
Pectobacterium versatile Bacteriophage Possum: A Complex Polysaccharide-Deacetylating Tail Fiber as a Tool for Host Recognition in Pectobacterial Schitoviridae

 ,
,  ,
,  ,
,  , and
, and {kind=link}
{kind=link}
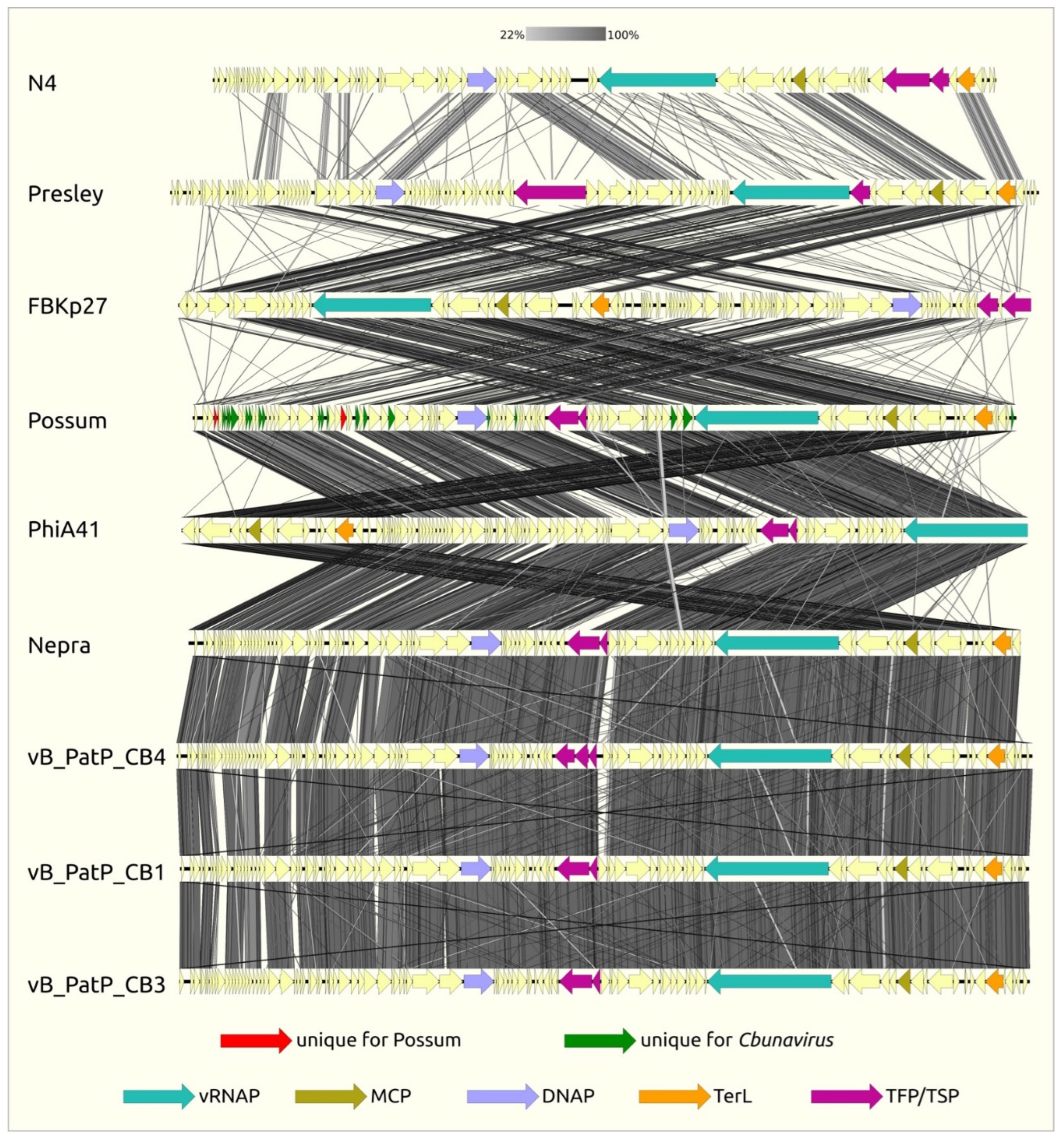{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. General Biological Features of Phages Horatius and Possum
2.2. Genomic Properties of Phages Horatius and Possum
2.3. Taxonomy and Phylogeny
2.4. Bioinformatic Analysis of the Adsorption Apparatus and Production of Recombinant Tail Fiber Protein
2.5. O-polysaccharide of Pectobacterium versatile F131 (VKM-3418)
2.6. Deacetylation of Bacterial O-polysaccharide by Tail Fiber Protein gp66
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Isolation and Purification of Phages Horatius and Possum
4.3. Electron Microscopy
4.4. Host Range and General Characterization of Phages Horatius and Possum
4.5. Genome Sequencing and Annotation
4.6. Phylogeny and Taxonomy Studies
4.7. Genome Comparison, Gene and Protein Analysis
4.8. Tail Fiber Protein Cloning, Expression, and Purification
4.9. Isolation and O-deacetylation of the O-polysaccharides
4.10. Sugar Analysis
4.11. NMR Spectroscopy
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pérombelon, M.C.M. Potato Diseases Caused by Soft Rot Erwinias: An Overview of Pathogenesis. Plant Pathol. 2002, 51, 1–12. [Google Scholar] [CrossRef]
- Gijsegem, F.; van Toth, I.K.; Wolf, J.M.; van der Wolf, J.M. Rot Pectobacteriaceae: A Brief Overview. In Plant Diseases Caused by Dickeya and Pectobacterium Species; Springer: Cham, Switzerland, 2021; pp. 1–11. ISBN 978-3-030-61458-4. [Google Scholar]
- Li, X.; Ma, Y.; Liang, S.; Tian, Y.; Yin, S.; Xie, S.; Xie, H. Comparative Genomics of 84 Pectobacterium Genomes Reveals the Variations Related to a Pathogenic Lifestyle. BMC Genom. 2018, 19, 889. [Google Scholar] [CrossRef] [PubMed]
- Toth, I.K.; Barny, M.; Czajkowski, R.; Elphinstone, J.G.; Li, X.; Pédron, J.; Pirhonen, M.; Van Gijsegem, F. Pectobacterium and Dickeya: Taxonomy and Evolution. In Plant Diseases Caused by Dickeya and Pectobacterium Species; Van Gijsegem, F., van der Wolf, J.M., Toth, I.K., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 13–37. ISBN 9783030614591. [Google Scholar]
- Zhang, Y.; Fan, Q.; Loria, R. A Re-Evaluation of the Taxonomy of Phytopathogenic Genera Dickeya and Pectobacterium Using Whole-Genome Sequencing Data. Syst. Appl. Microbiol. 2016, 39, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Evseev, P.; Ignatov, A.; Miroshnikov, K. Bioinformatic Basis to Define the Species Formation within Pectobacterium and Dickeya Bacterial Genera. In Proceedings of the 2020 Cognitive Sciences, Genomics and Bioinformatics (CSGB), Nososibirsk, Russia, 6–10 July 2020; pp. 47–52. [Google Scholar]
- Shirshikov, F.V.; Korzhenkov, A.A.; Miroshnikov, K.K.; Kabanova, A.P.; Barannik, A.P.; Ignatov, A.N.; Miroshnikov, K.A. Draft Genome Sequences of New Genomospecies “Candidatus Pectobacterium Maceratum” Strains, Which Cause Soft Rot in Plants. Genome Announc. 2018, 6, e00260–e00318. [Google Scholar] [CrossRef] [PubMed]
- Portier, P.; Pédron, J.; Taghouti, G.; Saux, M.F.-L.; Caullireau, E.; Bertrand, C.; Laurent, A.; Chawki, K.; Oulgazi, S.; Moumni, M.; et al. Elevation of Pectobacterium carotovorum Subsp. odoriferum to Species Level as Pectobacterium odoriferum Sp. Nov., Proposal of Pectobacterium brasiliense Sp. Nov. and Pectobacterium actinidiae Sp. Nov., Emended Description of Pectobacterium carotovorum and Description of Pectobacterium versatile Sp. Nov., Isolated from Streams and Symptoms on Diverse Plants. Int. J. Syst. Evol. Microbiol. 2019, 69, 3207–3216. [Google Scholar] [CrossRef]
- Curland, R.D.; Mainello, A.; Perry, K.L.; Hao, J.; Charkowski, A.O.; Bull, C.T.; McNally, R.R.; Johnson, S.B.; Rosenzweig, N.; Secor, G.A.; et al. Species of Dickeya and Pectobacterium Isolated during an Outbreak of Blackleg and Soft Rot of Potato in Northeastern and North Central United States. Microorganisms 2021, 9, 1733. [Google Scholar] [CrossRef]
- Han, W.; Wang, J.; Li, Z.; Pan, Y.; Zhang, D.; Zhao, D.; Li, Q.; Yang, Z.; Zhu, J. First Report of Pectobacterium versatile Causing Aerial Stem Rot of Potato in China. Plant Dis. 2022, 106, 756. [Google Scholar] [CrossRef]
- Jee, S.; Choi, J.-G.; Lee, Y.-G.; Kwon, M.; Hwang, I.; Heu, S. Distribution of Pectobacterium Species Isolated in South Korea and Comparison of Temperature Effects on Pathogenicity. Plant Pathol. J. 2020, 36, 346–354. [Google Scholar] [CrossRef]
- Ma, X.; Stodghill, P.; Gao, M.; Perry, K.L.; Swingle, B. Identification of Pectobacterium versatile Causing Blackleg of Potato in New York State. Plant Dis. 2021, 105, 2585–2594. [Google Scholar] [CrossRef]
- Ma, X.; Brazil, J.; Rivedal, H.; Perry, K.L.; Frost, K.; Swingle, B. First Report of Pectobacterium versatile Causing Potato Soft Rot in Oregon and Washington. Plant Dis. 2022, 106, 1292. [Google Scholar] [CrossRef]
- Oulghazi, S.; Moumni, M.; Khayi, S.; Robic, K.; Sarfraz, S.; Lopez-Roques, C.; Vandecasteele, C.; Faure, D. Diversity of Pectobacteriaceae Species in Potato Growing Regions in Northern Morocco. Microorganisms 2020, 8, 895. [Google Scholar] [CrossRef] [PubMed]
- Sarfraz, S.; Sahi, S.T.; Oulghazi, S.; Riaz, K.; Rajput, N.A.; Atiq, M.; Tufail, M.R.; Hameed, A.; Faure, D. Species Diversity of Dickeya and Pectobacterium Causing Potato Blackleg Disease in Pakistan. Plant Dis. 2020, 104, 1492–1499. [Google Scholar] [CrossRef] [PubMed]
- Portier, P.; Pédron, J.; Taghouti, G.; Dutrieux, C.; Barny, M.-A. Updated Taxonomy of Pectobacterium Genus in the CIRM-CFBP Bacterial Collection: When Newly Described Species Reveal “Old” Endemic Population. Microorganisms 2020, 8, 1141. [Google Scholar] [CrossRef] [PubMed]
- van der Wolf, J.M.; De Boer, S.H.; Czajkowski, R.; Cahill, G.; Van Gijsegem, F.; Davey, T.; Dupuis, B.; Ellicott, J.; Jafra, S.; Kooman, M.; et al. Management of Diseases Caused by Pectobacterium and Dickeya Species. In Plant Diseases Caused by Dickeya and Pectobacterium Species; Van Gijsegem, F., van der Wolf, J.M., Toth, I.K., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 175–214. ISBN 978-3-030-61459-1. [Google Scholar]
- Holtappels, D.; Fortuna, K.; Lavigne, R.; Wagemans, J. The Future of Phage Biocontrol in Integrated Plant Protection for Sustainable Crop Production. Curr. Opin. Biotechnol. 2021, 68, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Svircev, A.; Roach, D.; Castle, A. Framing the Future with Bacteriophages in Agriculture. Viruses 2018, 10, 218. [Google Scholar] [CrossRef]
- Casey, E.; van Sinderen, D.; Mahony, J. In Vitro Characteristics of Phages to Guide “Real Life” Phage Therapy Suitability. Viruses 2018, 10, 163. [Google Scholar] [CrossRef]
- Fernández, L.; Gutiérrez, D.; García, P.; Rodríguez, A. The Perfect Bacteriophage for Therapeutic Applications—A Quick Guide. Antibiotics 2019, 8, 126. [Google Scholar] [CrossRef]
- Harada, L.K.; Silva, E.C.; Campos, W.F.; Del Fiol, F.S.; Vila, M.; Dąbrowska, K.; Krylov, V.N.; Balcão, V.M. Biotechnological Applications of Bacteriophages: State of the Art. Microbiol. Res. 2018, 212–213, 38–58. [Google Scholar] [CrossRef]
- Buttimer, C.; Hendrix, H.; Lucid, A.; Neve, H.; Noben, J.; Franz, C.; O’Mahony, J.; Lavigne, R.; Coffey, A. Novel N4-Like Bacteriophages of Pectobacterium atrosepticum. Pharmaceuticals 2018, 11, 45. [Google Scholar] [CrossRef]
- Carstens, A.B.; Djurhuus, A.M.; Kot, W.; Hansen, L.H. A Novel Six-Phage Cocktail Reduces Pectobacterium atrosepticum Soft Rot Infection in Potato Tubers under Simulated Storage Conditions. FEMS Microbiol. Lett. 2019, 366, fnz101. [Google Scholar] [CrossRef]
- Zaczek-Moczydłowska, M.A.; Young, G.K.; Trudgett, J.; Plahe, C.; Fleming, C.C.; Campbell, K.; O’ Hanlon, R. Phage Cocktail Containing Podoviridae and Myoviridae Bacteriophages Inhibits the Growth of Pectobacterium Spp. under in Vitro and in Vivo Conditions. PLoS ONE 2020, 15, e0230842. [Google Scholar] [CrossRef] [PubMed]
- Voronina, M.V.; Bugaeva, E.N.; Vasiliev, D.M.; Kabanova, A.P.; Barannik, A.P.; Shneider, M.M.; Kulikov, E.E.; Korzhenkov, A.A.; Toschakov, S.V.; Ignatov, A.N.; et al. Characterization of Pectobacterium carotovorum Subsp. carotovorum Bacteriophage PP16 Prospective for Biocontrol of Potato Soft Rot. Microbiology 2019, 88, 451–460. [Google Scholar] [CrossRef]
- Lukianova, A.A.; Shneider, M.M.; Evseev, P.V.; Shpirt, A.M.; Bugaeva, E.N.; Kabanova, A.P.; Obraztsova, E.A.; Miroshnikov, K.K.; Senchenkova, S.N.; Shashkov, A.S.; et al. Morphologically Different Pectobacterium brasiliense Bacteriophages PP99 and PP101: Deacetylation of O-Polysaccharide by the Tail Spike Protein of Phage PP99 Accompanies the Infection. Front. Microbiol. 2020, 10, 3147. [Google Scholar] [CrossRef] [PubMed]
- Smolarska, A.; Rabalski, L.; Narajczyk, M.; Czajkowski, R. Isolation and Phenotypic and Morphological Characterization of the First Podoviridae Lytic Bacteriophages ΦA38 and ΦA41 Infecting Pectobacterium parmentieri (Former Pectobacterium wasabiae). Eur. J. Plant Pathol. 2018, 150, 413–425. [Google Scholar] [CrossRef]
- Gorbalenya, A.E.; Krupovic, M.; Mushegian, A.; Kropinski, A.M.; Siddell, S.G.; Varsani, A.; Adams, M.J.; Davison, A.J.; Dutilh, B.E.; Harrach, B.; et al. The New Scope of Virus Taxonomy: Partitioning the Virosphere into 15 Hierarchical Ranks. Nat. Microbiol. 2020, 5, 668–674. [Google Scholar] [CrossRef]
- Koonin, E.V.; Dolja, V.V.; Krupovic, M.; Varsani, A.; Wolf, Y.I.; Yutin, N.; Zerbini, F.M.; Kuhn, J.H. Global Organization and Proposed Megataxonomy of the Virus World. Microbiol. Mol. Biol. Rev. 2020, 84, e00061–e00119. [Google Scholar] [CrossRef]
- Wittmann, J.; Turner, D.; Millard, A.D.; Mahadevan, P.; Kropinski, A.M.; Adriaenssens, E.M. From Orphan Phage to a Proposed New Family–The Diversity of N4-Like Viruses. Antibiotics 2020, 9, 663. [Google Scholar] [CrossRef]
- Cairns, B.J.; Payne, R.J.H. Bacteriophage Therapy and the Mutant Selection Window. Antimicrob. Agents Chemother. 2008, 52, 4344. [Google Scholar] [CrossRef]
- Rothman-Denes, L.B.; Schito, G.C. Novel Transcribing Activities in N4-Infected Escherichia coli. Virology 1974, 60, 65–72. [Google Scholar] [CrossRef]
- Willis, S.H.; Kazmierczak, K.M.; Carter, R.H.; Rothman-Denes, L.B. N4 RNA Polymerase II, a Heterodimeric RNA Polymerase with Homology to the Single-Subunit Family of RNA Polymerases. J. Bacteriol. 2002, 184, 4952–4961. [Google Scholar] [CrossRef]
- Zivin, R.; Zehring, W.; Rothman-Denes, L.B. Transcriptional Map of Bacteriophage N4. Location and Polarity of N4 RNAs. J. Mol. Biol. 1981, 152, 335–356. [Google Scholar] [CrossRef]
- Drobysheva, A.V.; Panafidina, S.A.; Kolesnik, M.V.; Klimuk, E.I.; Minakhin, L.; Yakunina, M.V.; Borukhov, S.; Nilsson, E.; Holmfeldt, K.; Yutin, N.; et al. Structure and Function of Virion RNA Polymerase of a CrAss-like Phage. Nature 2021, 589, 306–309. [Google Scholar] [CrossRef]
- Lenneman, B.R.; Rothman-Denes, L.B. Structural and Biochemical Investigation of Bacteriophage N4-Encoded RNA Polymerases. Biomolecules 2015, 5, 647–667. [Google Scholar] [CrossRef] [PubMed]
- Carter, R.H.; Demidenko, A.A.; Hattingh-Willis, S.; Rothman-Denes, L.B. Phage N4 RNA Polymerase II Recruitment to DNA by a Single-Stranded DNA-Binding Protein. Genes Dev. 2003, 17, 2334–23445. [Google Scholar] [CrossRef] [PubMed]
- Weigele, P.; Raleigh, E.A. Biosynthesis and Function of Modified Bases in Bacteria and Their Viruses. Chem. Rev. 2016, 116, 12655–12687. [Google Scholar] [CrossRef]
- Lee, I.; Ouk Kim, Y.; Park, S.-C.; Chun, J. OrthoANI: An Improved Algorithm and Software for Calculating Average Nucleotide Identity. Int. J. Syst. Evol. Microbiol. 2016, 66, 1100–1103. [Google Scholar] [CrossRef]
- Moraru, C.; Varsani, A.; Kropinski, A.M. VIRIDIC—A Novel Tool to Calculate the Intergenomic Similarities of Prokaryote-Infecting Viruses. Viruses 2020, 12, 1268. [Google Scholar] [CrossRef]
- Heller, K.J. Identification of the Phage Gene for Host Receptor Specificity by Analyzing Hybrid Phages of T5 and BF23. Virology 1984, 139, 11–21. [Google Scholar] [CrossRef]
- Le, S.; He, X.; Tan, Y.; Huang, G.; Zhang, L.; Lux, R.; Shi, W.; Hu, F. Mapping the Tail Fiber as the Receptor Binding Protein Responsible for Differential Host Specificity of Pseudomonas aeruginosa Bacteriophages PaP1 and JG004. PLoS ONE 2013, 8, e68562. [Google Scholar] [CrossRef]
- Dupont, K.; Vogensen, F.K.; Neve, H.; Bresciani, J.; Josephsen, J. Identification of the Receptor-Binding Protein in 936-Species Lactococcal Bacteriophages. Appl. Environ. Microbiol. 2004, 70, 5818–5824. [Google Scholar] [CrossRef]
- Fokine, A.; Islam, M.Z.; Zhang, Z.; Bowman, V.D.; Rao, V.B.; Rossmann, M.G. Structure of the Three N-Terminal Immunoglobulin Domains of the Highly Immunogenic Outer Capsid Protein from a T4-Like Bacteriophage. J. Virol. 2011, 85, 8141–8148. [Google Scholar] [CrossRef] [PubMed]
- Cuervo, A.; Pulido-Cid, M.; Chagoyen, M.; Arranz, R.; González-García, V.A.; Garcia-Doval, C.; Castón, J.R.; Valpuesta, J.M.; van Raaij, M.J.; Martín-Benito, J.; et al. Structural Characterization of the Bacteriophage T7 Tail Machinery. J. Biol. Chem. 2013, 288, 26290–26299. [Google Scholar] [CrossRef] [PubMed]
- Latka, A.; Leiman, P.G.; Drulis-Kawa, Z.; Briers, Y. Modeling the Architecture of Depolymerase-Containing Receptor Binding Proteins in Klebsiella Phages. Front. Microbiol. 2019, 10, 2649. [Google Scholar] [CrossRef] [PubMed]
- Leiman, P.G.; Shneider, M.M. Contractile Tail Machines of Bacteriophages. Adv. Exp. Med. Biol. 2012, 726, 93–114. [Google Scholar] [CrossRef]
- Nobrega, F.L.; Vlot, M.; de Jonge, P.A.; Dreesens, L.L.; Beaumont, H.J.E.; Lavigne, R.; Dutilh, B.E.; Brouns, S.J.J. Targeting Mechanisms of Tailed Bacteriophages. Nat. Rev. Microbiol. 2018, 16, 760–773. [Google Scholar] [CrossRef]
- McPartland, J.; Rothman-Denes, L.B. The Tail Sheath of Bacteriophage N4 Interacts with the Escherichia coli Receptor. J. Bacteriol. 2009, 191, 525–532. [Google Scholar] [CrossRef]
- Choi, K.H.; McPartland, J.; Kaganman, I.; Bowman, V.D.; Rothman-Denes, L.B.; Rossmann, M.G. Insight into DNA and Protein Transport in Double-Stranded DNA Viruses: The Structure of Bacteriophage N4. J. Mol. Biol. 2008, 378, 726–736. [Google Scholar] [CrossRef]
- Zhang, Z.; Tian, C.; Zhao, J.; Chen, X.; Wei, X.; Li, H.; Lin, W.; Feng, R.; Jiang, A.; Yang, W.; et al. Characterization of Tail Sheath Protein of N4-Like Phage PhiAxp-3. Front. Microbiol. 2018, 9, 450. [Google Scholar] [CrossRef]
- Ma, Y.; Li, E.; Qi, Z.; Li, H.; Wei, X.; Lin, W.; Zhao, R.; Jiang, A.; Yang, H.; Yin, Z.; et al. Isolation and Molecular Characterisation of Achromobacter Phage PhiAxp-3, an N4-like Bacteriophage. Sci. Rep. 2016, 6, 24776. [Google Scholar] [CrossRef]
- Prokhorov, N.S.; Riccio, C.; Zdorovenko, E.L.; Shneider, M.M.; Browning, C.; Knirel, Y.A.; Leiman, P.G.; Letarov, A.V. Function of Bacteriophage G7C Esterase Tailspike in Host Cell Adsorption. Mol. Microbiol. 2017, 105, 385–398. [Google Scholar] [CrossRef]
- Bartnik, P.; Jafra, S.; Narajczyk, M.; Czaplewska, P.; Czajkowski, R. Pectobacterium parmentieri SCC 3193 Mutants with Altered Synthesis of Cell Surface Polysaccharides Are Resistant to N4-Like Lytic Bacteriophage ΦA38 (VB_Ppp_A38) but Express Decreased Virulence in Potato (Solanum tuberosum L.) Plants. Int. J. Mol. Sci. 2021, 22, 7346. [Google Scholar] [CrossRef] [PubMed]
- Knecht, L.E.; Heinrich, N.; Born, Y.; Felder, K.; Pelludat, C.; Loessner, M.J.; Fieseler, L. Bacteriophage S6 Requires Bacterial Cellulose for Erwinia amylovora Infection. Environ. Microbiol. 2022, 24, 3436–3450. [Google Scholar] [CrossRef] [PubMed]
- Römling, U. The Power of Unbiased Phenotypic Screens-Cellulose as a First Receptor for the Schitoviridae Phage S6 of Erwinia amylovora. Environ. Microbiol. 2022, 21, 3316–3321. [Google Scholar] [CrossRef] [PubMed]
- Plattner, M.; Shneider, M.M.; Arbatsky, N.P.; Shashkov, A.S.; Chizhov, A.O.; Nazarov, S.; Prokhorov, N.S.; Taylor, N.M.I.; Buth, S.A.; Gambino, M.; et al. Structure and Function of the Branched Receptor-Binding Complex of Bacteriophage CBA120. J. Mol. Biol. 2019, 431, 3718–3739. [Google Scholar] [CrossRef] [PubMed]
- Leiman, P.G.; Battisti, A.J.; Bowman, V.D.; Stummeyer, K.; Mühlenhoff, M.; Gerardy-Schahn, R.; Scholl, D.; Molineux, I.J. The Structures of Bacteriophages K1E and K1-5 Explain Processive Degradation of Polysaccharide Capsules and Evolution of New Host Specificities. J. Mol. Biol. 2007, 371, 836–849. [Google Scholar] [CrossRef]
- Taylor, N.M.I.; Prokhorov, N.S.; Guerrero-Ferreira, R.C.; Shneider, M.M.; Browning, C.; Goldie, K.N.; Stahlberg, H.; Leiman, P.G. Structure of the T4 Baseplate and Its Function in Triggering Sheath Contraction. Nature 2016, 533, 346–352. [Google Scholar] [CrossRef]
- Buth, S.A.; Shneider, M.M.; Scholl, D.; Leiman, P.G. Structure and Analysis of R1 and R2 Pyocin Receptor-Binding Fibers. Viruses 2018, 10, 427. [Google Scholar] [CrossRef]
- Olszak, T.; Shneider, M.M.; Latka, A.; Maciejewska, B.; Browning, C.; Sycheva, L.V.; Cornelissen, A.; Danis-Wlodarczyk, K.; Senchenkova, S.N.; Shashkov, A.S.; et al. The O-Specific Polysaccharide Lyase from the Phage LKA1 Tailspike Reduces Pseudomonas Virulence. Sci. Rep. 2017, 7, 16302. [Google Scholar] [CrossRef]
- Leonteln, K.; Lönngren, J. Determination of the Absolute Configuration of Sugars by Gas Liquid Chromatography of Their Acetylated 2-Octyl Glycosides. In Methods in Carbohydrate Chemistry; BeMiller, J.N., Whistler, R.L., Shaw, D.H., Eds.; John Wiley & Sons Inc.: New York, NY, USA, 1993; pp. 87–89. [Google Scholar]
- Shashkov, A.S.; Lipkind, G.M.; Knirel, Y.A.; Kochetkov, N.K. Stereochemical Factors Determining the Effects of Glycosylation on the 13C Chemical Shifts in Carbohydrates. Magn. Reson. Chem. 1988, 26, 735–747. [Google Scholar] [CrossRef]
- Lipkind, G.M.; Shashkov, A.S.; Knirel, Y.A.; Vinogradov, E.V.; Kochetkov, N.K. A Computer-Assisted Structural Analysis of Regular Polysaccharides on the Basis of 13C-n.m.r. Data. Carbohydr. Res. 1988, 175, 59–75. [Google Scholar] [CrossRef]
- Jansson, P.E.; Kenne, L.; Widmalm, G. Computer-Assisted Structural Analysis of Polysaccharides with an Extended Version of CASPER Using 1H- and 13C-n.m.r. Data. Carbohydr. Res. 1989, 188, 169–191. [Google Scholar] [CrossRef]
- Jansson, P.-E.; Kenne, L.; Schweda, E. Nuclear Magnetic Resonance and Conformational Studies on Monoacetylated Methyl D-Gluco- and D-Galacto-Pyranosides. J. Chem. Soc. Perkin. Trans. 1987, 10, 377–383. [Google Scholar] [CrossRef]
- Iyer, L.M.; Burroughs, A.M.; Anand, S.; de Souza, R.F.; Aravind, L. Polyvalent Proteins, a Pervasive Theme in the Intergenomic Biological Conflicts of Bacteriophages and Conjugative Elements. J. Bacteriol. 2017, 199, e00245–e00317. [Google Scholar] [CrossRef]
- Kazmierczak, K.M.; Davydova, E.K.; Mustaev, A.A.; Rothman-Denes, L.B. The Phage N4 Virion RNA Polymerase Catalytic Domain Is Related to Single-Subunit RNA Polymerases. EMBO J. 2002, 21, 5815–5823. [Google Scholar] [CrossRef]
- Chang, C.-Y.; Kemp, P.; Molineux, I.J. Gp15 and Gp16 Cooperate in Translocating Bacteriophage T7 DNA into the Infected Cell. Virology 2010, 398, 176. [Google Scholar] [CrossRef] [PubMed]
- Holtzman, T.; Globus, R.; Molshanski-Mor, S.; Ben-Shem, A.; Yosef, I.; Qimron, U. A Continuous Evolution System for Contracting the Host Range of Bacteriophage T7. Sci. Rep. 2020, 10, 307. [Google Scholar] [CrossRef]
- Bugaeva, E.N.; Voronina, M.V.; Vasiliev, D.M.; Lukianova, A.A.; Landyshev, N.N.; Ignatov, A.N.; Miroshnikov, K.A. Use of a Specific Phage Cocktail for Soft Rot Control on Ware Potatoes: A Case Study. Viruses 2021, 13, 1095. [Google Scholar] [CrossRef]
- Clokie, M.R.J.; Kropinski, A.M. Bacteriophages; Humana Press: Totowa, NJ, USA, 2009; ISBN 9781588296825. [Google Scholar]
- Ackermann, H.-W. Basic Phage Electron Microscopy. Bacteriophages 2009, 501, 113–126. [Google Scholar] [CrossRef]
- Czajkowski, R.; Ozymko, Z.; Lojkowska, E. Isolation and Characterization of Novel Soilborne Lytic Bacteriophages Infecting Dickeya Spp. Biovar 3 (‘D. Solani’). Plant Pathol. 2014, 63, 758–772. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. Available online: https://www.liebertpub.com/doi/10.1089/cmb.2012.0021 (accessed on 12 July 2022).
- Hyatt, D.; Chen, G.-L.; LoCascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic Gene Recognition and Translation Initiation Site Identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef]
- Besemer, J.; Lomsadze, A.; Borodovsky, M. GeneMarkS: A Self-Training Method for Prediction of Gene Starts in Microbial Genomes. Implications for Finding Sequence Motifs in Regulatory Regions. Nucleic Acids Res. 2001, 29, 2607. [Google Scholar] [CrossRef] [PubMed]
- Delcher, A.L.; Harmon, D.; Kasif, S.; White, O.; Salzberg, S.L. Improved Microbial Gene Identification with GLIMMER. Nucleic Acids Res. 1999, 27, 4636–4641. [Google Scholar] [CrossRef] [PubMed]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 Web Portal for Protein Modeling, Prediction and Analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.; Chang, H.-Y.; Daugherty, L.; Fraser, M.; Hunter, S.; Lopez, R.; McAnulla, C.; McMenamin, C.; Nuka, G.; Pesseat, S.; et al. The InterPro Protein Families Database: The Classification Resource after 15 Years. Nucleic Acids Res. 2015, 43, D213–D221. [Google Scholar] [CrossRef] [PubMed]
- Söding, J.; Biegert, A.; Lupas, A.N. The HHpred Interactive Server for Protein Homology Detection and Structure Prediction. Nucleic Acids Res. 2005, 33, W244–W248. [Google Scholar] [CrossRef] [PubMed]
- Schattner, P.; Brooks, A.N.; Lowe, T.M. The TRNAscan-SE, Snoscan and SnoGPS Web Servers for the Detection of TRNAs and SnoRNAs. Nucleic Acids Res. 2005, 33, W686–W689. [Google Scholar] [CrossRef]
- Laslett, D.; Canback, B. ARAGORN, a Program to Detect TRNA Genes and TmRNA Genes in Nucleotide Sequences. Nucleic Acids Res 2004, 32, 11–16. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A Novel Method for Rapid Multiple Sequence Alignment Based on Fast Fourier Transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Kumar, S.; Tamura, K.; Nei, M. MEGA: Molecular Evolutionary Genetics Analysis Software for Microcomputers. Comput. Appl. Biosci. 1994, 10, 189–191. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian Inference of Phylogenetic Trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian Phylogenetic Inference under Mixed Models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A Genome Comparison Visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef] [PubMed]
- Westphal, O. Bacterial Lipopolysaccharides: Extraction with Phenol-Water and Further Applications of the Procedure. Undefined 1965, 5, 83–91. [Google Scholar]
- Zych, K.; Toukach, F.V.; Arbatsky, N.P.; Kolodziejska, K.; Senchenkova, S.N.; Shashkov, A.S.; Knirel, Y.A.; Sidorczyk, Z. Structure of the O-Specific Polysaccharide of Proteus mirabilis D52 and Typing of This Strain to Proteus Serogroup O33. Eur. J. Biochem. 2001, 268, 4346–4351. [Google Scholar] [CrossRef] [PubMed]
- Sawardeker, J.S.; Sloneker, J.H.; Jeanes, A. Quantitative Determination of Monosaccharides as Their Alditol Acetates by Gas Liquid Chromatography. Anal. Chem. 1965, 37, 1602–1604. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lukianova, A.A.; Evseev, P.V.; Shneider, M.M.; Dvoryakova, E.A.; Tokmakova, A.D.; Shpirt, A.M.; Kabilov, M.R.; Obraztsova, E.A.; Shashkov, A.S.; Ignatov, A.N.; et al. Pectobacterium versatile Bacteriophage Possum: A Complex Polysaccharide-Deacetylating Tail Fiber as a Tool for Host Recognition in Pectobacterial Schitoviridae. Int. J. Mol. Sci. 2022, 23, 11043. https://doi.org/10.3390/ijms231911043
Lukianova AA, Evseev PV, Shneider MM, Dvoryakova EA, Tokmakova AD, Shpirt AM, Kabilov MR, Obraztsova EA, Shashkov AS, Ignatov AN, et al. Pectobacterium versatile Bacteriophage Possum: A Complex Polysaccharide-Deacetylating Tail Fiber as a Tool for Host Recognition in Pectobacterial Schitoviridae. International Journal of Molecular Sciences. 2022; 23(19):11043. https://doi.org/10.3390/ijms231911043
Chicago/Turabian StyleLukianova, Anna A., Peter V. Evseev, Mikhail M. Shneider, Elena A. Dvoryakova, Anna D. Tokmakova, Anna M. Shpirt, Marsel R. Kabilov, Ekaterina A. Obraztsova, Alexander S. Shashkov, Alexander N. Ignatov, and et al. 2022. "Pectobacterium versatile Bacteriophage Possum: A Complex Polysaccharide-Deacetylating Tail Fiber as a Tool for Host Recognition in Pectobacterial Schitoviridae" International Journal of Molecular Sciences 23, no. 19: 11043. https://doi.org/10.3390/ijms231911043
APA StyleLukianova, A. A., Evseev, P. V., Shneider, M. M., Dvoryakova, E. A., Tokmakova, A. D., Shpirt, A. M., Kabilov, M. R., Obraztsova, E. A., Shashkov, A. S., Ignatov, A. N., Knirel, Y. A., Dzhalilov, F. S.-U., & Miroshnikov, K. A. (2022). Pectobacterium versatile Bacteriophage Possum: A Complex Polysaccharide-Deacetylating Tail Fiber as a Tool for Host Recognition in Pectobacterial Schitoviridae. International Journal of Molecular Sciences, 23(19), 11043. https://doi.org/10.3390/ijms231911043