossRef]
- Grace, L.; Hescham, S.; Kellaway, L.A.; Bugarith, K.; Russell, V.A. Effect of Exercise on Learning and Memory in a Rat Model of Developmental Stress. Metab. Brain Dis. 2009, 24, 643–657. [Google Scholar] [CrossRef] [PubMed]
- Hendricks, S.; Ojuka, E.; Kellaway, L.A.; Mabandla, M.V.; Russell, V.A. Effect of Maternal Separation on Mitochondrial Function and Role of Exercise in a Rat Model of Parkinson’s Disease. Metab. Brain Dis. 2012, 27, 387–392. [Google Scholar] [CrossRef]
- Huang, T.Y.; Lin, C.H. A Comparison between Chronic Exercise Training and Desipramine as Treatments for the Depression-like Behavior of Early-Life Maternal Deprivation Rats. Neurosci. Lett. 2010, 480, 201–205. [Google Scholar] [CrossRef] [PubMed]
- James, M.H.; Campbell, E.J.; Walker, F.R.; Smith, D.W.; Richardson, H.N.; Hodgson, D.M.; Dayas, C.V. Exercise Reverses the Effects of Early Life Stress on Orexin Cell Reactivity in Male but Not Female Rats. Front. Behav. Neurosci. 2014, 8, 244. [Google Scholar] [CrossRef] [PubMed]
- Khorjahani, A.; Peeri, M.; Azarbayjani, M.A. The Therapeutic Effect of Exercise on Anxiety and Bowel Oxidative Stress in the Maternal Separation Animal Model. Basic Clin. Neurosci. 2020, 11, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Makena, N.; Bugarith, K.; Russell, V.A. Maternal Separation Enhances Object Location Memory and Prevents Exercise-Induced MAPK/ERK Signalling in Adult Sprague-Dawley Rats. Metab. Brain Dis. 2012, 27, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Maniam, J.; Morris, M.J. Voluntary Exercise and Palatable High-Fat Diet Both Improve Behavioural Profile and Stress Responses in Male Rats Exposed to Early Life Stress: Role of Hippocampus. Psychoneuroendocrinology 2010, 35, 1553–1564. [Google Scholar] [CrossRef]
- Marais, L.; Stein, D.J.; Daniels, W.M.U. Exercise Increases BDNF Levels in the Striatum and Decreases Depressive-like Behavior in Chronically Stressed Rats. Metab. Brain Dis. 2009, 24, 587–597. [Google Scholar] [CrossRef]
- Masrour, F.F.; Peeri, M.; Azarbayjani, M.A.; Hosseini, M.J. Voluntary Exercise During Adolescence Mitigated Negative the Effects of Maternal Separation Stress on the Depressive-Like Behaviors of Adult Male Rats: Role of NMDA Receptors. Neurochem. Res. 2018, 43, 1067–1074. [Google Scholar] [CrossRef]
- Park, S.S.; Park, H.S.; Kim, C.J.; Baek, S.S.; Kim, T.W. Exercise Attenuates Maternal Separation-Induced Mood Disorder-like Behaviors by Enhancing Mitochondrial Functions and Neuroplasticity in the Dorsal Raphe. Behav. Brain Res. 2019, 372, 112049. [Google Scholar] [CrossRef]
- Pierce, A.N.; Eller-Smith, O.C.; Christianson, J.A. Voluntary Wheel Running Attenuates Urinary Bladder Hypersensitivity and Dysfunction Following Neonatal Maternal Separation in Female Mice. Neurourol. Urodyn. 2018, 37, 1623–1632. [Google Scholar] [CrossRef] [PubMed]
- Sahafi, E.; Peeri, M.; Hosseini, M.J.; Azarbyjani, M.A. Cardiac Oxidative Stress Following Maternal Separation Stress Was Mitigated Following Adolescent Voluntary Exercise in Adult Male Rat. Physiol. Behav. 2018, 183, 39–45. [Google Scholar] [CrossRef]
- Wearick-Silva, L.E.; Marshall, P.; Viola, T.W.; Centeno-Silva, A.; de Azeredo, L.A.; Orso, R.; Li, X.; Donadio, M.V.; Bredy, T.W.; Grassi-Oliveira, R. Running during Adolescence Rescues a Maternal Separation-Induced Memory Impairment in Female Mice: Potential Role of Differential Exon-Specific BDNF Expression. Dev. Psychobiol. 2017, 59, 268–274. [Google Scholar] [CrossRef]
- Zolfaghari, F.S.; Pirri, F.; Gauvin, E.; Peeri, M.; Amiri, S. Exercise and Fluoxetine Treatment during Adolescence Protect against Early Life Stress-Induced Behavioral Abnormalities in Adult Rats. Pharmacol. Biochem. Behav. 2021, 205, 173190. [Google Scholar] [CrossRef]
- Garza, K.; Jovanovic, T. Impact of Gender on Child and Adolescent PTSD. Curr. Psychiatry Rep. 2017, 19, 1–6. [Google Scholar] [CrossRef]
- Glover, E.M.; Jovanovic, T.; Norrholm, S.D. Estrogen and Extinction of Fear Memories: Implications for Posttraumatic Stress Disorder Treatment. Biol. Psychiatry 2015, 78, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Ludyga, S.; Gerber, M.; Pühse, U.; Looser, V.N.; Kamijo, K. Systematic Review and Meta-Analysis Investigating Moderators of Long-Term Effects of Exercise on Cognition in Healthy Individuals. Nat. Hum. Behav. 2020, 4, 603–612. [Google Scholar] [CrossRef]
- Roth, T.L. Epigenetic Mechanisms in the Development of Behavior: Advances, Challenges, and Future Promises of a New Field. Dev. Psychopathol. 2013, 25, 1279. [Google Scholar] [CrossRef] [PubMed]
- Dor, Y.; Cedar, H. Principles of DNA Methylation and Their Implications for Biology and Medicine. Lancet 2018, 392, 777–786. [Google Scholar] [CrossRef]
- Martinowich, K.; Hattori, D.; Wu, H.; Fouse, S.; He, F.; Hu, Y.; Fan, G.; Sun, Y.E. DNA Methylation-Related Chromatin Remodeling in Activity-Dependent Bdnf Gene Regulation. Science 2003, 302, 890–893. [Google Scholar] [CrossRef]
- Casey, B.J.; Glatt, C.E.; Tottenham, N.; Soliman, F.; Bath, K.; Amso, D.; Altemus, M.; Pattwell, S.; Jones, R.; Levita, L.; et al. Brain-Derived Neurotrophic Factor as a Model System for Examining Gene by Environment Interactions across Development. Neuroscience 2009, 164, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Fumagalli, F.; Molteni, R.; Racagni, G.; Riva, M.A. Stress during Development: Impact on Neuroplasticity and Relevance to Psychopathology. Prog. Neurobiol. 2007, 81, 197–217. [Google Scholar] [CrossRef] [PubMed]
- Conner, J.M.; Lauterborn, J.C.; Yan, Q.; Gall, C.M.; Varon, S. Distribution of Brain-Derived Neurotrophic Factor (BDNF) Protein and MRNA in the Normal Adult Rat CNS: Evidence for Anterograde Axonal Transport. J. Neurosci. 1997, 17, 2295–2313. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, M.E.; Xu, B.; Lu, B.; Hempstead, B.L. New Insights in the Biology of BDNF Synthesis and Release: Implications in CNS Function. J. Neurosci. 2009, 29, 12764–12767. [Google Scholar] [CrossRef] [PubMed]
- Hennigan, A.; O’Callaghan, R.M.; Kelly, Á.M. Neurotrophins and Their Receptors: Roles in Plasticity, Neurodegeneration and Neuroprotection. Biochem. Soc. Trans. 2007, 35, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Nie, Z.; Shu, H.; Kuang, Y.; Chen, X.; Cheng, J.; Yu, S.; Liu, H. The Role of BDNF on Neural Plasticity in Depression. Front. Cell. Neurosci. 2020, 14, 82. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Nagappan, G.; Lu, Y. BDNF and Synaptic Plasticity, Cognitive Function, and Dysfunction. Handb. Exp. Pharmacol. 2015, 220, 223–250. [Google Scholar] [CrossRef]
- Aid, T.; Kazantseva, A.; Piirsoo, M.; Palm, K.; Timmusk, T. Mouse and Rat BDNF Gene Structure and Expression Revisited. J. Neurosci. Res. 2007, 85, 525–535. [Google Scholar] [CrossRef]
- Blaze, J.; Roth, T.L. Caregiver Maltreatment Causes Altered Neuronal DNA Methylation in Female Rodents. Dev. Psychopathol. 2017, 29, 477–489. [Google Scholar] [CrossRef]
- Doherty, T.S.; Forster, A.; Roth, T.L. Global and Gene-Specific DNA Methylation Alterations in the Adolescent Amygdala and Hippocampus in an Animal Model of Caregiver Maltreatment. Behav. Brain Res. 2016, 298, 55–61. [Google Scholar] [CrossRef]
- Dwivedi, Y.; Rizavi, H.S.; Conley, R.R.; Roberts, R.C.; Tamminga, C.A.; Pandey, G.N. Altered Gene Expression of Brain-Derived Neurotrophic Factor and Receptor Tyrosine Kinase B in Postmortem Brain of Suicide Subjects. Arch. Gen. Psychiatry 2003, 60, 804–815. [Google Scholar] [CrossRef]
- Ginsberg, S.D.; Malek-Ahmadi, M.H.; Alldred, M.J.; Chen, Y.; Chen, K.; Chao, M.V.; Counts, S.E.; Mufson, E.J. Brain-Derived Neurotrophic Factor (BDNF) and TrkB Hippocampal Gene Expression Are Putative Predictors of Neuritic Plaque and Neurofibrillary Tangle Pathology. Neurobiol. Dis. 2019, 132, 104540. [Google Scholar] [CrossRef]
- Hashimoto, T.; Bergen, S.E.; Nguyen, Q.L.; Xu, B.; Monteggia, L.M.; Pierri, J.N.; Sun, Z.; Sampson, A.R.; Lewis, D.A. Relationship of Brain-Derived Neurotrophic Factor and Its Receptor TrkB to Altered Inhibitory Prefrontal Circuitry in Schizophrenia. J. Neurosci. 2005, 25, 372–383. [Google Scholar] [CrossRef] [PubMed]
- Ray, M.T.; Weickert, C.S.; Wyatt, E.; Webster, M.J. Decreased BDNF, TrkB-TK+ and GAD67 MRNA Expression in the Hippocampus of Individuals with Schizophrenia and Mood Disorders. J. Psychiatry Neurosci. 2011, 36, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, D.; Chatterjee-Chakraborty, M.; Rees, S.; Cauchi, J.; de Medeiros, C.B.; Fleming, A.S. Maternal Isolation Alters the Expression of Neural Proteins during Development: ‘Stroking’ Stimulation Reverses These Effects. Brain Res. 2007, 1158, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Choy, K.H.C.; De Visser, Y.; Nichols, N.R.; Van Den Buuse, M. Combined Neonatal Stress and Young-Adult Glucocorticoid Stimulation in Rats Reduce BDNF Expression in Hippocampus: Effects on Learning and Memory. Hippocampus 2008, 18, 655–667. [Google Scholar] [CrossRef]
- Lippmann, M.; Bress, A.; Nemeroff, C.B.; Plotsky, P.M.; Monteggia, L.M. Long-Term Behavioural and Molecular Alterations Associated with Maternal Separation in Rats. Eur. J. Neurosci. 2007, 25, 3091–3098. [Google Scholar] [CrossRef]
- Boersma, G.J.; Lee, R.S.; Cordner, Z.A.; Ewald, E.R.; Purcell, R.H.; Moghadam, A.A.; Tamashiro, K.L. Prenatal Stress Decreases Bdnf Expression and Increases Methylation of Bdnf Exon IV in Rats. Epigenetics 2014, 9, 437–447. [Google Scholar] [CrossRef]
- Fachim, H.A.; Corsi-Zuelli, F.; Loureiro, C.M.; Iamjan, S.A.; Shuhama, R.; Joca, S.; Menezes, P.R.; Heald, A.; Louzada-Junior, P.; Dalton, C.F.; et al. Early-Life Stress Effects on BDNF DNA Methylation in First-Episode Psychosis and in Rats Reared in Isolation. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 108, 110188. [Google Scholar] [CrossRef]
- Seo, M.K.; Ly, N.N.; Lee, C.H.; Cho, H.Y.; Choi, C.M.; Nhu, L.H.; Lee, J.G.; Lee, B.J.; Kim, G.M.; Yoon, B.J.; et al. Early Life Stress Increases Stress Vulnerability through BDNF Gene Epigenetic Changes in the Rat Hippocampus. Neuropharmacology 2016, 105, 388–397. [Google Scholar] [CrossRef]
- Perroud, N.; Salzmann, A.; Prada, P.; Nicastro, R.; Hoeppli, M.E.; Furrer, S.; Ardu, S.; Krejci, I.; Karege, F.; Malafosse, A. Response to Psychotherapy in Borderline Personality Disorder and Methylation Status of the BDNF Gene. Transl. Psychiatry 2013, 3, 207. [Google Scholar] [CrossRef]
- Pilkay, S.R.; Combs-Orme, T.; Tylavsky, F.; Bush, N.; Smith, A.K. Maternal Trauma and Fear History Predict BDNF Methylation and Gene Expression in Newborns. PeerJ 2020, 2020, e8858. [Google Scholar] [CrossRef]
- Cammisuli, D.M.; Bonuccelli, U.; Daniele, S.; Martini, C.; Fusi, J.; Franzoni, F. Aerobic Exercise and Healthy Nutrition as Neuroprotective Agents for Brain Health in Patients with Parkinson’s Disease: A Critical Review of the Literature. Antioxidants 2020, 9, 380. [Google Scholar] [CrossRef] [PubMed]
- Marley, C.J.; Brugniaux, J.V.; Davis, D.; Calverley, T.A.; Owens, T.S.; Stacey, B.S.; Tsukamoto, H.; Ogoh, S.; Ainslie, P.N.; Bailey, D.M. Long-Term Exercise Confers Equivalent Neuroprotection in Females Despite Lower Cardiorespiratory Fitness. Neuroscience 2020, 427, 58–63. [Google Scholar] [CrossRef]
- Håkansson, K.; Ledreux, A.; Daffner, K.; Terjestam, Y.; Bergman, P.; Carlsson, R.; Kivipelto, M.; Winblad, B.; Granholm, A.C.; Mohammed, A.K.H. BDNF Responses in Healthy Older Persons to 35 Minutes of Physical Exercise, Cognitive Training, and Mindfulness: Associations with Working Memory Function. J. Alzheimers Dis. 2017, 55, 645. [Google Scholar] [CrossRef] [PubMed]
- Boschen, K.E.; McKeown, S.E.; Roth, T.L.; Klintsova, A.Y. Impact of Exercise and a Complex Environment on Hippocampal Dendritic Morphology, Bdnf Gene Expression, and DNA Methylation in Male Rat Pups Neonatally Exposed to Alcohol. Dev. Neurobiol. 2017, 77, 708–725. [Google Scholar] [CrossRef] [PubMed]
- Voisey, J.; Lawford, B.; Bruenig, D.; Harvey, W.; Morris, C.P.; Young, R.M.D.; Mehta, D. Differential BDNF Methylation in Combat Exposed Veterans and the Association with Exercise. Gene 2019, 698, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Piepmeier, A.T.; Etnier, J.L. Brain-Derived Neurotrophic Factor (BDNF) as a Potential Mechanism of the Effects of Acute Exercise on Cognitive Performance. J. Sport Health Sci. 2015, 4, 14–23. [Google Scholar] [CrossRef]
- Murawska-Ciałowicz, E.; Wiatr, M.; Ciałowicz, M.; de Assis, G.G.; Borowicz, W.; Rocha-Rodrigues, S.; Paprocka-Borowicz, M.; Marques, A. BDNF Impact on Biological Markers of Depression—Role of Physical Exercise and Training. Int. J. Environ. Res. Public Health 2021, 18, 7553. [Google Scholar] [CrossRef]
- Teicher, M.H.; Samson, J.A. Annual Research Review: Enduring Neurobiological Effects of Childhood Abuse and Neglect. J. Child Psychol. Psychiatry 2016, 57, 241–266. [Google Scholar] [CrossRef]
- Walsh, N.D.; Dalgleish, T.; Lombardo, M.V.; Dunn, V.J.; Van Harmelen, A.L.; Ban, M.; Goodyer, I.M. General and Specific Effects of Early-Life Psychosocial Adversities on Adolescent Grey Matter Volume. NeuroImage Clin. 2014, 4, 308–318. [Google Scholar] [CrossRef]
- Tan, Z.S.; Spartano, N.L.; Beiser, A.S.; DeCarli, C.; Auerbach, S.H.; Vasan, R.S.; Seshadri, S. Physical Activity, Brain Volume, and Dementia Risk: The Framingham Study. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 789–795. [Google Scholar] [CrossRef]
- Cefis, M.; Prigent-Tessier, A.; Quirié, A.; Pernet, N.; Marie, C.; Garnier, P. The Effect of Exercise on Memory and BDNF Signaling Is Dependent on Intensity. Brain Struct. Funct. 2019, 224, 1975–1985. [Google Scholar] [CrossRef] [PubMed]
- Lippi, G.; Mattiuzzi, C.; Sanchis-Gomar, F. Updated Overview on Interplay between Physical Exercise, Neurotrophins, and Cognitive Function in Humans. J. Sport Health Sci. 2020, 9, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Sanchéz, M.A.; Bustos-Cruz, R.H.; Velasco-Orjuela, G.P.; Quintero, A.P.; Tordecilla-Sanders, A.; Correa-Bautista, J.E.; Triana-Reina, H.R.; García-Hermoso, A.; González-Ruíz, K.; Peña-Guzmán, C.A.; et al. Acute Effects of High Intensity, Resistance, or Combined Protocol on the Increase of Level of Neurotrophic Factors in Physically Inactive Overweight Adults: The BrainFit Study. Front. Physiol. 2018, 9, 741. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Han, T.; Zou, X.; Zhang, H.; Feng, W.; Wang, H.; Shen, Y.; Zhang, L.; Fang, G. Long-Term High-Intensity Interval Training Increases Serum Neurotrophic Factors in Elderly Overweight and Obese Chinese Adults. Eur. J. Appl. Physiol. 2021, 121, 2773–2785. [Google Scholar] [CrossRef] [PubMed]
- Milbocker, K.A.; LeBlanc, G.L.; Brengel, E.K.; Hekmatyar, K.S.; Kulkarni, P.; Ferris, C.F.; Klintsova, A.Y. Reduced and Delayed Myelination and Volume of Corpus Callosum in an Animal Model of Fetal Alcohol Spectrum Disorders Partially Benefit from Voluntary Exercise. Sci. Rep. 2022, 12, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Varigonda, A.L.; Edgcomb, J.B.; Zima, B.T. The Impact of Exercise in Improving Executive Function Impairments among Children and Adolescents with ADHD, Autism Spectrum Disorder, and Fetal Alcohol Spectrum Disorder: A Systematic Review and Meta-Analysis. Arch. Clin. Psychiatry 2021, 47, 146–156. [Google Scholar] [CrossRef]
- Motl, R.W. Exercise and Multiple Sclerosis. Adv. Exp. Med. Biol. 2020, 1228, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Bahmani, E.; Hoseini, R.; Amiri, E. The Compensatory Increased BDNF and NGF in Patients with Multiple Sclerosis Following Home-Based Aerobic Training and Vitamin D Supplementation during COVID-19 Outbreak. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Kim, B.; Kang, S. Regular Leisure-Time Physical Activity Is Effective in Boosting Neurotrophic Factors and Alleviating Menopause Symptoms. Int. J. Environ. Res. Public Health 2020, 17, 8624. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).