
The Longitudinal Dividing Bacterium Candidatus Thiosymbion Oneisti Has a Natural Temperature-Sensitive FtsZ Protein with Low GTPase Activity



Abstract
:1. Introduction
2. Results
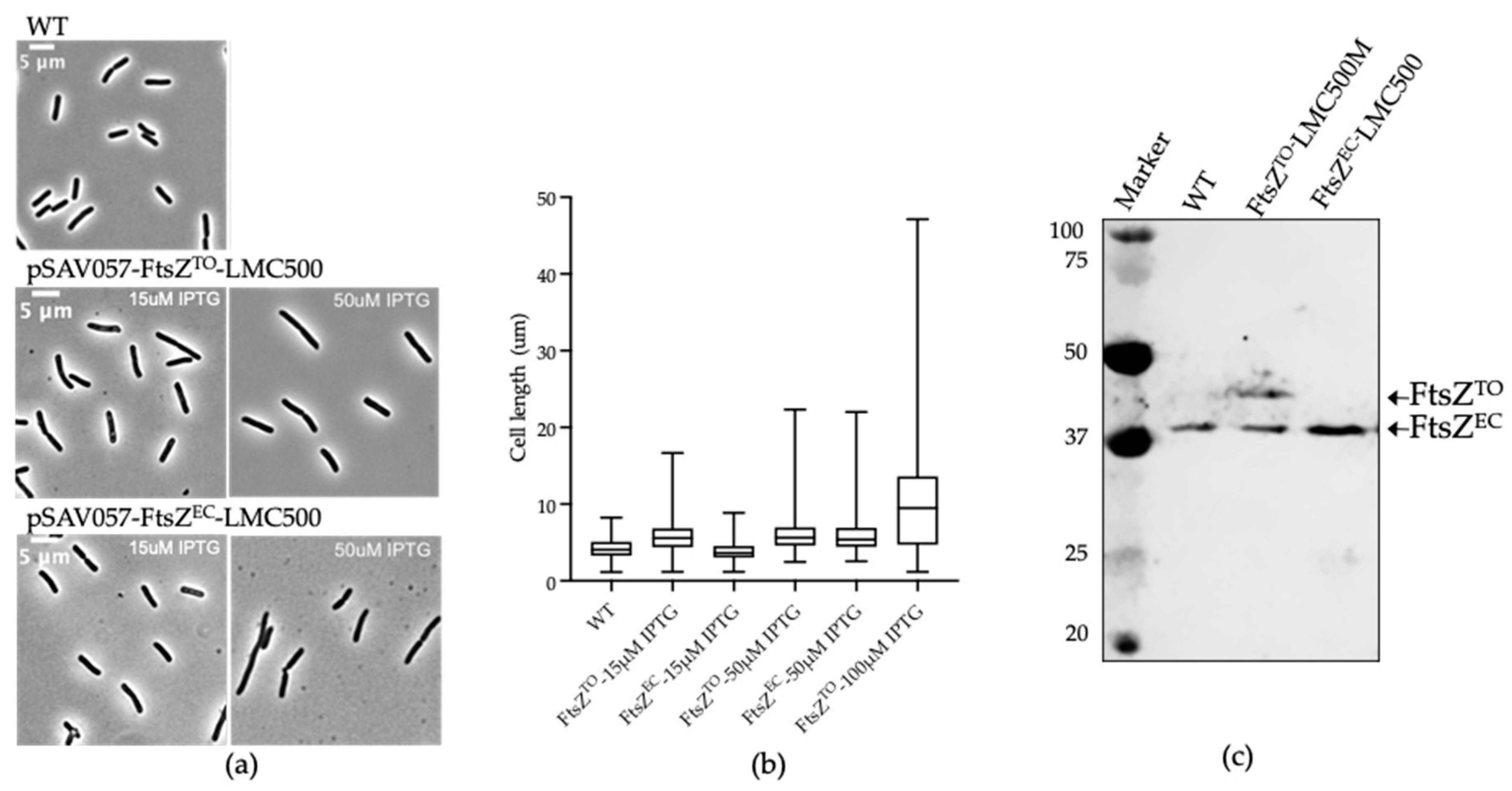
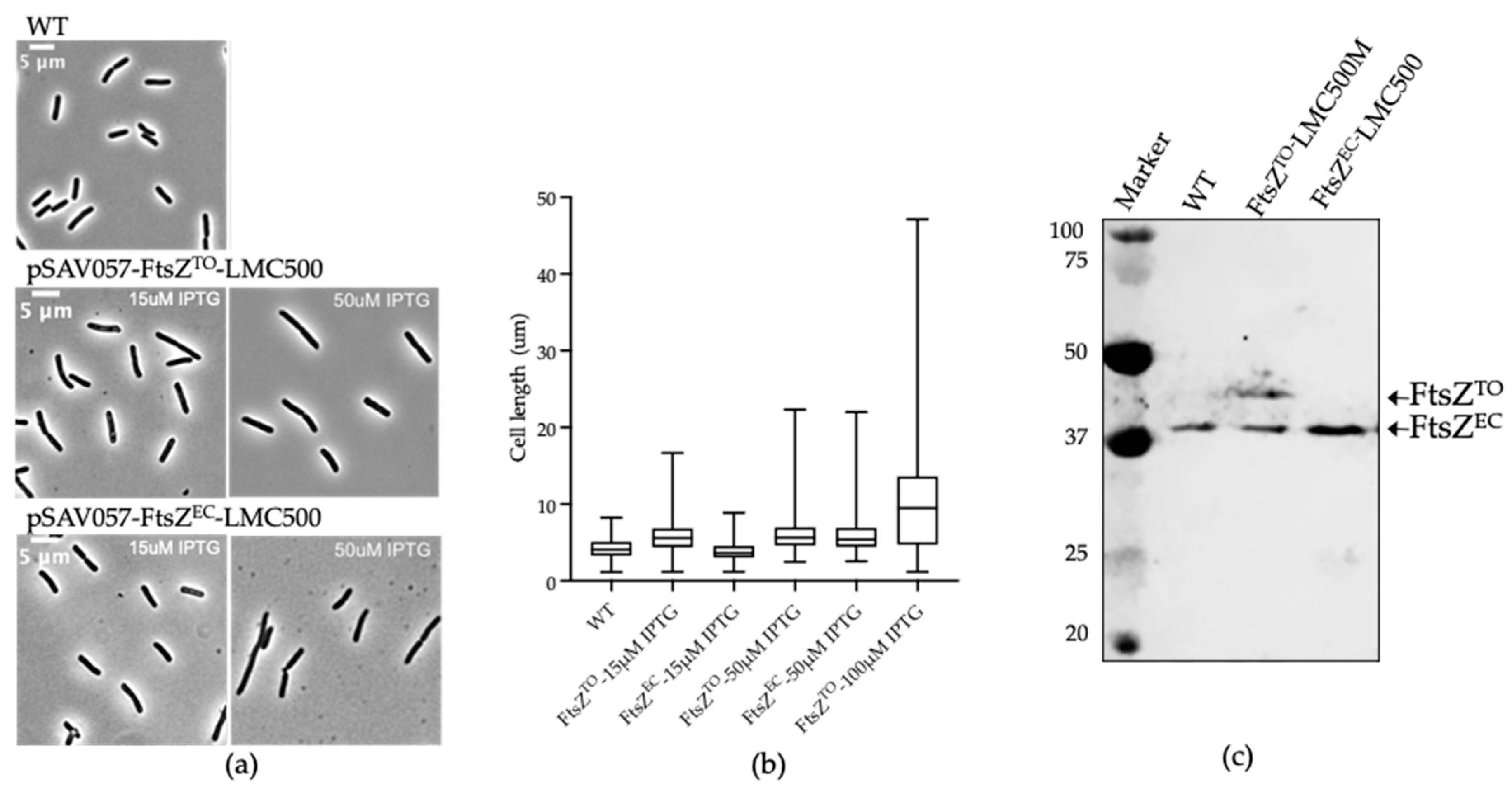
2.1. FtsZTO Interferes with Cell Division in E. coli and Overexpression of FtsZTO Results in Filamentous Cells
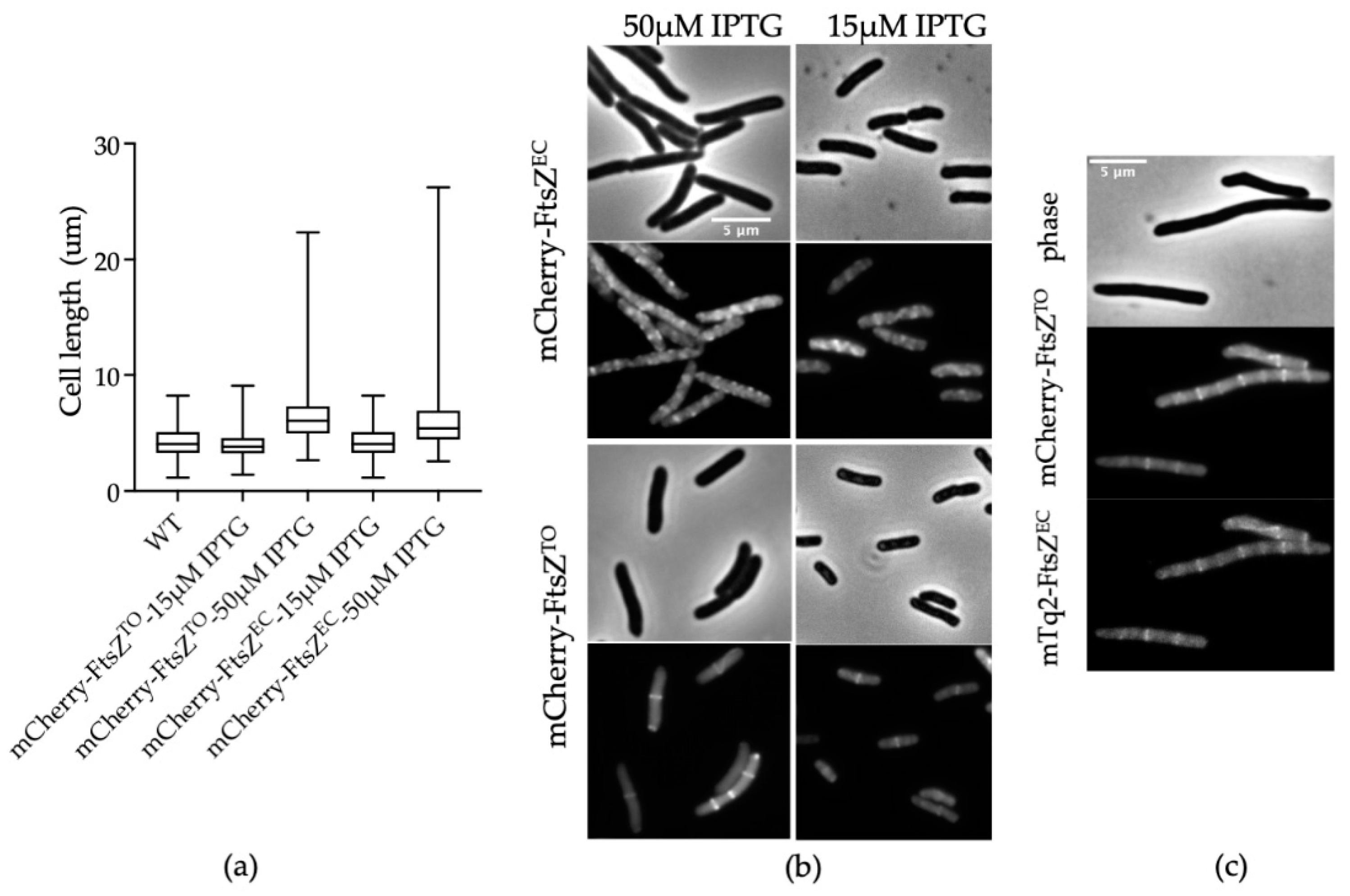
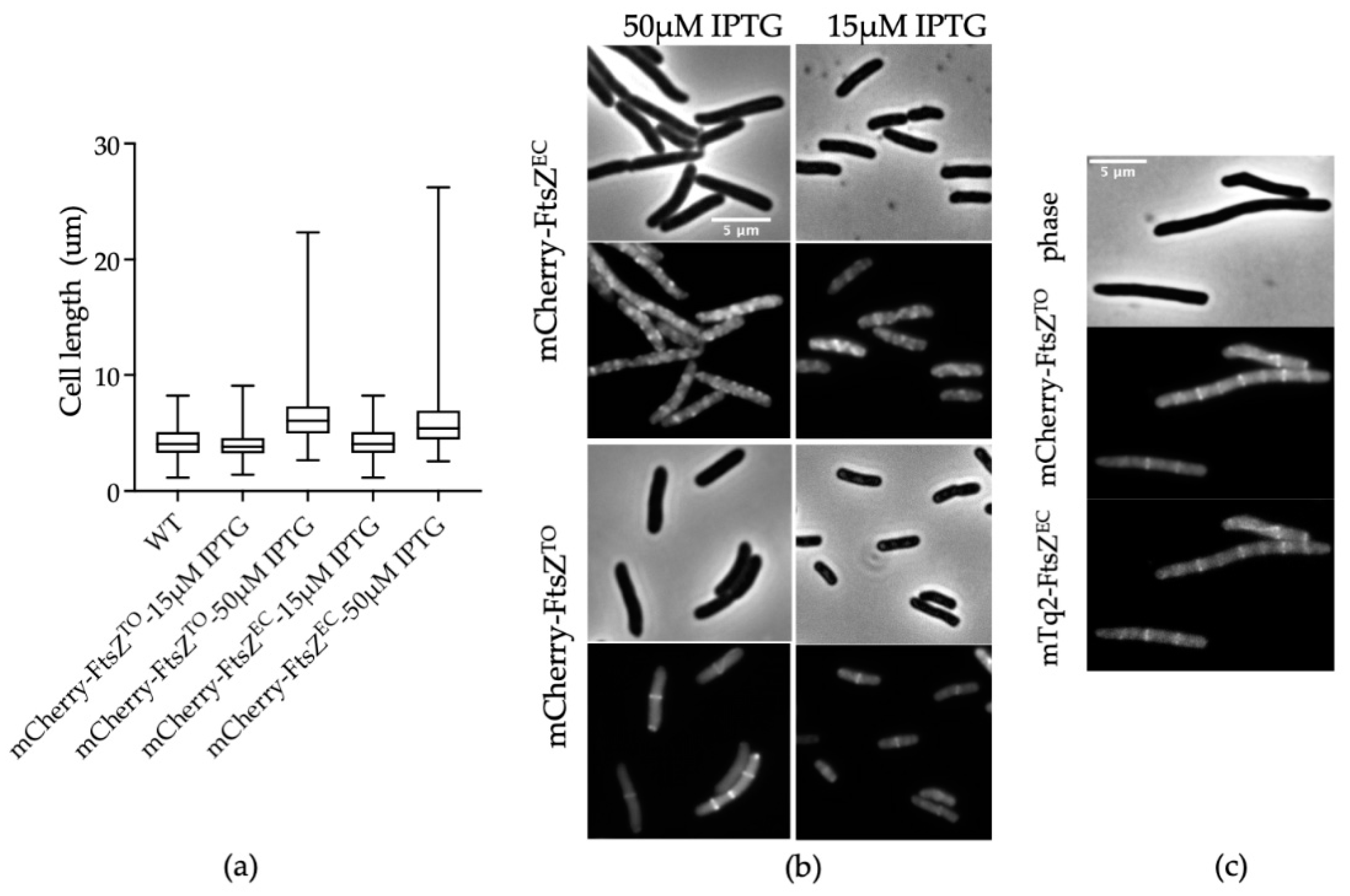
2.2. FtsZTO Colocalizes with FtsZEC Rings in E. coli
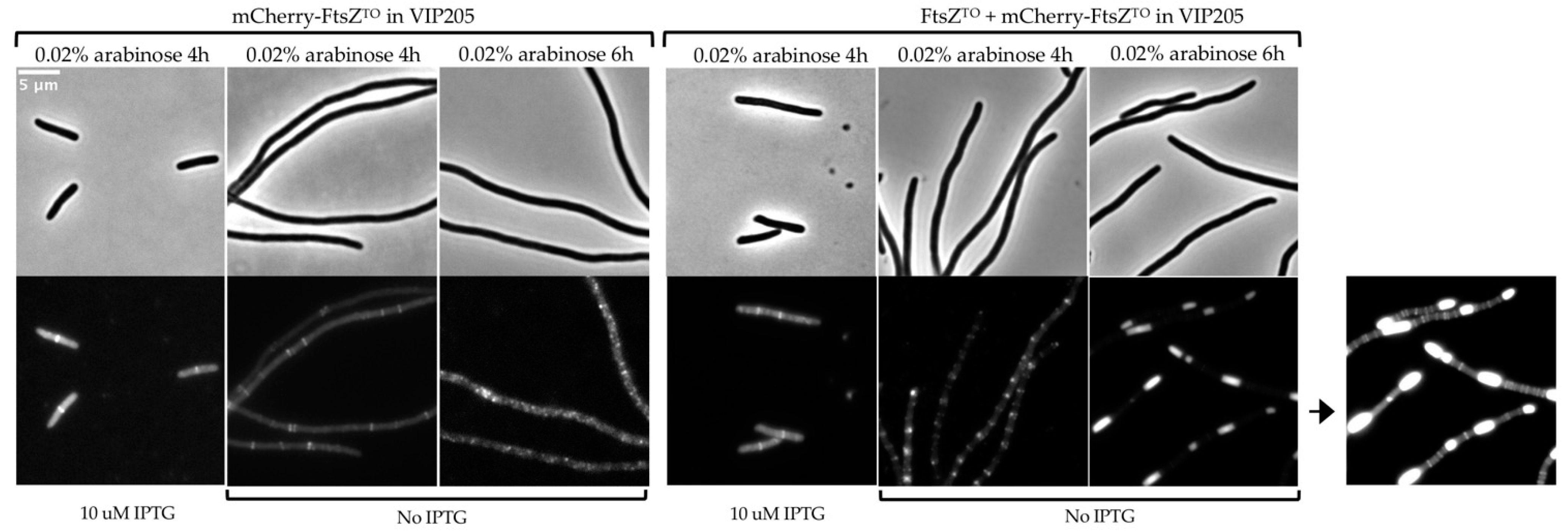
2.3. FtsZTO Forms Ring- and Barrel-like Structures Independently of FtsZEC in the E. coli FtsZ Depletion Strain VIP205
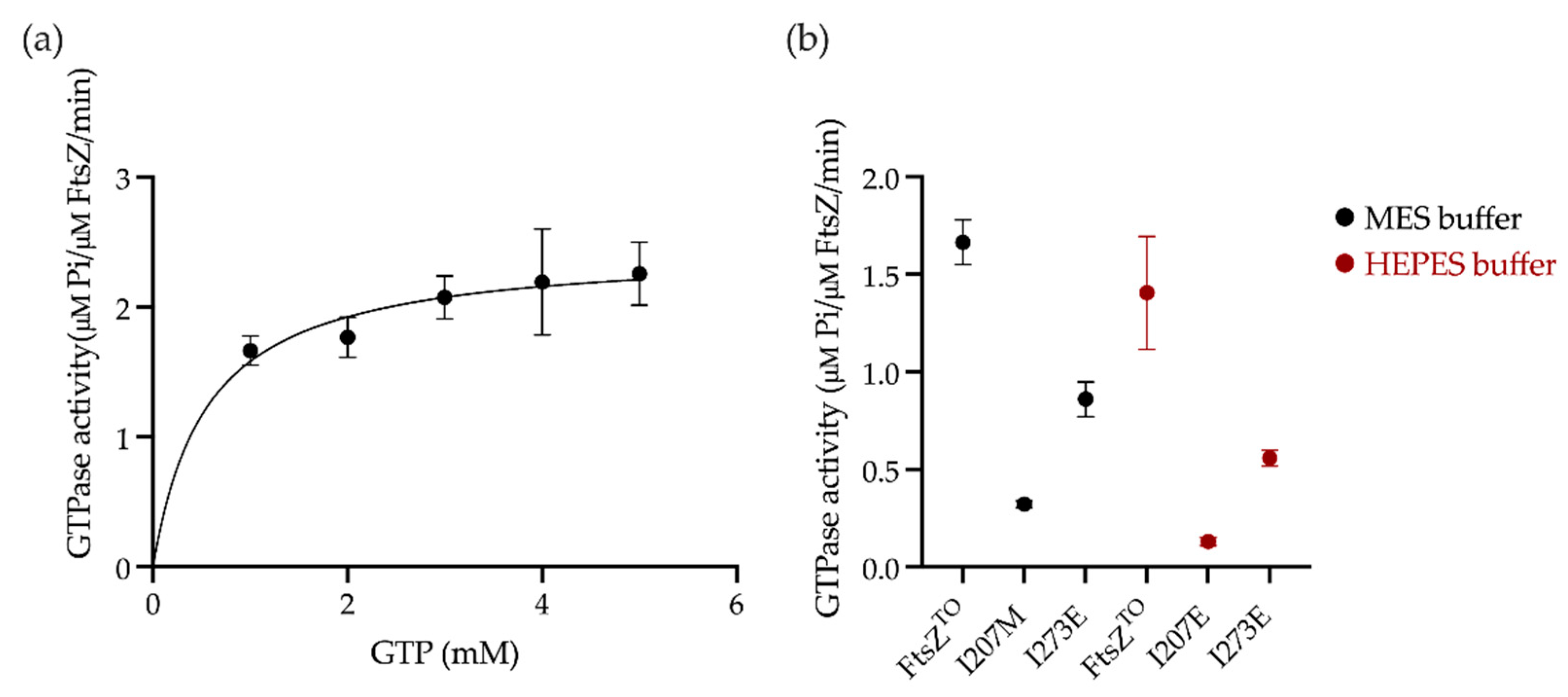
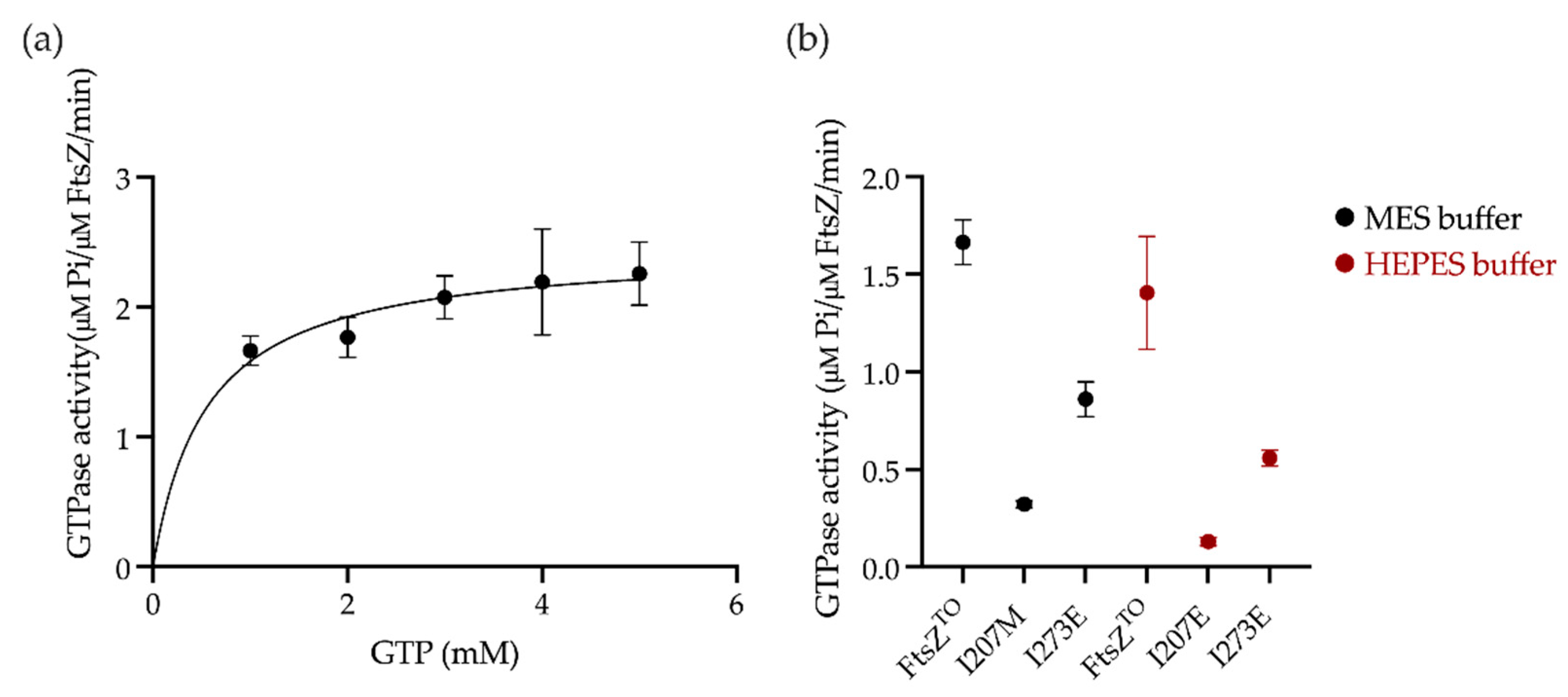
2.4. GTPase Activity Assay of FtsZTO In Vitro
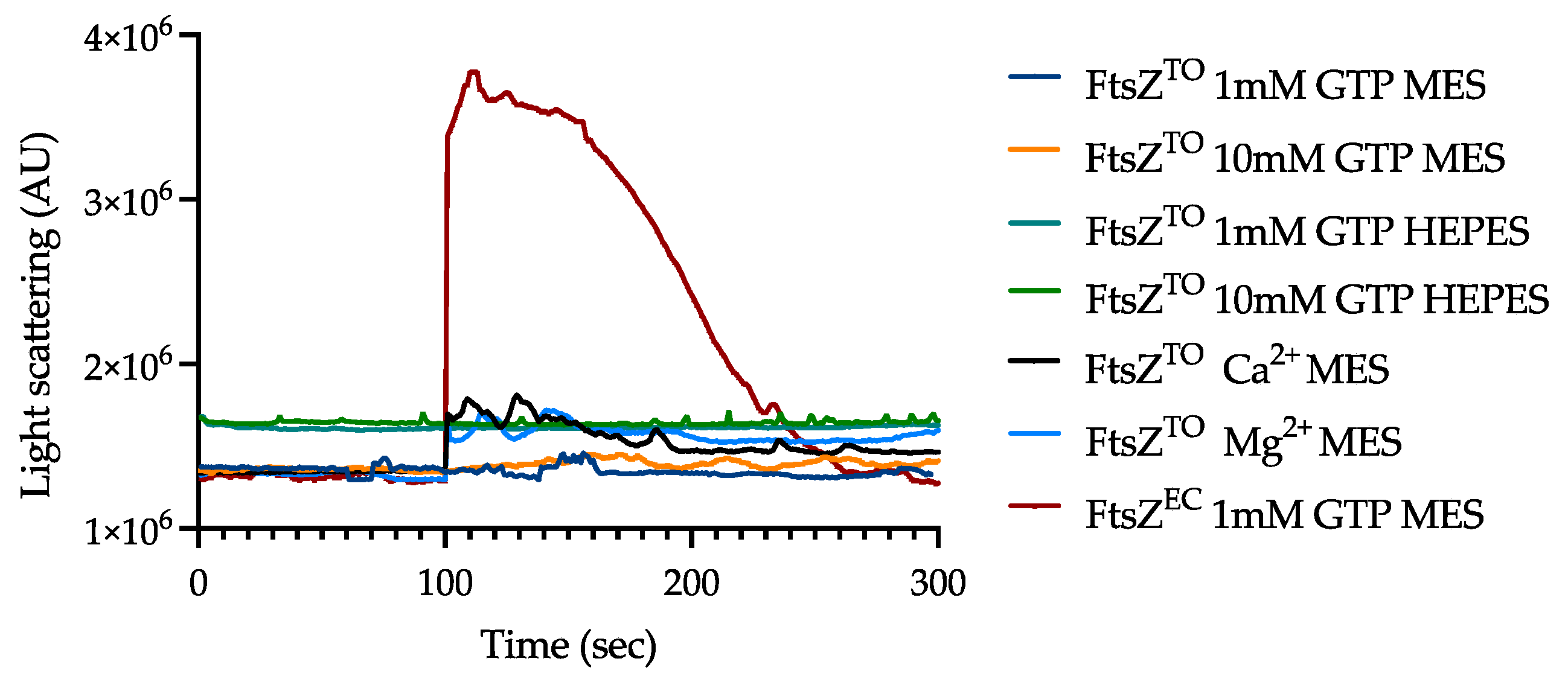
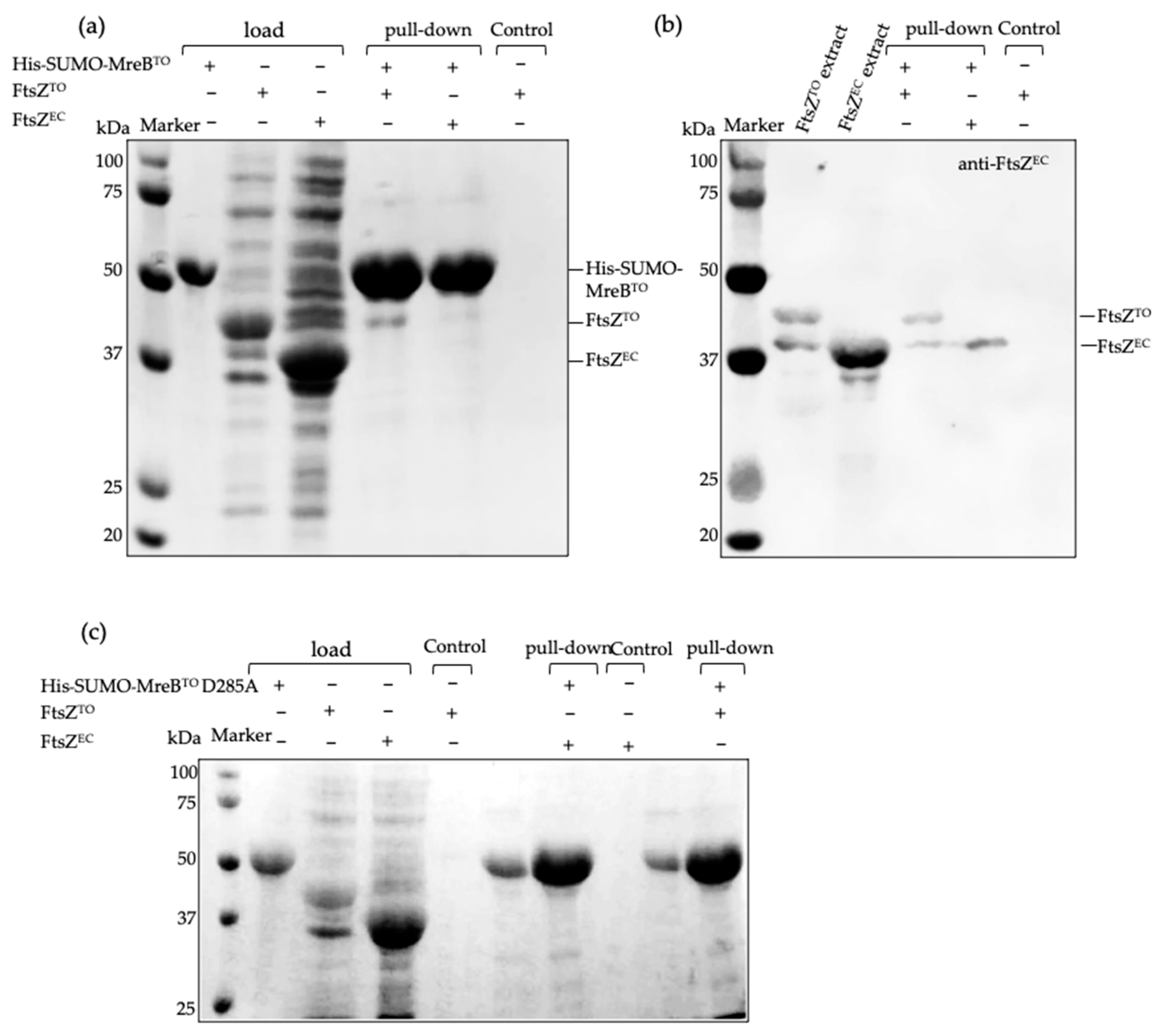
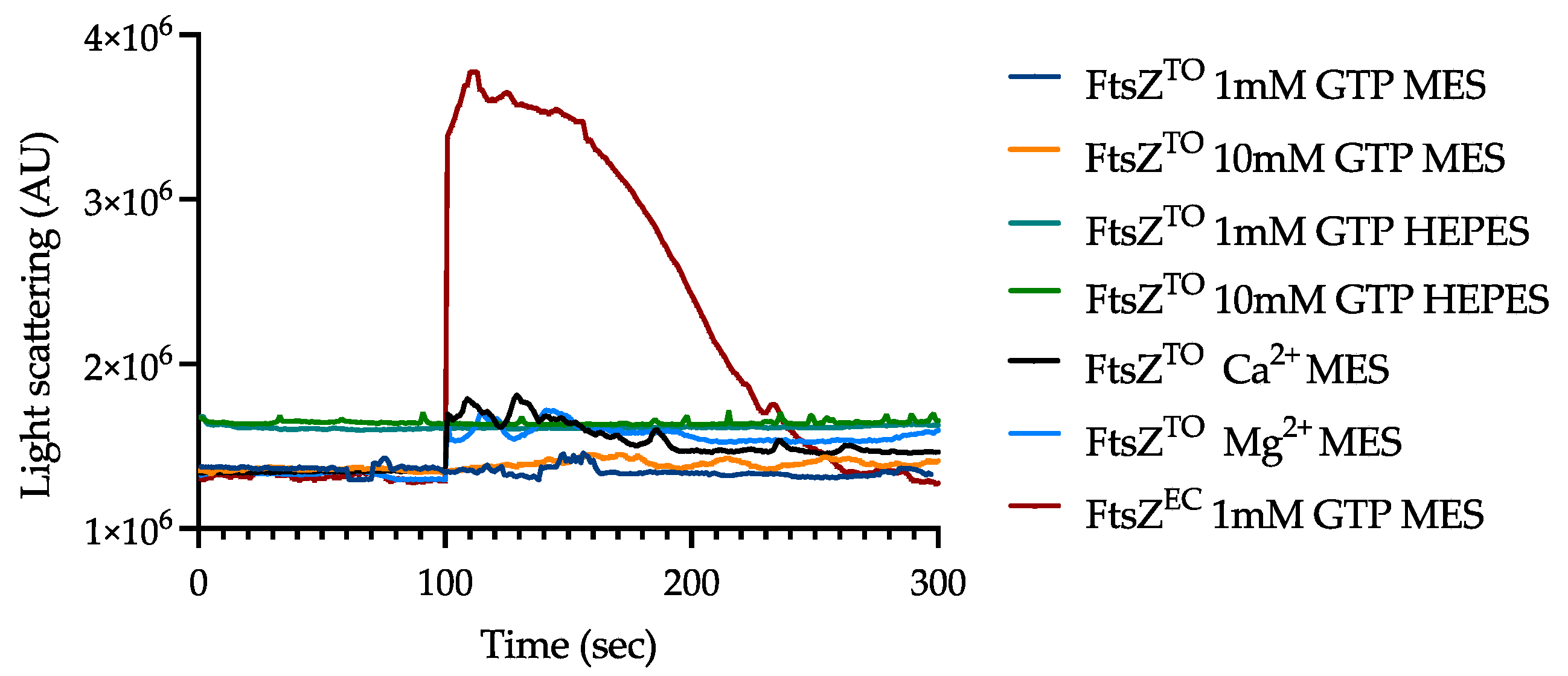
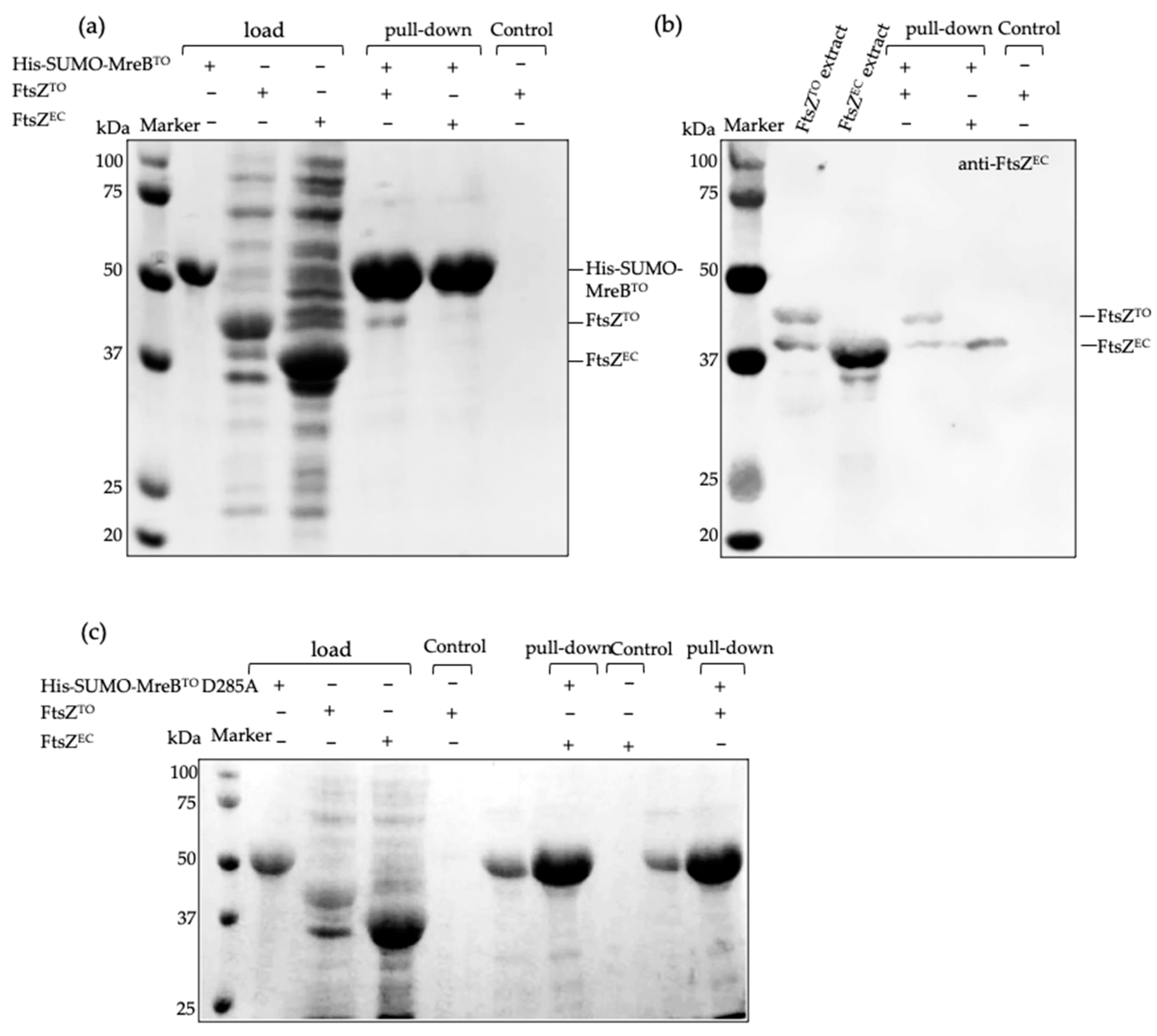
2.5. FtsZTO Fails to Polymerize but Interacts with MreBTO In Vitro
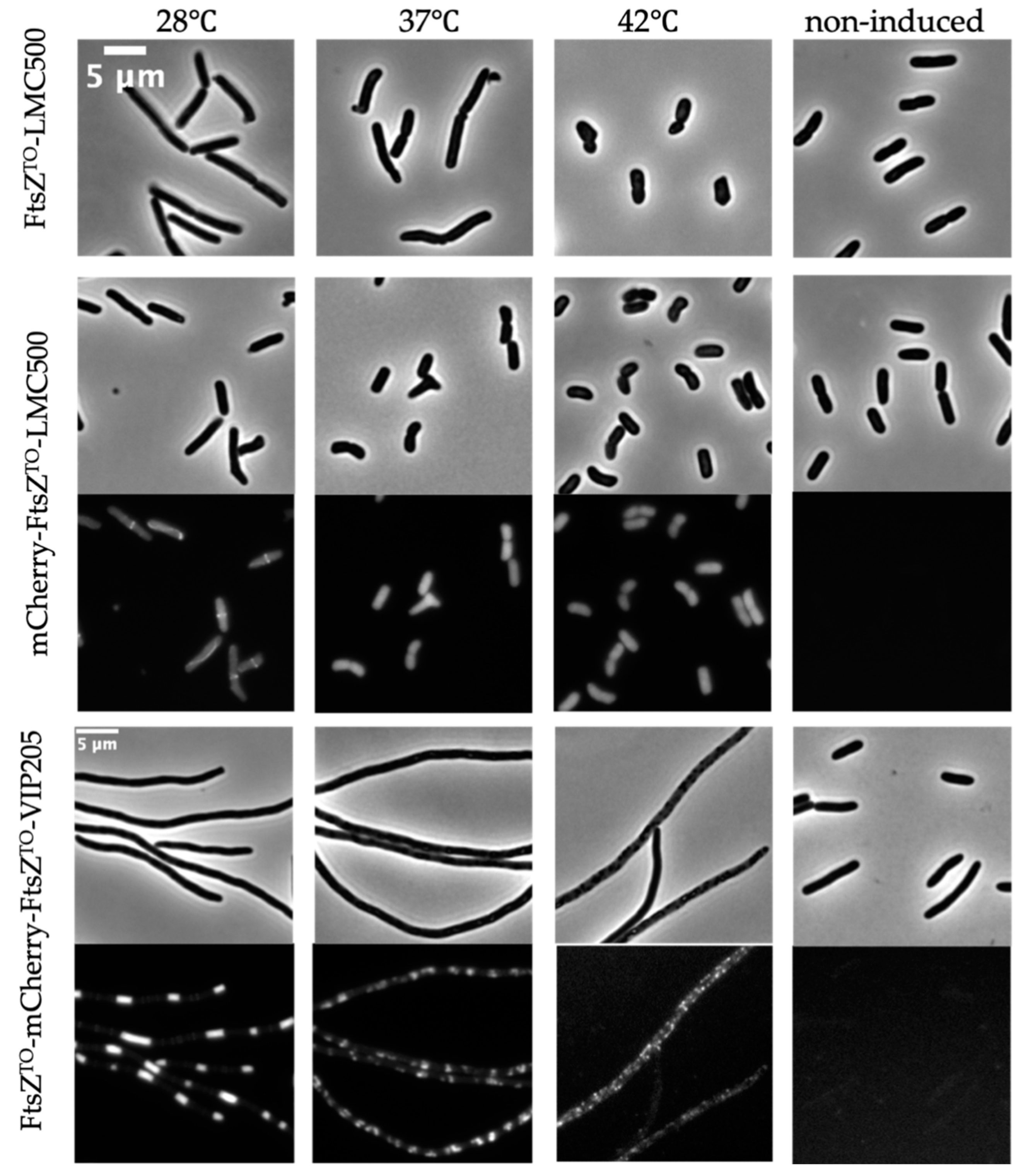
2.6. FtsZTO Shows Temperature-Sensitive Characteristics in E. coli
3. Discussion
3.1. FtsZTO Is Adapted to 28 °C and Temperature Increases Affect Its Localization
3.2. FtsZTO Can Form Rings in E. coli Independent from FtsZEC
3.3. Ca. T. oneisti Lacks Some of the Z-Associate Proteins
3.4. FtsZTO Interacts with MreBTO
3.5. Conclusions
4. Materials and Methods
4.1. Bacterial Strains and Plasmids
4.2. FtsZTO Expression and Purification
4.3. Circular Dichroism
4.4. Ninety-Degree Perpendicular Light Scattering Assay
4.5. GTPase Assay
4.6. Pull-Down
4.7. Microscopy and Image Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adams, D.W.; Errington, J. Bacterial cell division: Assembly, maintenance and disassembly of the Z ring. Nature 2009, 7, 642–653. [Google Scholar] [CrossRef] [PubMed]
- Kuempel, P.L. Temperature-sensitive initiation of chromosome replication in a mutant of Escherichia coli. J. Becteriol. 1969, 100, 1302–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broughton, C.E.; Roper, D.I.; van den Berg, H.A.; Rodger, A. Bacterial cell division: Experimental and theoretical approaches to the divisome. Sci. Prog. 2015, 98, 313–345. [Google Scholar] [CrossRef]
- RayChaudhuri, D.; Park, J.T. Escherichia coli cell-division gene ftsZ encodes a novel GTP-binding protein. Nature 1992, 359, 251–254. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.H.; Durand-Heredia, J.; Janakiraman, A. FtsZ ring stability: Of bundles, tubules, crosslinks, and curves. J. Bacteriol 2013, 195, 1859–1868. [Google Scholar] [CrossRef] [Green Version]
- Gueiros-Filho, F.J.; Losick, R. A widely conserved bacterial cell division protein that promotes assembly of the tubulin-like protein FtsZ. Genes Dev. 2002, 16, 2544–2556. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, A.; Lutkenhaus, J. Dynamic assembly of FtsZ regulated by GTP hydrolysis. EMBO J. 1998, 17, 462–469. [Google Scholar] [CrossRef]
- Scheffers, D.J.; den Blaauwen, T.; Driessen, A.J.M. Non-hydrolysable GTP-γ-S stabilizes the FtsZ polymer in a GDP-bound state. Mol. Microbiol. 2000, 35, 1211–1219. [Google Scholar] [CrossRef] [Green Version]
- Miraldi, E.R.; Thomas, P.J.; Romberg, L. Allosteric models for cooperative polymerization of linear polymers. Biophys. J. 2008, 95, 2470–2486. [Google Scholar] [CrossRef] [Green Version]
- Scheffers, D.; Driessen, A.J. The polymerization mechanism of the bacterial cell division protein FtsZ. FEBS Lett. 2001, 506, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Mendieta, J.; Rico, A.I.; López-Viñas, E.; Vicente, M.; Mingorance, J.; Gómez-Puertas, P. Structural and functional model for Ionic (K+/Na+) and pH dependence of GTPase activity and polymerization of FtsZ, the prokaryotic ortholog of tubulin. J. Mol. Biol. 2009, 390, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Huecas, S.; Llorca, O.; Boskovic, J.; Martín-Benito, J.; Valpuesta, J.M.; Andreu, J.M. Energetics and geometry of FtsZ polymers: Nucleated self-assembly of single protofilaments. Biophys. J. 2008, 94, 1796–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowlett, V.W.; Margolin, W. The Min system and other nucleoid-independent regulators of Z ring positioning. Front. Microbiol. 2015, 6, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichoff, S.; Lutkenhaus, J. Tethering the Z ring to the membrane through a conserved membrane targeting sequence in FtsA. Mol. Microbiol. 2005, 55, 1722–1734. [Google Scholar] [CrossRef]
- Geissler, B.; Elraheb, D.; Margolin, W. A gain-of-function mutation in ftsA bypasses the requirement for the essential cell division gene zipA in Escherichia coli. Proc. Natl. Acad. Sci. USA 2003, 100, 4197–4202. [Google Scholar] [CrossRef] [Green Version]
- Pichoff, S.; Lutkenhaus, J. Unique and overlapping roles for ZipA and FtsA in septal ring assembly in Escherichia coli. EMBO J. 2002, 21, 685–693. [Google Scholar] [CrossRef] [Green Version]
- Pichoff, S.; Du, S.; Lutkenhaus, J. The bypass of ZipA by overexpression of FtsN requires a previously unknown conserved FtsN motif essential for FtsA-FtsN interaction supporting a model in which FtsA monomers recruit late cell division proteins to the Z ring. Mol. Microbiol. 2015, 95, 971–987. [Google Scholar] [CrossRef]
- Marteyn, B.S.; Karimova, G.; Fenton, A.K.; Gazi, A.D.; West, N.; Touqui, L.; Prevost, M.C.; Betton, J.M.; Poyraz, O.; Ladant, D.; et al. ZapE is a novel cell division protein interacting with FtsZ and modulating the Z-ring dynamics. mBio 2014, 5, e00022-14. [Google Scholar] [CrossRef] [Green Version]
- McQuillen, R.; Xiao, J. Insights into the Structure, Function, and Dynamics of the Bacterial Cytokinetic FtsZ-Ring. Annu. Rev. Biophys. 2020, 49, 309–341. [Google Scholar] [CrossRef] [Green Version]
- Stricker, J.; Maddox, P.; Salmon, E.D.; Erickson, H.P. Rapid assembly dynamics of the Escherichia coli FtsZ-ring demonstrated by fluorescence recovery after photobleaching. Proc. Natl. Acad. Sci. USA 2002, 99, 3171–3175. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Lyu, Z.; Miguel, A.; McQuillen, R.; Huang, K.C.; Xiao, J. GTPase activity-coupled treadmilling of the bacterial tubulin FtsZ organizes septal cell wall synthesis. Science 2017, 355, 744–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bisson-Filho, A.W.; Hsu, Y.P.; Squyres, G.R.; Kuru, E.; Wu, F.; Jukes, C.; Sun, Y.; Dekker, C.; Holden, S.; VanNieuwenhze, M.S.; et al. Treadmilling by FtsZ filaments drives peptidoglycan synthesis and bacterial cell division. Science 2017, 355, 739–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez-Diaz, D.A.; García-Soriano, D.A.; Raso, A.; Mücksch, J.; Feingold, M.; Rivas, G.; Schwille, P. Treadmilling analysis reveals new insights into dynamic FtsZ ring architecture. PLoS Biol. 2018, 16, e2004845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, M.J.; Kemper, A.; Gruber-Vodicka, H.; Cardini, U.; van der Geest, M.; Kleiner, M.; Bulgheresi, S.; Mußmann, M.; Herbold, C.; Seah, K.B.B.; et al. Chemosynthetic symbionts of marine invertebrate animials are capable of nitrogen fixation. Nat. Microbiol. 2016, 2, 16195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paredes, F.G.; Viehboeck, T.; Lee, R.; Palatinszky, M.; Mausz, M.A.; Reipert, S.; Schintlmeister, A.; Maier, A.; Volland, J.M.; Hirschfeld, C.; et al. Anaerobic sulfur oxidation underlies adaptation of a chemosynthetic symbiont to oxic-anoxic interfaces. mSystems 2021, 6, e0118620. [Google Scholar] [CrossRef]
- Leisch, N.; Verheul, J.; Heindl, N.R.; Gruber-Vodicka, H.R.; Pende, N.; den Blaauwen, T.; Bulgheresi, S. Growth in width and FtsZ ring longitudinal positioning in a gamma- proteobacterial symbiont. Curr. Biol. 2012, 22, R831–R832. [Google Scholar] [CrossRef] [Green Version]
- Pende, N.; Wang, J.; Weber, P.M.; Verheul, J.; Kuru, E.; Rittmann, S.K.M.R.; Leisch, N.; VanNieuwenhze, M.S.; Brun, Y.V.; den Blaauwen, T.; et al. Host-polarized cell growth in animal symbionts. Curr. Biol. 2018, 28, 1039–1051.e5. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Bratton, B.P.; Gitai, Z.; Huang, K.C. How to build a bacterial cell: MreB as the foreman of E. coli construction. Cell 2018, 172, 1294–1305. [Google Scholar] [CrossRef]
- Wang, J.; Alvarez, L.; Bulgheresi, S.; Cava, F.; den Blaauwen, T. PBP4 is likely involved in cell division of the longitudinally dividing bacterium Candidatus Thiosymbion Oneisti. Antibiotics 2016, 10, 274. [Google Scholar] [CrossRef]
- Dai, K.; Lutkenhaus, J. The proper ratio of FtsZ to FtsA is required for cell division to occur in Escherichia coli. J. Bacteriol. 1992, 174, 6145–6151. [Google Scholar] [CrossRef] [Green Version]
- Haeusser, D.P.; Rowlett, V.W.; Margolin, W. A mutation in Escherichia coli FtsZ bypasses the requirement for the essential division gene zipA and confers resistance to FtsZ assembly inhibitors by stabilizing protofilament bundling. Mol. Microbiol. 2015, 97, 988–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido, T.; Sánchez, M.; Palacios, P.; Aldea, M.; Vicente, M. Transcription of ftsZ oscillates during the cell cycle of Escherichia coli. EMBO J. 1993, 12, 3957–3965. [Google Scholar] [CrossRef] [PubMed]
- van der Ploeg, R.; Verheul, J.; Vischer, N.O.E.; Alexeeva, S.; Hoogendoorn, E.; Postma, M.; Banzhaf, M.; Vollmer, W.; den Blaauwen, T. Colocalization and interaction between elongasome and divisome during a preparative cell division phase in Escherichia coli. Mol. Microbiol. 2013, 87, 1074–1087. [Google Scholar] [CrossRef] [PubMed]
- Fenton, A.K.; Gerdes, K. Direct interaction of FtsZ and MreB is required for septum synthesis and cell division in Escherichia coli. EMBO J. 2013, 32, 1953–1965. [Google Scholar] [CrossRef]
- Lee, C.D.; Sun, H.C.; Hu, S.M.; Chiu, C.F.; Homhuan, A.; Liang, S.M.; Leng, C.H.; Wang, T.F. An improved SUMO fusion protein system for effective production of native proteins. Protein Sci. 2008, 17, 1241–1248. [Google Scholar] [CrossRef] [Green Version]
- Ott, J.A. Sulphide symbioses in shallow sands. Oceanogr. Lit. Rev. 1997, 2, 128. [Google Scholar]
- Scheffers, D.J.; de Wit, J.G.; den Blaauwen, T.; Driessen, A.J.M. GTP hydrolysis of cell division protein FtsZ: Evidence that the active site is formed by the association of monomers. Biochemistry 2002, 41, 521–529. [Google Scholar] [CrossRef] [Green Version]
- Löwe, J.; Amos, L.A. Crystal structure of the bacterial cell-division protein FtsZ. Nature 1998, 391, 203–206. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [Green Version]
- Cordell, S.C.; Robinson, E.J.H.; Löwe, J. Crystal structure of the SOS cell division inhibitor SulA and in complex with FtsZ. Proc. Natl. Acad. Sci. USA 2003, 100, 7889–7894. [Google Scholar] [CrossRef] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Oliva, M.A.; Cordell, S.C.; Löwe, J. Structural insights into FtsZ protofilament formation. Nat. Struct. Mol. Biol. 2004, 11, 1243–1250. [Google Scholar] [CrossRef]
- Schumacher, M.A.; Zeng, W. Structures of the nucleoid occlusion protein SlmA bound to DNA and the C-terminal domain of the cytoskeletal protein FtsZ. Proc. Natl. Acad. Sci. USA 2016, 113, 4988–4993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addinall, S.G.; Small, E.; Whitaker, D.; Sturrock, S.; Donachie, W.D.; Khattar, M.M. New temperature-sensitive alleles of ftsZ in Escherichia coli. J. Bacteriol. 2005, 187, 358–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arjes, H.A.; Lai, B.; Emelue, E.; Steinbach, A.; Levin, P.A. Mutations in the bacterial cell division protein FtsZ highlight the role of GTP binding and longitudinal subunit interactions in assembly and function. BMC Microbiol. 2015, 15, 209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez-Diaz, D.A.; Merino-Salomón, A.; Meyer, F.; Heymann, M.; Rivas, G.; Bramkamp, M.; Schwille, P. FtsZ induces membrane deformations via torsional stress upon GTP hydrolysis. Nat. Commun. 2021, 12, 3310. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Han, X.; Yu, J.; Yao, M.; Tanaka, I. Structural change in FtsZ Induced by intermolecular interactions between bound GTP and the T7 loop. J. Biol. Chem. 2014, 289, 3501–3509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Trimble, M.J.; Brun, Y.V.; Jensen, G.J. The structure of FtsZ filaments in vivo suggests a force-generating role in cell division. EMBO J. 2007, 26, 4694–4708. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; McQuillen, R.; Lyu, Z.; Phillips-Mason, P.; De La Cruz, A.; McCausland, J.W.; Liang, H.; DeMeester, K.E.; Santiago, C.C.; Grimes, C.L.; et al. A two-track model for the spatiotemporal coordination of bacterial septal cell wall synthesis revealed by single-molecule imaging of FtsW. Nat. Microbiol. 2021, 6, 584–593. [Google Scholar] [CrossRef]
- Durand-Heredia, J.M.; Yu, H.H.; De Carlo, S.; Lesser, C.F.; Janakiraman, A. Identification and characterization of ZapC, a stabilizer of the FtsZ ring in Escherichia coli. J. Bacteriol. 2011, 193, 1405–1413. [Google Scholar] [CrossRef] [Green Version]
- Buss, J.A.; Peters, N.T.; Xiao, J.; Bernhardt, T.G. ZapA and ZapB form an FtsZ-independent structure at midcell. Mol. Microbiol. 2017, 104, 652–663. [Google Scholar] [CrossRef] [Green Version]
- Galli, E.; Gerdes, K. FtsZ-ZapA-ZapB interactome of Escherichia coli. J. Bacteriol. 2012, 194, 292–302. [Google Scholar] [CrossRef] [Green Version]
- Söderström, B.; Badrutdinov, A.; Chan, H.; Skoglund, U. Cell shape-independent FtsZ dynamics in synthetically remodeled bacterial cells. Nat. Commun. 2018, 9, 4323. [Google Scholar] [CrossRef] [Green Version]
- Taschner, P.E.; Verest, J.G.; Woldringh, C.L. Genetic and morphological characterization of ftsB and nrdB mutants of Escherichia coli. J. Bacteriol. 1987, 169, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Meiresonne, N.Y.; Consoli, E.; Mertens, L.M.Y.; Chertkova, A.O.; Goedhart, J.; den Blaauwen, T. Superfolder mTurquoise2ox optimized for the bacterial periplasm allows high efficiency in vivo FRET of cell division antibiotic targets. Mol. Microbiol. 2019, 111, 1025–1038. [Google Scholar] [CrossRef] [Green Version]
- Król, E.; Scheffers, D.J. FtsZ polymerization assays: Simple protocols and considerations. J. Vis. Exp. 2013, 81, e50844. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Vischer, N.O.E.; Verheul, J.; Postma, M.; van den Berg van Saparoea, B.; Galli, E.; Natale, P.; Gerdes, K.; Luirink, J.; Vollmer, W.; Vicente, M.; et al. Cell age dependent concentration of Escherichia coli divisome proteins analyzed with ImageJ and ObjectJ. Front. Microbiol. 2015, 6, 1631. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KM (mM) | Vmax (μM GTP/μM FtsZ/min) | kcat (min−1) |
|---|---|---|
| 0.55 ± 0.11 | 2.45 ± 0.24 | 0.49 ± 0.05 |
| E. coli Strain | Relevant Genotype | Reference or Source |
|---|---|---|
| LMC500 | MC4100 lysA | [54] |
| VIP205 | F-, araD139, Δ(ara-leu) 7697, Δ(lac)X74, galE15, galK16, rpsL150, ftsA::kan-Tu-lac9-ptac-ftsZ20 | [32] |
| BL21DE3 plysS | F-, ompT, hsdSB (rB−, mB−), gal dcm (DE3) | Invitrogen |
| plasmids | property | Reference or source |
| pJW11 | pTrc99A down, expressing FtsZTO, p15 origin, catR | This work |
| pJW12 | pTrc99A down, expressing FtsZEC, p15 origin, catR | This work |
| pJW13 | pTrc99A down, expressing mCherry-FtsZTO fusion, pBR322 origin, ampR | This work |
| pJW14 | pTrc99A down, expressing mCherry-FtsZEC fusion, pBR322 origin, ampR | This work |
| pJW15 | pBAD vector, expressing FtsZTO and mCherry-FtsZTO fusion in one operon, ampR | This work |
| pJW16 | pBAD vector, expressing mCherry-FtsZTO fusion, ampR | This work |
| pJW17 | pET11b vector (Novagen), expressing FtsZTO | This work |
| pJW18 | T7 promoter, expressing His tagged SUMO-MreBTO fusion, pBR322 origin, ampR | This work |
| pJW19 | T7 promoter, expressing His tagged-SUMO-MreBTO D285A fusion, pBR322 origin, ampR | This work |
| pJW20 | pET11b vector (Novagen), expressing FtsZTO I207M | This work |
| pJW21 | pET11b vector (Novagen), expressing FtsZTO I273E | This work |
| pNM046 | pTrc99Adown down, expressing mTq2-FtsZEC fusion, pBR322 origin, ampR | [55] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Bulgheresi, S.; den Blaauwen, T. The Longitudinal Dividing Bacterium Candidatus Thiosymbion Oneisti Has a Natural Temperature-Sensitive FtsZ Protein with Low GTPase Activity. Int. J. Mol. Sci. 2022, 23, 3016. https://doi.org/10.3390/ijms23063016
Wang J, Bulgheresi S, den Blaauwen T. The Longitudinal Dividing Bacterium Candidatus Thiosymbion Oneisti Has a Natural Temperature-Sensitive FtsZ Protein with Low GTPase Activity. International Journal of Molecular Sciences. 2022; 23(6):3016. https://doi.org/10.3390/ijms23063016
Chicago/Turabian StyleWang, Jinglan, Silvia Bulgheresi, and Tanneke den Blaauwen. 2022. "The Longitudinal Dividing Bacterium Candidatus Thiosymbion Oneisti Has a Natural Temperature-Sensitive FtsZ Protein with Low GTPase Activity" International Journal of Molecular Sciences 23, no. 6: 3016. https://doi.org/10.3390/ijms23063016
APA StyleWang, J., Bulgheresi, S., & den Blaauwen, T. (2022). The Longitudinal Dividing Bacterium Candidatus Thiosymbion Oneisti Has a Natural Temperature-Sensitive FtsZ Protein with Low GTPase Activity. International Journal of Molecular Sciences, 23(6), 3016. https://doi.org/10.3390/ijms23063016