
Increased Expression of N2BA Titin Corresponds to More Compliant Myofibrils in Athlete’s Heart

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Induction of Athlete’s Heart
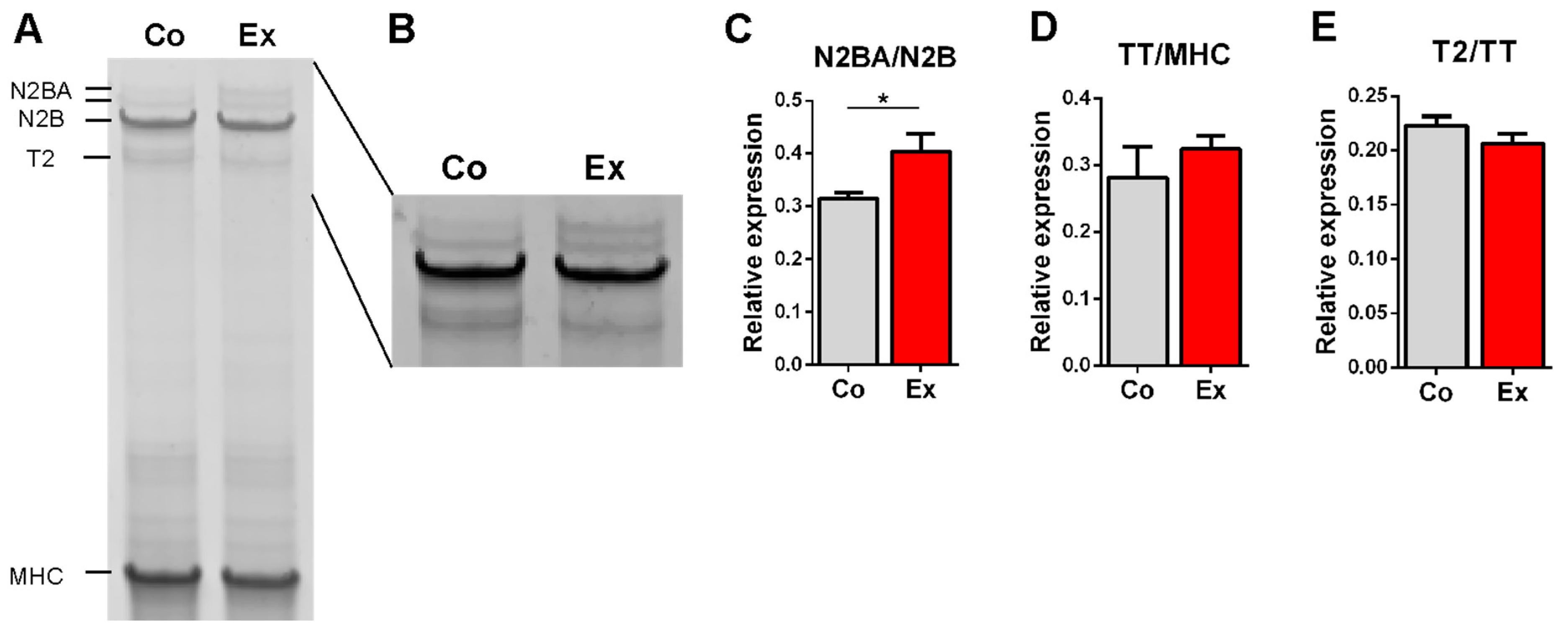
2.2. Effect of Long-Term Exercise on Titin Isoform Content
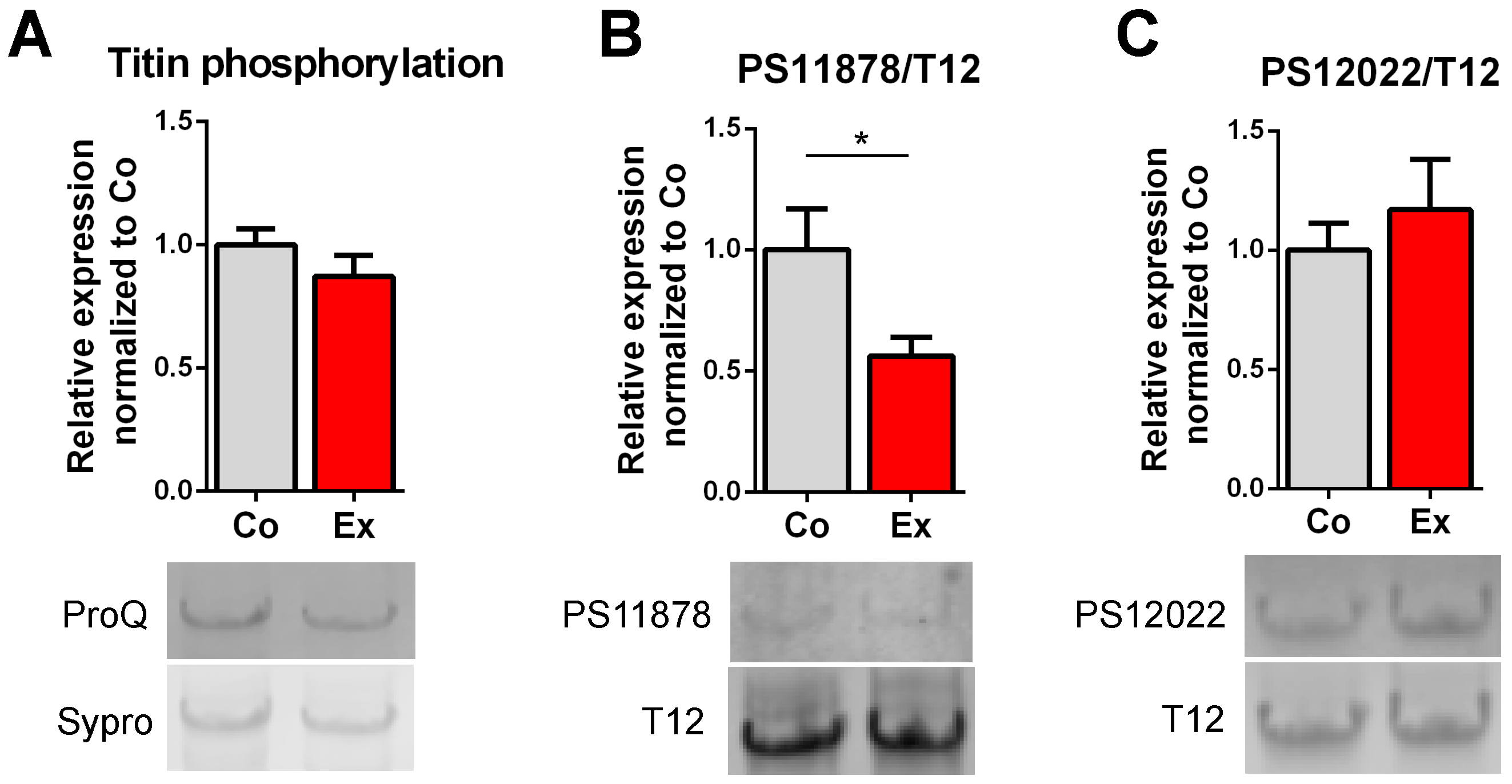
2.3. Exercise-Induced Phosphorylation Modifications on Titin
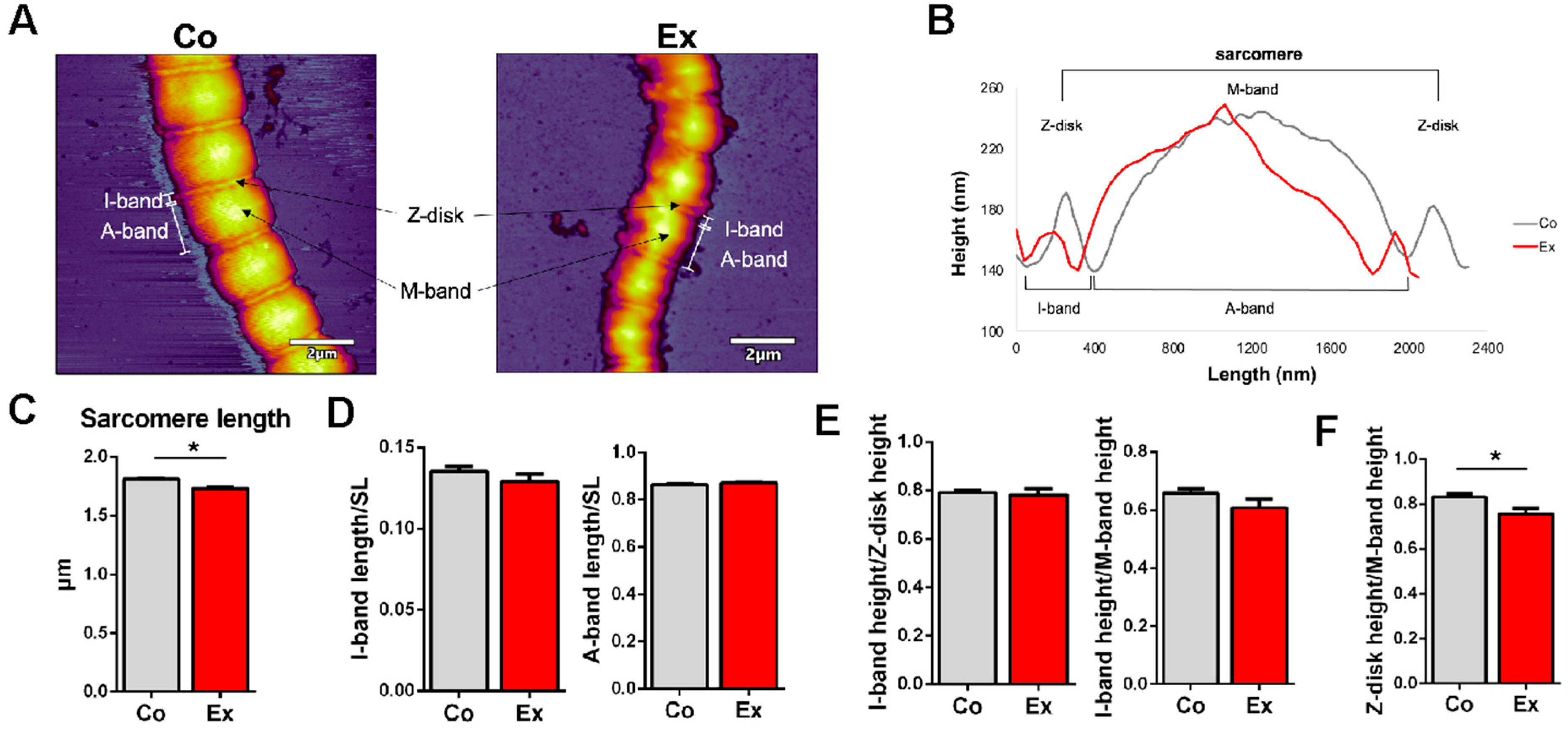
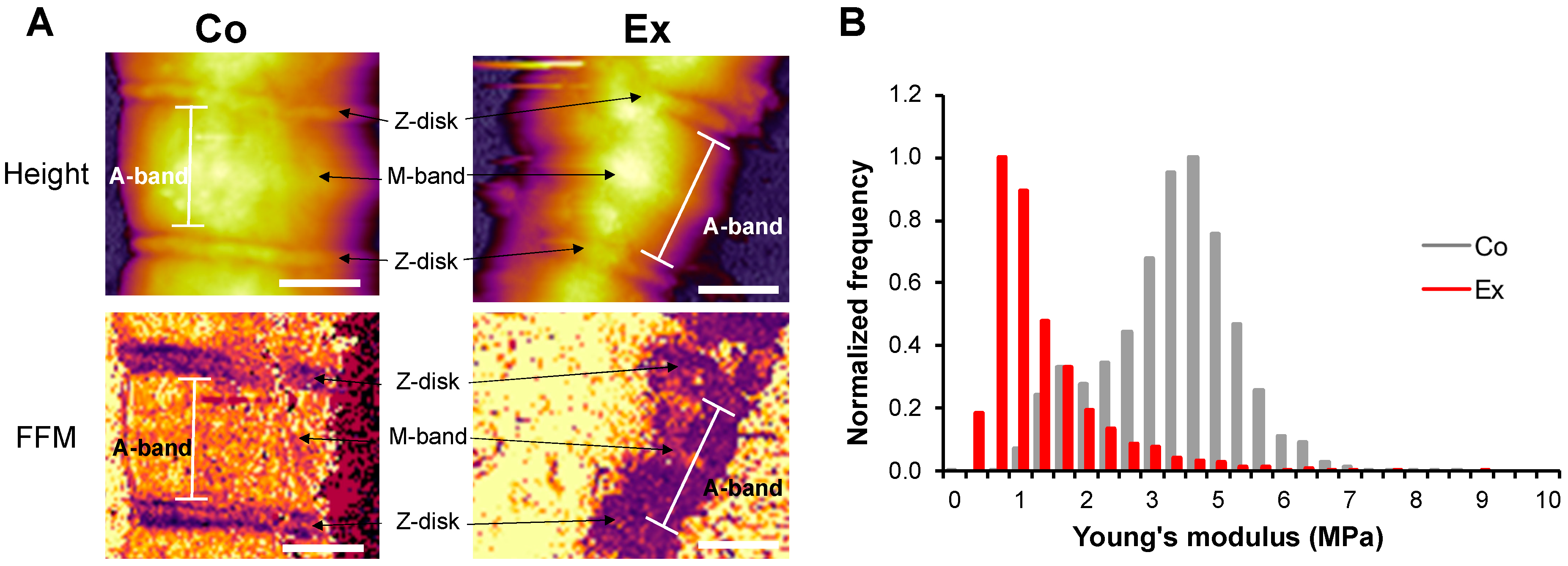
2.4. Myofibril Sarcomere Structure and Elasticity
3. Discussion
3.1. Athlete’s Heart Induced by 12-Week-Long Swimming Training
3.2. Exercise-Induced Titin Isoform Expression Alteration
3.3. Post-Translational Modifications of Titin after Long-Term Exercise
3.4. Structure and Elasticity of Exercise-Exposed Myofibrils
4. Materials and Methods
4.1. Animal Model and Ethical Approval
4.2. Sample Solubilization
4.3. Titin Isoform Analysis and Total Titin Phosphorylation
4.4. Titin’s PEVK Site Specific Phosphorylation
4.5. Preparation of Single Cardiac Myofibrils
4.6. Atomic Force Microscopy (AFM) Imaging and Force Spectroscopy
4.7. Data Analysis and Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zilinski, J.L.; Contursi, M.E.; Isaacs, S.K.; Deluca, J.R.; Lewis, G.D.; Weiner, R.B.; Hutter, A.M., Jr.; d’Hemecourt, P.A.; Troyanos, C.; Dyer, K.S.; et al. Myocardial adaptations to recreational marathon training among middle-aged men. Circ. Cardiovasc. Imaging 2015, 8, e002487. [Google Scholar] [CrossRef]
- Nystoriak, M.A.; Bhatnagar, A. Cardiovascular Effects and Benefits of Exercise. Front. Cardiovasc. Med. 2018, 5, 135. [Google Scholar] [CrossRef]
- Prior, D.L.; La Gerche, A. The athlete’s heart. Heart 2012, 98, 947–955. [Google Scholar] [CrossRef]
- Radovits, T.; Olah, A.; Lux, A.; Nemeth, B.T.; Hidi, L.; Birtalan, E.; Kellermayer, D.; Matyas, C.; Szabo, G.; Merkely, B. Rat model of exercise-induced cardiac hypertrophy: Hemodynamic characterization using left ventricular pressure-volume analysis. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, H124–H134. [Google Scholar] [CrossRef]
- Boldt, K.R.; Rios, J.L.; Joumaa, V.; Herzog, W. Force properties of skinned cardiac muscle following increasing volumes of aerobic exercise in rats. J. Appl. Physiol. 2018, 125, 495–503. [Google Scholar] [CrossRef]
- Diffee, G.M. Adaptation of cardiac myocyte contractile properties to exercise training. Exerc. Sport Sci. Rev. 2004, 32, 112–119. [Google Scholar] [CrossRef]
- Kemi, O.J.; Haram, P.M.; Loennechen, J.P.; Osnes, J.B.; Skomedal, T.; Wisloff, U.; Ellingsen, O. Moderate vs. high exercise intensity: Differential effects on aerobic fitness, cardiomyocyte contractility, and endothelial function. Cardiovasc. Res. 2005, 67, 161–172. [Google Scholar] [CrossRef]
- Apor, A.; Merkely, B.; Morrell, T.; Zhu, S.; Ghosh, E.; Vágó, H.; Andrássy, P.; Kovács, S.J. Diastolic function in Olympic athletes versus controls: Stiffness-based and relaxation-based echocardiographic comparisons. J. Exerc. Sci. Fit. 2013, 11, 29–34. [Google Scholar] [CrossRef]
- Lalande, S.; Mueller, P.J.; Chung, C.S. The link between exercise and titin passive stiffness. Exp. Physiol. 2017, 102, 1055–1066. [Google Scholar] [CrossRef]
- Arbab-Zadeh, A.; Dijk, E.; Prasad, A.; Fu, Q.; Torres, P.; Zhang, R.; Thomas, J.D.; Palmer, D.; Levine, B.D. Effect of aging and physical activity on left ventricular compliance. Circulation 2004, 110, 1799–1805. [Google Scholar] [CrossRef]
- Granzier, H.L.; Irving, T.C. Passive tension in cardiac muscle: Contribution of collagen, titin, microtubules, and intermediate filaments. Biophys. J. 1995, 68, 1027–1044. [Google Scholar] [CrossRef]
- Bang, M.L.; Centner, T.; Fornoff, F.; Geach, A.J.; Gotthardt, M.; McNabb, M.; Witt, C.C.; Labeit, D.; Gregorio, C.C.; Granzier, H.; et al. The complete gene sequence of titin, expression of an unusual approximately 700-kDa titin isoform, and its interaction with obscurin identify a novel Z-line to I-band linking system. Circ. Res. 2001, 89, 1065–1072. [Google Scholar] [CrossRef]
- Labeit, S.; Kolmerer, B. Titins: Giant proteins in charge of muscle ultrastructure and elasticity. Science 1995, 270, 293–296. [Google Scholar] [CrossRef]
- Granzier, H.L.; Labeit, S. The giant protein titin: A major player in myocardial mechanics, signaling, and disease. Circ. Res. 2004, 94, 284–295. [Google Scholar] [CrossRef]
- Kruger, M.; Kotter, S. Titin, a Central Mediator for Hypertrophic Signaling, Exercise-Induced Mechanosignaling and Skeletal Muscle Remodeling. Front. Physiol. 2016, 7, 76. [Google Scholar] [CrossRef]
- Chung, C.S.; Granzier, H.L. Contribution of titin and extracellular matrix to passive pressure and measurement of sarcomere length in the mouse left ventricle. J. Mol. Cell. Cardiol. 2011, 50, 731–739. [Google Scholar] [CrossRef]
- Guo, W.; Schafer, S.; Greaser, M.L.; Radke, M.H.; Liss, M.; Govindarajan, T.; Maatz, H.; Schulz, H.; Li, S.; Parrish, A.M.; et al. RBM20, a gene for hereditary cardiomyopathy, regulates titin splicing. Nat. Med. 2012, 18, 766–773. [Google Scholar] [CrossRef] [PubMed]
- Lahmers, S.; Wu, Y.; Call, D.R.; Labeit, S.; Granzier, H. Developmental control of titin isoform expression and passive stiffness in fetal and neonatal myocardium. Circ. Res. 2004, 94, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Nagueh, S.F.; Shah, G.; Wu, Y.; Torre-Amione, G.; King, N.M.; Lahmers, S.; Witt, C.C.; Becker, K.; Labeit, S.; Granzier, H.L. Altered titin expression, myocardial stiffness, and left ventricular function in patients with dilated cardiomyopathy. Circulation 2004, 110, 155–162. [Google Scholar] [CrossRef]
- Hidalgo, C.; Saripalli, C.; Granzier, H.L. Effect of exercise training on post-translational and post-transcriptional regulation of titin stiffness in striated muscle of wild type and IG KO mice. Arch. Biochem. Biophys. 2014, 552-553, 100–107. [Google Scholar] [CrossRef]
- Muller, A.E.; Kreiner, M.; Kotter, S.; Lassak, P.; Bloch, W.; Suhr, F.; Kruger, M. Acute exercise modifies titin phosphorylation and increases cardiac myofilament stiffness. Front. Physiol. 2014, 5, 449. [Google Scholar] [CrossRef] [PubMed]
- Slater, R.E.; Strom, J.G.; Granzier, H. Effect of exercise on passive myocardial stiffness in mice with diastolic dysfunction. J. Mol. Cell. Cardiol. 2017, 108, 24–33. [Google Scholar] [CrossRef]
- Koser, F.; Loescher, C.; Linke, W.A. Posttranslational modifications of titin from cardiac muscle: How, where, and what for? FEBS J. 2019, 286, 2240–2260. [Google Scholar] [CrossRef]
- Chung, C.S.; Hiske, M.A.; Chadha, A.; Mueller, P.J. Compliant Titin Isoform Content Is Reduced in Left Ventricles of Sedentary Versus Active Rats. Front. Physiol. 2020, 11, 15. [Google Scholar] [CrossRef] [PubMed]
- Methawasin, M.; Hutchinson, K.R.; Lee, E.J.; Smith, J.E., 3rd; Saripalli, C.; Hidalgo, C.G.; Ottenheijm, C.A.; Granzier, H. Experimentally increasing titin compliance in a novel mouse model attenuates the Frank-Starling mechanism but has a beneficial effect on diastole. Circulation 2014, 129, 1924–1936. [Google Scholar] [CrossRef] [PubMed]
- Ogneva, I.V.; Lebedev, D.V.; Shenkman, B.S. Transversal stiffness and Young’s modulus of single fibers from rat soleus muscle probed by atomic force microscopy. Biophys. J. 2010, 98, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Weiwad, W.K.; Linke, W.A.; Wussling, M.H. Sarcomere length-tension relationship of rat cardiac myocytes at lengths greater than optimum. J. Mol. Cell. Cardiol. 2000, 32, 247–259. [Google Scholar] [CrossRef]
- Kubler, J.; Burgstahler, C.; Brendel, J.M.; Gassenmaier, S.; Hagen, F.; Klingel, K.; Olthof, S.C.; Blume, K.; Wolfarth, B.; Mueller, K.A.L.; et al. Cardiac MRI findings to differentiate athlete’s heart from hypertrophic (HCM), arrhythmogenic right ventricular (ARVC) and dilated (DCM) cardiomyopathy. Int. J. Cardiovasc. Imaging 2021, 37, 2501–2515. [Google Scholar] [CrossRef]
- Olah, A.; Kellermayer, D.; Matyas, C.; Nemeth, B.T.; Lux, A.; Szabo, L.; Torok, M.; Ruppert, M.; Meltzer, A.; Sayour, A.A.; et al. Complete Reversion of Cardiac Functional Adaptation Induced by Exercise Training. Med. Sci. Sports Exerc. 2017, 49, 420–429. [Google Scholar] [CrossRef]
- Olah, A.; Barta, B.A.; Sayour, A.A.; Ruppert, M.; Virag-Tulassay, E.; Novak, J.; Varga, Z.V.; Ferdinandy, P.; Merkely, B.; Radovits, T. Balanced Intense Exercise Training Induces Atrial Oxidative Stress Counterbalanced by the Antioxidant System and Atrial Hypertrophy That Is Not Associated with Pathological Remodeling or Arrhythmogenicity. Antioxidants 2021, 10, 452. [Google Scholar] [CrossRef]
- Wang, Y.; Wisloff, U.; Kemi, O.J. Animal models in the study of exercise-induced cardiac hypertrophy. Physiol. Res. 2010, 59, 633–644. [Google Scholar] [CrossRef]
- Kovacs, A.; Olah, A.; Lux, A.; Matyas, C.; Nemeth, B.T.; Kellermayer, D.; Ruppert, M.; Torok, M.; Szabo, L.; Meltzer, A.; et al. Strain and strain rate by speckle-tracking echocardiography correlate with pressure-volume loop-derived contractility indices in a rat model of athlete’s heart. Am. J. Physiol. Heart Circ. Physiol. 2015, 308, H743–H748. [Google Scholar] [CrossRef] [PubMed]
- Olah, A.; Kovacs, A.; Lux, A.; Tokodi, M.; Braun, S.; Lakatos, B.K.; Matyas, C.; Kellermayer, D.; Ruppert, M.; Sayour, A.A.; et al. Characterization of the dynamic changes in left ventricular morphology and function induced by exercise training and detraining. Int. J. Cardiol. 2019, 277, 178–185. [Google Scholar] [CrossRef]
- Olah, A.; Matyas, C.; Kellermayer, D.; Ruppert, M.; Barta, B.A.; Sayour, A.A.; Torok, M.; Koncsos, G.; Giricz, Z.; Ferdinandy, P.; et al. Sex Differences in Morphological and Functional Aspects of Exercise-Induced Cardiac Hypertrophy in a Rat Model. Front. Physiol. 2019, 10, 889. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, C.; Hudson, B.; Bogomolovas, J.; Zhu, Y.; Anderson, B.; Greaser, M.; Labeit, S.; Granzier, H. PKC phosphorylation of titin’s PEVK element: A novel and conserved pathway for modulating myocardial stiffness. Circ. Res. 2009, 105, 631–638. [Google Scholar] [CrossRef]
- Fukuda, N.; Wu, Y.; Nair, P.; Granzier, H.L. Phosphorylation of titin modulates passive stiffness of cardiac muscle in a titin isoform-dependent manner. J. Gen. Physiol. 2005, 125, 257–271. [Google Scholar] [CrossRef] [PubMed]
- Hudson, B.; Hidalgo, C.; Saripalli, C.; Granzier, H. Hyperphosphorylation of mouse cardiac titin contributes to transverse aortic constriction-induced diastolic dysfunction. Circ. Res. 2011, 109, 858–866. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lang, P.; Linke, W.A. Titin stiffness modifies the force-generating region of muscle sarcomeres. Sci. Rep. 2016, 6, 24492. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, N.; Ohnuki, Y.; Kunioka, Y.; Saeki, Y.; Yamada, T. Transverse stiffness of myofibrils of skeletal and cardiac muscles studied by atomic force microscopy. J. Physiol. Sci. 2006, 56, 145–151. [Google Scholar] [CrossRef]
- Warren, C.M.; Krzesinski, P.R.; Greaser, M.L. Vertical agarose gel electrophoresis and electroblotting of high-molecular-weight proteins. Electrophoresis 2003, 24, 1695–1702. [Google Scholar] [CrossRef]
- Berggren, K.; Chernokalskaya, E.; Steinberg, T.H.; Kemper, C.; Lopez, M.F.; Diwu, Z.; Haugland, R.P.; Patton, W.F. Background-free, high sensitivity staining of proteins in one- and two-dimensional sodium dodecyl sulfate-polyacrylamide gels using a luminescent ruthenium complex. Electrophoresis 2000, 21, 2509–2521. [Google Scholar] [CrossRef]
- Freiburg, A.; Trombitas, K.; Hell, W.; Cazorla, O.; Fougerousse, F.; Centner, T.; Kolmerer, B.; Witt, C.; Beckmann, J.S.; Gregorio, C.C.; et al. Series of exon-skipping events in the elastic spring region of titin as the structural basis for myofibrillar elastic diversity. Circ. Res. 2000, 86, 1114–1121. [Google Scholar] [CrossRef] [PubMed]
- Furst, D.O.; Osborn, M.; Nave, R.; Weber, K. The organization of titin filaments in the half-sarcomere revealed by monoclonal antibodies in immunoelectron microscopy: A map of ten nonrepetitive epitopes starting at the Z line extends close to the M line. J. Cell Biol. 1988, 106, 1563–1572. [Google Scholar] [CrossRef]
- Knight, P.J.; Trinick, J.A. Preparation of myofibrils. Methods Enzymol. 1982, 85, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Vikhorev, P.G.; Ferenczi, M.A.; Marston, S.B. Instrumentation to study myofibril mechanics from static to artificial simulations of cardiac cycle. MethodsX 2016, 3, 156–170. [Google Scholar] [CrossRef] [PubMed]
- Kellermayer, M.S.Z.; Voros, Z.; Csik, G.; Herenyi, L. Forced phage uncorking: Viral DNA ejection triggered by a mechanically sensitive switch. Nanoscale 2018, 10, 1898–1904. [Google Scholar] [CrossRef]
- Hutter, J.L.; Bechhoefer, J. Calibration of atomic-force microscope tips. Rev. Sci. Instrum. 1993, 64, 1868–1873. [Google Scholar] [CrossRef]
- Moreno-Herrero, F.; Perez, M.; Baro, A.M.; Avila, J. Characterization by atomic force microscopy of Alzheimer paired helical filaments under physiological conditions. Biophys. J. 2004, 86, 517–525. [Google Scholar] [CrossRef][Green Version]
- Kiss, B.; Kis, Z.; Palyi, B.; Kellermayer, M.S.Z. Topography, Spike Dynamics, and Nanomechanics of Individual Native SARS-CoV-2 Virions. Nano Lett. 2021, 21, 2675–2680. [Google Scholar] [CrossRef] [PubMed]
- Bouchonville, N.; Nicolas, A. Quantification of the Elastic Properties of Soft and Sticky Materials Using AFM. Methods Mol. Biol. 2019, 1886, 281–290. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Co (n = 6) | Ex (n = 6) | p-Value | |
|---|---|---|---|
| BW (g) | 483 ± 24 | 417 ± 18 | 0.06 |
| TL (cm) | 4.33 ± 0.05 | 4.18 ± 0.06 | 0.09 |
| HW (g) | 1.23 ± 0.05 | 1.45 ± 0.08 * | 0.04 |
| HW/BW (g/kg) | 2.55 ± 0.08 | 3.47 ± 0.09 * | <0.01 |
| HW/TL (g/cm) | 0.28 ± 0.01 | 0.34 ± 0.01 * | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kellermayer, D.; Kiss, B.; Tordai, H.; Oláh, A.; Granzier, H.L.; Merkely, B.; Kellermayer, M.; Radovits, T. Increased Expression of N2BA Titin Corresponds to More Compliant Myofibrils in Athlete’s Heart. Int. J. Mol. Sci. 2021, 22, 11110. https://doi.org/10.3390/ijms222011110
Kellermayer D, Kiss B, Tordai H, Oláh A, Granzier HL, Merkely B, Kellermayer M, Radovits T. Increased Expression of N2BA Titin Corresponds to More Compliant Myofibrils in Athlete’s Heart. International Journal of Molecular Sciences. 2021; 22(20):11110. https://doi.org/10.3390/ijms222011110
Chicago/Turabian StyleKellermayer, Dalma, Bálint Kiss, Hedvig Tordai, Attila Oláh, Henk L. Granzier, Béla Merkely, Miklós Kellermayer, and Tamás Radovits. 2021. "Increased Expression of N2BA Titin Corresponds to More Compliant Myofibrils in Athlete’s Heart" International Journal of Molecular Sciences 22, no. 20: 11110. https://doi.org/10.3390/ijms222011110
APA StyleKellermayer, D., Kiss, B., Tordai, H., Oláh, A., Granzier, H. L., Merkely, B., Kellermayer, M., & Radovits, T. (2021). Increased Expression of N2BA Titin Corresponds to More Compliant Myofibrils in Athlete’s Heart. International Journal of Molecular Sciences, 22(20), 11110. https://doi.org/10.3390/ijms222011110