
A Dynamic Network of Proteins Facilitate Cell Envelope Biogenesis in Gram-Negative Bacteria

 , ,
, ,
Abstract
: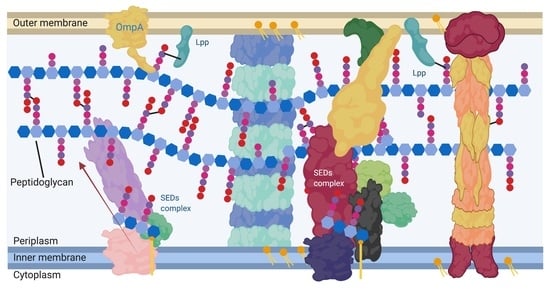
1. Peptidoglycan in Gram-Negatives
2. Cell Wall Modifying Enzymes and Complexes Have Altered Localisation during Growth Which Is Essential for Specialised Peptidoglycan Biosynthesis
3. Regulation of Peptidoglycan Modifying Enzymes by Their Interacting Partners
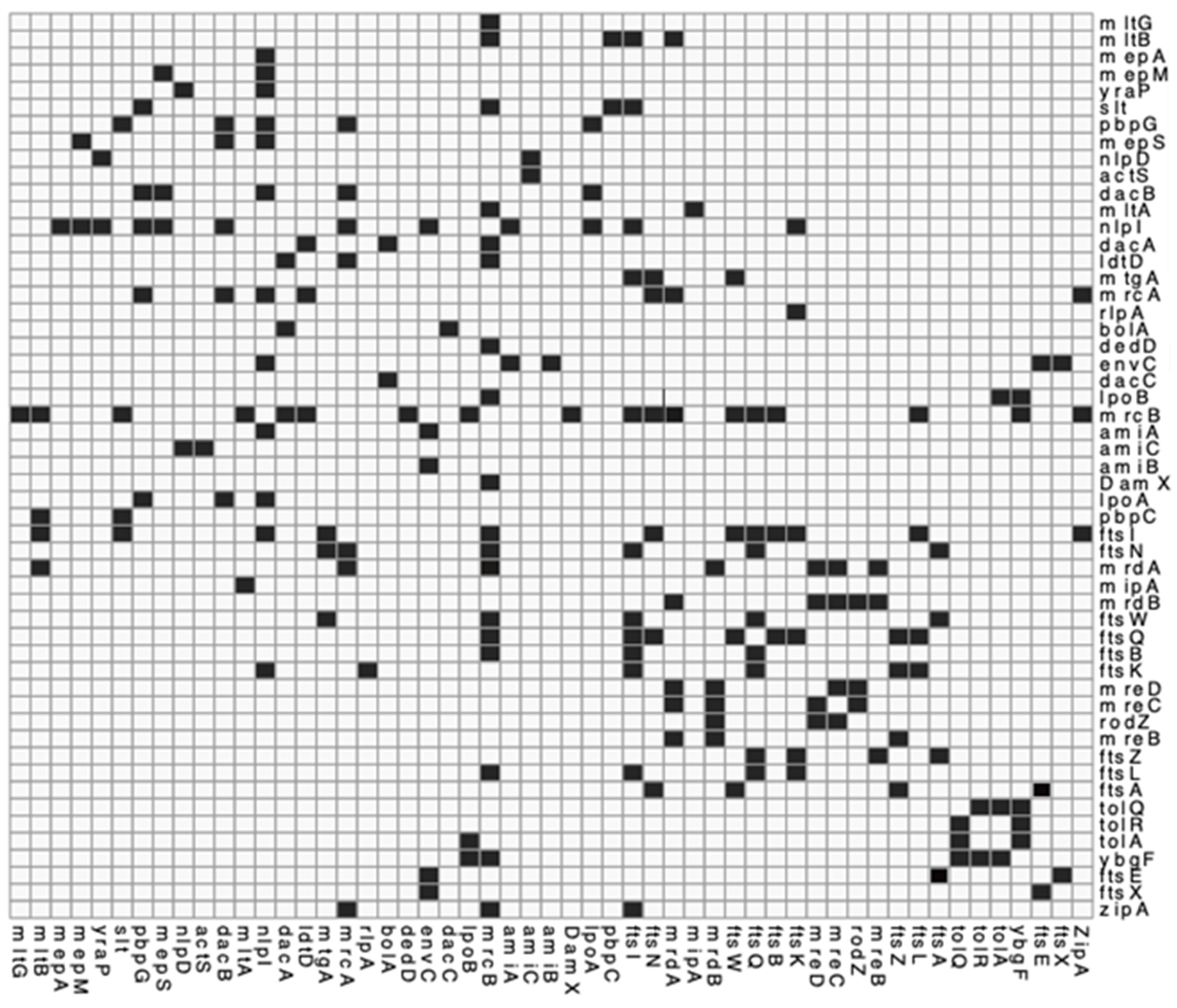
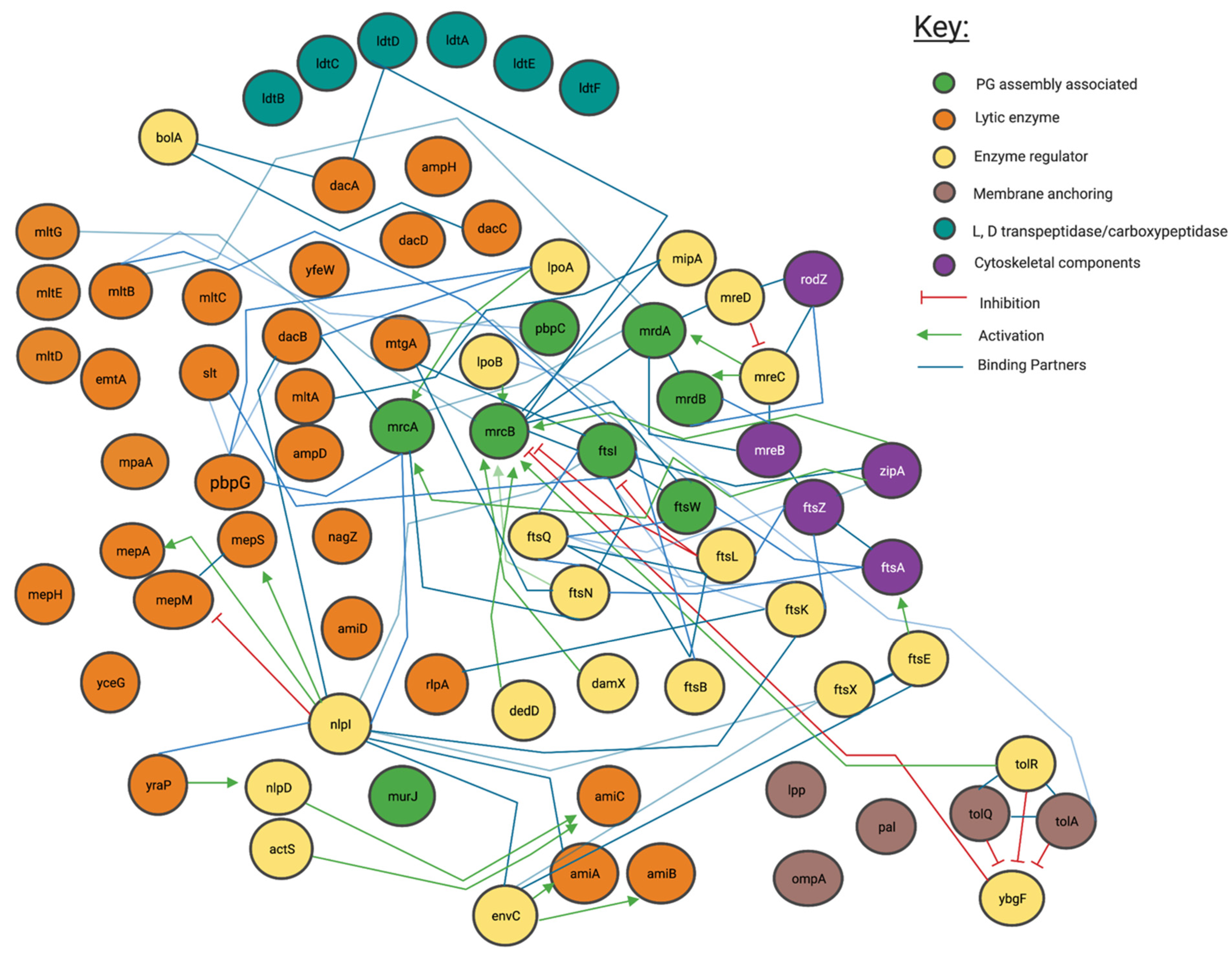
4. Method Used to Visualise PG Synthesis Networks for This Meta-Review
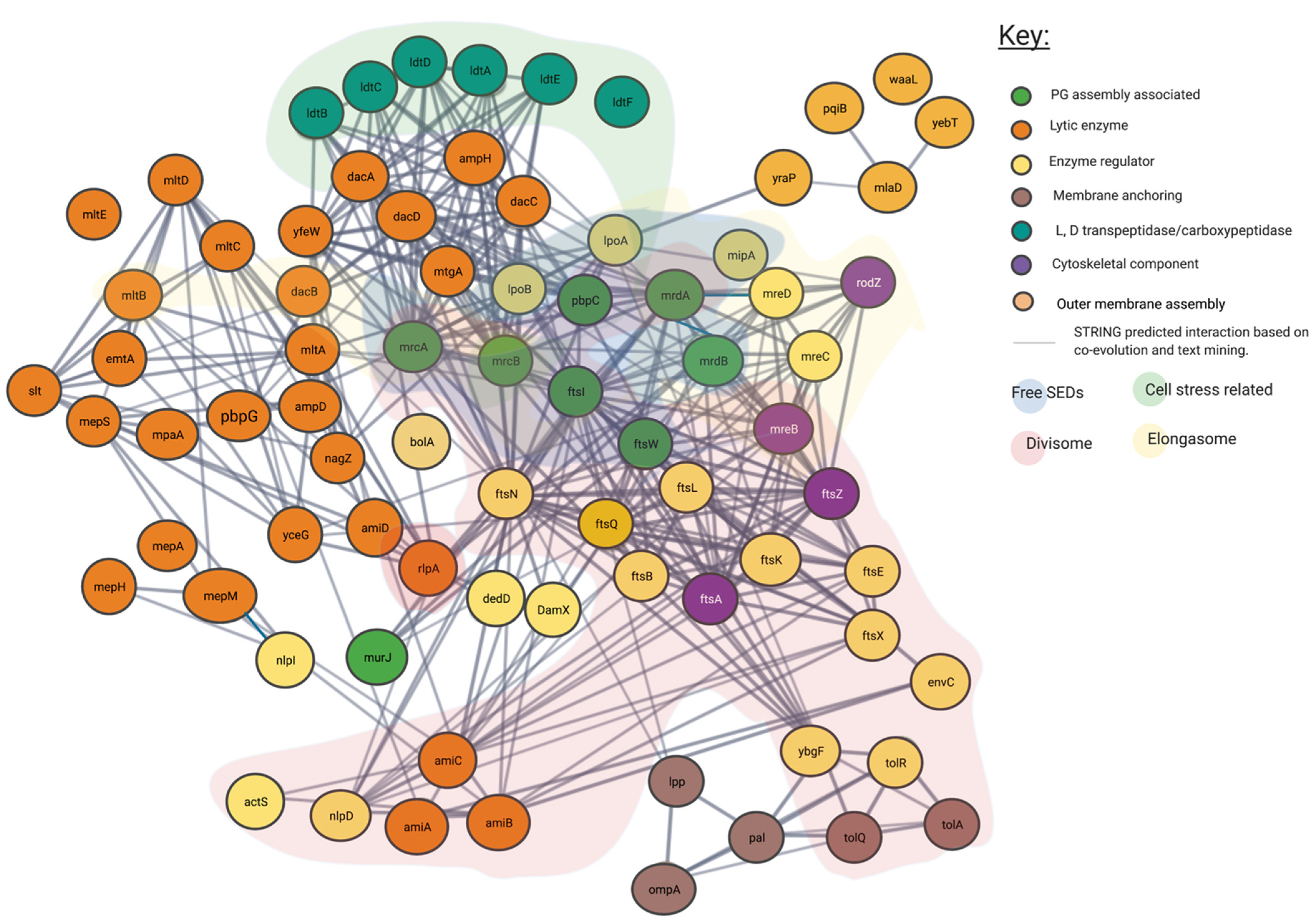
4.1. Genetic and Protein Interactions Confirmed by the Literature
4.2. Proposed Genetic Interactions
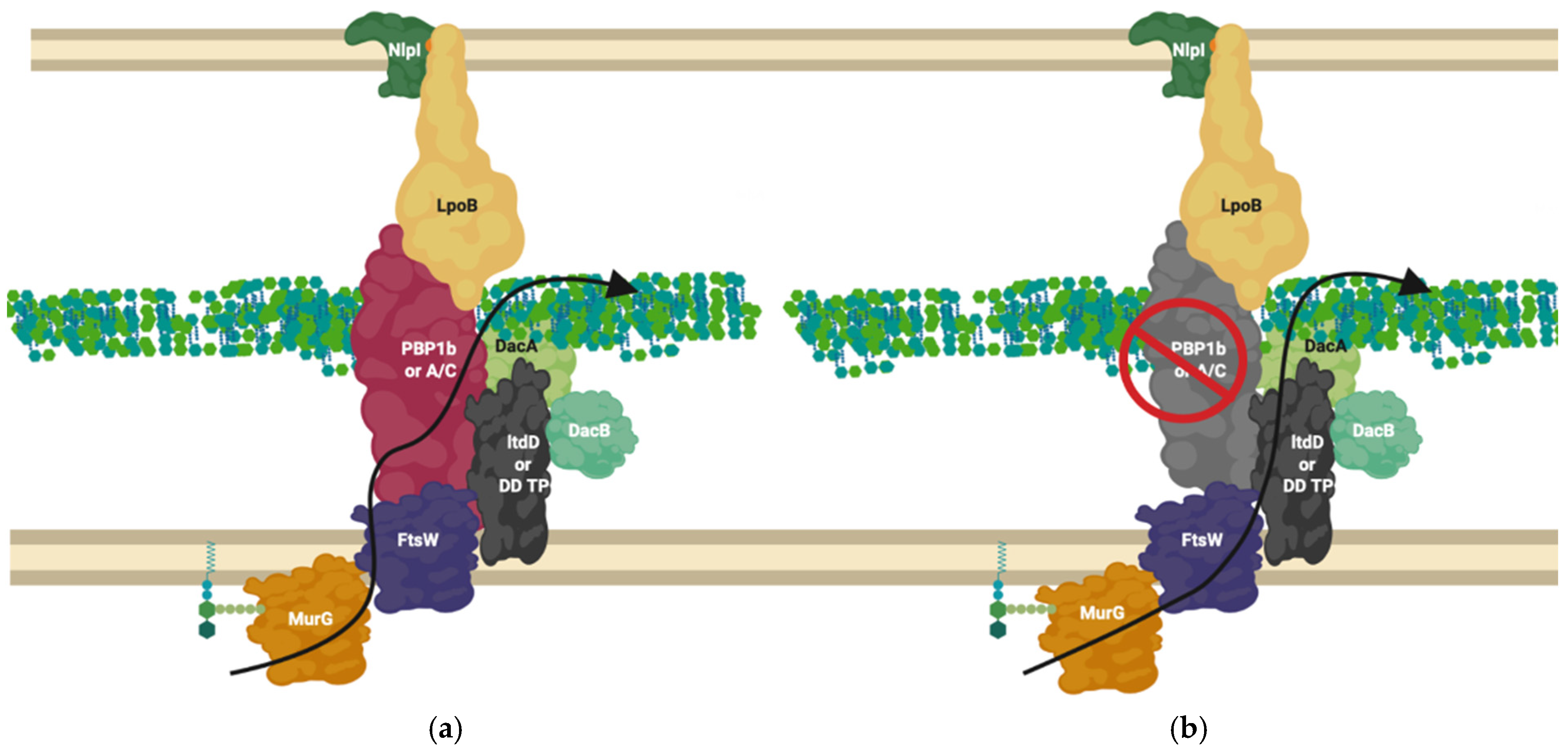
5. Most Peptidoglycan Synthases and Modifiers Are Members of Multiple Local Complexes as Predicted by Genetic Interactions and Confirmed by Literature
6. Cytoskeletal Proteins Create Nodes of Complex Formation
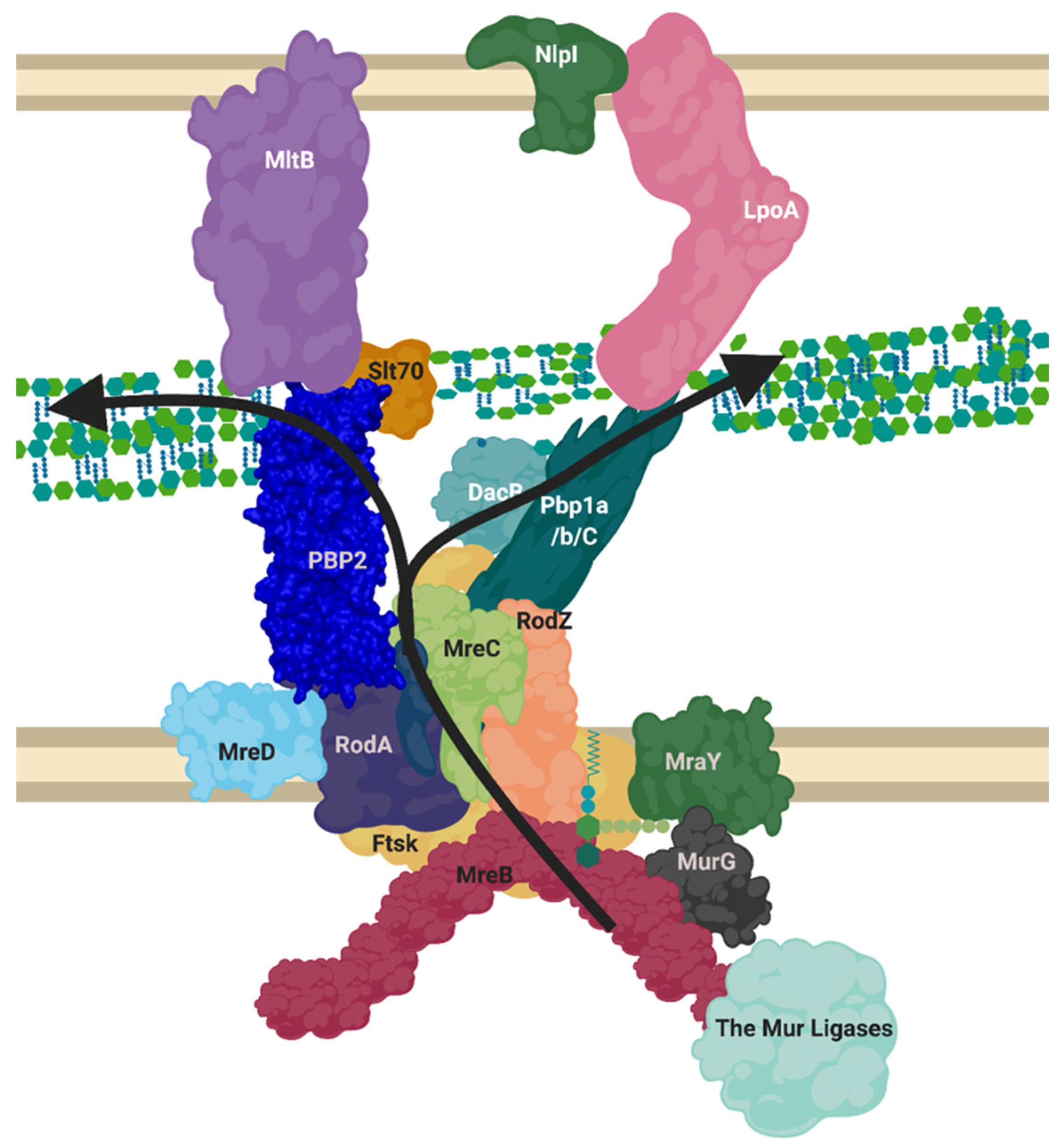
7. The “Elongasome” Is a Collection of Multiple Complexes
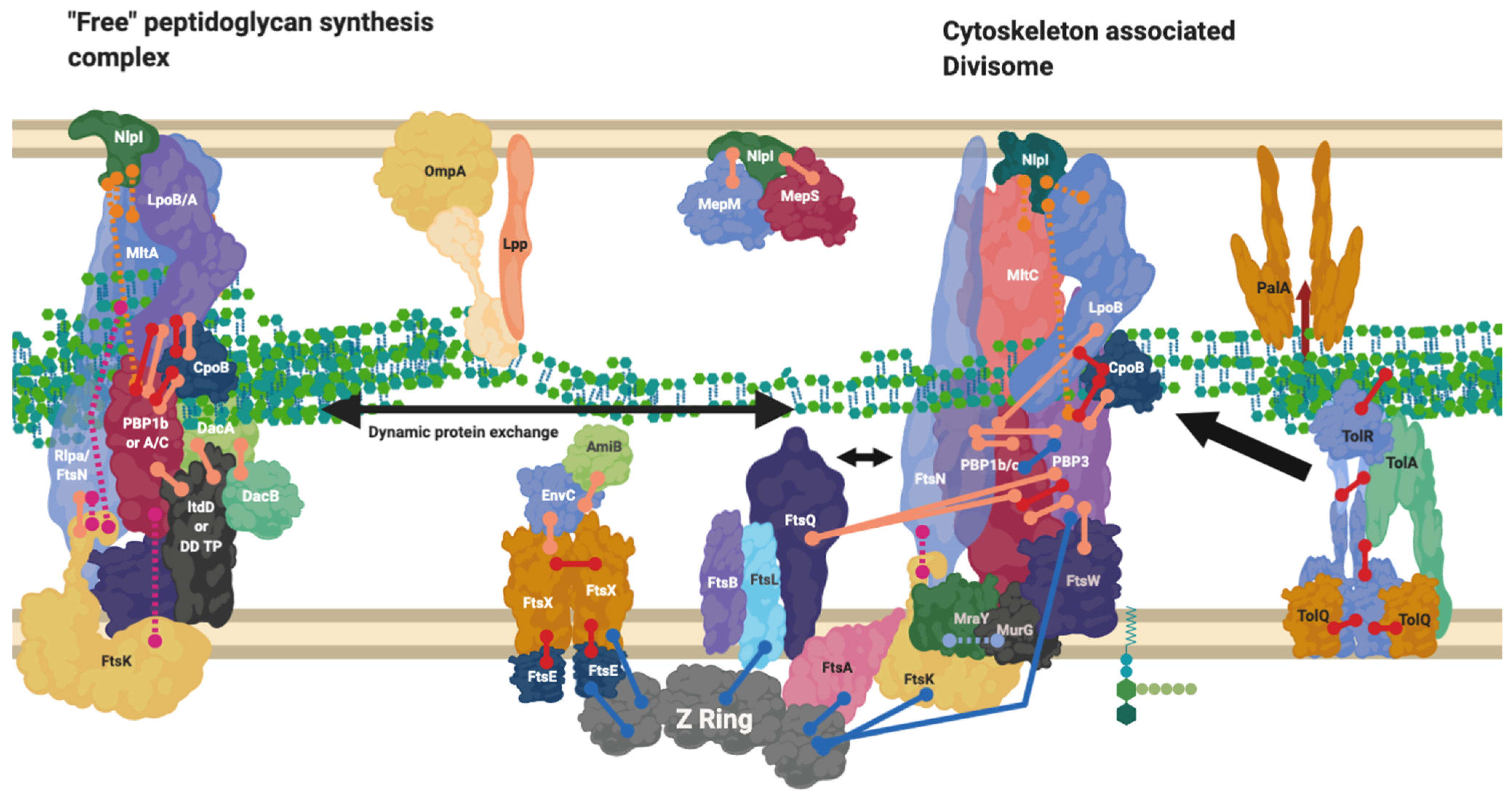
8. NlpI Acts as a Facilitator of PBP Nucleation and Complex Interaction
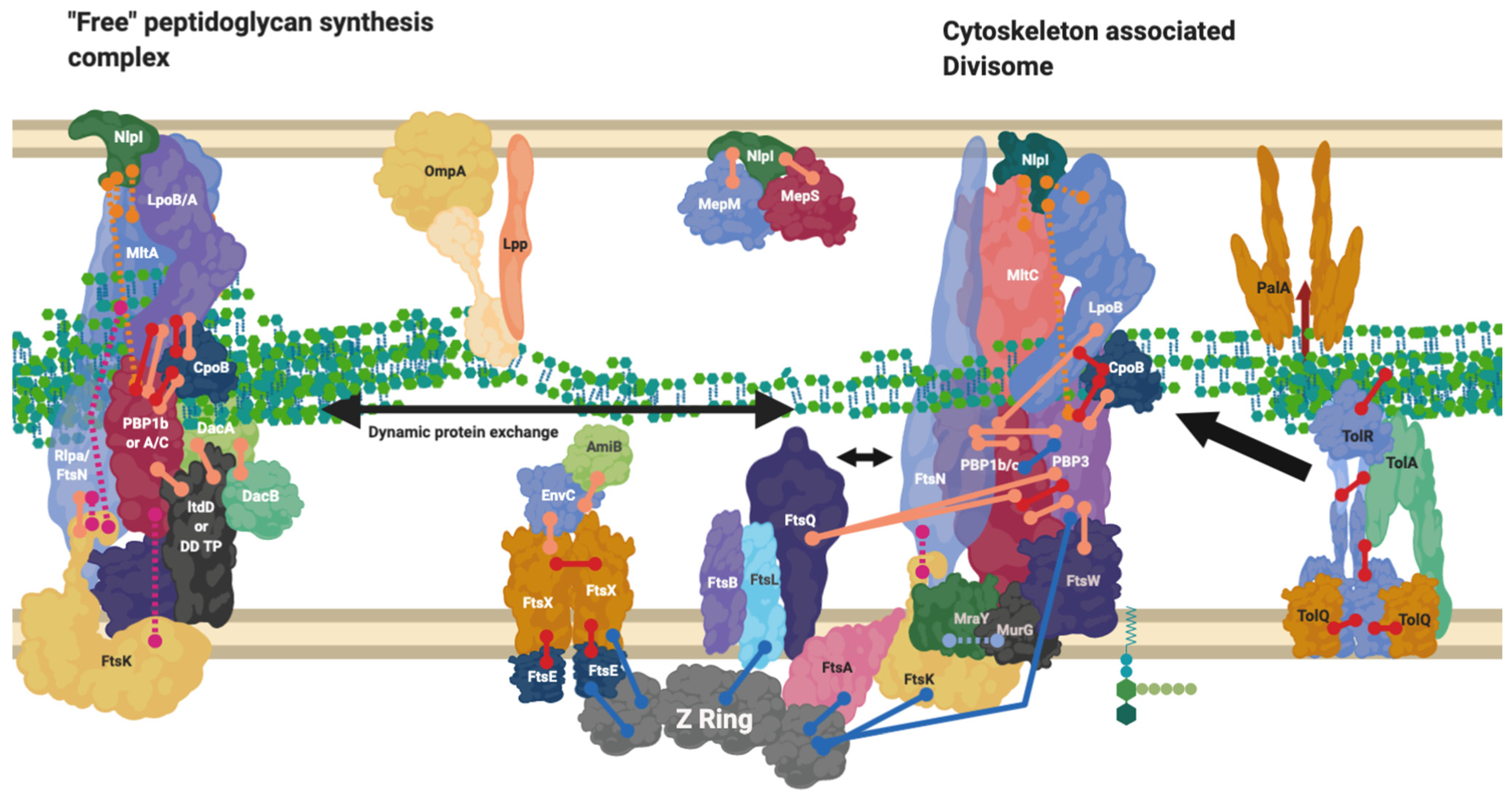
9. The Divisome Is a Series of Complexes Controlled by Cytoplasmic Events
10. Proteins Interchange between Complexes, and Complexes Interact
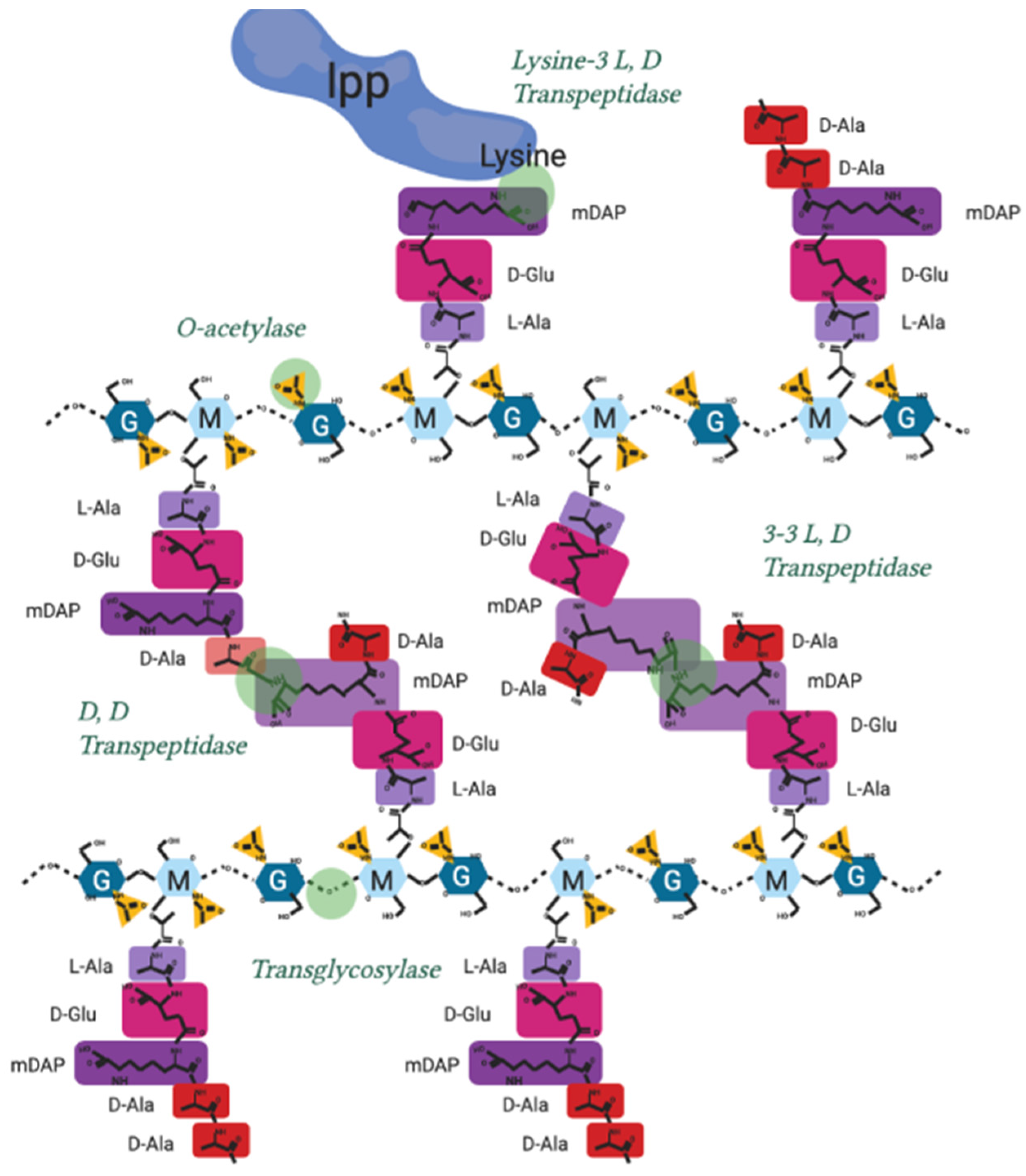
11. Alternate Protein Complexes Exist, Containing 3-3 Crosslinking L, D Transpeptidases as an Alternative to 3-4 Crosslinking PBPs Important for Antibiotic Resistance
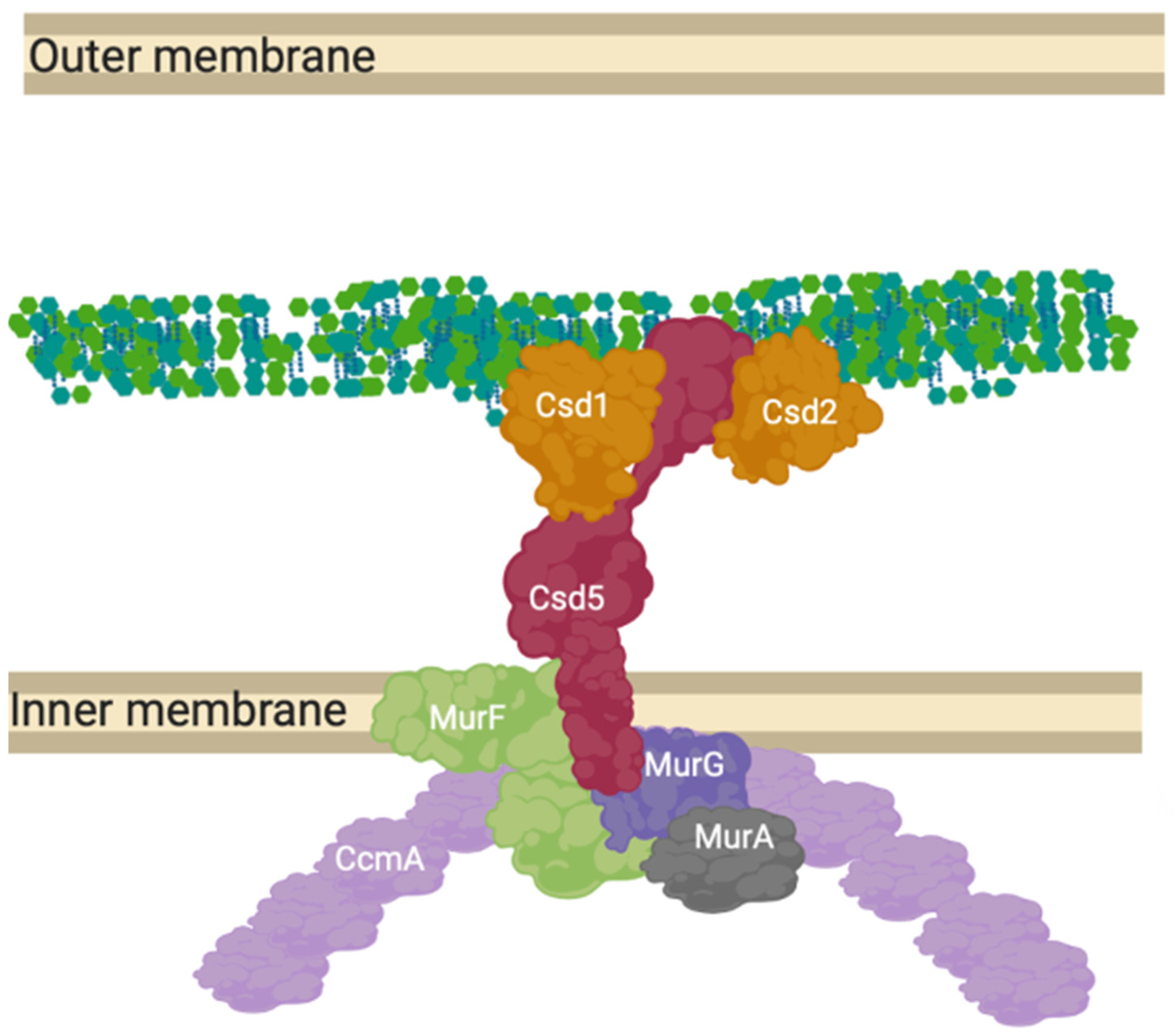
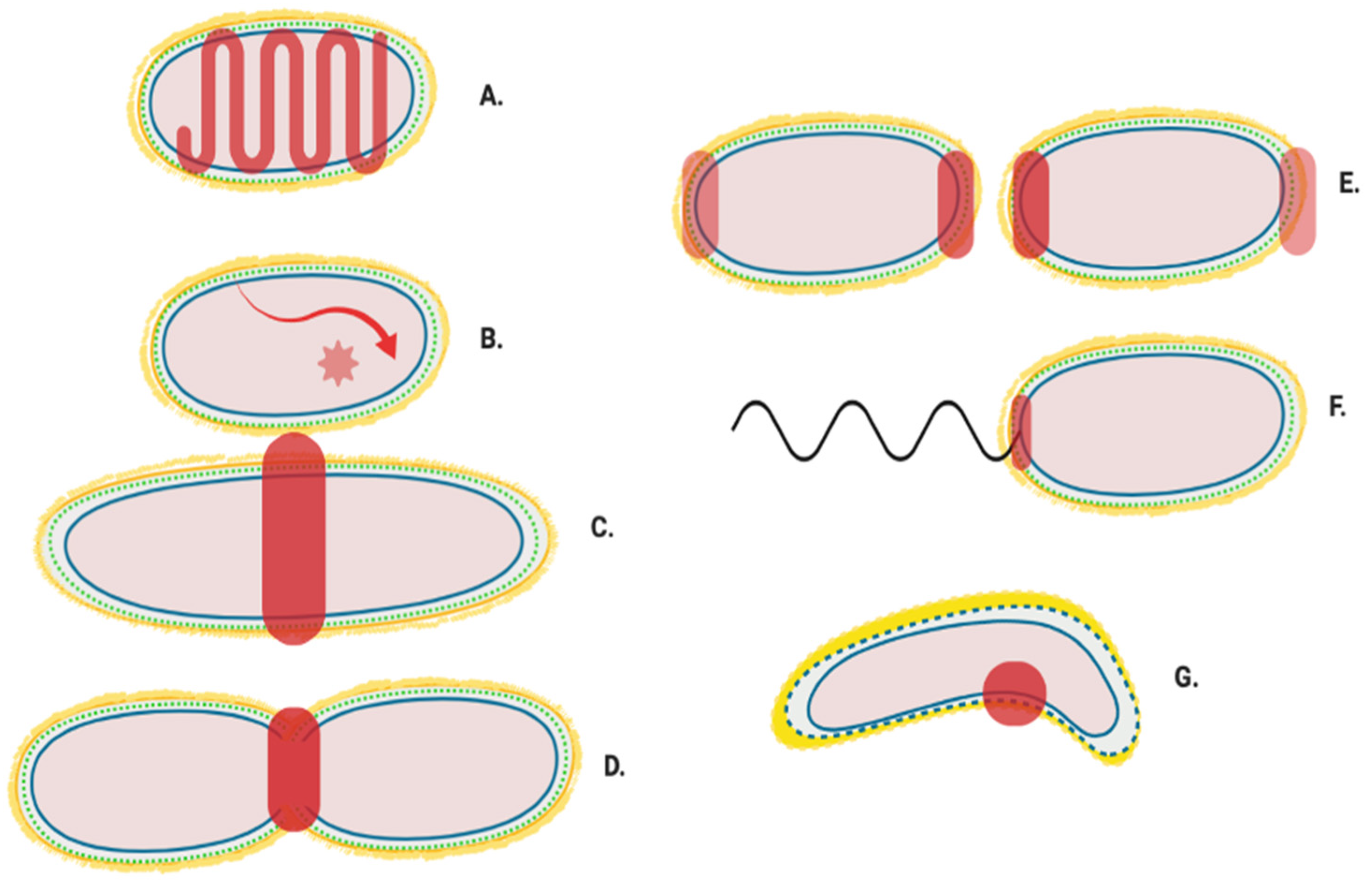
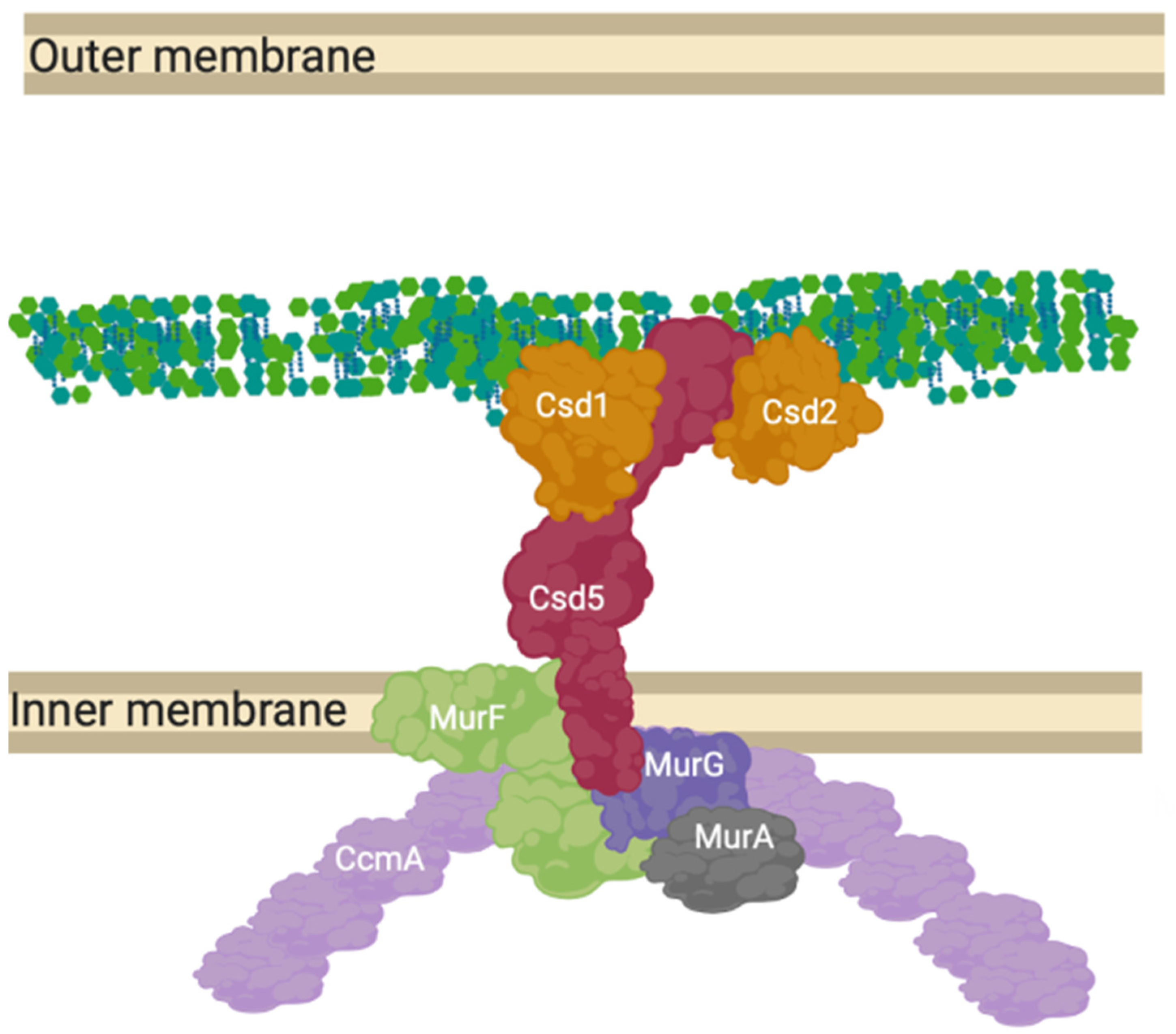
12. A “Shapeosome” Complex Synthesises Peptidoglycan in Curved Gram-Negatives
13. Unrelated Cell Envelope Proteins must Affect Peptidoglycan-Membrane Linkage
14. Review Summary
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
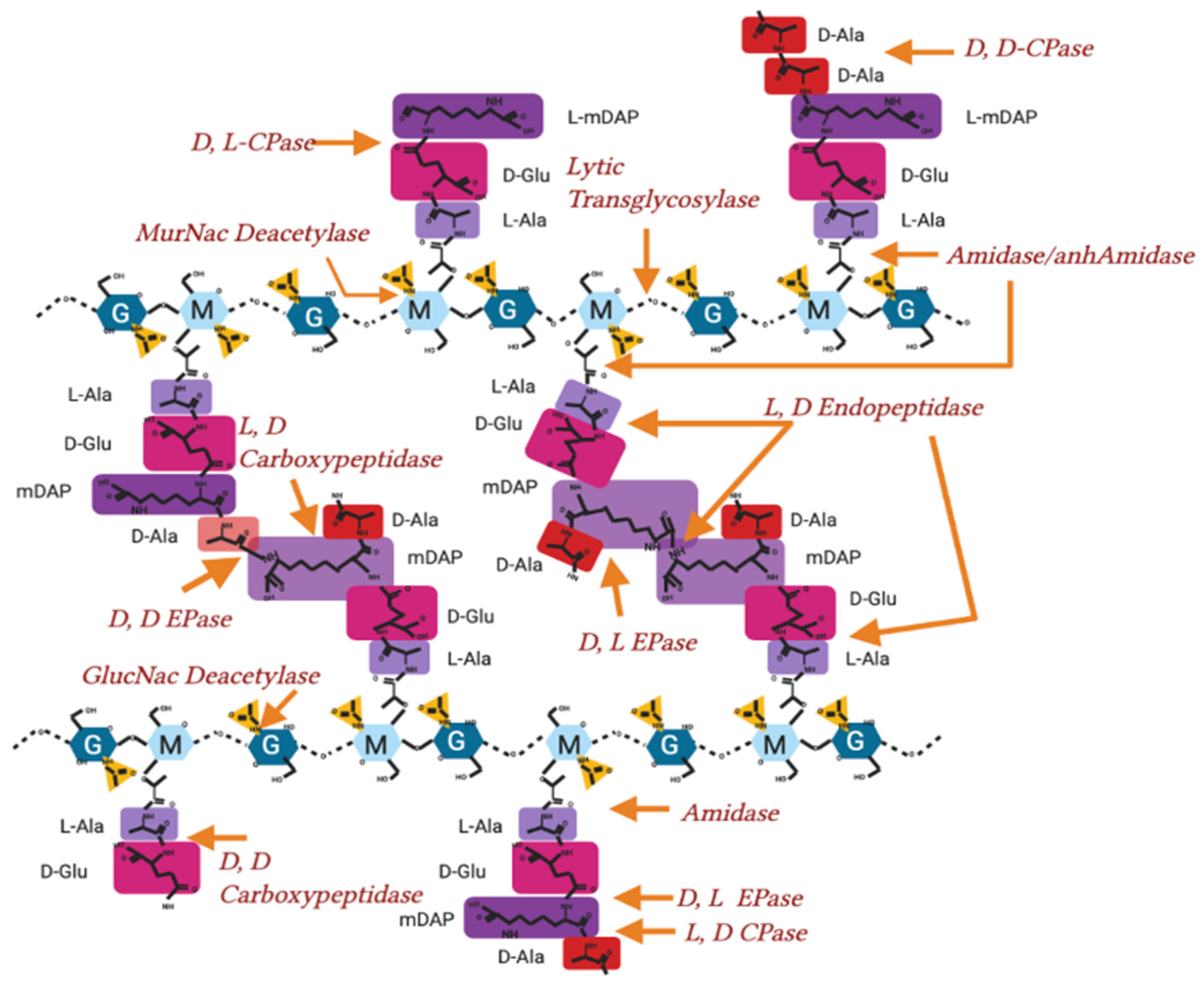
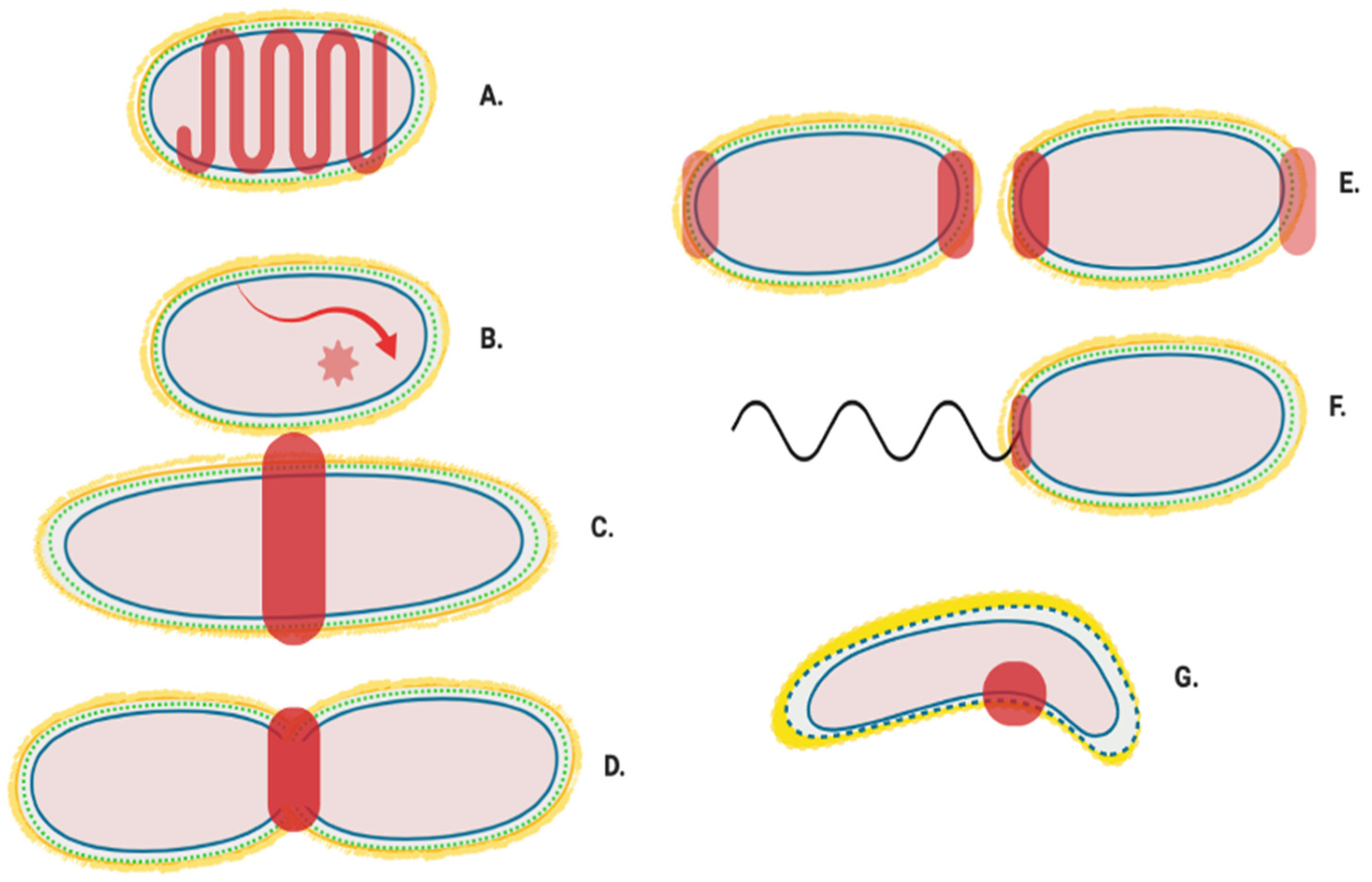
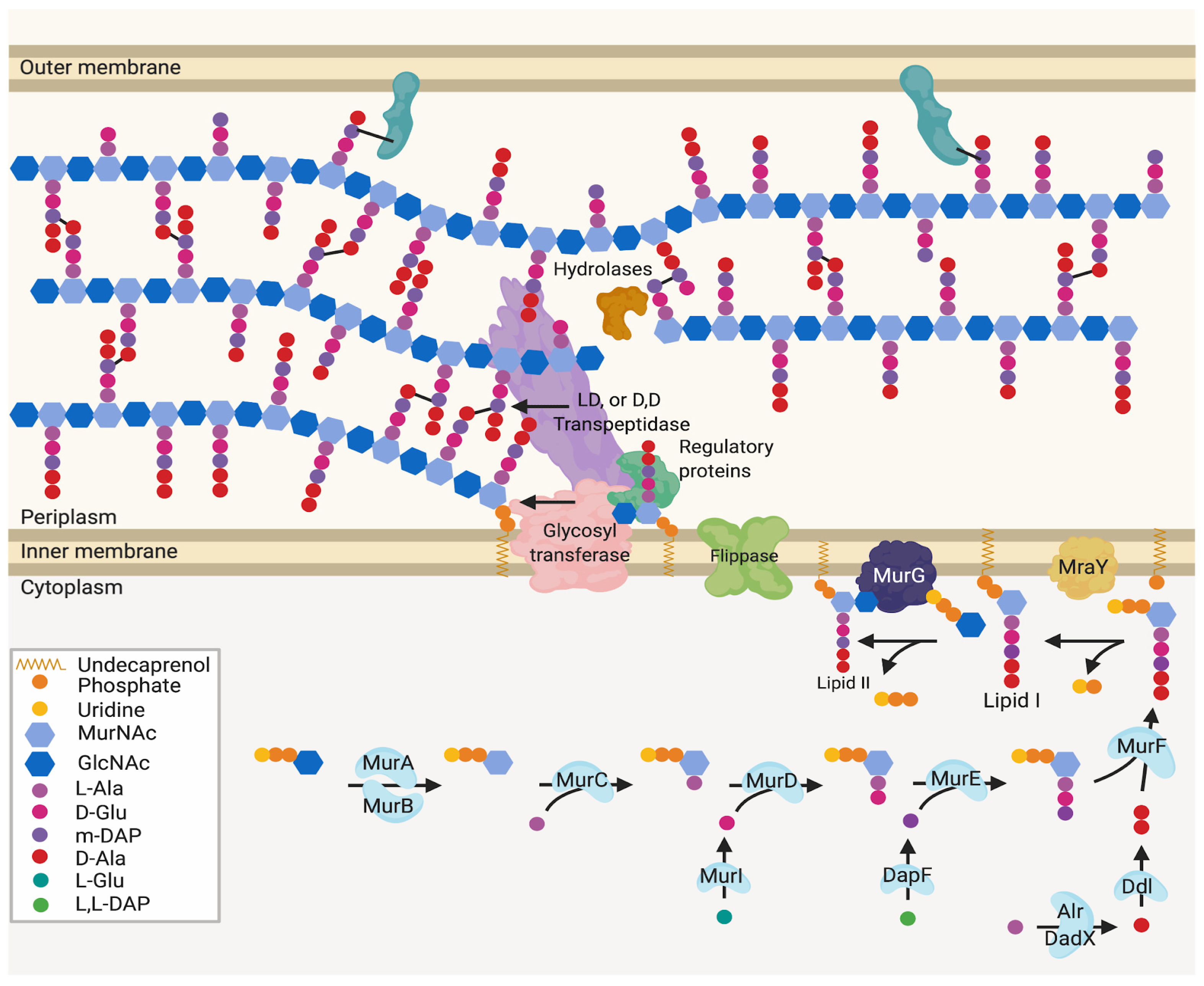
Appendix A.1. Mur Ligase Pathway
Appendix A.2. After Initial Synthesis, Peptidoglycan Is Modified

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptidoglycan Degradation/Hydrolases | |||
|---|---|---|---|
| Function | Enzymatic Action | Known Genes/Protein | References |
| D,D Carboxypeptidases | D-Ala D-Ala Cleavage 4-5 | dacA, yfeW, dacC, dacD, vanY, ampH, Csd3 * | [69,89,90,91] |
| MurNac de-Acetylase | Deacetylation of N-acetyl Muramic acid | pgdA | [92] |
| GlutNac de-acetylase | Deacetylation of N-acetyl Glucosamine | ||
| Amidase | Cleavage of peptide stem from Glycan strand | amiA, amiB, amiC, amiD, ampD, mpaA | [88,90,93,94] |
| Lytic Transglycocylase | Breaking Glycan strand at GlucNac-MurNac (endo) | Slt, MltA, MltB, MltC, MtD, MltE, PilT, traB, virB1, rlpA, MltG | [16,58,79,82,83,86,95,96] |
| Breaking Glycan strand at GlucNac-MurNac(exo) | NagZ | [97] | |
| L, D Carboxy/Endopeptidase | mDAP mDAP cleavage 3-3 | mepA | [98] |
| mDAP-Lpp Cleavage | YafK/LdtF | [80] | |
| mDAP D-Ala cleavage 3-4 | pgp2 *, csd6 * | [71,91] | |
| mDAP-D Glucosamine cleave 3-2 | csd4 * | [72] | |
| D, D Endopeptidase | Cleavage of D-Ala-mDAP crosslink 3-4 | dacB, pbpG, MepS, MepM, PBP7, MepH | [31,99] |
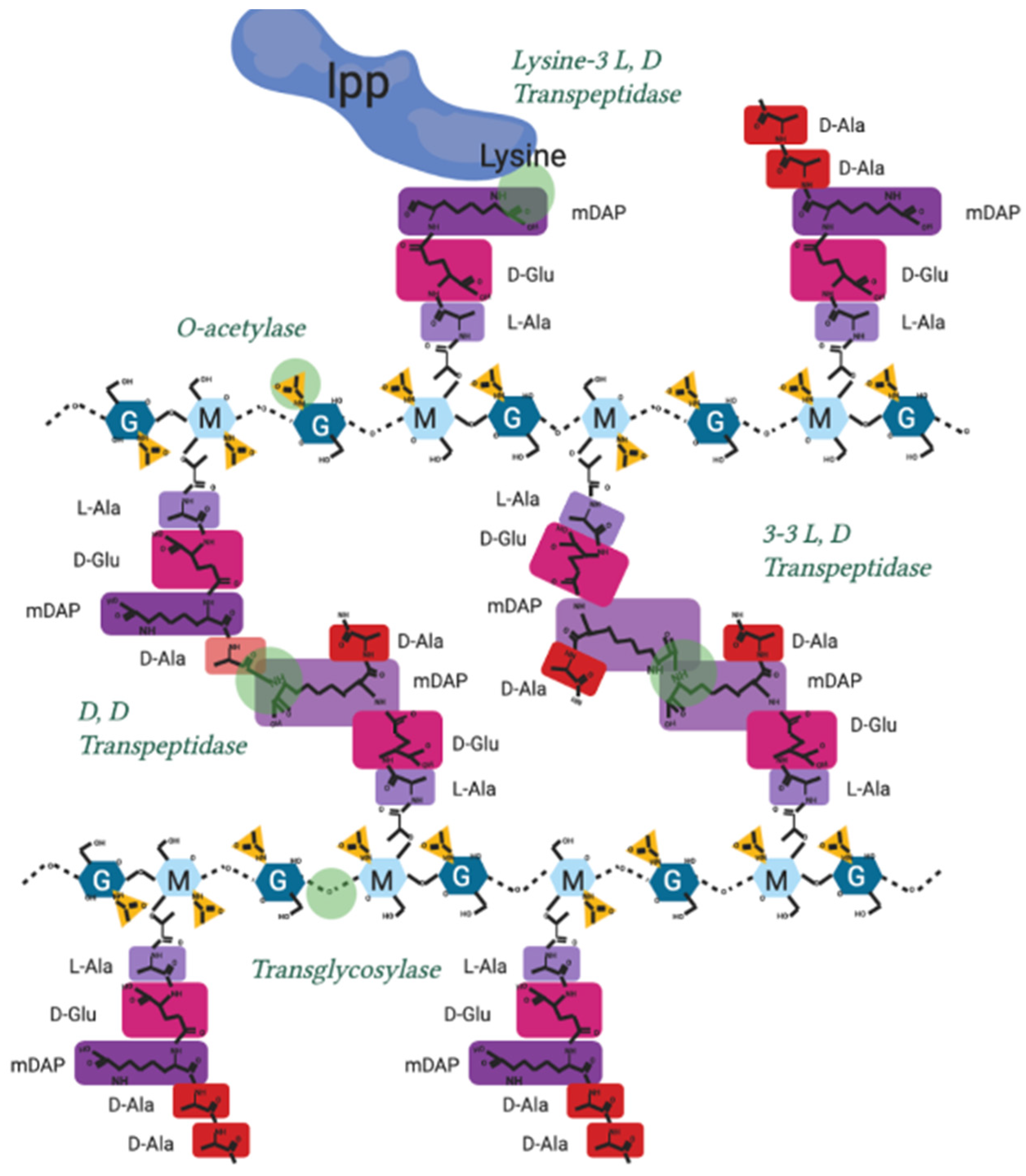
| Function | Enzymatic Action | Known Genes/Protein (E. coli) | References |
|---|---|---|---|
| D, D Transpeptidase and Transglycocylase | Adds lipid II to nascent strand and crosslinks into existing PG | mrcA/PBP1a | [29,100] |
| mrcB/PBP1b | [78] | ||
| D, D Transpeptidase | Crosslinks nascent strand into existing peptidoglycan | mrdA/PBP2, FtsI/PBP3 | [11,101] |
| Transglycosylase | Adds lipid II to nascent strand | mtgA, rodA, ftsW | [23,102,103] |
| Flippase | Flips Lipid II to periplasm | murJ | [104] |
| L, D Transpeptidases | Peptidoglycan Brauns lipoprotein crosslinkers | LdtA/ErfK, | [105] |
| YbiS/LdtB | |||
| Ycfs/LdtC | |||
| Peptidoglycan 3mDAP-3mDAP crosslinkers | YnhG/LdtE | ||
| YcbB/LdtD | |||
| O-acetylation | O-acetylates nam | oatA * | [2,92] |
| adr | |||
| pacA |
| Function | Known Genes/Protein (E. coli) | Reference |
|---|---|---|
| Moderate class A PBP activity | LpoA, LpoB | [9] |
| Alter interactor ability | CpoB | [9] |
| Bind OM with peptidoglycan | OmpA | [106] |
| Moderate OM linkage with peptidoglycan | lpp | [73] |
| Periskeletal elongasome component, moderates PBP2 activity | MreC | [23] |
| Periskeletal elongasome component, moderate PBP2 activity | MreD | |
| Treadmilling Cytoskeletal elongasome component | MreB | [15] |
| Elongasome staple component | RodZ | [57] |
| SPOR domain containing proteins, protein interaction | Rlpa, FtsN, DamX, SpoX | [61,85,107] |
| Hydrolase binding activity | NlpI | [55] |
| ActS | [108] | |
| NlpD | [109] | |
| EnvC | [93] | |
| Inner membrane peptidoglycan moderation | TolA, TolR, TolQ, palA, palB | [9,109] |
| Division moderation and EnvC control | FtsX, FtsE | [110] |
| Treadmilling cytoskeletal component for divisome | FtsZ | [44,56] |
| Division moderation | FtsA | [56] |
| Helicase and PBP interactor | FtsK | [58] |
| FtsN interactor and division start | FtsB, FtsL, FtsQ | [25] |
| FtsZ interactor and PIPs mediator | ZipA | [39] |
References
- Egan, A.J.; Cleverley, R.M.; Peters, K.; Lewis, R.J.; Vollmer, W. Regulation of bacterial cell wall growth. FEBS J. 2017, 284, 851–867. [Google Scholar] [CrossRef]
- Vollmer, W. Structural variation in the glycan strands of bacterial peptidoglycan. FEMS Microbiol. Rev. 2008, 32, 287–306. [Google Scholar] [CrossRef] [PubMed]
- Egan, A.J.; Errington, J.; Vollmer, W. Regulation of peptidoglycan synthesis and remodelling. Nat. Rev. Microbiol. 2020, 18, 446–460. [Google Scholar] [CrossRef] [PubMed]
- Barreteau, H.; Kovač, A.; Boniface, A.; Sova, M.; Gobec, S.; Blanot, D. Cytoplasmic steps of peptidoglycan biosynthesis. FEMS Microbiol. Rev. 2008, 32, 168–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernal-Cabas, M.; Ayala, J.A.; Raivio, T.L. The Cpx envelope stress response modifies peptidoglycan cross-linking via the L,D-transpeptidase LdtD and the novel protein YgaU. J. Bacteriol. 2015, 197, 603–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pazos, M.; Peters, K.; Vollmer, W. Robust peptidoglycan growth by dynamic and variable multi-protein complexes. Curr. Opin. Microbiol. 2017, 36, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Mueller, E.A.; Egan, A.J.; Breukink, E.; Vollmer, W.; Levin, P.A. Plasticity of Escherichia coli cell wall metabolism promotes fitness and antibiotic resistance across environmental conditions. eLife 2019, 8, e40754. [Google Scholar] [CrossRef] [PubMed]
- Lambert, C.; Chang, C.Y.; Capeness, M.J.; Sockett, R.E. The first bite—Profiling the predatosome in the bacterial pathogen Bdellovibrio. PLoS ONE 2010, 5, e8599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, A.N.; Egan, A.J.F.; van’t Veer, I.L.; Verheul, J.; Colavin, A.; Koumoutsi, A.; Biboy, J.; Altelaar, M.A.; Damen, M.J.; Huang, K.C.; et al. Coordination of peptidoglycan synthesis and outer membrane constriction during Escherichia coli cell division. eLife 2015, 4, e07118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.; Wivagg, C.N.; Kapoor, M.; Barry, Z.; Rohs, P.D.; Suh, H.; Marto, J.A.; Garner, E.C.; Bernhardt, T.G. Bacterial cell wall biogenesis is mediated by SEDS and PBP polymerase families functioning semi-Autonomously. Nat. Microbiol. 2016, 1, 16172. [Google Scholar] [CrossRef] [PubMed]
- Wollrab, E.; Özbaykal, G.; Vigouroux, A.; Cordier, B.; Simon, F.; Chaze, T.; Matondo, M.; van Teeffelen, S. Transpeptidase PBP2 governs initial localization and activity of major cell-wall synthesis machinery in Escherichia coli. bioRxiv 2019, 716407. [Google Scholar] [CrossRef] [Green Version]
- Vischer, N.O.; Verheul, J.; Postma, M.; van den Berg van Saparoea, B.; Galli, E.; Natale, P.; Gerdes, K.; Luirink, J.; Vollmer, W.; Vicente, M.; et al. Cell age dependent concentration of Escherichia coli divisome proteins analyzed with ImageJ and ObjectJ. Front. Microbiol. 2015, 6, 586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Ploeg, R.; Verheul, J.; Vischer, N.O.; Alexeeva, S.; Hoogendoorn, E.; Postma, M.; Banzhaf, M.; Vollmer, W.; Den Blaauwen, T. Colocalization and interaction between elongasome and divisome during a preparative cell division phase in Escherichia coli. Mol. Microbiol. 2013, 87, 1074–1087. [Google Scholar] [CrossRef]
- van der Ploeg, R.; Goudelis, S.T.; den Blaauwen, T. Validation of FRET assay for the screening of growth inhibitors of Escherichia coli reveals elongasome assembly dynamics. Int. J. Mol. Sci. 2015, 16, 17637–17654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniel, R.A.; Errington, J. Control of cell morphogenesis in bacteria: Two distinct ways to make a rod-shaped cell. Cell 2003, 113, 767–776. [Google Scholar] [CrossRef] [Green Version]
- Roure, S.; Bonis, M.; Chaput, C.; Ecobichon, C.; Mattox, A.; Barrière, C.; Geldmacher, N.; Guadagnini, S.; Schmitt, C.; Prévost, M.C.; et al. Peptidoglycan maturation enzymes affect flagellar functionality in bacteria. Mol. Microbiol. 2012, 86, 845–856. [Google Scholar] [CrossRef]
- Sycuro, L.K.; Pincus, Z.; Gutierrez, K.D.; Biboy, J.; Stern, C.A.; Vollmer, W.; Salama, N.R. Peptidoglycan Crosslinking Relaxation Promotes Helicobacter pylori’s Helical Shape and Stomach Colonization. Cell 2010, 141, 822–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blair, K. Exploring Mechanisms of Cell Shape Control in Helicobacter pylori. Ph.D Thesis, University of Washington, Seattle, WA, USA, 2018. [Google Scholar]
- Turner, R.D.; Vollmer, W.; Foster, S.J. Different walls for rods and balls: The diversity of peptidoglycan. Mol. Microbiol. 2014, 91, 862–874. [Google Scholar] [CrossRef] [Green Version]
- Turner, R.D.; Mesnage, S.; Hobbs, J.K.; Foster, S.J. Molecular imaging of glycan chains couples cell-wall polysaccharide architecture to bacterial cell morphology. Nat. Commun. 2018, 9, 1263. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Jericho, M.; Pink, D.; Beveridge, T. Thickness and elasticity of gram-negative murein sacculi measured by atomic force microscopy. J. Bacteriol. 1999, 181, 6865–6875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vollmer, W.; Blanot, D.; De Pedro, M.A. Peptidoglycan structure and architecture. FEMS Microbiol. Rev. 2008, 32, 149–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Biboy, J.; Vollmer, W.; den Blaauwen, T. MreC and MreD balance the interaction between the elongasome proteins PBP2 and RodA. PLoS Genet. 2020, 16, e1009276. [Google Scholar] [CrossRef] [PubMed]
- den Blaauwen, T.; Luirink, J. Checks and Balances in Bacterial Cell Division. MBio 2019, 10, e00149-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Typas, A.; Banzhaf, M.; van den Berg van Saparoea, B.; Verheul, J.; Biboy, J.; Nichols, R.J.; Zietek, M.; Beilharz, K.; Kannenberg, K.; von Rechenberg, M.; et al. Regulation of Peptidoglycan Synthesis by Outer-Membrane Proteins. Cell 2010, 143, 1097–1109. [Google Scholar] [CrossRef] [Green Version]
- Moynihan, P.J.; Clarke, A.J. O-Acetylated peptidoglycan: Controlling the activity of bacterial autolysins and lytic enzymes of innate immune systems. Int. J. Biochem. Cell Biol. 2011, 43, 1655–1659. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef]
- Zhang, B.; Horvath, S. A general framework for weighted gene co-expression network analysis. Stat. Appl. Genet. Mol. Biol. 2005, 4, 17. [Google Scholar] [CrossRef]
- Boes, A.; Olatunji, S.; Breukink, E.; Terrak, M. Regulation of the Peptidoglycan Polymerase Activity of PBP1b by Antagonist Actions of the Core Divisome Proteins FtsBLQ and FtsN. MBio 2019, 10, e01912-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohs, P.D.; Buss, J.; Sim, S.I.; Squyres, G.R.; Srisuknimit, V.; Smith, M.; Cho, H.; Sjodt, M.; Kruse, A.C.; Garner, E.C.; et al. A central role for PBP2 in the activation of peptidoglycan polymerization by the bacterial cell elongation machinery. PLoS Genet. 2018, 14, e1007726. [Google Scholar] [CrossRef] [PubMed]
- Sjodt, M.; Rohs, P.D.; Gilman, M.S.; Erlandson, S.C.; Zheng, S.; Green, A.G.; Brock, K.P.; Taguchi, A.; Kahne, D.; Walker, S.; et al. Structural coordination of polymerization and crosslinking by a SEDS–bPBP peptidoglycan synthase complex. Nat. Microbiol. 2020, 5, 813–820. [Google Scholar] [CrossRef]
- Meeske, A.J.; Riley, E.P.; Robins, W.P.; Uehara, T.; Mekalanos, J.J.; Kahne, D.; Walker, S.; Kruse, A.C.; Bernhardt, T.G.; Rudner, D.Z. SEDS proteins are a widespread family of bacterial cell wall polymerases. Nature 2016, 537, 634–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranjit, D.K.; Jorgenson, M.A.; Young, K.D. PBP1B glycosyltransferase and transpeptidase activities play different essential roles during the de novo regeneration of rod morphology in Escherichia coli. J. Bacteriol. 2017, 199, e00612-16. [Google Scholar] [CrossRef] [Green Version]
- Müller, P.; Ewers, C.; Bertsche, U.; Anstett, M.; Kallis, T.; Breukink, E.; Fraipont, C.; Terrak, M.; Nguyen-Distèche, M.; Vollmer, W. The essential cell division protein FtsN interacts with the murein (peptidoglycan) synthase PBP1B in Escherichia coli. J. Biol. Chem. 2007, 282, 36394–36402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; McQuillen, R.; Lyu, Z.; Phillips-Mason, P.; De La Cruz, A.; McCausland, J.W.; Liang, H.; DeMeester, K.E.; Santiago, C.C.; Grimes, C.L.; et al. A two-track model for the spatiotemporal coordination of bacterial septal cell wall synthesis revealed by single-molecule imaging of FtsW. Nat. Microbiol. 2021, 6, 584–593. [Google Scholar] [CrossRef]
- Kim, S.Y.; Gitai, Z.; Kinkhabwala, A.; Shapiro, L.; Moerner, W.E. Single molecules of the bacterial actin MreB undergo directed treadmilling motion in Caulobacter crescentus. Proc. Natl. Acad. Sci. USA 2006, 103, 10929–10934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, D.W.; Errington, J. Bacterial cell division: Assembly, maintenance and disassembly of the Z ring. Nat. Rev. Microbiol. 2009, 7, 642–653. [Google Scholar] [CrossRef]
- Lutkenhaus, J.; Pichoff, S.; Du, S. Bacterial cytokinesis: From Z ring to divisome. Cytoskeleton 2012, 69, 778–790. [Google Scholar] [CrossRef] [PubMed]
- Potluri, L.P.; Kannan, S.; Young, K.D. ZipA is required for FtsZ-dependent preseptal peptidoglycan synthesis prior to invagination during cell division. J. Bacteriol. 2012, 194, 5334–5342. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Galgoci, A.; Kodali, S.; Herath, K.B.; Jayasuriya, H.; Dorso, K.; Vicente, F.; González, A.; Cully, D.; Bramhill, D.; et al. Discovery of a Small Molecule that Inhibits Cell Division by Blocking FtsZ, a Novel Therapeutic Target of Antibiotics. J. Biol. Chem. 2003, 278, 44424–44428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Ithurbide, S.; Löwe, J.; Duggin, I.G. Two FtsZ proteins orchestrate archaeal cell division through distinct functions in ring assembly and constriction. bioRxiv 2021. [Google Scholar] [CrossRef]
- Van Teeffelen, S.; Gitai, Z. Rotate into shape: MreB and bacterial morphogenesis. EMBO J. 2011, 30, 4856–4857. [Google Scholar] [CrossRef] [PubMed]
- Bean, G.J.; Flickinger, S.T.; Westler, W.M.; McCully, M.E.; Sept, D.; Weibel, D.B.; Amann, K.J. A22 disrupts the bacterial actin cytoskeleton by directly binding and inducing a low-affinity state in MreB. Biochemistry 2009, 48, 4852–4857. [Google Scholar] [CrossRef] [Green Version]
- McCausland, J.W.; Yang, X.; Lyu, Z.; Söderström, B.; Xiao, J.; Liu, J. Treadmilling FtsZ polymers drive the directional movement of sPG-synthesis enzymes via Brownian ratchet mechanism. bioRxiv 2019. [Google Scholar] [CrossRef]
- Contreras-Martel, C.; Martins, A.; Ecobichon, C.; Trindade, D.M.; Matteï, P.J.; Hicham, S.; Hardouin, P.; El Ghachi, M.; Boneca, I.G.; Dessen, A. Molecular architecture of the PBP2-MreC core bacterial cell wall synthesis complex. Nat. Commun. 2017, 8, 776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banzhaf, M.; van den Berg van Saparoea, B.; Terrak, M.; Fraipont, C.; Egan, A.; Philippe, J.; Zapun, A.; Breukink, E.; Nguyen-Distèche, M.; den Blaauwen, T.; et al. Cooperativity of peptidoglycan synthases active in bacterial cell elongation. Mol. Microbiol. 2012, 85, 179–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Divakaruni, A.V.; Baida, C.; White, C.L.; Gober, J.W. The cell shape proteins MreB and MreC control cell morphogenesis by positioning cell wall synthetic complexes. Mol. Microbiol. 2007, 66, 174–188. [Google Scholar] [CrossRef]
- Martins, A.; Contreras-Martel, C.; Janet-Maitre, M.; Miyachiro, M.M.; Estrozi, L.F.; Trindade, D.M.; Malospirito, C.C.; Rodrigues-Costa, F.; Imbert, L.; Job, V.; et al. Self-association of MreC as a regulatory signal in bacterial cell wall elongation. Nat. Commun. 2021, 12, 2987. [Google Scholar] [CrossRef] [PubMed]
- Jean, N.L.; Bougault, C.M.; Lodge, A.; Derouaux, A.; Callens, G.; Egan, A.J.; Ayala, I.; Lewis, R.J.; Vollmer, W.; Simorre, J.P. Elongated structure of the outer-membrane activator of peptidoglycan synthesis LpoA: Implications for PBP1A stimulation. Structure 2014, 22, 1047–1054. [Google Scholar] [CrossRef] [Green Version]
- Paradis-Bleau, C.; Markovski, M.; Uehara, T.; Lupoli, T.J.; Walker, S.; Kahne, D.E.; Bernhardt, T.G. Lipoprotein Cofactors Located in the Outer Membrane Activate Bacterial Cell Wall Polymerases. Cell 2010, 143, 1110–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catherwood, A.C.; Lloyd, A.J.; Tod, J.A.; Chauhan, S.; Slade, S.E.; Walkowiak, G.P.; Galley, N.F.; Punekar, A.S.; Smart, K.; Rea, D.; et al. Substrate and Stereochemical Control of Peptidoglycan Cross-Linking by Transpeptidation by Escherichia coli PBP1B. J. Am. Chem. Soc. 2020, 142, 5034–5048. [Google Scholar] [CrossRef]
- White, C.L.; Kitich, A.; Gober, J.W. Positioning cell wall synthetic complexes by the bacterial morphogenetic proteins MreB and MreD. Mol. Microbiol. 2010, 76, 616–633. [Google Scholar] [CrossRef]
- Banzhaf, M.; Yau, H.C.; Verheul, J.; Lodge, A.; Kritikos, G.; Mateus, A.; Hov, A.K.; Stein, F.; Wartel, M.; Pazos, M.; et al. The outer membrane lipoprotein NlpI nucleates hydrolases within peptidoglycan multi-enzyme complexes in Escherichia coli. bioRxiv 2019, 609503. [Google Scholar] [CrossRef] [Green Version]
- Fenton, A.K.; Gerdes, K. Direct interaction of FtsZ and MreB is required for septum synthesis and cell division in Escherichia coli. EMBO J. 2013, 32, 1953–1965. [Google Scholar] [CrossRef] [PubMed]
- Schwechheimer, C.; Rodriguez, D.L.; Kuehn, M.J. NlpI-mediated modulation of outer membrane vesicle production through peptidoglycan dynamics in Escherichia coli. Microbiologyopen 2015, 4, 375–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bendezú, F.O.; Hale, C.A.; Bernhardt, T.G.; De Boer, P.A. RodZ (YfgA) is required for proper assembly of the MreB actin cytoskeleton and cell shape in E. coli. EMBO J. 2009, 28, 193–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berezuk, A.M.; Glavota, S.; Roach, E.J.; Goodyear, M.C.; Krieger, J.R.; Khursigara, C.M. Outer membrane lipoprotein RlpA is a novel periplasmic interaction partner of the cell division protein FtsK in Escherichia coli. Sci. Rep. 2018, 8, 12933. [Google Scholar] [CrossRef]
- Caveney, N.A.; Caballero, G.; Voedts, H.; Niciforovic, A.; Worrall, L.J.; Vuckovic, M.; Fonvielle, M.; Hugonnet, J.E.; Arthur, M.; Strynadka, N.C. Structural insight into YcbB-mediated beta-lactam resistance in Escherichia coli. Nat. Commun. 2019, 10, 1849. [Google Scholar] [CrossRef] [PubMed]
- Laddomada, F.; Miyachiro, M.M.; Jessop, M.; Patin, D.; Job, V.; Mengin-Lecreulx, D.; Le Roy, A.; Ebel, C.; Breyton, C.; Gutsche, I.; et al. The MurG glycosyltransferase provides an oligomeric scaffold for the cytoplasmic steps of peptidoglycan biosynthesis in the human pathogen Bordetella pertussis. Sci. Rep. 2019, 9, 4656. [Google Scholar] [CrossRef]
- Gerding, M.A.; Liu, B.; Bendezú, F.O.; Hale, C.A.; Bernhardt, T.G.; De Boer, P.A. Self-enhanced accumulation of FtsN at division sites and roles for other proteins with a SPOR domain (DamX, DedD, and RlpA) in Escherichia coli cell constriction. J. Bacteriol. 2009, 191, 7383–7401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petiti, M.; Serrano, B.; Faure, L.; Lloubes, R.; Mignot, T.; Duché, D. Tol Energy-Driven Localization of Pal and Anchoring to the Peptidoglycan Promote Outer-Membrane Constriction. J. Mol. Biol. 2019, 431, 3275–3288. [Google Scholar] [CrossRef]
- Bisson-Filho, A.W.; Hsu, Y.P.; Squyres, G.R.; Kuru, E.; Wu, F.; Jukes, C.; Sun, Y.; Dekker, C.; Holden, S.; VanNieuwenhze, M.S.; et al. Treadmilling by FtsZ filaments drives peptidoglycan synthesis and bacterial cell division. Science 2017, 355, 739–743. [Google Scholar] [CrossRef] [Green Version]
- Bendezú, F.O.; De Boer, P.A. Conditional lethality, division defects, membrane involution, and endocytosis in mre and mrd shape mutants of Escherichia coli. J. Bacteriol. 2008, 190, 1792–1811. [Google Scholar] [CrossRef] [Green Version]
- Pazos, M.; Peters, K.; Casanova, M.; Palacios, P.; VanNieuwenhze, M.; Breukink, E.; Vicente, M.; Vollmer, W. Z-ring membrane anchors associate with cell wall synthases to initiate bacterial cell division. Nat. Commun. 2018, 9, 5090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eberhardt, C.; Kuerschner, L.; Weiss, D.S. Probing the Catalytic Activity of a Cell Division-Specific Transpeptidase In Vivo with β-Lactams. J. Bacteriol. 2003, 185, 3726–3734. [Google Scholar] [CrossRef] [Green Version]
- Varma, A.; Young, K.D. In Escherichia coli, MreB and FtsZ direct the synthesis of lateral cell wall via independent pathways that require PBP 2. J. Bacteriol. 2009, 191, 3526–3533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dion, M.F.; Kapoor, M.; Sun, Y.; Wilson, S.; Ryan, J.; Vigouroux, A.; van Teeffelen, S.; Oldenbourg, R.; Garner, E.C. Bacillus subtilis cell diameter is determined by the opposing actions of two distinct cell wall synthetic systems. Nat. Microbiol. 2019, 4, 1294–1305. [Google Scholar] [CrossRef]
- Shiomi, D.; Toyoda, A.; Aizu, T.; Ejima, F.; Fujiyama, A.; Shini, T.; Kohara, Y.; Niki, H. Mutations in cell elongation genes mreB, mrdA and mrdB suppress the shape defect of RodZ-deficient cells. Mol. Microbiol. 2013, 87, 1029–1044. [Google Scholar] [CrossRef] [Green Version]
- Bernal-Cabas, M. The Cpx Pathway Causes Changes in the Peptidoglycan Structure, Turnover, and Recycling. Master’s Thesis, University of Alberta, Edmonton, AB, Canada, 2014. [Google Scholar]
- Hugonnet, J.E.; Mengin-Lecreulx, D.; Monton, A.; den Blaauwen, T.; Carbonnelle, E.; Veckerlé, C.; Yves, V.B.; van Nieuwenhze, M.; Bouchier, C.; Tu, K.; et al. Factors essential for L,D-transpeptidase-mediated peptidoglycan cross-linking and β-lactam resistance in Escherichia coli. eLife 2016, 5, e19469. [Google Scholar] [CrossRef]
- Mohammadi, T.; Karczmarek, A.; Crouvoisier, M.; Bouhss, A.; Mengin-Lecreulx, D.; Den Blaauwen, T. The essential peptidoglycan glycosyltransferase MurG forms a complex with proteins involved in lateral envelope growth as well as with proteins involved in cell division in Escherichia coli. Mol. Microbiol. 2007, 65, 1106–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.C.; Blair, K.M.; Salama, N.R. Staying in Shape: The Impact of Cell Shape on Bacterial Survival in Diverse Environments. Microbiol. Mol. Biol. Rev. 2016, 80, 187–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frirdich, E.; Vermeulen, J.; Biboy, J.; Soares, F.; Taveirne, M.E.; Johnson, J.G.; DiRita, V.J.; Girardin, S.E.; Vollmer, W.; Gaynor, E.C. Peptidoglycan LD-carboxypeptidase Pgp2 influences Campylobacter jejuni helical cell shape and pathogenic properties and provides the substrate for the DL-carboxypeptidase Pgp1. J. Biol. Chem. 2014, 289, 8007–8018. [Google Scholar] [CrossRef] [Green Version]
- Blair, K.M.; Mears, K.S.; Taylor, J.A.; Fero, J.; Jones, L.A.; Gafken, P.R.; Whitney, J.C.; Salama, N.R. The Helicobacter pylori cell shape promoting protein Csd5 interacts with the cell wall, MurF, and the bacterial cytoskeleton. Mol. Microbiol. 2018, 110, 114–127. [Google Scholar] [CrossRef]
- Samsudin, F.; Boags, A.; Piggot, T.J.; Khalid, S. Braun’s Lipoprotein Facilitates OmpA Interaction with the Escherichia coli Cell Wall. Biophys. J. 2017, 113, 1496–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, V.; Sieglin, U. The Covalent Murein-Lipoprotin Structure of the Escherichia coli Cell Wall: The Attachment Site of the Lipoprotein on the Murein. Eur. J. Biochem. 1970, 13, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Zahrl, D.; Wagner, M.; Bischof, K.; Bayer, M.; Zavecz, B.; Beranek, A.; Ruckenstuhl, C.; Zarfel, G.E.; Koraimann, G. Peptidoglycan degradation by specialized lytic transglycosylases associated with type III and type IV secretion systems. Microbiology 2005, 151, 3455–3467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.C.; Lin, C.N.; Wang, S.H.; Ng, S.C.; Hu, W.S.; Syu, W.J. A putative lytic transglycosylase tightly regulated and critical for the EHEC type three secretion. J. Biomed. Sci. 2010, 17, 52. [Google Scholar] [CrossRef] [Green Version]
- Sandoz, K.M.; Moore, R.A.; Beare, P.A.; Patel, A.V.; Smith, R.E.; Bern, M.; Hwang, H.; Cooper, C.J.; Priola, S.A.; Parks, J.M.; et al. β-Barrel proteins tether the outer membrane in many Gram-negative bacteria. Nat. Microbiol. 2021, 6, 19–26. [Google Scholar] [CrossRef]
- Turner, R.D.; Hurd, A.F.; Cadby, A.; Hobbs, J.K.; Foster, S.J. Cell wall elongation mode in Gram-negative bacteria is determined by peptidoglycan architecture. Nat. Commun. 2013, 4, 1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahadur, R.; Chodisetti, P.K.; Reddy, M. Cleavage of Braun’s lipoprotein Lpp from the bacterial peptidoglycan by a paralog of L,D-transpeptidases, LdtF. Proc. Natl. Acad. Sci. USA 2021, 118, e2101989118. [Google Scholar] [CrossRef]
- Kaplan, E.; Greene, N.P.; Crow, A.; Koronakis, V. Insights into bacterial lipoprotein trafficking from a structure of LolA bound to the LolC periplasmic domain. Proc. Natl. Acad. Sci. USA 2018, 115, E7389–E7397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isom, G.L.; Davies, N.J.; Chong, Z.S.; Bryant, J.A.; Jamshad, M.; Sharif, M.; Cunningham, A.F.; Knowles, T.J.; Chng, S.S.; Cole, J.A.; et al. MCE domain proteins: Conserved inner membrane lipid-binding proteins required for outer membrane homeostasis. Sci. Rep. 2017, 7, 8608. [Google Scholar] [CrossRef] [PubMed]
- Yahashiri, A.; Jorgenson, M.A.; Weiss, D.S. Bacterial SPOR domains are recruited to septal peptidoglycan by binding to glycan strands that lack stem peptides. Proc. Natl. Acad. Sci. USA 2015, 112, 11347–11352. [Google Scholar] [CrossRef] [Green Version]
- Born, P.; Breukink, E.; Vollmer, W. In vitro synthesis of cross-linked murein and its attachment to sacculi by PBP1A from Escherichia coli. J. Biol. Chem. 2006, 281, 26985–26993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidrich, C.; Ursinus, A.; Berger, J.; Schwarz, H.; Höltje, J.V. Effects of multiple deletions of murein hydrolases on viability, septum cleavage, and sensitivity to large toxic molecules in Escherichia coli. J. Bacteriol. 2002, 184, 6093–6099. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.M.; Otten, C.; Biboy, J.; Breukink, E.; Van Nieuwenhze, M.; Vollmer, W.; den Blaauwen, T. The fluorescent D-Amino Acid NADA as a tool to study the conditional activity of transpeptidases in Escherichia coli. Front. Microbiol. 2018, 9, 2101. [Google Scholar] [CrossRef]
- Scheurwater, E.; Reid, C.W.; Clarke, A.J. Lytic transglycosylases: Bacterial space-making autolysins. Int. J. Biochem. Cell Biol. 2008, 40, 586–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vollmer, W. Structure and Biosynthesis of the Murein (Peptidoglycan) Sacculus. In The Periplasm; ASM Press: Washington, DC, USA, 2014; Chapter 11. [Google Scholar] [CrossRef]
- Morè, N.; Martorana, A.M.; Biboy, J.; Otten, C.; Winkle, M.; Serrano, C.K.; Montón Silva, A.; Atkinson, L.; Yau, H.; Breukink, E.; et al. Peptidoglycan remodeling enables Escherichia coli to survive severe outer membrane assembly defect. MBio 2019, 10, e02729-18. [Google Scholar] [CrossRef] [Green Version]
- Lambert, C.; Lerner, T.R.; Bui, N.K.; Somers, H.; Aizawa, S.I.; Liddell, S.; Clark, A.; Vollmer, W.; Lovering, A.L.; Sockett, R.E. Interrupting peptidoglycan deacetylation during Bdellovibrio predator-prey interaction prevents ultimate destruction of prey wall, liberating bacterial-ghosts. Sci. Rep. 2016, 6, 26010. [Google Scholar] [CrossRef] [Green Version]
- Meisel, U.; Höltje, J.V.; Vollmer, W. Overproduction of inactive variants of the murein synthase PBP1B causes lysis in Escherichia coli. J. Bacteriol. 2003, 185, 5342–5348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vermassen, A.; Leroy, S.; Talon, R.; Provot, C.; Popowska, M.; Desvaux, M. Cell Wall Hydrolases in Bacteria: Insight on the Diversity of Cell Wall Amidases, Glycosidases and Peptidases Toward Peptidoglycan. Front. Microbiol. 2019, 10, 331. [Google Scholar] [CrossRef] [PubMed]
- Ahangar, M.S.; Furze, C.M.; Guy, C.S.; Cooper, C.; Maskew, K.S.; Graham, B.; Cameron, A.D.; Fullam, X.E. Structural and functional determination of homologs of the Mycobacterium tuberculosis N-acetylglucosamine-6-phosphate deacetylase (NagA). J. Biol. Chem. 2018, 293, 9770–9783. [Google Scholar] [CrossRef] [Green Version]
- van Heijenoort, J. Peptidoglycan Hydrolases of Escherichia coli. Microbiol. Mol. Biol. Rev. 2011, 75, 636–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sycuro, L.K.; Rule, C.S.; Petersen, T.W.; Wyckoff, T.J.; Sessler, T.; Nagarkar, D.B.; Khalid, F.; Pincus, Z.; Biboy, J.; Vollmer, W.; et al. Flow cytometry-based enrichment for cell shape mutants identifies multiple genes that influence Helicobacter pylori morphology. Mol. Microbiol. 2013, 90, 869–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rae, C.S.; Geissler, A.; Adamson, P.C.; Portnoy, D.A. Mutations of the Listeria monocytogenes peptidoglycan N-Deacetylase and O-acetylase result in enhanced lysozyme sensitivity, bacteriolysis, and hyperinduction of innate immune pathways. Infect. Immun. 2011, 79, 3596–3606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocaboy, M.; Herman, R.; Sauvage, E.; Remaut, H.; Moonens, K.; Terrak, M.; Charlier, P.; Kerff, F. The crystal structure of the cell division amidase amic reveals the fold of the AMIN domain, a new peptidoglycan binding domain. Mol. Microbiol. 2013, 90, 267–277. [Google Scholar] [CrossRef] [Green Version]
- Bernhardt, T.G.; De Boer, P.A. The Escherichia coli amidase AmiC is a periplasmic septal ring component exported via the twin-arginine transport pathway. Mol. Microbiol. 2003, 48, 1171–1182. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.C.; Tan, K.; Joachimiak, A.; Bernhardt, T.G. A conformational switch controls cell wall-remodelling enzymes required for bacterial cell division. Mol. Microbiol. 2012, 85, 768–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derouaux, A.; Wolf, B.; Fraipont, C.; Breukink, E.; Nguyen-Distèche, M.; Terrak, M. The monofunctional glycosyltransferase of Escherichia coli localizes to the cell division site and interacts with penicillin-binding protein 3, FtsW, and FtsN. J. Bacteriol. 2008, 190, 1831–1834. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, T.; Van Dam, V.; Sijbrandi, R.; Vernet, T.; Zapun, A.; Bouhss, A.; Diepeveen-De Bruin, M.; Nguyen-Distéche, M.; De Kruijff, B.; Breukink, E. Identification of FtsW as a transporter of lipid-linked cell wall precursors across the membrane. EMBO J. 2011, 30, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Crépin, S.; Ottosen, E.N.; Peters, K.; Smith, S.N.; Himpsl, S.D.; Vollmer, W.; Mobley, H.L. The lytic transglycosylase MltB connects membrane homeostasis and in vivo fitness of Acinetobacter baumannii. Mol. Microbiol. 2018, 109, 745–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moynihan, P.J.; Cadby, I.T.; Veerapen, N.; Jankute, M.; Crosatti, M.; Mukamolova, G.V.; Lovering, A.L.; Besra, G.S. The hydrolase LpqI primes mycobacterial peptidoglycan recycling. Nat. Commun. 2019, 10, 2647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keck, W.; Van Leeuwen, A.M.; Huber, M.; Goodell, E.W. Cloning and characterization of mepA, the structural gene of the penicillin-insensitive murein endopeptidase from Escherichia coli. Mol. Microbiol. 1990, 4, 209–219. [Google Scholar] [CrossRef]
- Liu, B.; Hale, C.A.; Persons, L.; Phillips-Mason, P.J.; de Boer, P.A. Roles of the DedD Protein in Escherichia coli Cell Constriction. J. Bacteriol. 2019, 201, e00698-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcorlo, M.; Martínez-Caballero, S.; Molina, R.; Hermoso, J.A. Carbohydrate recognition and lysis by bacterial peptidoglycan hydrolases. Curr. Opin. Struct. Biol. 2017, 44, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Kocaoglu, O.; Carlson, E.E. Profiling of β-lactam selectivity for penicillin-binding proteins in Escherichia coli strain DC2. Antimicrob. Agents Chemother. 2015, 59, 2785–2790. [Google Scholar] [CrossRef] [Green Version]
- Egan, A.J.; Biboy, J.; van’t Veer, I.; Breukink, E.; Vollmer, W. Activities and regulation of peptidoglycan synthases. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20150031. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Graham, C.L.B.; Newman, H.; Gillett, F.N.; Smart, K.; Briggs, N.; Banzhaf, M.; Roper, D.I. A Dynamic Network of Proteins Facilitate Cell Envelope Biogenesis in Gram-Negative Bacteria. Int. J. Mol. Sci. 2021, 22, 12831. https://doi.org/10.3390/ijms222312831
Graham CLB, Newman H, Gillett FN, Smart K, Briggs N, Banzhaf M, Roper DI. A Dynamic Network of Proteins Facilitate Cell Envelope Biogenesis in Gram-Negative Bacteria. International Journal of Molecular Sciences. 2021; 22(23):12831. https://doi.org/10.3390/ijms222312831
Chicago/Turabian StyleGraham, Chris L. B., Hector Newman, Francesca N. Gillett, Katie Smart, Nicholas Briggs, Manuel Banzhaf, and David I. Roper. 2021. "A Dynamic Network of Proteins Facilitate Cell Envelope Biogenesis in Gram-Negative Bacteria" International Journal of Molecular Sciences 22, no. 23: 12831. https://doi.org/10.3390/ijms222312831
APA StyleGraham, C. L. B., Newman, H., Gillett, F. N., Smart, K., Briggs, N., Banzhaf, M., & Roper, D. I. (2021). A Dynamic Network of Proteins Facilitate Cell Envelope Biogenesis in Gram-Negative Bacteria. International Journal of Molecular Sciences, 22(23), 12831. https://doi.org/10.3390/ijms222312831