
Microbiota Signals during the Neonatal Period Forge Life-Long Immune Responses

 , , , and
, , , and
Abstract
1. Introduction
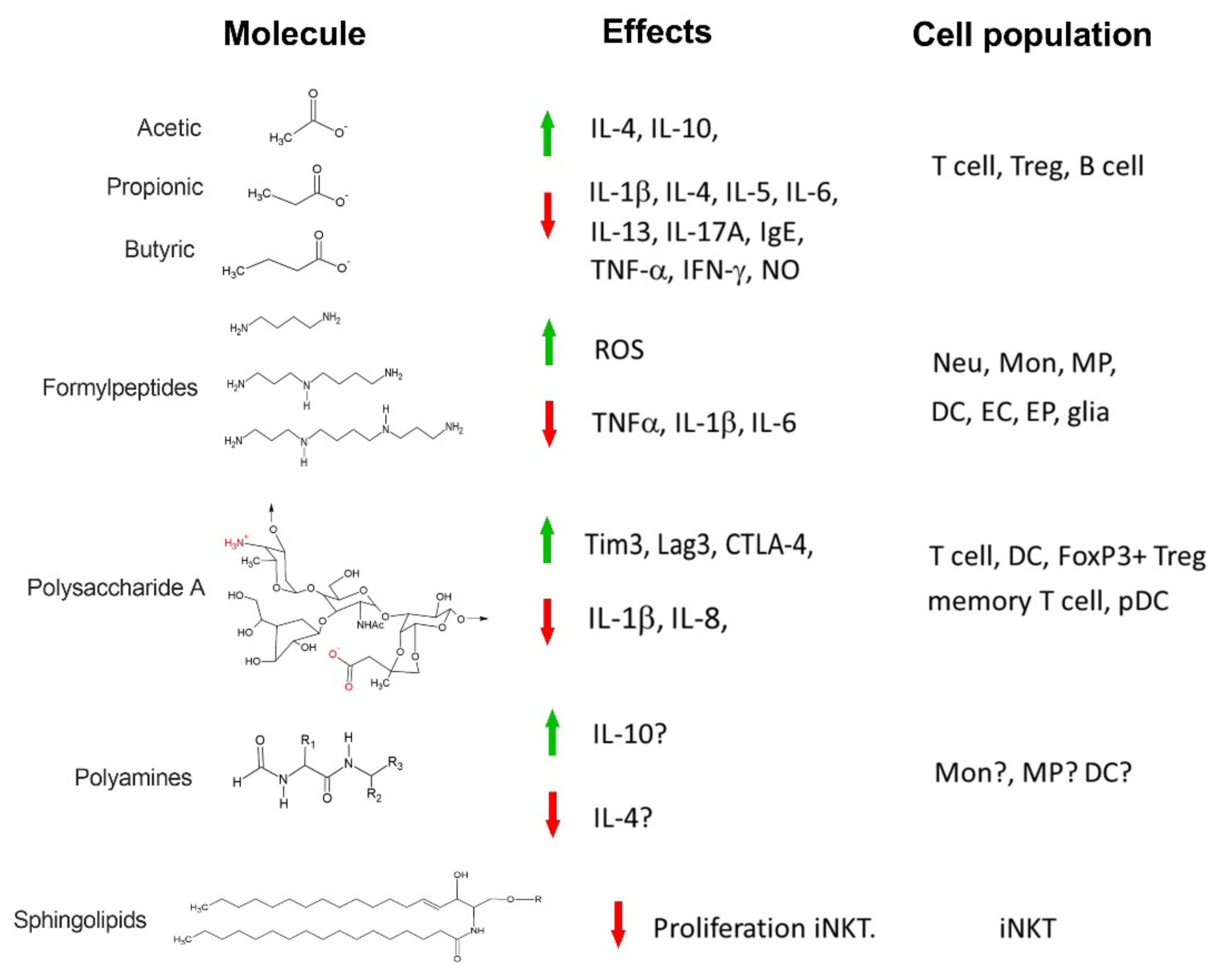
2. Short-Chain Fatty Acids
Immune Intermediaries Affected by SCFAs
3. Important Immune Mediators during the Neonatal Period
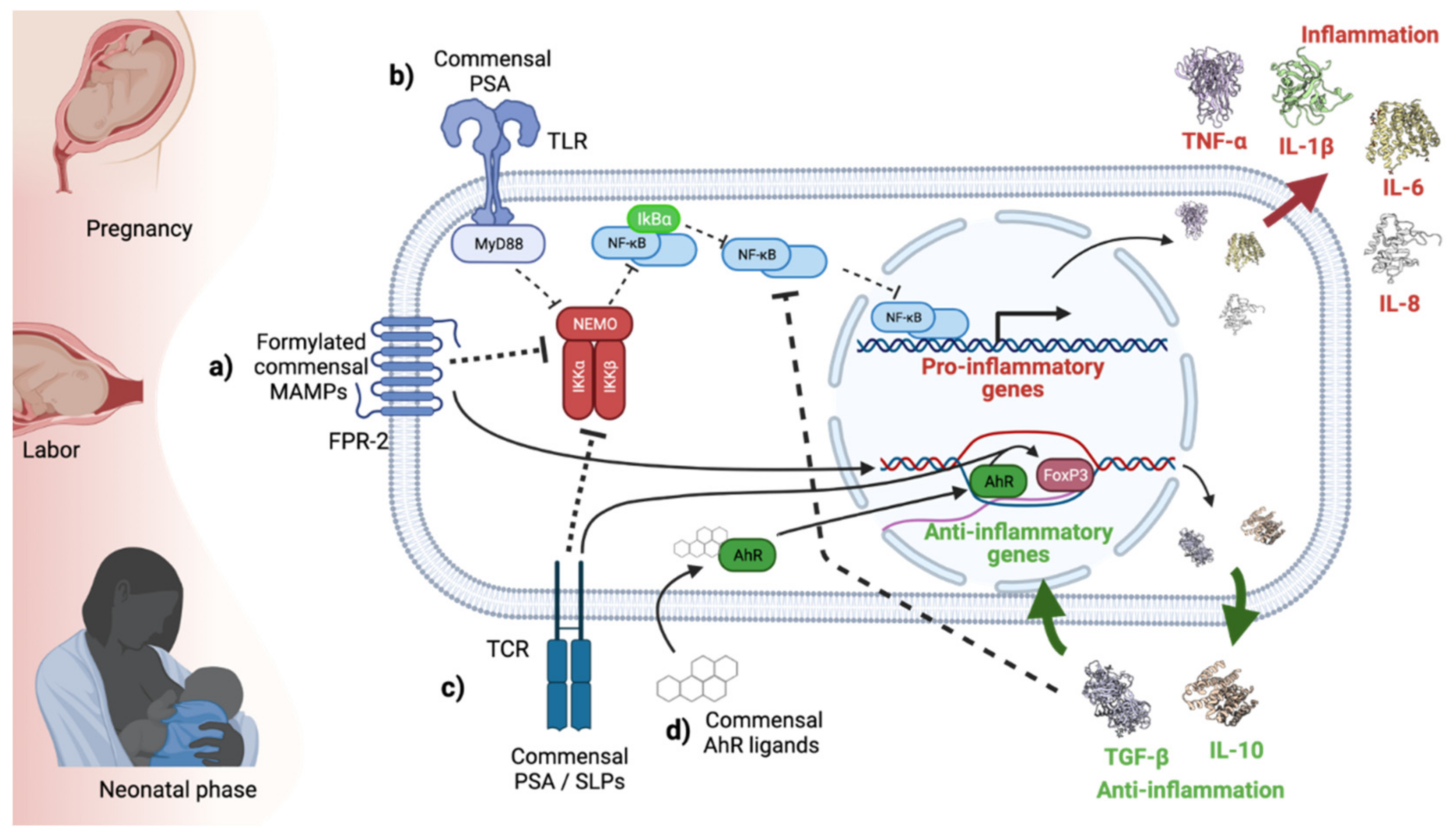
4. Formyl Peptides
5. Polysaccharide A
Immune Molecules Affected by PSA
6. Polyamines
7. Sphingolipids
8. Aryl Hydrocarbon Receptor Ligands
9. Opportunities to Modulate Newborn Microbiota
10. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Gensollen, T.; Iyer, S.S.; Kasper, D.L.; Blumberg, R.S. How colonization by microbiota in early life shapes the immune system. Science 2016, 352, 539–544. [Google Scholar] [CrossRef]
- Renz, H.; Adkins, B.D.; Bartfeld, S.; Blumberg, R.S.; Farber, D.L.; Garssen, J.; Ghazal, P.; Hackam, D.J.; Marsland, B.J.; McCoy, K.D.; et al. The neonatal window of opportunity-early priming for life. J. Allergy Clin. Immunol. 2018, 141, 1212–1214. [Google Scholar] [CrossRef]
- Jimenez, E.; Fernandez, L.; Marin, M.L.; Martin, R.; Odriozola, J.M.; Nueno-Palop, C.; Narbad, A.; Olivares, M.; Xaus, J.; Rodriguez, J.M. Isolation of commensal bacteria from umbilical cord blood of healthy neonates born by cesarean section. Curr. Microbiol. 2005, 51, 270–274. [Google Scholar] [CrossRef]
- Borre, Y.E.; O’Keeffe, G.W.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Microbiota and neurodevelopmental windows: Implications for brain disorders. Trends Mol. Med. 2014, 20, 509–518. [Google Scholar] [CrossRef]
- Torow, N.; Hornef, M.W. The Neonatal Window of Opportunity: Setting the Stage for Life-Long Host-Microbial Interaction and Immune Homeostasis. J. Immunol. 2017, 198, 557–563. [Google Scholar] [CrossRef]
- Moles, L.; Gomez, M.; Heilig, H.; Bustos, G.; Fuentes, S.; de Vos, W.; Fernandez, L.; Rodriguez, J.M.; Jimenez, E. Bacterial diversity in meconium of preterm neonates and evolution of their fecal microbiota during the first month of life. PLoS ONE 2013, 8, e66986. [Google Scholar] [CrossRef]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.; Oehrle, K.; Worthen, G.; Alenghat, T.; Whitsett, J.; Deshmukh, H. Intestinal commensal bacteria mediate lung mucosal immunity and promote resistance of newborn mice to infection. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Ganal-Vonarburg, S.C.; Fuhrer, T.; Gomez de Aguero, M. Maternal microbiota and antibodies as advocates of neonatal health. Gut Microbes 2017, 8, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Jennewein, M.F.; Abu-Raya, B.; Jiang, Y.; Alter, G.; Marchant, A. Transfer of maternal immunity and programming of the newborn immune system. Semin Immunopathol. 2017, 39, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Stinson, L.F.; Payne, M.S.; Keelan, J.A. Planting the seed: Origins, composition, and postnatal health significance of the fetal gastrointestinal microbiota. Crit. Rev. Microbiol. 2017, 43, 352–369. [Google Scholar] [CrossRef]
- Vuillermin, P.J.; Macia, L.; Nanan, R.; Tang, M.L.; Collier, F.; Brix, S. The maternal microbiome during pregnancy and allergic disease in the offspring. Semin Immunopathol. 2017, 39, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.B.; Davis, S.; Sharma, S. Maternal-fetal cross talk through cell-free fetal DNA, telomere shortening, microchimerism, and inflammation. Am. J. Reprod. Immunol. 2018, 79, e12851. [Google Scholar] [CrossRef]
- Walker, R.W.; Clemente, J.C.; Peter, I.; Loos, R.J.F. The prenatal gut microbiome: Are we colonized with bacteria in utero? Pediatr. Obes. 2017, 12 (Suppl. 1), 3–17. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.A.; Hoffmann, C.; Abt, M.C.; Du, Y.; Kobuley, D.; Kirn, T.J.; Bushman, F.D.; Artis, D. Metagenomic analyses reveal antibiotic-induced temporal and spatial changes in intestinal microbiota with associated alterations in immune cell homeostasis. Mucosal. Immunol. 2010, 3, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Tuteja, S.; Ferguson, J.F. Gut Microbiome and Response to Cardiovascular Drugs. Circ. Genom. Precis. Med. 2019, 12, 421–429. [Google Scholar] [CrossRef]
- Wu, H.; Esteve, E.; Tremaroli, V.; Khan, M.T.; Caesar, R.; Mannerås-Holm, L.; Ståhlman, M.; Olsson, L.M.; Serino, M.; Planas-Fèlix, M.; et al. Metformin alters the gut microbiome of individuals with treatment-naive type 2 diabetes, contributing to the therapeutic effects of the drug. Nat. Med. 2017, 23, 850–858. [Google Scholar] [CrossRef]
- O’Sullivan, O.; Cronin, O.; Clarke, S.F.; Murphy, E.F.; Molloy, M.G.; Shanahan, F.; Cotter, P.D. Exercise and the microbiota. Gut Microbes 2015, 6, 131–136. [Google Scholar] [CrossRef]
- Davis, E.C.; Dinsmoor, A.M.; Wang, M.; Donovan, S.M. Microbiome Composition in Pediatric Populations from Birth to Adolescence: Impact of Diet and Prebiotic and Probiotic Interventions. Dig. Dis. Sci. 2020, 65, 706–722. [Google Scholar] [CrossRef]
- Romano-Keeler, J.; Weitkamp, J.H. Maternal influences on fetal microbial colonization and immune development. Pediatr. Res. 2015, 77, 189–195. [Google Scholar] [CrossRef]
- Chu, D.M.; Meyer, K.M.; Prince, A.L.; Aagaard, K.M. Impact of maternal nutrition in pregnancy and lactation on offspring gut microbial composition and function. Gut Microbes 2016, 7, 459–470. [Google Scholar] [CrossRef]
- Forsthuber, T.; Yip, H.C.; Lehmann, P.V. Induction of TH1 and TH2 immunity in neonatal mice. Science 1996, 271, 1728–1730. [Google Scholar] [CrossRef] [PubMed]
- Ridge, J.P.; Fuchs, E.J.; Matzinger, P. Neonatal tolerance revisited: Turning on newborn T cells with dendritic cells. Science 1996, 271, 1723–1726. [Google Scholar] [CrossRef] [PubMed]
- Sarzotti, M.; Robbins, D.S.; Hoffman, P.M. Induction of protective CTL responses in newborn mice by a murine retrovirus. Science 1996, 271, 1726–1728. [Google Scholar] [CrossRef] [PubMed]
- Menard, S.; Forster, V.; Lotz, M.; Gutle, D.; Duerr, C.U.; Gallo, R.L.; Henriques-Normark, B.; Putsep, K.; Andersson, M.; Glocker, E.O.; et al. Developmental switch of intestinal antimicrobial peptide expression. J. Exp. Med. 2008, 205, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Bhat, N.M.; Kantor, A.B.; Bieber, M.M.; Stall, A.M.; Herzenberg, L.A.; Teng, N.N. The ontogeny and functional characteristics of human B-1 (CD5+ B) cells. Int. Immunol. 1992, 4, 243–252. [Google Scholar] [CrossRef]
- Gavin, M.A.; Bevan, M.J. Increased peptide promiscuity provides a rationale for the lack of N regions in the neonatal T cell repertoire. Immunity 1995, 3, 793–800. [Google Scholar] [CrossRef]
- Rios-Covian, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; de Los Reyes-Gavilan, C.G.; Salazar, N. Intestinal Short Chain Fatty Acids and their Link with Diet and Human Health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef]
- Li, H.; Myeroff, L.; Smiraglia, D.; Romero, M.F.; Pretlow, T.P.; Kasturi, L.; Lutterbaugh, J.; Rerko, R.M.; Casey, G.; Issa, J.P.; et al. SLC5A8, a sodium transporter, is a tumor suppressor gene silenced by methylation in human colon aberrant crypt foci and cancers. Proc. Natl. Acad. Sci. USA 2003, 100, 8412–8417. [Google Scholar] [CrossRef] [PubMed]
- Yanase, H.; Takebe, K.; Nio-Kobayashi, J.; Takahashi-Iwanaga, H.; Iwanaga, T. Cellular expression of a sodium-dependent monocarboxylate transporter (Slc5a8) and the MCT family in the mouse kidney. Histochem. Cell Biol. 2008, 130, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Derrien, M.; Vaughan, E.E.; Plugge, C.M.; de Vos, W.M. Akkermansia muciniphila gen. nov., sp. nov., a human intestinal mucin-degrading bacterium. Int. J. Syst. Evol. Microbiol. 2004, 54, 1469–1476. [Google Scholar] [CrossRef]
- Louis, P.; Young, P.; Holtrop, G.; Flint, H.J. Diversity of human colonic butyrate-producing bacteria revealed by analysis of the butyryl-CoA:acetate CoA-transferase gene. Environ. Microbiol. 2010, 12, 304–314. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [PubMed]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, G.T.; Cummings, J.H.; Allison, C. Protein degradation by human intestinal bacteria. J. Gen. Microbiol. 1986, 132, 1647–1656. [Google Scholar] [CrossRef]
- Donohoe, D.R.; Garge, N.; Zhang, X.; Sun, W.; O’Connell, T.M.; Bunger, M.K.; Bultman, S.J. The microbiome and butyrate regulate energy metabolism and autophagy in the mammalian colon. Cell Metab. 2011, 13, 517–526. [Google Scholar] [CrossRef]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef]
- Deehan, E.C.; Yang, C.; Perez-Muñoz, M.E.; Nguyen, N.K.; Cheng, C.C.; Triador, L.; Zhang, Z.; Bakal, J.A.; Walter, J. Precision Microbiome Modulation with Discrete Dietary Fiber Structures Directs Short-Chain Fatty Acid Production. Cell Host Microbe 2020, 27, 389–404.e6. [Google Scholar] [CrossRef]
- Topping, D.L.; Clifton, P.M. Short-chain fatty acids and human colonic function: Roles of resistant starch and nonstarch polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar] [CrossRef]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef]
- Bloemen, J.G.; Venema, K.; van de Poll, M.C.; Olde Damink, S.W.; Buurman, W.A.; Dejong, C.H. Short chain fatty acids exchange across the gut and liver in humans measured at surgery. Clin. Nutr. 2009, 28, 657–661. [Google Scholar] [CrossRef] [PubMed]
- Boets, E.; Gomand, S.V.; Deroover, L.; Preston, T.; Vermeulen, K.; De Preter, V.; Hamer, H.M.; Van den Mooter, G.; De Vuyst, L.; Courtin, C.M.; et al. Systemic availability and metabolism of colonic-derived short-chain fatty acids in healthy subjects: A stable isotope study. J. Physiol. 2017, 595, 541–555. [Google Scholar] [CrossRef] [PubMed]
- Frost, G.; Sleeth, M.L.; Sahuri-Arisoylu, M.; Lizarbe, B.; Cerdan, S.; Brody, L.; Anastasovska, J.; Ghourab, S.; Hankir, M.; Zhang, S.; et al. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat. Commun. 2014, 5, 3611. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.; Goldsworthy, S.M.; Barnes, A.A.; Eilert, M.M.; Tcheang, L.; Daniels, D.; Muir, A.I.; Wigglesworth, M.J.; Kinghorn, I.; Fraser, N.J.; et al. The Orphan G protein-coupled receptors GPR41 and GPR43 are activated by propionate and other short chain carboxylic acids. J. Biol. Chem. 2003, 278, 11312–11319. [Google Scholar] [CrossRef]
- Le Poul, E.; Loison, C.; Struyf, S.; Springael, J.Y.; Lannoy, V.; Decobecq, M.E.; Brezillon, S.; Dupriez, V.; Vassart, G.; Van Damme, J.; et al. Functional characterization of human receptors for short chain fatty acids and their role in polymorphonuclear cell activation. J. Biol. Chem. 2003, 278, 25481–25489. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I.; Inoue, D.; Maeda, T.; Hara, T.; Ichimura, A.; Miyauchi, S.; Kobayashi, M.; Hirasawa, A.; Tsujimoto, G. Short-chain fatty acids and ketones directly regulate sympathetic nervous system via G protein-coupled receptor 41 (GPR41). Proc. Natl. Acad. Sci. USA 2011, 108, 8030–8035. [Google Scholar] [CrossRef]
- Tolhurst, G.; Heffron, H.; Lam, Y.S.; Parker, H.E.; Habib, A.M.; Diakogiannaki, E.; Cameron, J.; Grosse, J.; Reimann, F.; Gribble, F.M. Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via the G-protein-coupled receptor FFAR2. Diabetes 2012, 61, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Pluznick, J.L.; Protzko, R.J.; Gevorgyan, H.; Peterlin, Z.; Sipos, A.; Han, J.; Brunet, I.; Wan, L.X.; Rey, F.; Wang, T.; et al. Olfactory receptor responding to gut microbiota-derived signals plays a role in renin secretion and blood pressure regulation. Proc. Natl. Acad. Sci. USA 2013, 110, 4410–4415. [Google Scholar] [CrossRef]
- Tazoe, H.; Otomo, Y.; Karaki, S.; Kato, I.; Fukami, Y.; Terasaki, M.; Kuwahara, A. Expression of short-chain fatty acid receptor GPR41 in the human colon. Biomed. Res. 2009, 30, 149–156. [Google Scholar] [CrossRef]
- Nøhr, M.K.; Egerod, K.L.; Christiansen, S.H.; Gille, A.; Offermanns, S.; Schwartz, T.W.; Møller, M. Expression of the short chain fatty acid receptor GPR41/FFAR3 in autonomic and somatic sensory ganglia. Neuroscience 2015, 290, 126–137. [Google Scholar] [CrossRef]
- Nilsson, N.E.; Kotarsky, K.; Owman, C.; Olde, B. Identification of a free fatty acid receptor, FFA2R, expressed on leukocytes and activated by short-chain fatty acids. Biochem. Biophys. Res. Commun. 2003, 303, 1047–1052. [Google Scholar] [CrossRef]
- Brunkhorst, B.A.; Kraus, E.; Coppi, M.; Budnick, M.; Niederman, R. Propionate induces polymorphonuclear leukocyte activation and inhibits formylmethionyl-leucyl-phenylalanine-stimulated activation. Infect. Immun. 1992, 60, 2957–2968. [Google Scholar] [CrossRef]
- Senga, T.; Iwamoto, S.; Yoshida, T.; Yokota, T.; Adachi, K.; Azuma, E.; Hamaguchi, M.; Iwamoto, T. LSSIG is a novel murine leukocyte-specific GPCR that is induced by the activation of STAT3. Blood 2003, 101, 1185–1187. [Google Scholar] [CrossRef] [PubMed]
- Karaki, S.; Tazoe, H.; Hayashi, H.; Kashiwabara, H.; Tooyama, K.; Suzuki, Y.; Kuwahara, A. Expression of the short-chain fatty acid receptor, GPR43, in the human colon. J. Mol. Histol. 2008, 39, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Karaki, S.; Mitsui, R.; Hayashi, H.; Kato, I.; Sugiya, H.; Iwanaga, T.; Furness, J.B.; Kuwahara, A. Short-chain fatty acid receptor, GPR43, is expressed by enteroendocrine cells and mucosal mast cells in rat intestine. Cell Tissue Res. 2006, 324, 353–360. [Google Scholar] [CrossRef]
- D’Souza, W.N.; Douangpanya, J.; Mu, S.; Jaeckel, P.; Zhang, M.; Maxwell, J.R.; Rottman, J.B.; Labitzke, K.; Willee, A.; Beckmann, H.; et al. Differing roles for short chain fatty acids and GPR43 agonism in the regulation of intestinal barrier function and immune responses. PLoS ONE 2017, 12, e0180190. [Google Scholar] [CrossRef] [PubMed]
- Taggart, A.K.; Kero, J.; Gan, X.; Cai, T.Q.; Cheng, K.; Ippolito, M.; Ren, N.; Kaplan, R.; Wu, K.; Wu, T.J.; et al. (D)-beta-Hydroxybutyrate inhibits adipocyte lipolysis via the nicotinic acid receptor PUMA-G. J. Biol. Chem. 2005, 280, 26649–26652. [Google Scholar] [CrossRef]
- Yousefi, S.; Cooper, P.R.; Mueck, B.; Potter, S.L.; Jarai, G. cDNA representational difference analysis of human neutrophils stimulated by GM-CSF. Biochem. Biophys. Res. Commun. 2000, 277, 401–409. [Google Scholar] [CrossRef]
- Schaub, A.; Fütterer, A.; Pfeffer, K. PUMA-G, an IFN-gamma-inducible gene in macrophages is a novel member of the seven transmembrane spanning receptor superfamily. Eur. J. Immunol. 2001, 31, 3714–3725. [Google Scholar] [CrossRef]
- Tunaru, S.; Kero, J.; Schaub, A.; Wufka, C.; Blaukat, A.; Pfeffer, K.; Offermanns, S. PUMA-G and HM74 are receptors for nicotinic acid and mediate its anti-lipolytic effect. Nat. Med. 2003, 9, 352–355. [Google Scholar] [CrossRef]
- Thangaraju, M.; Cresci, G.A.; Liu, K.; Ananth, S.; Gnanaprakasam, J.P.; Browning, D.D.; Mellinger, J.D.; Smith, S.B.; Digby, G.J.; Lambert, N.A.; et al. GPR109A is a G-protein-coupled receptor for the bacterial fermentation product butyrate and functions as a tumor suppressor in colon. Cancer Res. 2009, 69, 2826–2832. [Google Scholar] [CrossRef] [PubMed]
- Li, H.M.; Zhang, M.; Xu, S.T.; Li, D.Z.; Zhu, L.Y.; Peng, S.W.; Chen, G.Q.; Martin, P.M.; Ganapathy, V.; Wei, C.J. Nicotinic acid inhibits glucose-stimulated insulin secretion via the G protein-coupled receptor PUMA-G in murine islet β cells. Pancreas 2011, 40, 615–621. [Google Scholar] [CrossRef]
- Li, G.; Shi, Y.; Huang, H.; Zhang, Y.; Wu, K.; Luo, J.; Sun, Y.; Lu, J.; Benovic, J.L.; Zhou, N. Internalization of the human nicotinic acid receptor GPR109A is regulated by G(i), GRK2, and arrestin3. J. Biol. Chem. 2010, 285, 22605–22618. [Google Scholar] [CrossRef] [PubMed]
- Conzelmann, S.; Levai, O.; Bode, B.; Eisel, U.; Raming, K.; Breer, H.; Strotmann, J. A novel brain receptor is expressed in a distinct population of olfactory sensory neurons. Eur. J. Neurosci. 2000, 12, 3926–3934. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.; Pehl, U.; Breer, H.; Strotmann, J. Olfactory receptor expressed in ganglia of the autonomic nervous system. J. Neurosci. Res. 2002, 68, 176–184. [Google Scholar] [CrossRef]
- Lévai, O.; Feistel, T.; Breer, H.; Strotmann, J. Cells in the vomeronasal organ express odorant receptors but project to the accessory olfactory bulb. J. Comp. Neurol. 2006, 498, 476–490. [Google Scholar] [CrossRef]
- Fleischer, J.; Bumbalo, R.; Bautze, V.; Strotmann, J.; Breer, H. Expression of odorant receptor Olfr78 in enteroendocrine cells of the colon. Cell Tissue Res. 2015, 361, 697–710. [Google Scholar] [CrossRef]
- Chang, A.J.; Ortega, F.E.; Riegler, J.; Madison, D.V.; Krasnow, M.A. Oxygen regulation of breathing through an olfactory receptor activated by lactate. Nature 2015, 527, 240–244. [Google Scholar] [CrossRef]
- Jovancevic, N.; Wunderlich, K.A.; Haering, C.; Flegel, C.; Maßberg, D.; Weinrich, M.; Weber, L.; Tebbe, L.; Kampik, A.; Gisselmann, G.; et al. Deep Sequencing of the Human Retinae Reveals the Expression of Odorant Receptors. Front. Cell Neurosci. 2017, 11, 3. [Google Scholar] [CrossRef]
- Aisenberg, W.H.; Huang, J.; Zhu, W.; Rajkumar, P.; Cruz, R.; Santhanam, L.; Natarajan, N.; Yong, H.M.; De Santiago, B.; Oh, J.J.; et al. Defining an olfactory receptor function in airway smooth muscle cells. Sci. Rep. 2016, 6, 38231. [Google Scholar] [CrossRef]
- Torres-Torrelo, H.; Ortega-Sáenz, P.; Macías, D.; Omura, M.; Zhou, T.; Matsunami, H.; Johnson, R.S.; Mombaerts, P.; López-Barneo, J. The role of Olfr78 in the breathing circuit of mice. Nature 2018, 561, E33–E40. [Google Scholar] [CrossRef]
- Mosienko, V.; Chang, A.J.; Alenina, N.; Teschemacher, A.G.; Kasparov, S. Rodents and humans are able to detect the odour of L-Lactate. PLoS ONE 2017, 12, e0178478. [Google Scholar] [CrossRef] [PubMed]
- Imai, T.; Suzuki, M.; Sakano, H. Odorant receptor-derived cAMP signals direct axonal targeting. Science 2006, 314, 657–661. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Si, H.; Liu, D.; Jiang, H. Butyrate activates the cAMP-protein kinase A-cAMP response element-binding protein signaling pathway in Caco-2 cells. J. Nutr. 2012, 142, 1–6. [Google Scholar] [CrossRef]
- Wise, A.; Foord, S.M.; Fraser, N.J.; Barnes, A.A.; Elshourbagy, N.; Eilert, M.; Ignar, D.M.; Murdock, P.R.; Steplewski, K.; Green, A.; et al. Molecular identification of high and low affinity receptors for nicotinic acid. J. Biol. Chem. 2003, 278, 9869–9874. [Google Scholar] [CrossRef]
- Ahmed, K.; Tunaru, S.; Offermanns, S. GPR109A, GPR109B and GPR81, a family of hydroxy-carboxylic acid receptors. Trends. Pharm. Sci. 2009, 30, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Digby, J.E.; McNeill, E.; Dyar, O.J.; Lam, V.; Greaves, D.R.; Choudhury, R.P. Anti-inflammatory effects of nicotinic acid in adipocytes demonstrated by suppression of fractalkine, RANTES, and MCP-1 and upregulation of adiponectin. Atherosclerosis 2010, 209, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Wanders, D.; Graff, E.C.; Judd, R.L. Effects of high fat diet on GPR109A and GPR81 gene expression. Biochem. Biophys. Res. Commun. 2012, 425, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Deng, X.; Wu, C.; Zhou, Q.; Chen, L.; Shi, Y.; Huang, H.; Zhou, N. Distinct kinetic and spatial patterns of protein kinase C (PKC)- and epidermal growth factor receptor (EGFR)-dependent activation of extracellular signal-regulated kinases 1 and 2 by human nicotinic acid receptor GPR109A. J. Biol. Chem. 2011, 286, 31199–31212. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Lai, X.; Ye, L.; Chen, K.; Cao, Z.; Gong, W.; Jin, L.; Wang, C.; Liu, M.; Liao, Y.; et al. Activated niacin receptor HCA2 inhibits chemoattractant-mediated macrophage migration via Gβγ/PKC/ERK1/2 pathway and heterologous receptor desensitization. Sci. Rep. 2017, 7, 42279. [Google Scholar] [CrossRef] [PubMed]
- Ang, Z.; Xiong, D.; Wu, M.; Ding, J.L. FFAR2-FFAR3 receptor heteromerization modulates short-chain fatty acid sensing. Faseb. J. 2018, 32, 289–303. [Google Scholar] [CrossRef]
- Gomes, S.D.; Oliveira, C.S.; Azevedo-Silva, J.; Casanova, M.R.; Barreto, J.; Pereira, H.; Chaves, S.R.; Rodrigues, L.R.; Casal, M.; Côrte-Real, M.; et al. The Role of Diet Related Short-Chain Fatty Acids in Colorectal Cancer Metabolism and Survival: Prevention and Therapeutic Implications. Curr. Med. Chem. 2020, 27, 4087–4108. [Google Scholar] [CrossRef]
- Stumpff, F. A look at the smelly side of physiology: Transport of short chain fatty acids. Pflug. Arch. 2018, 470, 571–598. [Google Scholar] [CrossRef]
- Zhan, K.; Jiang, M.; Gong, X.; Zhao, G. Effect of short-chain fatty acids on the expression of genes involved in short-chain fatty acid transporters and inflammatory response in goat jejunum epithelial cells. In Vitr. Cell Dev. Biol. Anim. 2018, 54, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Waldecker, M.; Kautenburger, T.; Daumann, H.; Busch, C.; Schrenk, D. Inhibition of histone-deacetylase activity by short-chain fatty acids and some polyphenol metabolites formed in the colon. J. Nutr. Biochem. 2008, 19, 587–593. [Google Scholar] [CrossRef]
- Fellows, R.; Denizot, J.; Stellato, C.; Cuomo, A.; Jain, P.; Stoyanova, E.; Balazsi, S.; Hajnady, Z.; Liebert, A.; Kazakevych, J.; et al. Microbiota derived short chain fatty acids promote histone crotonylation in the colon through histone deacetylases. Nat. Commun. 2018, 9, 105. [Google Scholar] [CrossRef]
- Baradari, V.; Huether, A.; Hopfner, M.; Schuppan, D.; Scherubl, H. Antiproliferative and proapoptotic effects of histone deacetylase inhibitors on gastrointestinal neuroendocrine tumor cells. Endocr. Relat. Cancer 2006, 13, 1237–1250. [Google Scholar] [CrossRef]
- De Vadder, F.; Kovatcheva-Datchary, P.; Goncalves, D.; Vinera, J.; Zitoun, C.; Duchampt, A.; Backhed, F.; Mithieux, G. Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell 2014, 156, 84–96. [Google Scholar] [CrossRef]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef]
- Erny, D.; Hrabe de Angelis, A.L.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; Keren-Shaul, H.; Mahlakoiv, T.; Jakobshagen, K.; Buch, T.; et al. Host microbiota constantly control maturation and function of microglia in the CNS. Nat. Neurosci. 2015, 18, 965–977. [Google Scholar] [CrossRef]
- Tahara, Y.; Yamazaki, M.; Sukigara, H.; Motohashi, H.; Sasaki, H.; Miyakawa, H.; Haraguchi, A.; Ikeda, Y.; Fukuda, S.; Shibata, S. Gut microbiota-derived short chain fatty acids induce circadian clock entrainment in mouse peripheral tissue. Sci. Rep. 2018, 8, 1395. [Google Scholar] [CrossRef]
- Roduit, C.; Frei, R.; Ferstl, R.; Loeliger, S.; Westermann, P.; Rhyner, C.; Schiavi, E.; Barcik, W.; Rodriguez-Perez, N.; Wawrzyniak, M.; et al. High levels of butyrate and propionate in early life are associated with protection against atopy. Allergy 2019, 74, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.F.; Wang, J.; Yan, X.L.; Tian, F.; Zhao, J.B.; Wang, Y.J.; Jiang, T. Histone deacetylase inhibitor, butyrate, attenuates lipopolysaccharide-induced acute lung injury in mice. Respir. Res. 2010, 11, 33. [Google Scholar] [CrossRef] [PubMed]
- Cahenzli, J.; Koller, Y.; Wyss, M.; Geuking, M.B.; McCoy, K.D. Intestinal microbial diversity during early-life colonization shapes long-term IgE levels. Cell Host Microbe 2013, 14, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Schwarzer, M.; Srutkova, D.; Hermanova, P.; Leulier, F.; Kozakova, H.; Schabussova, I. Diet Matters: Endotoxin in the Diet Impacts the Level of Allergic Sensitization in Germ-Free Mice. PLoS ONE 2017, 12, e0167786. [Google Scholar] [CrossRef] [PubMed]
- Pacifico, L.G.; Marinho, F.A.; Fonseca, C.T.; Barsante, M.M.; Pinho, V.; Sales-Junior, P.A.; Cardoso, L.S.; Araujo, M.I.; Carvalho, E.M.; Cassali, G.D.; et al. Schistosoma mansoni antigens modulate experimental allergic asthma in a murine model: A major role for CD4+ CD25+ Foxp3+ T cells independent of interleukin-10. Infect. Immun. 2009, 77, 98–107. [Google Scholar] [CrossRef]
- Cardoso, L.S.; Oliveira, S.C.; Goes, A.M.; Oliveira, R.R.; Pacifico, L.G.; Marinho, F.V.; Fonseca, C.T.; Cardoso, F.C.; Carvalho, E.M.; Araujo, M.I. Schistosoma mansoni antigens modulate the allergic response in a murine model of ovalbumin-induced airway inflammation. Clin. Exp. Immunol. 2010, 160, 266–274. [Google Scholar] [CrossRef]
- Resende, S.D.; Magalhaes, F.C.; Rodrigues-Oliveira, J.L.; Castro, V.N.; Souza, C.S.A.; Oliveira, E.J.; Carneiro, M.; Geiger, S.M.; Negrao-Correa, D.A. Modulation of Allergic Reactivity in Humans Is Dependent on Schistosoma mansoni Parasite Burden, Low Levels of IL-33 or TNF-alpha and High Levels of IL-10 in Serum. Front. Immunol. 2018, 9, 3158. [Google Scholar] [CrossRef] [PubMed]
- Huuskonen, J.; Suuronen, T.; Nuutinen, T.; Kyrylenko, S.; Salminen, A. Regulation of microglial inflammatory response by sodium butyrate and short-chain fatty acids. Br. J. Pharmacol. 2004, 141, 874–880. [Google Scholar] [CrossRef]
- MacFabe, D.F.; Cain, D.P.; Rodriguez-Capote, K.; Franklin, A.E.; Hoffman, J.E.; Boon, F.; Taylor, A.R.; Kavaliers, M.; Ossenkopp, K.P. Neurobiological effects of intraventricular propionic acid in rats: Possible role of short chain fatty acids on the pathogenesis and characteristics of autism spectrum disorders. Behav. Brain Res. 2007, 176, 149–169. [Google Scholar] [CrossRef]
- MacFabe, D.F.; Cain, N.E.; Boon, F.; Ossenkopp, K.P.; Cain, D.P. Effects of the enteric bacterial metabolic product propionic acid on object-directed behavior, social behavior, cognition, and neuroinflammation in adolescent rats: Relevance to autism spectrum disorder. Behav. Brain Res. 2011, 217, 47–54. [Google Scholar] [CrossRef]
- Yang, L.L.; Millischer, V.; Rodin, S.; MacFabe, D.F.; Villaescusa, J.C.; Lavebratt, C. Enteric short-chain fatty acids promote proliferation of human neural progenitor cells. J. Neurochem. 2020, 154, 635–646. [Google Scholar] [CrossRef]
- Wenzel, T.J.; Gates, E.J.; Ranger, A.L.; Klegeris, A. Short-chain fatty acids (SCFAs) alone or in combination regulate select immune functions of microglia-like cells. Mol. Cell Neurosci. 2020, 105, 103493. [Google Scholar] [CrossRef]
- Abdel-Haq, R.; Schlachetzki, J.C.M.; Glass, C.K.; Mazmanian, S.K. Microbiome-microglia connections via the gut-brain axis. J. Exp. Med. 2019, 216, 41–59. [Google Scholar] [CrossRef]
- Han, S.; Lu, J.; Zhang, Y.; Cheng, C.; Li, L.; Han, L.; Huang, B. HDAC inhibitors TSA and sodium butyrate enhanced the human IL-5 expression by altering histone acetylation status at its promoter region. Immunol. Lett. 2007, 108, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kim, M.; Kang, S.G.; Jannasch, A.H.; Cooper, B.; Patterson, J.; Kim, C.H. Short-chain fatty acids induce both effector and regulatory T cells by suppression of histone deacetylases and regulation of the mTOR-S6K pathway. Mucosal. Immunol. 2015, 8, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Shu, D.; Zheng, M.; Wang, J.; Luo, C.; Wang, Y.; Guo, F.; Zou, X.; Lv, X.; Li, Y.; et al. Microbial metabolite butyrate facilitates M2 macrophage polarization and function. Sci. Rep. 2016, 6, 24838. [Google Scholar] [CrossRef] [PubMed]
- Usami, M.; Kishimoto, K.; Ohata, A.; Miyoshi, M.; Aoyama, M.; Fueda, Y.; Kotani, J. Butyrate and trichostatin A attenuate nuclear factor kappaB activation and tumor necrosis factor alpha secretion and increase prostaglandin E2 secretion in human peripheral blood mononuclear cells. Nutr. Res. 2008, 28, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Kendrick, S.F.; O’Boyle, G.; Mann, J.; Zeybel, M.; Palmer, J.; Jones, D.E.; Day, C.P. Acetate, the key modulator of inflammatory responses in acute alcoholic hepatitis. Hepatology 2010, 51, 1988–1997. [Google Scholar] [CrossRef]
- Vinolo, M.A.; Rodrigues, H.G.; Hatanaka, E.; Sato, F.T.; Sampaio, S.C.; Curi, R. Suppressive effect of short-chain fatty acids on production of proinflammatory mediators by neutrophils. J. Nutr. Biochem. 2011, 22, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.V.; Hao, L.; Offermanns, S.; Medzhitov, R. The microbial metabolite butyrate regulates intestinal macrophage function via histone deacetylase inhibition. Proc. Natl. Acad. Sci. USA 2014, 111, 2247–2252. [Google Scholar] [CrossRef] [PubMed]
- Saemann, M.D.; Bohmig, G.A.; Osterreicher, C.H.; Burtscher, H.; Parolini, O.; Diakos, C.; Stockl, J.; Horl, W.H.; Zlabinger, G.J. Anti-inflammatory effects of sodium butyrate on human monocytes: Potent inhibition of IL-12 and up-regulation of IL-10 production. FASEB J. 2000, 14, 2380–2382. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly, Y.M.; Glickman, J.N.; Garrett, W.S. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Gurav, A.; Sivaprakasam, S.; Brady, E.; Padia, R.; Shi, H.; Thangaraju, M.; Prasad, P.D.; Manicassamy, S.; Munn, D.H.; et al. Activation of Gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis. Immunity 2014, 40, 128–139. [Google Scholar] [CrossRef]
- Shi, Y.; Xu, L.Z.; Peng, K.; Wu, W.; Wu, R.; Liu, Z.Q.; Yang, G.; Geng, X.R.; Liu, J.; Liu, Z.G.; et al. Specific immunotherapy in combination with Clostridium butyricum inhibits allergic inflammation in the mouse intestine. Sci. Rep. 2015, 5, 17651. [Google Scholar] [CrossRef]
- Liao, H.Y.; Tao, L.; Zhao, J.; Qin, J.; Zeng, G.C.; Cai, S.W.; Li, Y.; Zhang, J.; Chen, H.G. Clostridium butyricum in combination with specific immunotherapy converts antigen-specific B cells to regulatory B cells in asthmatic patients. Sci. Rep. 2016, 6, 20481. [Google Scholar] [CrossRef]
- Xu, L.Z.; Yang, L.T.; Qiu, S.Q.; Yang, G.; Luo, X.Q.; Miao, B.P.; Geng, X.R.; Liu, Z.Q.; Liu, J.; Wen, Z.; et al. Combination of specific allergen and probiotics induces specific regulatory B cells and enhances specific immunotherapy effect on allergic rhinitis. Oncotarget 2016, 7, 54360–54369. [Google Scholar] [CrossRef]
- Fernando, M.R.; Saxena, A.; Reyes, J.L.; McKay, D.M. Butyrate enhances antibacterial effects while suppressing other features of alternative activation in IL-4-induced macrophages. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G822–G831. [Google Scholar] [CrossRef]
- Needell, J.C.; Ir, D.; Robertson, C.E.; Kroehl, M.E.; Frank, D.N.; Zipris, D. Maternal treatment with short-chain fatty acids modulates the intestinal microbiota and immunity and ameliorates type 1 diabetes in the offspring. PLoS ONE 2017, 12, e0183786. [Google Scholar] [CrossRef]
- Jimenez, E.; Sacedon, R.; Vicente, A.; Hernandez-Lopez, C.; Zapata, A.G.; Varas, A. Rat peripheral CD4+CD8+ T lymphocytes are partially immunocompetent thymus-derived cells that undergo post-thymic maturation to become functionally mature CD4+ T lymphocytes. J. Immunol. 2002, 168, 5005–5013. [Google Scholar] [CrossRef]
- Marchant, A.; Appay, V.; van der Sande, M.; Dulphy, N.; Liesnard, C.; Kidd, M.; Kaye, S.; Ojuola, O.; Gillespie, G.M.A.; Vargas Cuero, A.L.; et al. Mature CD8+ T lymphocyte response to viral infection during fetal life. J. Clin. Investig. 2003, 111, 1747–1755. [Google Scholar] [CrossRef] [PubMed]
- Cupedo, T.; Nagasawa, M.; Weijer, K.; Blom, B.; Spits, H. Development and activation of regulatory T cells in the human fetus. Eur. J. Immunol. 2005, 35, 383–390. [Google Scholar] [CrossRef]
- Ota, M.O.; Vekemans, J.; Schlegel-Haueter, S.E.; Fielding, K.; Whittle, H.; Lambert, P.H.; McAdam, K.P.; Siegrist, C.A.; Marchant, A. Hepatitis B immunisation induces higher antibody and memory Th2 responses in new-borns than in adults. Vaccine 2004, 22, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Mold, J.E.; Michaelsson, J.; Burt, T.D.; Muench, M.O.; Beckerman, K.P.; Busch, M.P.; Lee, T.H.; Nixon, D.F.; McCune, J.M. Maternal alloantigens promote the development of tolerogenic fetal regulatory T cells in utero. Science 2008, 322, 1562–1565. [Google Scholar] [CrossRef] [PubMed]
- Torow, N.; Yu, K.; Hassani, K.; Freitag, J.; Schulz, O.; Basic, M.; Brennecke, A.; Sparwasser, T.; Wagner, N.; Bleich, A.; et al. Active suppression of intestinal CD4(+)TCRalphabeta(+) T-lymphocyte maturation during the postnatal period. Nat. Commun. 2015, 6, 7725. [Google Scholar] [CrossRef]
- Rogier, E.W.; Frantz, A.L.; Bruno, M.E.; Wedlund, L.; Cohen, D.A.; Stromberg, A.J.; Kaetzel, C.S. Secretory antibodies in breast milk promote long-term intestinal homeostasis by regulating the gut microbiota and host gene expression. Proc. Natl. Acad. Sci. USA 2014, 111, 3074–3079. [Google Scholar] [CrossRef]
- Koch, M.A.; Reiner, G.L.; Lugo, K.A.; Kreuk, L.S.; Stanbery, A.G.; Ansaldo, E.; Seher, T.D.; Ludington, W.B.; Barton, G.M. Maternal IgG and IgA Antibodies Dampen Mucosal T Helper Cell Responses in Early Life. Cell 2016, 165, 827–841. [Google Scholar] [CrossRef]
- Wilcox, C.R.; Holder, B.; Jones, C.E. Factors Affecting the FcRn-Mediated Transplacental Transfer of Antibodies and Implications for Vaccination in Pregnancy. Front. Immunol. 2017, 8, 1294. [Google Scholar] [CrossRef]
- Giles, M.L.; Krishnaswamy, S.; Wallace, E.M. Maternal immunisation: What have been the gains? Where are the gaps? What does the future hold? F1000Res 2018, 7. [Google Scholar] [CrossRef]
- Mold, J.E.; Venkatasubrahmanyam, S.; Burt, T.D.; Michaelsson, J.; Rivera, J.M.; Galkina, S.A.; Weinberg, K.; Stoddart, C.A.; McCune, J.M. Fetal and adult hematopoietic stem cells give rise to distinct T cell lineages in humans. Science 2010, 330, 1695–1699. [Google Scholar] [CrossRef]
- Yang, S.; Fujikado, N.; Kolodin, D.; Benoist, C.; Mathis, D. Immune tolerance. Regulatory T cells generated early in life play a distinct role in maintaining self-tolerance. Science 2015, 348, 589–594. [Google Scholar] [CrossRef]
- Wang, J.; Wissink, E.M.; Watson, N.B.; Smith, N.L.; Grimson, A.; Rudd, B.D. Fetal and adult progenitors give rise to unique populations of CD8+ T cells. Blood 2016, 128, 3073–3082. [Google Scholar] [CrossRef] [PubMed]
- Michaelsson, J.; Mold, J.E.; McCune, J.M.; Nixon, D.F. Regulation of T cell responses in the developing human fetus. J. Immunol. 2006, 176, 5741–5748. [Google Scholar] [CrossRef] [PubMed]
- Lathrop, S.K.; Bloom, S.M.; Rao, S.M.; Nutsch, K.; Lio, C.W.; Santacruz, N.; Peterson, D.A.; Stappenbeck, T.S.; Hsieh, C.S. Peripheral education of the immune system by colonic commensal microbiota. Nature 2011, 478, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Cebula, A.; Seweryn, M.; Rempala, G.A.; Pabla, S.S.; McIndoe, R.A.; Denning, T.L.; Bry, L.; Kraj, P.; Kisielow, P.; Ignatowicz, L. Thymus-derived regulatory T cells contribute to tolerance to commensal microbiota. Nature 2013, 497, 258–262. [Google Scholar] [CrossRef]
- Kim, K.S.; Hong, S.W.; Han, D.; Yi, J.; Jung, J.; Yang, B.G.; Lee, J.Y.; Lee, M.; Surh, C.D. Dietary antigens limit mucosal immunity by inducing regulatory T cells in the small intestine. Science 2016, 351, 858–863. [Google Scholar] [CrossRef]
- Witkin, S.S.; Linhares, I.M.; Bongiovanni, A.M.; Herway, C.; Skupski, D. Unique alterations in infection-induced immune activation during pregnancy. BJOG 2011, 118, 145–153. [Google Scholar] [CrossRef]
- Shi, Z.; Vasquez-Vivar, J.; Luo, K.; Yan, Y.; Northington, F.; Mehrmohammadi, M.; Tan, S. Ascending Lipopolysaccharide-Induced Intrauterine Inflammation in Near-Term Rabbits Leading to Newborn Neurobehavioral Deficits. Dev. Neurosci. 2018, 40, 534–546. [Google Scholar] [CrossRef]
- Roy, S.K.; Meng, Q.; Sadowitz, B.D.; Kollisch-Singule, M.; Yepuri, N.; Satalin, J.; Gatto, L.A.; Nieman, G.F.; Cooney, R.N.; Clark, D. Enteral administration of bacteria fermented formula in newborn piglets: A high fidelity model for necrotizing enterocolitis (NEC). PLoS ONE 2018, 13, e0201172. [Google Scholar] [CrossRef]
- Raymond, S.L.; Hawkins, R.B.; Murphy, T.J.; Rincon, J.C.; Stortz, J.A.; Lopez, M.C.; Ungaro, R.; Ellett, F.; Baker, H.V.; Wynn, J.L.; et al. Impact of toll-like receptor 4 stimulation on human neonatal neutrophil spontaneous migration, transcriptomics, and cytokine production. J. Mol. Med. 2018, 96, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Glaser, K.; Schlegel, N.; Claus, H.; Speer, C.P. An underestimated pathogen: Staphylococcus epidermidis induces pro-inflammatory responses in human alveolar epithelial cells. Cytokine 2019, 123, 154761. [Google Scholar] [CrossRef] [PubMed]
- Rich, B.S.; Dolgin, S.E. Necrotizing Enterocolitis. Pediatr. Rev. 2017, 38, 552–559. [Google Scholar] [CrossRef]
- Spehlmann, M.E.; Eckmann, L. Nuclear factor-kappa B in intestinal protection and destruction. Curr. Opin. Gastroenterol. 2009, 25, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Nafday, S.M.; Chen, W.; Peng, L.; Babyatsky, M.W.; Holzman, I.R.; Lin, J. Short-chain fatty acids induce colonic mucosal injury in rats with various postnatal ages. Pediatr. Res. 2005, 57, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Thymann, T.; Moller, H.K.; Stoll, B.; Stoy, A.C.; Buddington, R.K.; Bering, S.B.; Jensen, B.B.; Olutoye, O.O.; Siggers, R.H.; Molbak, L.; et al. Carbohydrate maldigestion induces necrotizing enterocolitis in preterm pigs. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, G1115–G1125. [Google Scholar] [CrossRef] [PubMed]
- Prado, C.; Michels, M.; Avila, P.; Burger, H.; Milioli, M.V.M.; Dal-Pizzol, F. The protective effects of fecal microbiota transplantation in an experimental model of necrotizing enterocolitis. J. Pediatr. Surg. 2019, 54, 1578–1583. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Hui, Y.; Goericke-Pesch, S.; Pankratova, S.; Kot, W.; Pan, X.; Thymann, T.; Sangild, P.T.; Nguyen, D.N. Gut and immune effects of bioactive milk factors in preterm pigs exposed to prenatal inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 317, G67–G77. [Google Scholar] [CrossRef]
- Hoang, T.K.; He, B.; Wang, T.; Tran, D.Q.; Rhoads, J.M.; Liu, Y. Protective effect of Lactobacillus reuteri DSM 17938 against experimental necrotizing enterocolitis is mediated by Toll-like receptor 2. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G231–G240. [Google Scholar] [CrossRef]
- Laskowska, E.; Jarosz, L.; Gradzki, Z. Effect of Multi-Microbial Probiotic Formulation Bokashi on Pro- and Anti-Inflammatory Cytokines Profile in the Serum, Colostrum and Milk of Sows, and in a Culture of Polymorphonuclear Cells Isolated from Colostrum. Probiotics Antimicrob. Proteins 2019, 11, 220–232. [Google Scholar] [CrossRef]
- Culley, F.J.; Pollott, J.; Openshaw, P.J. Age at first viral infection determines the pattern of T cell-mediated disease during reinfection in adulthood. J. Exp. Med. 2002, 196, 1381–1386. [Google Scholar] [CrossRef] [PubMed]
- Dakhama, A.; Park, J.W.; Taube, C.; Joetham, A.; Balhorn, A.; Miyahara, N.; Takeda, K.; Gelfand, E.W. The enhancement or prevention of airway hyperresponsiveness during reinfection with respiratory syncytial virus is critically dependent on the age at first infection and IL-13 production. J. Immunol. 2005, 175, 1876–1883. [Google Scholar] [CrossRef] [PubMed]
- Dakhama, A.; Lee, Y.M.; Ohnishi, H.; Jing, X.; Balhorn, A.; Takeda, K.; Gelfand, E.W. Virus-specific IgE enhances airway responsiveness on reinfection with respiratory syncytial virus in newborn mice. J. Allergy Clin. Immunol. 2009, 123, 138–145.e5. [Google Scholar] [CrossRef] [PubMed]
- Saglani, S.; Gregory, L.G.; Manghera, A.K.; Branchett, W.J.; Uwadiae, F.; Entwistle, L.J.; Oliver, R.A.; Vasiliou, J.E.; Sherburn, R.; Lui, S.; et al. Inception of early-life allergen-induced airway hyperresponsiveness is reliant on IL-13(+)CD4(+) T cells. Sci. Immunol. 2018, 3. [Google Scholar] [CrossRef]
- Tasker, L.; Lindsay, R.W.; Clarke, B.T.; Cochrane, D.W.; Hou, S. Infection of mice with respiratory syncytial virus during neonatal life primes for enhanced antibody and T cell responses on secondary challenge. Clin. Exp. Immunol. 2008, 153, 277–288. [Google Scholar] [CrossRef]
- Lee, Y.M.; Miyahara, N.; Takeda, K.; Prpich, J.; Oh, A.; Balhorn, A.; Joetham, A.; Gelfand, E.W.; Dakhama, A. IFN-gamma production during initial infection determines the outcome of reinfection with respiratory syncytial virus. Am. J. Respir. Crit. Care Med. 2008, 177, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Tregoning, J.S.; Yamaguchi, Y.; Harker, J.; Wang, B.; Openshaw, P.J. The role of T cells in the enhancement of respiratory syncytial virus infection severity during adult reinfection of neonatally sensitized mice. J. Virol. 2008, 82, 4115–4124. [Google Scholar] [CrossRef]
- Ruckwardt, T.J.; Malloy, A.M.; Gostick, E.; Price, D.A.; Dash, P.; McClaren, J.L.; Thomas, P.G.; Graham, B.S. Neonatal CD8 T-cell hierarchy is distinct from adults and is influenced by intrinsic T cell properties in respiratory syncytial virus infected mice. PLoS Pathog. 2011, 7, e1002377. [Google Scholar] [CrossRef]
- Lynch, J.P.; Werder, R.B.; Loh, Z.; Sikder, M.A.A.; Curren, B.; Zhang, V.; Rogers, M.J.; Lane, K.; Simpson, J.; Mazzone, S.B.; et al. Plasmacytoid dendritic cells protect from viral bronchiolitis and asthma through semaphorin 4a-mediated T reg expansion. J. Exp. Med. 2018, 215, 537–557. [Google Scholar] [CrossRef]
- Glocker, E.O.; Kotlarz, D.; Klein, C.; Shah, N.; Grimbacher, B. IL-10 and IL-10 receptor defects in humans. Ann. N. Y. Acad. Sci. 2011, 1246, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Campos-Alberto, E.; Morita, Y.; Yamaguchi, M.; Toshimitsu, T.; Kimura, K.; Ikegami, S.; Katsuki, T.; Kohno, Y.; Shimojo, N. Low Interleukin 10 Production at Birth Is a Risk Factor for Atopic Dermatitis in Neonates with Bifidobacterium Colonization. Int. Arch. Allergy Immunol. 2018, 177, 342–349. [Google Scholar] [CrossRef]
- Genovese, F.; Mancuso, G.; Cuzzola, M.; Biondo, C.; Beninati, C.; Delfino, D.; Teti, G. Role of IL-10 in a neonatal mouse listeriosis model. J. Immunol. 1999, 163, 2777–2782. [Google Scholar] [PubMed]
- Byun, H.J.; Jung, W.W.; Lee, J.B.; Chung, H.Y.; Sul, D.; Kim, S.J.; Park, C.G.; Choi, I.; Hwang, K.W.; Chun, T. An evaluation of the neonatal immune system using a listeria infection model. Neonatology 2007, 92, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Lantier, L.; Lacroix-Lamande, S.; Potiron, L.; Metton, C.; Drouet, F.; Guesdon, W.; Gnahoui-David, A.; Le Vern, Y.; Deriaud, E.; Fenis, A.; et al. Intestinal CD103+ dendritic cells are key players in the innate immune control of Cryptosporidium parvum infection in neonatal mice. PLoS Pathog. 2013, 9, e1003801. [Google Scholar] [CrossRef]
- Altobelli, A.; Bauer, M.; Velez, K.; Cover, T.L.; Müller, A. Helicobacter pylori VacA Targets Myeloid Cells in the Gastric Lamina Propria To Promote Peripherally Induced Regulatory T-Cell Differentiation and Persistent Infection. mBio 2019, 10, e00261–e00219. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Gong, M.; Colliou, N.; Zadeh, M.; Li, J.; Jones, D.P.; Li, S.; Mohamadzadeh, M. Neonatal intestinal immune regulation by the commensal bacterium, P. UF1. Mucosal. Immunol. 2019, 12, 434–444. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Tian, S.; Yu, H.; Wang, J.; Zhu, W. Response of Colonic Mucosa-Associated Microbiota Composition, Mucosal Immune Homeostasis, and Barrier Function to Early Life Galactooligosaccharides Intervention in Suckling Piglets. J. Agric. Food Chem. 2019, 67, 578–588. [Google Scholar] [CrossRef] [PubMed]
- Hrdy, J.; Vlasakova, K.; Cerny, V.; Sukenikova, L.; Novotna, O.; Petraskova, P.; Borakova, K.; Lodinova-Zadnikova, R.; Kolarova, L.; Prokesova, L. Decreased allergy incidence in children supplemented with E. coli O83:K24:H31 and its possible modes of action. Eur. J. Immunol. 2018, 48, 2015–2030. [Google Scholar] [CrossRef]
- Cheng, R.Y.; Yao, J.R.; Wan, Q.; Guo, J.W.; Pu, F.F.; Shi, L.; Hu, W.; Yang, Y.H.; Li, L.; Li, M.; et al. Oral administration of Bifidobacterium bifidum TMC3115 to neonatal mice may alleviate IgE-mediated allergic risk in adulthood. Benef. Microbes 2018, 9, 815–828. [Google Scholar] [CrossRef] [PubMed]
- Echeverry, A.; Schesser, K.; Adkins, B. Murine neonates are highly resistant to Yersinia enterocolitica following orogastric exposure. Infect. Immun. 2007, 75, 2234–2243. [Google Scholar] [CrossRef]
- Echeverry, A.; Saijo, S.; Schesser, K.; Adkins, B. Yersinia enterocolitica promotes robust mucosal inflammatory T-cell immunity in murine neonates. Infect. Immun. 2010, 78, 3595–3608. [Google Scholar] [CrossRef]
- Siefker, D.T.; Adkins, B. Rapid CD8(+) Function Is Critical for Protection of Neonatal Mice from an Extracellular Bacterial Enteropathogen. Front. Pediatr. 2016, 4, 141. [Google Scholar] [CrossRef] [PubMed]
- Haas, J.D.; Ravens, S.; Düber, S.; Sandrock, I.; Oberdörfer, L.; Kashani, E.; Chennupati, V.; Föhse, L.; Naumann, R.; Weiss, S.; et al. Development of interleukin-17-producing γδ T cells is restricted to a functional embryonic wave. Immunity 2012, 37, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, H.S.; Liu, Y.; Menkiti, O.R.; Mei, J.; Dai, N.; O’Leary, C.E.; Oliver, P.M.; Kolls, J.K.; Weiser, J.N.; Worthen, G.S. The microbiota regulates neutrophil homeostasis and host resistance to Escherichia coli K1 sepsis in neonatal mice. Nat. Med. 2014, 20, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Chen, K.; Xiang, Y.; Yoshimura, T.; Su, S.; Zhu, J.; Bian, X.w.; Wang, J.M. New development in studies of formyl-peptide receptors: Critical roles in host defense. J. Leukoc. Biol. 2015, 99, 425–435. [Google Scholar] [CrossRef]
- Jones, R.M.; Neish, A.S. Recognition of bacterial pathogens and mucosal immunity. Cell. Microbiol. 2011, 13, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Aymeric, L.; Sansonetti, P. Discriminating Pathogens from Commensals at Mucosal Surfaces. Mucosal Immunol. 2015, 1, 975–984. [Google Scholar]
- Ye, R.D.; Boulay, F.; Wang, J.M.; Dahlgren, C.; Gerard, C.; Parmentier, M.; Serhan, C.N.; Murphy, P.M. International Union of Basic and Clinical Pharmacology. LXXIII. Nomenclature for the Formyl Peptide Receptor (FPR) Family. Pharmacol. Rev. 2009, 61, 119–161. [Google Scholar] [CrossRef] [PubMed]
- Ward, P.A.; Lepow, I.H.; Newman, L.J. Bacterial factors chemotactic for polymorphonuclear leukocytes. Am. J. Pathol. 1968, 52, 725–736. [Google Scholar] [PubMed]
- Schiffmann, E.; Showell, H.V.; Corcoran, B.A.; Ward, P.A.; Smith, E.; Becker, E.L. The isolation and partial characterization of neutrophil chemotactic factors from Escherichia coli. J. Immunol. 1975, 114, 1831–1837. [Google Scholar]
- Spurr, L.; Nadkarni, S.; Pederzoli-Ribeil, M.; Goulding, N.J.; Perretti, M.; D’Acquisto, F. Comparative analysis of Annexin A1-formyl peptide receptor 2/ALX expression in human leukocyte subsets. Int. Immunopharmacol. 2011, 11, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Chen, Q.; Le, Y.; Wang, J.M.; Oppenheim, J.J. Differential Regulation of Formyl Peptide Receptor-Like 1 Expression During the Differentiation of Monocytes to Dendritic Cells and Macrophages. J. Immunol. 2001, 166, 4092–4098. [Google Scholar] [CrossRef]
- Coers, J.; Gemperle, C.; Schmid, M.; Herova, M.; Marti-Jaun, J.; Wuest, S.J.A.; Loretz, C.; Hersberger, M. Regulation of the Formyl Peptide Receptor 1 (FPR1) Gene in Primary Human Macrophages. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Yang, D.; Chen, Q.; Gertz, B.; He, R.; Phulsuksombati, M.; Ye, R.D.; Oppenheim, J.J. Human dendritic cells express functional formyl peptide receptor-like-2 (FPRL2) throughout maturation. J. Leukoc. Biol. 2002, 72, 598–607. [Google Scholar] [PubMed]
- Anton, P.; O’Connell, J.; O’Connell, D.; Whitaker, L.; O’Sullivan, G.C.; Collins, J.K.; Shanahan, F. Mucosal subepithelial binding sites for the bacterial chemotactic peptide, formyl-methionyl-leucyl-phenylalanine (FMLP). Gut 1998, 42, 374–379. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Keitoku, M.; Kohzuki, M.; Katoh, H.; Funakoshi, M.; Suzuki, S.; Takeuchi, M.; Karibe, A.; Horiguchi, S.; Watanabe, J.; Satoh, S.; et al. FMLP Actions and its Binding Sites in Isolated Human Coronary Arteries. J. Mol. Cell. Cardiol. 1997, 29, 881–894. [Google Scholar] [CrossRef]
- Chen, K.; Iribarren, P.; Huang, J.; Zhang, L.; Gong, W.; Cho, E.H.; Lockett, S.; Dunlop, N.M.; Wang, J.M. Induction of the Formyl Peptide Receptor 2 in Microglia by IFN-γ and Synergy with CD40 Ligand. J. Immunol. 2007, 178, 1759–1766. [Google Scholar] [CrossRef]
- Wentworth, C.C.; Jones, R.M.; Kwon, Y.M.; Nusrat, A.; Neish, A.S. Commensal-Epithelial Signaling Mediated via Formyl Peptide Receptors. Am. J. Pathol. 2010, 177, 2782–2790. [Google Scholar] [CrossRef]
- Leoni, G.; Alam, A.; Neumann, P.-A.; Lambeth, J.D.; Cheng, G.; McCoy, J.; Hilgarth, R.S.; Kundu, K.; Murthy, N.; Kusters, D.; et al. Annexin A1, formyl peptide receptor, and NOX1 orchestrate epithelial repair. J. Clin. Investig. 2012, 123, 443–454. [Google Scholar] [CrossRef]
- Swanson, P.A.; Kumar, A.; Samarin, S.; Vijay-Kumar, M.; Kundu, K.; Murthy, N.; Hansen, J.; Nusrat, A.; Neish, A.S. Enteric commensal bacteria potentiate epithelial restitution via reactive oxygen species-mediated inactivation of focal adhesion kinase phosphatases. Proc. Natl. Acad. Sci. USA 2011, 108, 8803–8808. [Google Scholar] [CrossRef]
- Alam, A.; Leoni, G.; Wentworth, C.C.; Kwal, J.M.; Wu, H.; Ardita, C.S.; Swanson, P.A.; Lambeth, J.D.; Jones, R.M.; Nusrat, A.; et al. Redox signaling regulates commensal-mediated mucosal homeostasis and restitution and requires formyl peptide receptor 1. Mucosal. Immunol. 2014, 7, 645–655. [Google Scholar] [CrossRef]
- Babbin, B.A.; Jesaitis, A.J.; Ivanov, A.I.; Kelly, D.; Laukoetter, M.; Nava, P.; Parkos, C.A.; Nusrat, A. Formyl Peptide Receptor-1 Activation Enhances Intestinal Epithelial Cell Restitution through Phosphatidylinositol 3-Kinase-Dependent Activation of Rac1 and Cdc42. J. Immunol. 2007, 179, 8112–8121. [Google Scholar] [CrossRef] [PubMed]
- Babbin, B.A.; Lee, W.Y.; Parkos, C.A.; Winfree, L.M.; Akyildiz, A.; Perretti, M.; Nusrat, A. Annexin I Regulates SKCO-15 Cell Invasion by Signaling through Formyl Peptide Receptors. J. Biol. Chem. 2006, 281, 19588–19599. [Google Scholar] [CrossRef] [PubMed]
- Canny, G.; Levy, O.; Furuta, G.T.; Narravula-Alipati, S.; Sisson, R.B.; Serhan, C.N.; Colgan, S.P. Lipid mediator-induced expression of bactericidal/ permeability-increasing protein (BPI) in human mucosal epithelia. Proc. Natl. Acad. Sci. USA 2002, 99, 3902–3907. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, D.; Gleske, A.-K.; Rautenberg, M.; Wang, R.; Köberle, M.; Bohn, E.; Schöneberg, T.; Rabiet, M.-J.; Boulay, F.; Klebanoff, S.J.; et al. Human Formyl Peptide Receptor 2 Senses Highly Pathogenic Staphylococcus aureus. Cell Host Microbe 2010, 7, 463–473. [Google Scholar] [CrossRef]
- Fillion, I.; Ouellet, N.; Simard, M.; Bergeron, Y.; Sato, S.; Bergeron, M.G. Role of Chemokines and Formyl Peptides in Pneumococcal Pneumonia-Induced Monocyte/Macrophage Recruitment. J. Immunol. 2001, 166, 7353–7361. [Google Scholar] [CrossRef]
- Lala, A.; Amano, A.; Sojar, H.T.; Radel, S.J.; Denardin, E. Porphyromonas gingivalis Trypsin-Like Protease: A Possible Natural Ligand for the Neutrophil Formyl Peptide Receptor. Biochem. Biophys. Res. Commun. 1994, 199, 1489–1496. [Google Scholar] [CrossRef]
- Uriarte, S.M.; Edmisson, J.S.; Jimenez-Flores, E. Human neutrophils and oral microbiota: A constant tug-of-war between a harmonious and a discordant coexistence. Immunol. Rev. 2016, 273, 282–298. [Google Scholar] [CrossRef]
- Basha, S.; Surendran, N.; Pichichero, M. Immune responses in neonates. Expert Rev. Clin. Immunol. 2014, 10, 1171–1184. [Google Scholar] [CrossRef]
- Dzidic, M.; Boix-Amorós, A.; Selma-Royo, M.; Mira, A.; Collado, M. Gut microbiota and mucosal immunity in the neonate. Med. Sci. 2018, 6, 56. [Google Scholar] [CrossRef]
- Stålhammar, M.E.; Douhan Håkansson, L.; Jonzon, A.; Sindelar, R. Differential neutrophil chemotactic response towards IL-8 and bacterial N-formyl peptides in term newborn infants. Upsala J. Med. Sci. 2016, 122, 35–42. [Google Scholar] [CrossRef]
- Stålhammar, M.E.; Douhan Håkansson, L.; Sindelar, R. Bacterial N-formyl Peptides Reduce PMA- and Escherichia coli-Induced Neutrophil Respiratory Burst in Term Neonates and Adults. Scand. J. Immunol. 2017, 85, 365–371. [Google Scholar] [CrossRef]
- Liu, W.; Huang, J.; Doycheva, D.; Gamdzyk, M.; Tang, J.; Zhang, J.H. RvD1binding with FPR2 attenuates inflammation via Rac1/NOX2 pathway after neonatal hypoxic-ischemic injury in rats. Exp. Neurol. 2019, 320. [Google Scholar] [CrossRef] [PubMed]
- Rubino, T.; Gallo, I.; Rattazzi, L.; Piras, G.; Gobbetti, T.; Panza, E.; Perretti, M.; Dalley, J.W.; D’Acquisto, F. Formyl Peptide Receptor as a Novel Therapeutic Target for Anxiety-Related Disorders. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Kim, S.D.; Kim, Y.-K.; Lee, H.Y.; Kim, Y.-S.; Jeon, S.G.; Baek, S.-H.; Song, D.-K.; Ryu, S.H.; Bae, Y.-S. The Agonists of Formyl Peptide Receptors Prevent Development of Severe Sepsis after Microbial Infection. J. Immunol. 2010, 185, 4302–4310. [Google Scholar] [CrossRef]
- Kao, W.; Gu, R.; Jia, Y.; Wei, X.; Fan, H.; Harris, J.; Zhang, Z.; Quinn, J.; Morand, E.F.; Yang, Y.H. A formyl peptide receptor agonist suppresses inflammation and bone damage in arthritis. Br. J. Pharmacol. 2014, 171, 4087–4096. [Google Scholar] [CrossRef] [PubMed]
- Linehan, J.L.; Harrison, O.J.; Han, S.-J.; Byrd, A.L.; Vujkovic-Cvijin, I.; Villarino, A.V.; Sen, S.K.; Shaik, J.; Smelkinson, M.; Tamoutounour, S.; et al. Non-classical immunity controls microbiota impact on skin immunity and tissue repair. Cell 2018, 172, 784–796.e18. [Google Scholar] [CrossRef]
- Dominguez-Andres, J.; Netea, M.G. Long-term reprogramming of the innate immune system. J. Leukoc. Biol. 2018, 105, 329–338. [Google Scholar] [CrossRef]
- Wang, Y.; Kalka-Moll, W.M.; Roehrl, M.H.; Kasper, D.L. Structural basis of the abscess-modulating polysaccharide A2 from Bacteroides fragilis. Proc. Natl. Acad. Sci. USA 2000, 97, 13478–13483. [Google Scholar] [CrossRef]
- Cobb, B.A.; Kasper, D.L. Characteristics of carbohydrate antigen binding to the presentation protein HLA-DR. Glycobiology 2008, 18, 707–718. [Google Scholar] [CrossRef] [PubMed]
- Kalka-Moll, W.M.; Tzianabos, A.O.; Bryant, P.W.; Niemeyer, M.; Ploegh, H.L.; Kasper, D.L. Zwitterionic polysaccharides stimulate T cells by MHC class II-dependent interactions. J. Immunol. 2002, 169, 6149–6153. [Google Scholar] [CrossRef] [PubMed]
- Baumann, H.; Tzianabos, A.O.; Brisson, J.R.; Kasper, D.L.; Jennings, H.J. Structural elucidation of two capsular polysaccharides from one strain of Bacteroides fragilis using high-resolution NMR spectroscopy. Biochemistry 1992, 31, 4081–4089. [Google Scholar] [CrossRef]
- Jost, T.; Lacroix, C.; Braegger, C.P.; Chassard, C. New insights in gut microbiota establishment in healthy breast fed neonates. PLoS ONE 2012, 7, e44595. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, C.A.; Jones, M.B.; Hambor, J.; Cobb, B.A. Characterization of Polysaccharide A Response Reveals Interferon Responsive Gene Signature and Immunomodulatory Marker Expression. Front. Immunol. 2020, 11, 556813. [Google Scholar] [CrossRef]
- Sharpe, A.H.; Pauken, K.E. The diverse functions of the PD1 inhibitory pathway. Nat. Rev. Immunol. 2018, 18, 153–167. [Google Scholar] [CrossRef]
- Sofi, M.H.; Johnson, B.M.; Gudi, R.R.; Jolly, A.; Gaudreau, M.C.; Vasu, C. Polysaccharide A-Dependent Opposing Effects of Mucosal and Systemic Exposures to Human Gut Commensal Bacteroides fragilis in Type 1 Diabetes. Diabetes 2019, 68, 1975–1989. [Google Scholar] [CrossRef]
- Ramakrishna, C.; Kujawski, M.; Chu, H.; Li, L.; Mazmanian, S.K.; Cantin, E.M. Bacteroides fragilis polysaccharide A induces IL-10 secreting B and T cells that prevent viral encephalitis. Nat. Commun. 2019, 10, 2153. [Google Scholar] [CrossRef]
- Ryan, S.O.; Bonomo, J.A.; Zhao, F.; Cobb, B.A. MHCII glycosylation modulates Bacteroides fragilis carbohydrate antigen presentation. J. Exp. Med. 2011, 208, 1041–1053. [Google Scholar] [CrossRef]
- Cobb, B.A.; Wang, Q.; Tzianabos, A.O.; Kasper, D.L. Polysaccharide processing and presentation by the MHCII pathway. Cell 2004, 117, 677–687. [Google Scholar] [CrossRef]
- Johnson, J.L.; Jones, M.B.; Cobb, B.A. Polysaccharide A from the capsule of Bacteroides fragilis induces clonal CD4+ T cell expansion. J. Biol. Chem. 2015, 290, 5007–5014. [Google Scholar] [CrossRef]
- Johnson, J.L.; Jones, M.B.; Cobb, B.A. Bacterial capsular polysaccharide prevents the onset of asthma through T-cell activation. Glycobiology 2015, 25, 368–375. [Google Scholar] [CrossRef]
- Jiang, F.; Meng, D.; Weng, M.; Zhu, W.; Wu, W.; Kasper, D.; Walker, W.A. The symbiotic bacterial surface factor polysaccharide A on Bacteroides fragilis inhibits IL-1β-induced inflammation in human fetal enterocytes via toll receptors 2 and 4. PLoS ONE 2017, 12, e0172738. [Google Scholar] [CrossRef]
- Dasgupta, S.; Erturk-Hasdemir, D.; Ochoa-Reparaz, J.; Reinecker, H.C.; Kasper, D.L. Plasmacytoid dendritic cells mediate anti-inflammatory responses to a gut commensal molecule via both innate and adaptive mechanisms. Cell Host Microbe 2014, 15, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.K.; Kasper, D.L.; Wang, B.; Forsythe, P.; Bienenstock, J.; Kunze, W.A. Bacteroides fragilis polysaccharide A is necessary and sufficient for acute activation of intestinal sensory neurons. Nat. Commun. 2013, 4, 1465. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Repáraz, J.; Mielcarz, D.W.; Ditrio, L.E.; Burroughs, A.R.; Begum-Haque, S.; Dasgupta, S.; Kasper, D.L.; Kasper, L.H. Central nervous system demyelinating disease protection by the human commensal Bacteroides fragilis depends on polysaccharide A expression. J. Immunol. 2010, 185, 4101–4108. [Google Scholar] [CrossRef] [PubMed]
- Rogers, A.C.; McDermott, F.D.; Mohan, H.M.; O’Connell, P.R.; Winter, D.C.; Baird, A.W. The effects of polyamines on human colonic mucosal function. Eur. J. Pharmacol. 2015, 764, 157–163. [Google Scholar] [CrossRef]
- Sakanaka, M.; Sugiyama, Y.; Kitakata, A.; Katayama, T.; Kurihara, S. Carboxyspermidine decarboxylase of the prominent intestinal microbiota species Bacteroides thetaiotaomicron is required for spermidine biosynthesis and contributes to normal growth. Amino Acids 2016, 48, 2443–2451. [Google Scholar] [CrossRef]
- Dufour, C.; Dandrifosse, G.; Forget, P.; Vermesse, F.; Romain, N.; Lepoint, P. Spermine and spermidine induce intestinal maturation in the rat. Gastroenterology 1988, 95, 112–116. [Google Scholar] [CrossRef]
- Löser, C.; Eisel, A.; Harms, D.; Fölsch, U.R. Dietary polyamines are essential luminal growth factors for small intestinal and colonic mucosal growth and development. Gut 1999, 44, 12–16. [Google Scholar] [CrossRef]
- Gangoiti, P.; Camacho, L.; Arana, L.; Ouro, A.; Granado, M.H.; Brizuela, L.; Casas, J.; Fabriás, G.; Abad, J.L.; Delgado, A.; et al. Control of metabolism and signaling of simple bioactive sphingolipids: Implications in disease. Prog. Lipid. Res. 2010, 49, 316–334. [Google Scholar] [CrossRef]
- Olsen, I.; Jantzen, E. Sphingolipids in Bacteria and Fungi. Anaerobe 2001, 7, 103–112. [Google Scholar] [CrossRef]
- Harrison, P.J.; Dunn, T.M.; Campopiano, D.J. Sphingolipid biosynthesis in man and microbes. Nat. Prod. Rep. 2018, 35, 921–954. [Google Scholar] [CrossRef] [PubMed]
- An, D.; Oh, S.F.; Olszak, T.; Neves, J.F.; Avci, F.Y.; Erturk-Hasdemir, D.; Lu, X.; Zeissig, S.; Blumberg, R.S.; Kasper, D.L. Sphingolipids from a symbiotic microbe regulate homeostasis of host intestinal natural killer T cells. Cell 2014, 156, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.M.; Ke, X.; Hitchcock, D.; Jeanfavre, S.; Avila-Pacheco, J.; Nakata, T.; Arthur, T.D.; Fornelos, N.; Heim, C.; Franzosa, E.A.; et al. Bacteroides-derived sphingolipids are critical for maintaining intestinal homeostasis and symbiosis. Cell Host Microbe 2019, 25, 668–680.e7. [Google Scholar] [CrossRef]
- Stockinger, B.; Shah, K.; Wincent, E. AHR in the intestinal microenvironment: Safeguarding barrier function. Nat. Rev. Gastroenterol. Hepatol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Juricek, L.; Coumoul, X. The Aryl Hydrocarbon Receptor and the Nervous System. Int. J. Mol. Sci. 2018, 19, 2504. [Google Scholar] [CrossRef] [PubMed]
- Kimura, E.; Tohyama, C. Embryonic and Postnatal Expression of Aryl Hydrocarbon Receptor mRNA in Mouse Brain. Front. Neuroanat. 2017, 11, 4. [Google Scholar] [CrossRef]
- Wong, C.B.; Tanaka, A.; Kuhara, T.; Xiao, J.Z. Potential Effects of Indole-3-Lactic Acid, a Metabolite of Human Bifidobacteria, on NGF-induced Neurite Outgrowth in PC12 Cells. Microorganisms 2020, 8, 398. [Google Scholar] [CrossRef]
- Li, Y.; Wang, K.; Zou, Q.Y.; Zhou, C.; Magness, R.R.; Zheng, J. A possible role of aryl hydrocarbon receptor in spontaneous preterm birth. Med. Hypotheses 2015, 84, 494–497. [Google Scholar] [CrossRef]
- Vorderstrasse, B.A.; Cundiff, J.A.; Lawrence, B.P. Developmental exposure to the potent aryl hydrocarbon receptor agonist 2,3,7,8-tetrachlorodibenzo-p-dioxin Impairs the cell-mediated immune response to infection with influenza a virus, but enhances elements of innate immunity. J. Immunotoxicol. 2004, 1, 103–112. [Google Scholar] [CrossRef][Green Version]
- Moura-Alves, P.; Faé, K.; Houthuys, E.; Dorhoi, A.; Kreuchwig, A.; Furkert, J.; Barison, N.; Diehl, A.; Munder, A.; Constant, P.; et al. AhR sensing of bacterial pigments regulates antibacterial defence. Nature 2014, 512, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Bock, K.W. Aryl hydrocarbon receptor (AHR)-mediated inflammation and resolution: Non-genomic and genomic signaling. Biochem. Pharmacol. 2020, 182, 114220. [Google Scholar] [CrossRef] [PubMed]
- Gomez de Agüero, M.; Ganal-Vonarburg, S.C.; Fuhrer, T.; Rupp, S.; Uchimura, Y.; Li, H.; Steinert, A.; Heikenwalder, M.; Hapfelmeier, S.; Sauer, U.; et al. The maternal microbiota drives early postnatal innate immune development. Science 2016, 351, 1296–1302. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Yamaguchi, Y.; Fulton, W.B.; Wang, S.; Zhou, Q.; Jia, H.; Kovler, M.L.; Salazar, A.G.; Sampah, M.; Prindle, T., Jr.; et al. Maternal aryl hydrocarbon receptor activation protects newborns against necrotizing enterocolitis. Nat. Commun. 2021, 12, 1042. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, X.; Zhang, J.; Luo, Y.; Xu, B.; Ling, S.; Zhang, Y.; Li, W.; Yao, X. Activation of aryl hydrocarbon receptor in Langerhans cells by a microbial metabolite of tryptophan negatively regulates skin inflammation. J. Dermatol. Sci. 2020, 100, 192–200. [Google Scholar] [CrossRef]
- Kimura, A.; Abe, H.; Tsuruta, S.; Chiba, S.; Fujii-Kuriyama, Y.; Sekiya, T.; Morita, R.; Yoshimura, A. Aryl hydrocarbon receptor protects against bacterial infection by promoting macrophage survival and reactive oxygen species production. Int. Immunol. 2014, 26, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.Y.; Tung, Y.T.; Yang, Y.S.H.; Hsu, J.B.; Lee, C.Y.; Chang, T.H.; Su, E.C.; Hsieh, R.H.; Chen, Y.C. Maternal vegetable and fruit consumption during pregnancy and its effects on infant gut microbiome. Nutrients 2021, 13, 1559. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.J.; Yeung, C.Y. Recent advance in infant nutrition: Human milk oligosaccharides. Pediatr. Neonatol. 2021. [Google Scholar] [CrossRef]
- Gómez-Gallego, C.; Collado, M.C.; Pérez, G.; Ilo, T.; Jaakkola, U.M.; Bernal, M.J.; Periago, M.J.; Frias, R.; Ros, G.; Salminen, S. Resembling breast milk: Influence of polyamine-supplemented formula on neonatal BALB/cOlaHsd mouse microbiota. Br. J. Nutr. 2014, 111, 1050–1058. [Google Scholar] [CrossRef]
- Gomez-Gallego, C.; Garcia Romo, M.; Frias, R.; Periago, M.J.; Ros, G.; Salminen, S.; Collado, M.C. Mice exposed to infant formula enriched with polyamines: Impact on host transcriptome and microbiome. Food Funct. 2017, 8, 1622–1626. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.J.; Lynch, D.B.; Murphy, K.; Ulaszewska, M.; Jeffery, I.B.; O’Shea, C.A.; Watkins, C.; Dempsey, E.; Mattivi, F.; Tuohy, K.; et al. Evolution of gut microbiota composition from birth to 24 weeks in the INFANTMET Cohort. Microbiome 2017, 5, 4. [Google Scholar] [CrossRef]
- Korpela, K.; Helve, O.; Kolho, K.L.; Saisto, T.; Skogberg, K.; Dikareva, E.; Stefanovic, V.; Salonen, A.; Andersson, S.; de Vos, W.M. Maternal Fecal Microbiota Transplantation in Cesarean-Born Infants Rapidly Restores Normal Gut Microbial Development: A Proof-of-Concept Study. Cell 2020, 183, 324–334.e5. [Google Scholar] [CrossRef]
- Dominguez-Bello, M.G.; De Jesus-Laboy, K.M.; Shen, N.; Cox, L.M.; Amir, A.; Gonzalez, A.; Bokulich, N.A.; Song, S.J.; Hoashi, M.; Rivera-Vinas, J.I.; et al. Partial restoration of the microbiota of cesarean-born infants via vaginal microbial transfer. Nat. Med. 2016, 22, 250–253. [Google Scholar] [CrossRef]
- Butler, É.M.; Chiavaroli, V.; Derraik, J.G.B.; Grigg, C.P.; Wilson, B.C.; Walker, N.; O’Sullivan, J.M.; Cutfield, W.S. Maternal bacteria to correct abnormal gut microbiota in babies born by C-section. Medicine 2020, 99, e21315. [Google Scholar] [CrossRef] [PubMed]
- Athalye-Jape, G.; Rao, S.; Patole, S. Effects of probiotics on experimental necrotizing enterocolitis: A systematic review and meta-analysis. Pediatr Res 2018, 83, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.P.; Chow, S.S.W.; Alsweiler, J.; Bourchier, D.; Broadbent, R.; Knight, D.; Lynn, A.M.; Patel, H. Probiotics for Prevention of Severe Necrotizing Enterocolitis: Experience of New Zealand Neonatal Intensive Care Units. Front Pediatr 2020, 8, 119. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Metabolite | Localization | Concentration | References |
|---|---|---|---|
| Total SCFAs | Caecum | 131 mmol/kg (dry weight) | [42,43] |
| Portal | 323–375 µmol/L | ||
| Hepatic | 148–238 µmol/L | ||
| Blood | 79–184 µmol/L |
| Metabolite | Receptor | EC50 | References |
|---|---|---|---|
| Acetate | GPR 41 | 1.02–2.99 mM | [50,51,53,56,61] |
| GPR 43 | 52 µM–4.46 mM | ||
| GPR 109A | >10 mM | ||
| OR51E2 | 2.93 mM | ||
| Propionate | GPR 41 | 11.6 µM–5.21 mM | |
| GPR 43 | 31 µM–4.85 mM | ||
| GPR 109A | >10 mM | ||
| OR51E2 | ~100 µM–2.16 mM | ||
| Butyrate | GPR 41 | 158 µM–4.38 mM | |
| GPR 43 | 100 µM–4.55 mM | ||
| GPR 109A | 1.6 mM | ||
| OR51E2 | unknown |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phillips-Farfán, B.; Gómez-Chávez, F.; Medina-Torres, E.A.; Vargas-Villavicencio, J.A.; Carvajal-Aguilera, K.; Camacho, L. Microbiota Signals during the Neonatal Period Forge Life-Long Immune Responses. Int. J. Mol. Sci. 2021, 22, 8162. https://doi.org/10.3390/ijms22158162
Phillips-Farfán B, Gómez-Chávez F, Medina-Torres EA, Vargas-Villavicencio JA, Carvajal-Aguilera K, Camacho L. Microbiota Signals during the Neonatal Period Forge Life-Long Immune Responses. International Journal of Molecular Sciences. 2021; 22(15):8162. https://doi.org/10.3390/ijms22158162
Chicago/Turabian StylePhillips-Farfán, Bryan, Fernando Gómez-Chávez, Edgar Alejandro Medina-Torres, José Antonio Vargas-Villavicencio, Karla Carvajal-Aguilera, and Luz Camacho. 2021. "Microbiota Signals during the Neonatal Period Forge Life-Long Immune Responses" International Journal of Molecular Sciences 22, no. 15: 8162. https://doi.org/10.3390/ijms22158162
APA StylePhillips-Farfán, B., Gómez-Chávez, F., Medina-Torres, E. A., Vargas-Villavicencio, J. A., Carvajal-Aguilera, K., & Camacho, L. (2021). Microbiota Signals during the Neonatal Period Forge Life-Long Immune Responses. International Journal of Molecular Sciences, 22(15), 8162. https://doi.org/10.3390/ijms22158162