
Caffeine-Induced Acute and Delayed Responses in Cerebral Metabolism of Control and Schizophrenia-like Wisket Rats

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
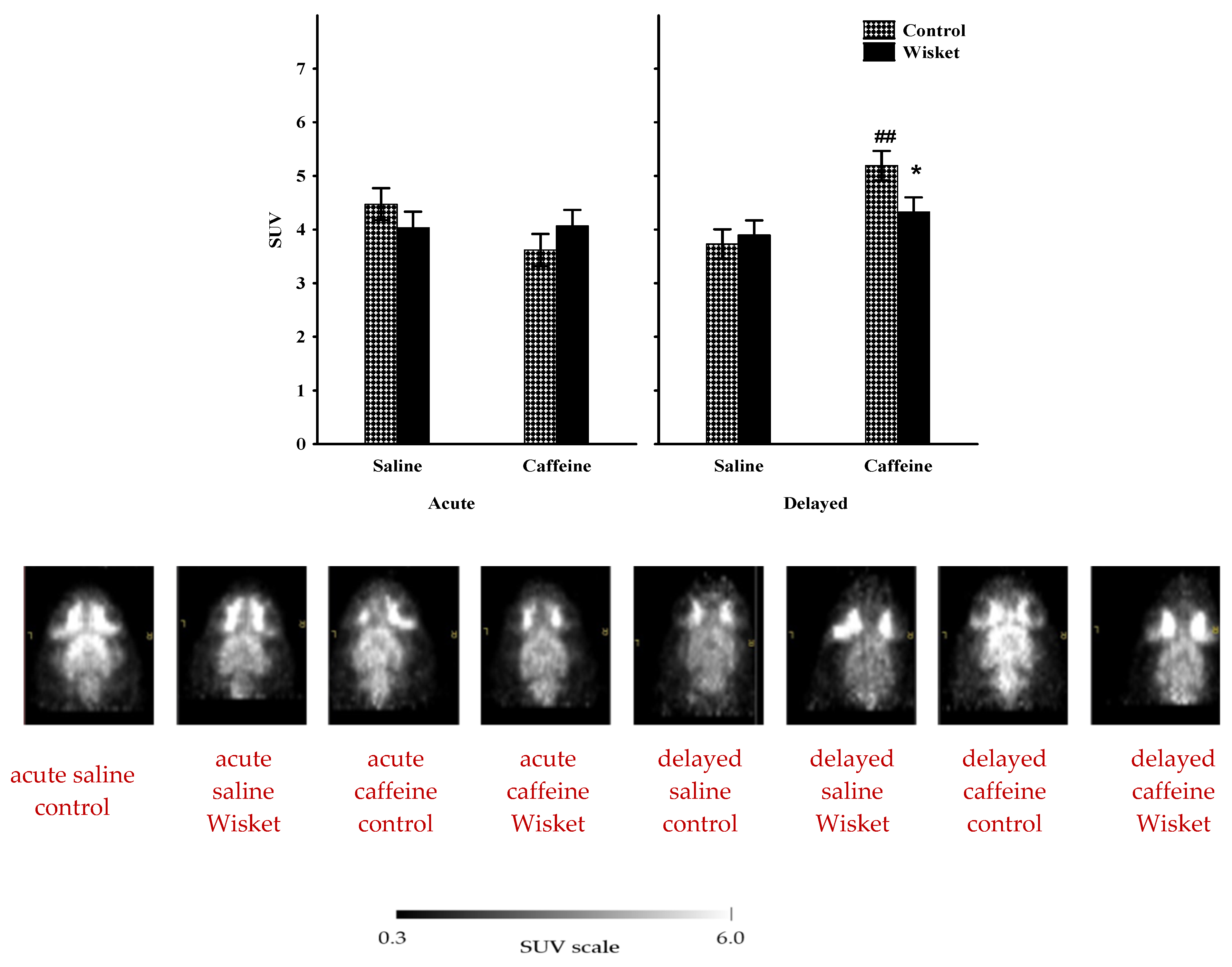
2.1. Whole-Brain Metabolic Activity
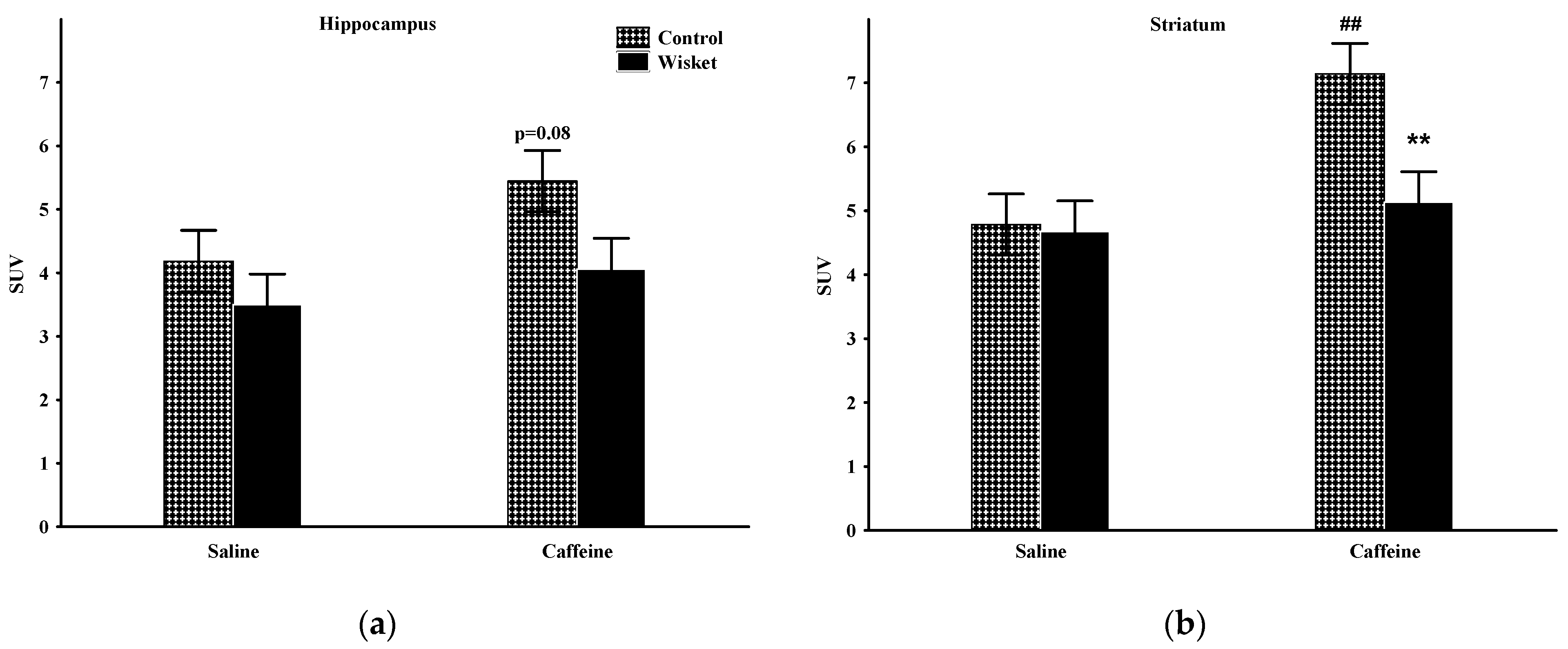
2.2. Hippocampus
2.3. Striatum
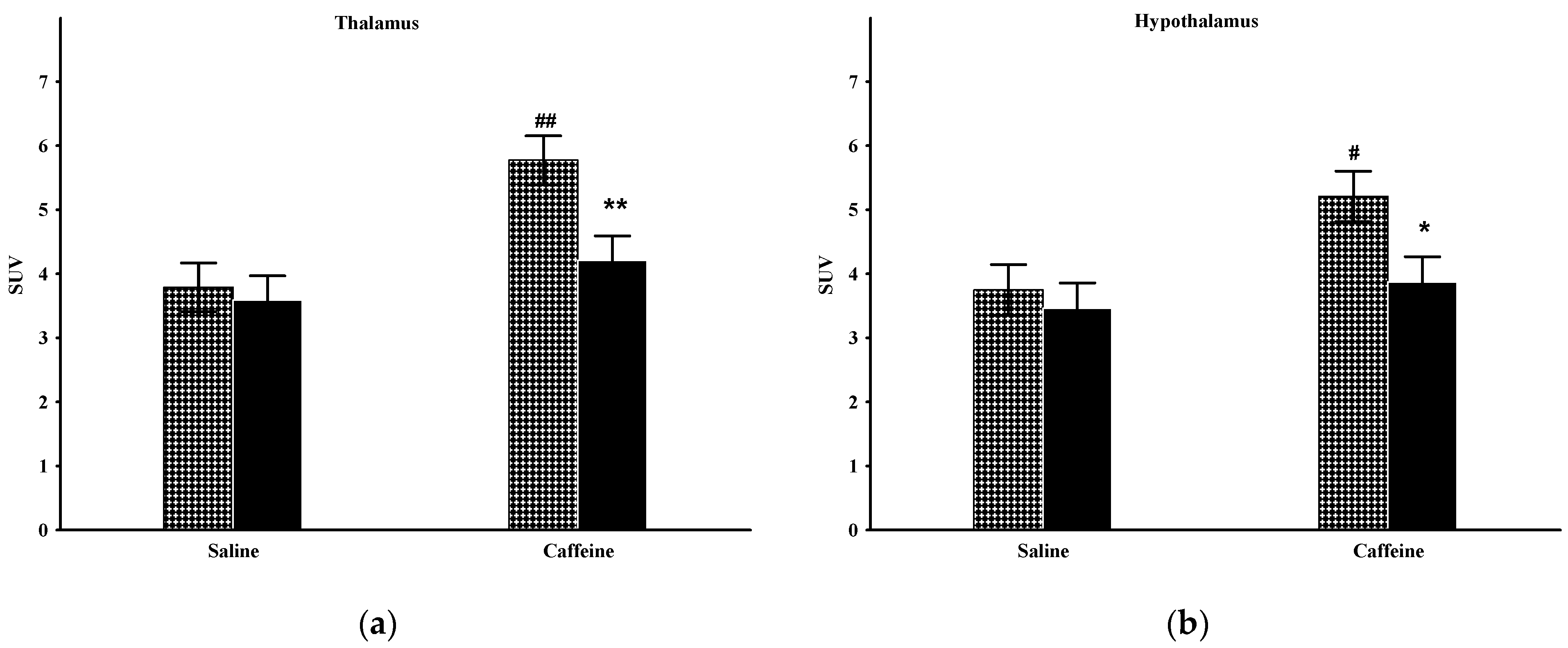
2.4. Thalamus
2.5. Hypothalamus
3. Discussion
4. Materials and Methods
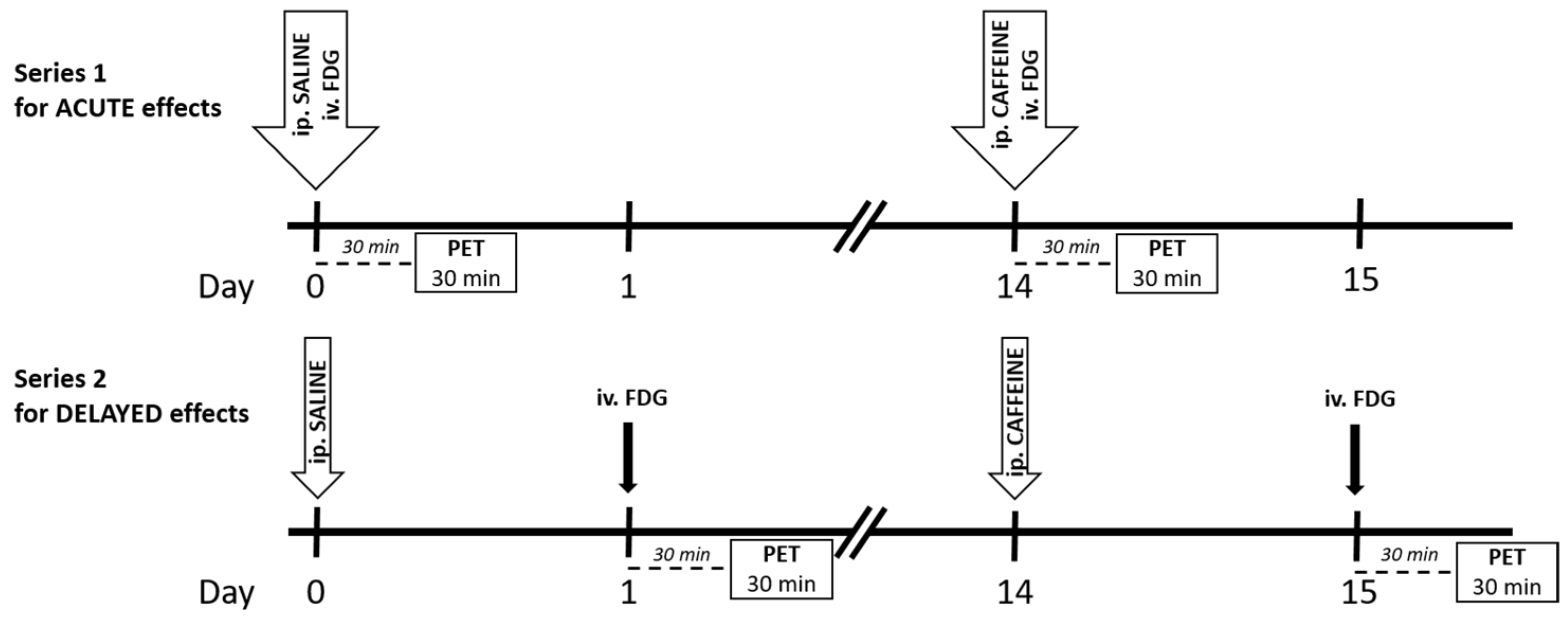
4.1. Experimental Paradigm
4.2. In Vivo PET Imaging
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Bralet, M.C.; Buchsbaum, M.S.; DeCastro, A.; Shihabuddin, L.; Mitelman, S.A. FDG-PET Scans in Patients with Kraepelinian and Non-Kraepelinian Schizophrenia. Eur. Arch. Psychiatry Clin. Neurosci. 2016, 266, 481–494. [Google Scholar] [CrossRef]
- Mitelman, S.A.; Bralet, M.C.; Mehmet Haznedar, M.; Hollander, E.; Shihabuddin, L.; Hazlett, E.A.; Buchsbaum, M.S. Positron Emission Tomography Assessment of Cerebral Glucose Metabolic Rates in Autism Spectrum Disorder and Schizophrenia. Brain Imaging Behav. 2018, 12, 532–546. [Google Scholar] [CrossRef]
- Buchsbaum, M.S.; Hazlett, E.A. Positron Emission Tomography Studies of Abnormal Glucose Metabolism in Schizophrenia. Schizophr. Bull. 1998, 24, 343–364. [Google Scholar] [CrossRef]
- Roldán, A.; Portella, M.J.; Sampedro, F.; Alonso-Solís, A.; Sarró, S.; Rabella, M.; Grasa, E.M.; Álvarez, E.; Rodríguez, R.; Camacho, V.; et al. Brain Metabolic Changes in Patients with Treatment Resistant Schizophrenia Treated with Deep Brain Stimulation: A Series of Cases. J. Psychiatr. Res. 2020, 127, 57–61. [Google Scholar] [CrossRef]
- Sukumar, N.; Sabesan, P.; Anazodo, U.; Palaniyappan, L. Neurovascular Uncoupling in Schizophrenia: A Bimodal Meta-Analysis of Brain Perfusion and Glucose Metabolism. Front. Psychiatry 2020, 11, 754. [Google Scholar] [CrossRef]
- Tomasella, E.; Falasco, G.; Urrutia, L.; Bechelli, L.; Padilla, L.; Gelman, D.M. Impaired Brain Glucose Metabolism and Presynaptic Dopaminergic Functioning in a Mouse Model of Schizophrenia. EJNMMI Res. 2020, 10, 39. [Google Scholar] [CrossRef]
- Hadar, R.; Soto-Montenegro, M.L.; Götz, T.; Wieske, F.; Sohr, R.; Desco, M.; Hamani, C.; Weiner, I.; Pascau, J.; Winter, C. Using a Maternal Immune Stimulation Model of Schizophrenia to Study Behavioral and Neurobiological Alterations over the Developmental Course. Schizophr. Res. 2015, 166, 238–247. [Google Scholar] [CrossRef]
- Kosten, L.; Verhaeghe, J.; Verkerk, R.; Thomae, D.; De Picker, L.; Wyffels, L.; Van Eetveldt, A.; Dedeurwaerdere, S.; Stroobants, S.; Staelens, S. Multiprobe Molecular Imaging of an NMDA Receptor Hypofunction Rat Model for Glutamatergic Dysfunction. Psychiatry Res. Neuroimaging 2016, 248, 1–11. [Google Scholar] [CrossRef]
- François, J.; Koning, E.; Ferrandon, A.; Sandner, G.; Nehlig, A. Metabolic Activity in the Brain of Juvenile and Adult Rats with a Neonatal Ventral Hippocampal Lesion. Hippocampus 2010, 20, 841–851. [Google Scholar] [CrossRef]
- Hanaya, R.; Koning, E.; Ferrandon, A.; Schweitzer, A.; Andrieux, A.; Nehlig, A. Deletion of the STOP Gene, a Microtubule Stabilizing Factor, Leads Only to Discrete Cerebral Metabolic Changes in Mice. J. Neurosci. Res. 2008, 86, 813–820. [Google Scholar] [CrossRef][Green Version]
- Joo, Y.H.; Kim, Y.K.; Choi, I.G.; Kim, H.J.; Son, Y.D.; Kim, H.K.; Cumming, P.; Kim, J.H. In Vivo Glucose Metabolism and Glutamate Levels in MGluR5 Knockout Mice: A Multimodal Neuroimaging Study Using [18F]FDG MicroPET and MRS. EJNMMI Res. 2020, 10, 116. [Google Scholar] [CrossRef]
- Casquero-Veiga, M.; García-García, D.; MacDowell, K.S.; Pérez-Caballero, L.; Torres-Sánchez, S.; Fraguas, D.; Berrocoso, E.; Leza, J.C.; Arango, C.; Desco, M.; et al. Risperidone Administered during Adolescence Induced Metabolic, Anatomical and Inflammatory/Oxidative Changes in Adult Brain: A PET and MRI Study in the Maternal Immune Stimulation Animal Model. Eur. Neuropsychopharmacol. 2019, 29, 880–896. [Google Scholar] [CrossRef]
- Bikovsky, L.; Hadar, R.; Soto-Montenegro, M.L.; Klein, J.; Weiner, I.; Desco, M.; Pascau, J.; Winter, C.; Hamani, C. Deep Brain Stimulation Improves Behavior and Modulates Neural Circuits in a Rodent Model of Schizophrenia. Exp. Neurol. 2016, 283, 142–150. [Google Scholar] [CrossRef]
- Horvath, G.; Kis, G.; Kekesi, G.; Büki, A.; Adlan, L.G.; Szűcs, E.; Heni, H.E.; Benyhe, S. Interaction of Clozapine with Metformin in a Schizophrenia Rat Model. Sci. Rep. 2021, 11, 16862. [Google Scholar] [CrossRef]
- Kekesi, G.; Petrovszki, Z.; Benedek, G.; Horvath, G. Sex-Specific Alterations in Behavioral and Cognitive Functions in a “Three Hit” Animal Model of Schizophrenia. Behav. Brain Res. 2015, 284, 85–93. [Google Scholar] [CrossRef]
- Petrovszki, Z.; Adam, G.; Tuboly, G.; Kekesi, G.; Benedek, G.; Keri, S.; Horvath, G. Characterization of Gene-Environment Interactions by Behavioral Profiling of Selectively Bred Rats: The Effect of NMDA Receptor Inhibition and Social Isolation. Behav. Brain Res. 2013, 240, 134–145. [Google Scholar] [CrossRef]
- Horvath, G.; Liszli, P.; Kekesi, G.; Büki, A.; Benedek, G. Characterization of Exploratory Activity and Learning Ability of Healthy and “Schizophrenia-like” Rats in a Square Corridor System (AMBITUS). Physiol. Behav. 2017, 169, 155–164. [Google Scholar] [CrossRef]
- Banki, L.; Büki, A.; Horvath, G.; Kekesi, G.; Kis, G.; Somogyvári, F.; Jancsó, G.; Vécsei, L.; Varga, E.; Tuboly, G. Distinct Changes in Chronic Pain Sensitivity and Oxytocin Receptor Expression in a New Rat Model (Wisket) of Schizophrenia. Neurosci. Lett. 2020, 714, 134561. [Google Scholar] [CrossRef]
- Horvath, G.; Adam, G.; Tuboly, G.; Kekesi, G.; Büki, A.; Ducza, E.; Szűcs, E.; Benyhe, S.; Benedek, G. Caffeine—Treat or Trigger? Disparate Behavioral and Long-Term Dopaminergic Changes in Control and Schizophrenia-like Wisket Rats. Physiol. Behav. 2021, 236, 113410. [Google Scholar] [CrossRef]
- Szűcs, E.; Dvorácskó, S.; Tömböly, C.; Büki, A.; Kékesi, G.; Horváth, G.; Benyhe, S. Decreased CB Receptor Binding and Cannabinoid Signaling in Three Brain Regions of a Rat Model of Schizophrenia. Neurosci. Lett. 2016, 633, 87–93. [Google Scholar] [CrossRef]
- Horvath, G.; Petrovszki, Z.; Kekesi, G.; Tuboly, G.; Bodosi, B.; Horvath, J.; Gombköto, P.; Benedek, G.; Nagy, A. Electrophysiological Alterations in a Complex Rat Model of Schizophrenia. Behav. Brain Res. 2016, 307, 65–72. [Google Scholar] [CrossRef]
- Szűcs, E.; Ducza, E.; Büki, A.; Kekesi, G.; Benyhe, S.; Horvath, G. Characterization of Dopamine D2 Receptor Binding, Expression and Signaling in Different Brain Regions of Control and Schizophrenia-Model Wisket Rats. Brain Res. 2020, 1748, 147074. [Google Scholar] [CrossRef]
- Adlan, L.G.; Csordás-Nagy, M.; Bodosi, B.; Kalmár, G.; Nyúl, L.G.; Nagy, A.; Kekesi, G.; Büki, A.; Horvath, G. Sleep-Wake Rhythm and Oscillatory Pattern Analysis in a Multiple Hit Schizophrenia Rat Model (Wisket). Front. Behav. Neurosci. 2022, 15, 799271. [Google Scholar] [CrossRef]
- Büki, A.; Bohár, Z.; Kekesi, G.; Vécsei, L.; Horvath, G. Wisket Rat Model of Schizophrenia: Impaired Motivation and, Altered Brain Structure, but No Anhedonia. Physiol. Behav. 2021, 244, 113651. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Bättig, K.; Holmén, J.; Nehlig, A.; Zvartau, E.E. Actions of Caffeine in the Brain with Special Reference to Factors That Contribute to Its Widespread Use. Pharmacol. Rev. 1999, 51, 83–133. [Google Scholar]
- Dall’Igna, O.P.; Da Silva, A.L.; Dietrich, M.O.; Hoffmann, A.; De Oliveira, R.V.; Souza, D.O.; Lara, D.R. Chronic Treatment with Caffeine Blunts the Hyperlocomotor but Not Cognitive Effects of the N-Methyl-D-Aspartate Receptor Antagonist MK-801 in Mice. Psychopharmacology 2003, 166, 258–263. [Google Scholar] [CrossRef]
- Lara, D.R.; Souza, D.O. Schizophrenia: A Purinergic Hypothesis. Med. Hypotheses 2000, 54, 157–166. [Google Scholar] [CrossRef]
- De Oliveira, R.V.; Dall’Igna, O.P.; Tort, A.B.L.; Schuh, J.F.; Neto, P.F.; Gomes, M.W.S.; Souza, D.O.; Lara, D.R. Effect of Subchronic Caffeine Treatment on MK-801-Induced Changes in Locomotion, Cognition and Ataxia in Mice. Behav. Pharmacol. 2005, 16, 79–84. [Google Scholar] [CrossRef]
- Hughes, J.R.; McHugh, P.; Holtzman, S.; Frances, R.J. Caffeine and Schizophrenia. Psychiatr. Serv. 1998, 49, 1415–1417. [Google Scholar] [CrossRef]
- Huang, L.; Sperlágh, B. Caffeine Consumption and Schizophrenia: A Highlight on Adenosine Receptor–Independent Mechanisms. Curr. Opin. Pharmacol. 2021, 61, 106–113. [Google Scholar] [CrossRef]
- Gandhi, K.K.; Williams, J.M.; Menza, M.; Galazyn, M.; Benowitz, N.L. Higher Serum Caffeine in Smokers with Schizophrenia Compared to Smoking Controls. Drug Alcohol Depend. 2010, 110, 151–155. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cameron, O.G.; Modell, J.G.; Hariharan, M. Caffeine and Human Cerebral Blood Flow: A Positron Emission Tomography Study. Life Sci. 1990, 47, 1141–1146. [Google Scholar] [CrossRef]
- Park, C.A.; Kang, C.K.; Son, Y.D.; Choi, E.J.; Kim, S.H.; Oh, S.T.; Kim, Y.B.; Park, C.W.; Cho, Z.H. The Effects of Caffeine Ingestion on Cortical Areas: Functional Imaging Study. Magn. Reson. Imaging 2014, 32, 366–371. [Google Scholar] [CrossRef]
- Nehlig, A.; Daval, J.L.; Boyet, S.; Vert, P. Comparative Effects of Acute and Chronic Administration of Caffeine on Local Cerebral Glucose Utilization in the Conscious Rat. Eur. J. Pharmacol. 1986, 129, 93–103. [Google Scholar] [CrossRef]
- Nehlig, A.; Lucignani, G.; Kadekaro, M.; Porrino, L.J.; Sokoloff, L. Effects of Acute Administration of Caffeine on Local Cerebral Glucose Utilization in the Rat. Eur. J. Pharmacol. 1984, 101, 91–100. [Google Scholar] [CrossRef]
- Nehlig, A.; Boyet, S. Dose-Response Study of Caffeine Effects on Cerebral Functional Activity with a Specific Focus on Dependence. Brain Res. 2000, 858, 71–77. [Google Scholar] [CrossRef]
- Peng, S.L.; Chiu, H.; Wu, C.Y.; Huang, C.W.; Chung, Y.H.; Shih, C.T.; Shen, W.C. The Effect of Caffeine on Cerebral Metabolism during Alpha-Chloralose Anesthesia Differs from Isoflurane Anesthesia in the Rat Brain. Psychopharmacology 2019, 236, 1749–1757. [Google Scholar] [CrossRef]
- Davidson, L.L.; Heinrichs, R.W. Quantification of Frontal and Temporal Lobe Brain-Imaging Findings in Schizophrenia: A Meta-Analysis. Psychiatry Res. Neuroimaging 2003, 122, 69–87. [Google Scholar] [CrossRef]
- Molina, V.; Sanz, J.; Sarramea, F.; Benito, C.; Palomo, T. Prefrontal Atrophy in First Episodes of Schizophrenia Associated with Limbic Metabolic Hyperactivity. J. Psychiatr. Res. 2005, 39, 117–127. [Google Scholar] [CrossRef]
- Lehrer, D.S.; Christian, B.T.; Mantil, J.; Murray, A.C.; Buchsbaum, B.R.; Oakes, T.R.; Byne, W.; Kemether, E.M.; Buchsbaum, M.S. Thalamic and Prefrontal FDG Uptake in Never Medicated Patients with Schizophrenia. Am. J. Psychiatry 2005, 162, 931–938. [Google Scholar] [CrossRef]
- Potkin, S.G.; Alva, G.; Fleming, K.; Anand, R.; Keator, D.; Carreon, D.; Doo, M.; Jin, Y.; Wu, J.C.; Fallon, J.H. A PET Study of the Pathophysiology of Negative Symptoms in Schizophrenia. Am. J. Psychiatry 2002, 159, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Bonab, A.A.; Fricchione, J.G.; Gorantla, S.; Vitalo, A.G.; Auster, M.E.; Levine, S.J.; Scichilone, J.M.; Hegde, M.; Foote, W.; Fricchione, G.L.; et al. Isolation Rearing Significantly Perturbs Brain Metabolism in the Thalamus and Hippocampus. Neuroscience 2012, 223, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, N.R. Integrative Circuit Models and Their Implications for the Pathophysiologies and Treatments of the Schizophrenias. Curr. Top. Behav. Neurosci. 2010, 4, 555–583. [Google Scholar] [CrossRef]
- Horvath, G.; Kekesi, G.; Petrovszki, Z.; Benedek, G. Abnormal Motor Activity and Thermoregulation in a Schizophrenia Rat Model for Translational Science. PLoS ONE 2015, 10, e0143751. [Google Scholar] [CrossRef] [PubMed]
- McLellan, T.M.; Caldwell, J.A.; Lieberman, H.R. A Review of Caffeine’s Effects on Cognitive, Physical and Occupational Performance. Neurosci. Biobehav. Rev. 2016, 71, 294–312. [Google Scholar] [CrossRef]
- Ongini, E. Pharmacology of Adenosine A2A Receptors. Trends Pharmacol. Sci. 1996, 17, 364–372. [Google Scholar] [CrossRef]
- Boison, D.; Singer, P.; Shen, H.-Y.; Feldon, J.; Yee, B.K. Adenosine Hypothesis of Schizophrenia—Opportunities for Pharmacotherapy. Neuropharmacology 2012, 62, 1527–1543. [Google Scholar] [CrossRef] [PubMed]
- Quarta, D.; Ferré, S.; Solinas, M.; You, Z.B.; Hockemeyer, J.; Popoli, P.; Goldberg, S.R. Opposite Modulatory Roles for Adenosine A1 and A2A Receptors on Glutamate and Dopamine Release in the Shell of the Nucleus Accumbens. Effects of Chronic Caffeine Exposure. J. Neurochem. 2004, 88, 1151–1158. [Google Scholar] [CrossRef]
- Prediger, R.D.S.; Pamplona, F.A.; Fernandes, D.; Takahashi, R.N. Caffeine Improves Spatial Learning Deficits in an Animal Model of Attention Deficit Hyperactivity Disorder (ADHD)—The Spontaneously Hypertensive Rat (SHR). Int. J. Neuropsychopharmacol. 2005, 8, 583–594. [Google Scholar] [CrossRef]
- Prediger, R.D.S.; Batista, L.C.; Takahashi, R.N. Caffeine Reverses Age-Related Deficits in Olfactory Discrimination and Social Recognition Memory in Rats: Involvement of Adenosine A1 and A2A Receptors. Neurobiol. Aging 2005, 26, 957–964. [Google Scholar] [CrossRef]
- Lucas, P.B.; Pickar, D.; Kelsoe, J.; Rapaport, M.; Pato, C.; Hommer, D. Effects of the Acute Administration of Caffeine in Patients with Schizophrenia. Biol. Psychiatry 1990, 28, 35–40. [Google Scholar] [CrossRef]
- Núñez, C.; Stephan-Otto, C.; Cuevas-Esteban, J.; Maria Haro, J.; Huerta-Ramos, E.; Ochoa, S.; Usall, J.; Brébion, G. Effects of Caffeine Intake and Smoking on Neurocognition in Schizophrenia. Psychiatry Res. 2015, 230, 924–931. [Google Scholar] [CrossRef] [PubMed]
- SanMiguel, N.; Pardo, M.; Carratalá-Ros, C.; López-Cruz, L.; Salamone, J.D.; Correa, M. Individual Differences in the Energizing Effects of Caffeine on Effort-Based Decision-Making Tests in Rats. Pharmacol. Biochem. Behav. 2018, 169, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Angelucci, M.E.M.; Cesário, C.; Hiroi, R.H.; Rosalen, P.L.; Da Cunha, C. Effects of Caffeine on Learning and Memory in Rats Tested in the Morris Water Maze. Braz. J. Med. Biol. Res. 2002, 35, 1201–1208. [Google Scholar] [CrossRef]
- Dubroqua, S.; Low, S.R.L.; Yee, B.K.; Singer, P. Caffeine Impairs the Acquisition and Retention, but Not the Consolidation of Pavlovian Conditioned Freezing in Mice. Psychopharmacology 2015, 232, 721–731. [Google Scholar] [CrossRef]
- Pedraza, L.K.; Sierra, R.O.; Lotz, F.N.; Alvares, L.D.O. Periodical Reactivation under the Effect of Caffeine Attenuates Fear Memory Expression in Rats. Sci. Rep. 2018, 8, 7260. [Google Scholar] [CrossRef]
- Griffeth, V.E.M.; Perthen, J.E.; Buxton, R.B. Prospects for Quantitative FMRI: Investigating the Effects of Caffeine on Baseline Oxygen Metabolism and the Response to a Visual Stimulus in Humans. Neuroimage 2011, 57, 809–816. [Google Scholar] [CrossRef]
- Merola, A.; Germuska, M.A.; Warnert, E.A.; Richmond, L.; Helme, D.; Khot, S.; Murphy, K.; Rogers, P.J.; Hall, J.E.; Wise, R.G. Mapping the Pharmacological Modulation of Brain Oxygen Metabolism: The Effects of Caffeine on Absolute CMRO2 Measured Using Dual Calibrated FMRI. Neuroimage 2017, 155, 331–343. [Google Scholar] [CrossRef]
- Xu, F.; Liu, P.; Pekar, J.J.; Lu, H. Does Acute Caffeine Ingestion Alter Brain Metabolism in Young Adults? Neuroimage 2015, 110, 39–47. [Google Scholar] [CrossRef]
- Nehlig, A.; de Vasconcelos, A.P.; Dumont, I.; Boyet, S. Effects of Caffeine, L-Phenylisopropyladenosine and Their Combination on Local Cerebral Blood Flow in the Rat. Eur. J. Pharmacol. 1990, 179, 271–280. [Google Scholar] [CrossRef]
- Nabbi-Schroeter, D.; Elmenhorst, D.; Oskamp, A.; Laskowski, S.; Bauer, A.; Kroll, T. Effects of Long-Term Caffeine Consumption on the Adenosine A1 Receptor in the Rat Brain: An in Vivo PET Study with [18F]CPFPX. Mol. Imaging Biol. 2018, 20, 284–291. [Google Scholar] [CrossRef]
- Kaplan, G.B.; Greenblatt, D.J.; Kent, M.A.; Cotreau-Bibbo, M.M. Caffeine Treatment and Withdrawal in Mice: Relationships between Dosage, Concentrations, Locomotor Activity and A1 Adenosine Receptor Binding. J. Pharmacol. Exp. Ther. 1993, 266, 1563–1572. [Google Scholar] [PubMed]
- Lemos, C.; Pinheiro, B.S.; Beleza, R.O.; Marques, J.M.; Rodrigues, R.J.; Cunha, R.A.; Rial, D.; Köfalvi, A. Adenosine A2B Receptor Activation Stimulates Glucose Uptake in the Mouse Forebrain. Purinergic Signal. 2015, 11, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, E.R.; Parent, M.J.; Souza, D.G.; Leuzy, A.; Lecrux, C.; Kim, H.I.; Gauthier, S.; Pellerin, L.; Hamel, E.; Rosa-Neto, P. [18F]FDG PET Signal Is Driven by Astroglial Glutamate Transport. Nat. Neurosci. 2017, 20, 393–395. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Chang, L.; Wang, G.J.; Fowler, J.S.; Ding, Y.S.; Sedler, M.; Logan, J.; Franceschi, D.; Gatley, J.; Hitzemann, R.; et al. Low Level of Brain Dopamine D2 Receptors in Methamphetamine Abusers: Association with Metabolism in the Orbitofrontal Cortex. Am. J. Psychiatry 2001, 158, 2015–2021. [Google Scholar] [CrossRef]
- Mitelman, S.A.; Buchsbaum, M.S.; Christian, B.T.; Merrill, B.M.; Buchsbaum, B.R.; Mukherjee, J.; Lehrer, D.S. Positive Association between Cerebral Grey Matter Metabolism and Dopamine D2/D3 Receptor Availability in Healthy and Schizophrenia Subjects: An 18F-Fluorodeoxyglucose and 18F-Fallypride Positron Emission Tomography Study. World J. Biol. Psychiatry 2020, 21, 368–382. [Google Scholar] [CrossRef] [PubMed]
- Hanna, C.; Hamilton, J.; Arnavut, E.; Blum, K.; Thanos, P.K. Brain Mapping the Effects of Chronic Aerobic Exercise in the Rat Brain Using FDG PET. J. Pers. Med. 2022, 12, 860. [Google Scholar] [CrossRef] [PubMed]
- Marsteller, D.A.; Barbarich-Marsteller, N.C.; Fowler, J.S.; Schiffer, W.K.; Alexoff, D.L.; Rubins, D.J.; Dewey, S.L. Reproducibility of Intraperitoneal 2-Deoxy-2-[18F]-Fluoro-D-Glucose Cerebral Uptake in Rodents through Time. Nucl. Med. Biol. 2006, 33, 71–79. [Google Scholar] [CrossRef]
- McGregor, M.; Richer, K.; Ananth, M.; Thanos, P.K. The Functional Networks of a Novel Environment: Neural Activity Mapping in Awake Unrestrained Rats Using Positron Emission Tomography. Brain Behav. 2020, 10, e01646. [Google Scholar] [CrossRef]
- Kurucz, A.; Bombicz, M.; Kiss, R.; Priksz, D.; Varga, B.; Hortobágyi, T.; Trencsényi, G.; Szabó, R.; Pósa, A.; Gesztelyi, R.; et al. Heme Oxygenase-1 Activity as a Correlate to Exercise-Mediated Amelioration of Cognitive Decline and Neuropathological Alterations in an Aging Rat Model of Dementia. Biomed Res. Int. 2018, 2018, 7212861. [Google Scholar] [CrossRef]
- Trencsényi, G.; Márián, T.; Lajtos, I.; Krasznai, Z.; Balkay, L.; Emri, M.; Mikecz, P.; Goda, K.; Szalóki, G.; Juhász, I.; et al. 18FDG, [18F]FLT, [18F]FAZA, and 11C-Methionine Are Suitable Tracers for the Diagnosis and in Vivo Follow-up of the Efficacy of Chemotherapy by MiniPET in Both Multidrug Resistant and Sensitive Human Gynecologic Tumor Xenografts. Biomed Res. Int. 2014, 2014, 787365. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horvath, G.; Kertész, I.; Nagy, T.; Adlan, L.G.; Kekesi, G.; Büki, A.; Tuboly, G.; Trencsényi, G. Caffeine-Induced Acute and Delayed Responses in Cerebral Metabolism of Control and Schizophrenia-like Wisket Rats. Int. J. Mol. Sci. 2022, 23, 8186. https://doi.org/10.3390/ijms23158186
Horvath G, Kertész I, Nagy T, Adlan LG, Kekesi G, Büki A, Tuboly G, Trencsényi G. Caffeine-Induced Acute and Delayed Responses in Cerebral Metabolism of Control and Schizophrenia-like Wisket Rats. International Journal of Molecular Sciences. 2022; 23(15):8186. https://doi.org/10.3390/ijms23158186
Chicago/Turabian StyleHorvath, Gyöngyi, István Kertész, Tamás Nagy, Leatitia Gabriella Adlan, Gabriella Kekesi, Alexandra Büki, Gabor Tuboly, and György Trencsényi. 2022. "Caffeine-Induced Acute and Delayed Responses in Cerebral Metabolism of Control and Schizophrenia-like Wisket Rats" International Journal of Molecular Sciences 23, no. 15: 8186. https://doi.org/10.3390/ijms23158186
APA StyleHorvath, G., Kertész, I., Nagy, T., Adlan, L. G., Kekesi, G., Büki, A., Tuboly, G., & Trencsényi, G. (2022). Caffeine-Induced Acute and Delayed Responses in Cerebral Metabolism of Control and Schizophrenia-like Wisket Rats. International Journal of Molecular Sciences, 23(15), 8186. https://doi.org/10.3390/ijms23158186