
Epigenetic Regulation of microRNAs in Cancer: Shortening the Distance from Bench to Bedside

 , ,
, ,
Abstract
:1. Introduction
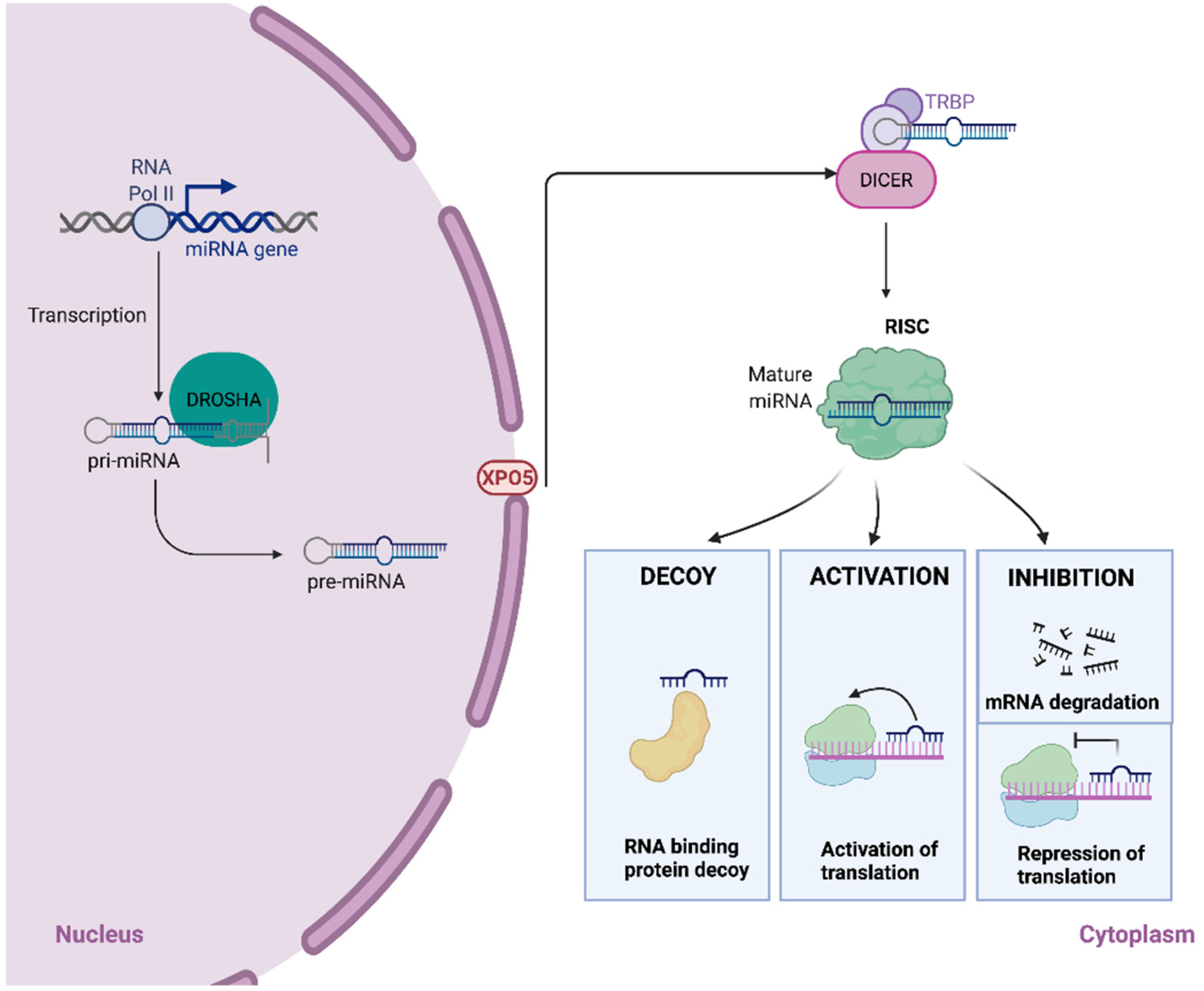
2. The Biogenesis of miRNAs
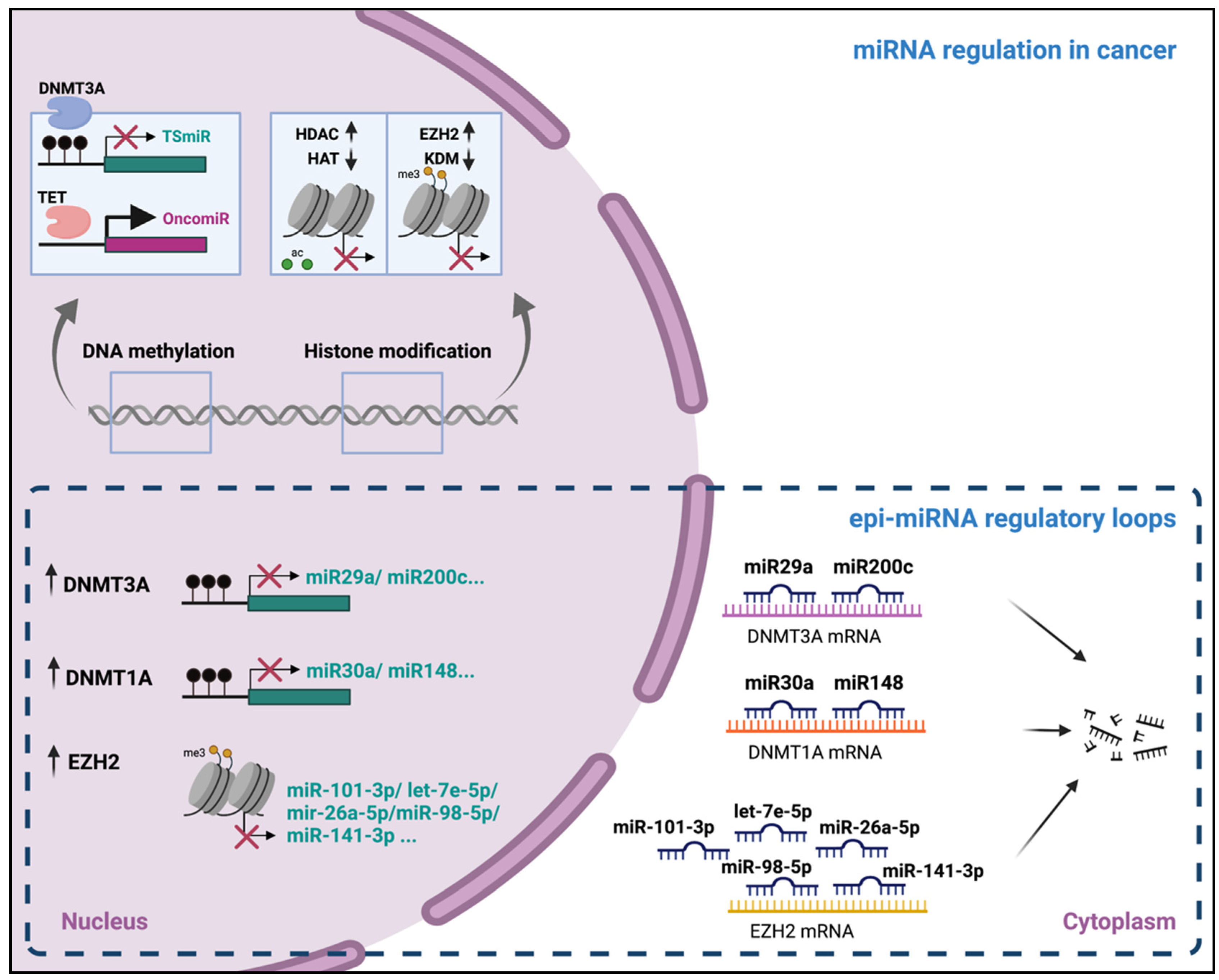
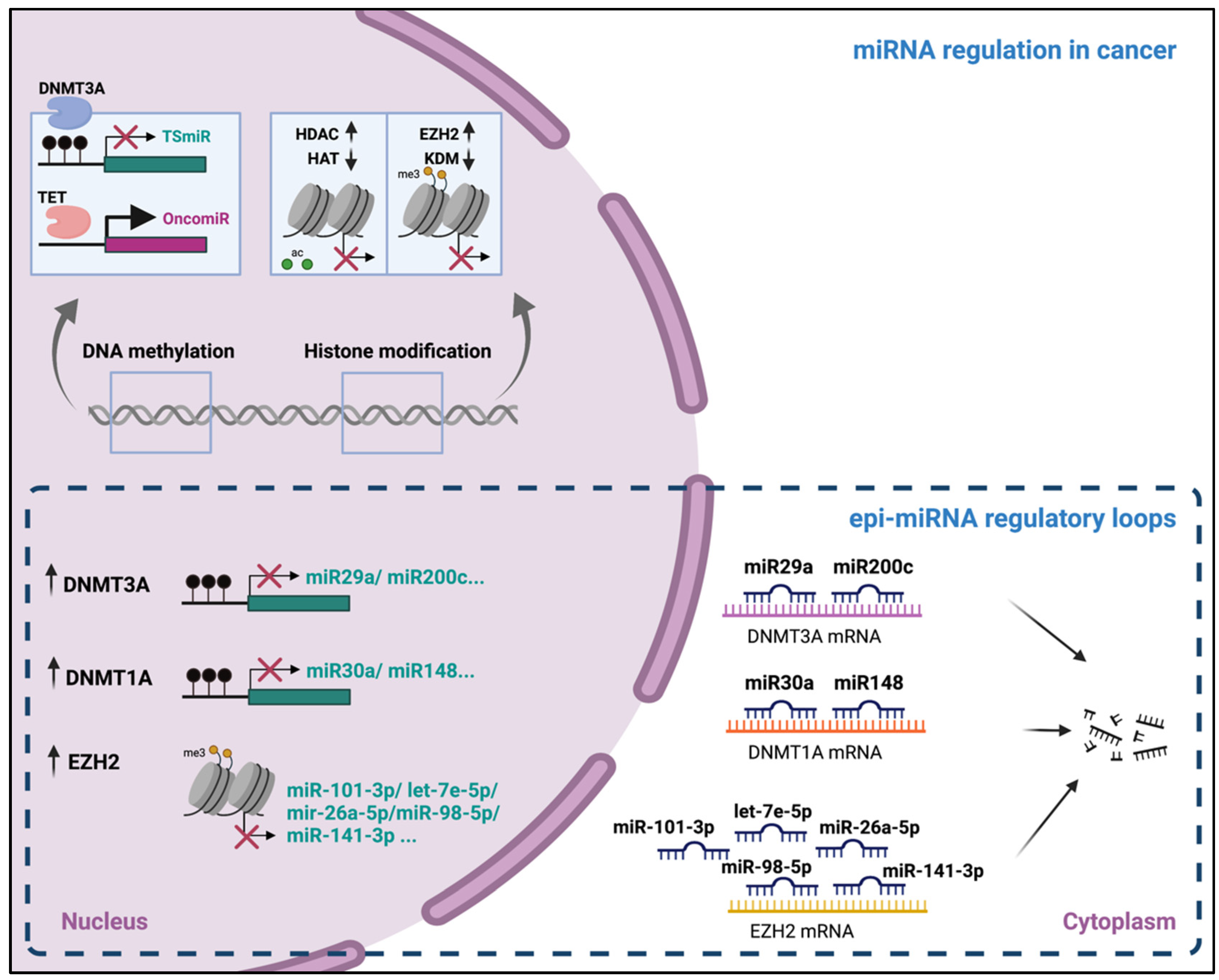
3. Epigenetic Regulation of miRNAs Expression
3.1. DNA Methylation
3.2. Histone Modifications
3.2.1. Histone Acetylation
3.2.2. Histone Methylation
3.3. RNA Modifications
4. miRNAs as Epigenetic Regulators
5. Epigenetic Alteration and microRNA Dysregulation in Cancer
5.1. The Cancer Epigenome Landscape
5.2. miRNAs in the Control of Critical Cancer-Related Pathways
5.3. Bidirectional Relationship between Epigenetic Alterations and miRNA Dysregulation: Cases with Biological Relevance in Cancer Diseases
5.3.1. miRNAs and Lung Cancer
5.3.2. miRNAs and Gastric Cancer
5.3.3. miRNAs and Ovarian Cancer
5.3.4. miRNAs and Breast Cancer
6. Clinical Applications: miRNAs Epigenetics in Cancer
6.1. miRNAs Methylation as Diagnostic Biomarkers
6.2. Epigenetic Regulation of miRNAs as Prognostic Biomarkers
6.3. Epigenetic Regulation of miRNAs as a Therapeutic Strategy in Cancer
7. Conclusions
Funding
Conflicts of Interest
References
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Kota, J.; Chivukula, R.R.; O’Donnell, K.A.; Wentzel, E.A.; Montgomery, C.L.; Hwang, H.-W.; Chang, T.-C.; Vivekanandan, P.; Torbenson, M.; Clark, K.R.; et al. Therapeutic microRNA Delivery Suppresses Tumorigenesis in a Murine Liver Cancer Model. Cell 2009, 137, 1005–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tay, Y.; Zhang, J.; Thomson, A.M.; Lim, B.; Rigoutsos, I. MicroRNAs to Nanog, Oct4 and Sox2 coding regions modulate embryonic stem cell differentiation. Nature 2008, 455, 1124–1128. [Google Scholar] [CrossRef]
- Tapeh, B.E.G.; Mosayyebi, B.; Samei, M.; Basmenj, H.B.; Mohammadi, A.; Alivand, M.R.; Hassanpour, P.; Solali, S. microRNAs involved in T-cell development, selection, activation, and hemostasis. J. Cell. Physiol. 2020, 235, 8461–8471. [Google Scholar] [CrossRef] [PubMed]
- Ozsolak, F.; Poling, L.L.; Wang, Z.; Liu, H.; Liu, X.S.; Roeder, R.G.; Zhang, X.; Song, J.S.; Fisher, D.E. Chromatin structure analyses identify miRNA promoters. Genes Dev. 2008, 22, 3172–3183. [Google Scholar] [CrossRef] [Green Version]
- Iorio, M.V.; Piovan, C.; Croce, C.M. Interplay between microRNAs and the epigenetic machinery: An intricate network. Biochim. Biophys. Acta (BBA) Gene Regul. Mech. 2010, 1799, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Cai, X.; Yung, M.M.H.; Zhou, W.; Li, J.; Zhang, Y.; Li, Z.; Liu, S.S.; Cheung, A.N.Y.; Ngan, H.Y.S.; et al. miR-137 mediates the functional link between c-Myc and EZH2 that regulates cisplatin resistance in ovarian cancer. Oncogene 2019, 38, 564–580. [Google Scholar] [CrossRef]
- Syeda, Z.A.; Langden, S.S.S.; Munkhzul, C.; Lee, M.; Song, S.J. Regulatory Mechanism of MicroRNA Expression in Cancer. Int. J. Mol. Sci. 2020, 21, 1723. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, H.; Maruyama, R.; Yamamoto, E.; Kai, M. Epigenetic alteration and microRNA dysregulation in cancer. Front. Genet. 2013, 4, 258. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.-H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA genes are transcribed by RNA polymerase II. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Ahn, C.; Kim, V.N.; Han, J.; Choi, H.; Kim, J.; Yim, J.; Lee, J.; Provost, P.; Rådmark, O.; et al. The nuclear RNase III Drosha initiates microRNA processing. Nature 2003, 425, 415–419. [Google Scholar] [CrossRef]
- Han, J.; Lee, Y.; Yeom, K.-H.; Kim, Y.K.; Jin, H.; Kim, V.N. The Drosha-DGCR8 complex in primary microRNA processing. Genes Dev. 2004, 18, 3016–3027. [Google Scholar] [CrossRef] [Green Version]
- Gregory, R.I.; Yan, K.-P.; Amuthan, G.; Chendrimada, T.; Doratotaj, B.; Cooch, N.; Shiekhattar, R. The Microprocessor complex mediates the genesis of microRNAs. Nat. Cell Biol. 2004, 432, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Yi, R.; Qin, Y.; Macara, I.G.; Cullen, B.R. Exportin-5 mediates the nuclear export of pre-microRNAs and short hairpin RNAs. Genes Dev. 2003, 17, 3011–3016. [Google Scholar] [CrossRef] [Green Version]
- Lund, E.; Güttinger, S.; Calado, Â.; Dahlberg, J.E.; Kutay, U. Nuclear Export of MicroRNA Precursors. Science 2004, 303, 95–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutvagner, G.; McLachlan, J.; Pasquinelli, A.E.; Bálint, É.; Tuschl, T.; Zamore, P.D. A Cellular Function for the RNA-Interference Enzyme Dicer in the Maturation of the let-7 Small Temporal RNA. Science 2001, 293, 834–838. [Google Scholar] [CrossRef] [Green Version]
- Martinez, J.; Patkaniowska, A.; Urlaub, H.; Lührmann, R.; Tuschl, T. Single-Stranded Antisense siRNAs Guide Target RNA Cleavage in RNAi. Cell 2002, 110, 563–574. [Google Scholar] [CrossRef] [Green Version]
- Gregory, R.I.; Chendrimada, T.P.; Cooch, N.; Shiekhattar, R. Human RISC Couples MicroRNA Biogenesis and Posttranscriptional Gene Silencing. Cell 2005, 123, 631–640. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Love, T.M.; Call, M.E.; Doench, J.G.; Novina, C.D. Recapitulation of Short RNA-Directed Translational Gene Silencing In Vitro. Mol. Cell 2006, 22, 553–560. [Google Scholar] [CrossRef]
- Vasudevan, S.; Tong, Y.; Steitz, J.A. Switching from Repression to Activation: MicroRNAs Can Up-Regulate Translation. Science 2007, 318, 1931–1934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moretti, F.; Thermann, R.; Hentze, M.W. Mechanism of translational regulation by miR-2 from sites in the 5′ untranslated region or the open reading frame. RNA 2010, 16, 2493–2502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ørom, U.A.; Nielsen, F.C.; Lund, A.H. MicroRNA-10a Binds the 5′UTR of Ribosomal Protein mRNAs and Enhances Their Translation. Mol. Cell 2008, 30, 460–471. [Google Scholar] [CrossRef] [PubMed]
- Eiring, A.M.; Harb, J.G.; Neviani, P.; Garton, C.; Oaks, J.J.; Spizzo, R.; Liu, S.; Schwind, S.; Santhanam, R.; Hickey, C.J.; et al. miR-328 Functions as an RNA Decoy to Modulate hnRNP E2 Regulation of mRNA Translation in Leukemic Blasts. Cell 2010, 140, 652–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruby, J.G.; Jan, C.H.; Bartel, D.P. Intronic microRNA precursors that bypass Drosha processing. Nature 2007, 448, 83–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havens, M.A.; Reich, A.A.; Duelli, D.M.; Hastings, M.L. Biogenesis of mammalian microRNAs by a non-canonical processing pathway. Nucleic Acids Res. 2012, 40, 4626–4640. [Google Scholar] [CrossRef] [Green Version]
- Cheloufi, S.; Dos Santos, C.O.; Chong, M.M.W.; Hannon, G.J. A dicer-independent miRNA biogenesis pathway that requires Ago catalysis. Nature 2010, 465, 584–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antequera, F. Structure, function and evolution of CpG island promoters. Cell. Mol. Life Sci. CMLS 2003, 60, 1647–1658. [Google Scholar] [CrossRef]
- Jones, P.A. Functions of DNA methylation: Islands, start sites, gene bodies and beyond. Nat. Rev. Genet. 2012, 13, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.S.K.; Vertino, P.M. Orphan CpG islands define a novel class of highly active enhancers. Epigenetics 2017, 12, 449–464. [Google Scholar] [CrossRef] [Green Version]
- Neri, F.; Rapelli, S.; Krepelova, A.; Incarnato, D.; Parlato, C.; Basile, G.; Maldotti, M.; Anselmi, F.; Oliviero, S. Intragenic DNA methylation prevents spurious transcription initiation. Nature 2017, 543, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Piletič, K.; Kunej, T. MicroRNA epigenetic signatures in human disease. Arch. Toxicol. 2016, 90, 2405–2419. [Google Scholar] [CrossRef]
- Maiti, A. Mechanism of Active DNA Demethylation: Recent Progress in Epigenetics. J. Biomol. Res. Ther. 2012, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Saito, Y.; Liang, G.; Egger, G.; Friedman, J.M.; Chuang, J.C.; Coetzee, G.A.; Jones, P.A. Specific activation of microRNA-127 with downregulation of the proto-oncogene BCL6 by chromatin-modifying drugs in human cancer cells. Cancer Cell 2006, 9, 435–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuna, M.; Machado, A.S.; Calin, G.A. Genetic and epigenetic alterations of microRNAs and implications for human cancers and other diseases. Genes Chromosom. Cancer 2016, 55, 193–214. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.A.; Baylin, S.B. The Epigenomics of Cancer. Cell 2007, 128, 683–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audia, J.E.; Campbell, R.M. Histone Modifications and Cancer. Cold Spring Harb. Perspect. Biol. 2016, 8, a019521. [Google Scholar] [CrossRef] [PubMed]
- Kouzarides, T. Chromatin Modifications and Their Function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Paredes, M.; Esteller, M. Cancer epigenetics reaches mainstream oncology. Nat. Med. 2011, 17, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Eades, G.; Yang, M.; Yao, Y.; Zhang, Y.; Zhou, Q. miR-200a Regulates Nrf2 Activation by Targeting Keap1 mRNA in Breast Cancer Cells. J. Biol. Chem. 2011, 286, 40725–40733. [Google Scholar] [CrossRef] [Green Version]
- Bian, X.; Liang, Z.; Feng, A.; Salgado, E.; Shim, H. HDAC inhibitor suppresses proliferation and invasion of breast cancer cells through regulation of miR-200c targeting CRKL. Biochem. Pharmacol. 2018, 147, 30–37. [Google Scholar] [CrossRef]
- Nalls, D.; Tang, S.-N.; Rodova, M.; Srivastava, R.K.; Shankar, S. Targeting Epigenetic Regulation of miR-34a for Treatment of Pancreatic Cancer by Inhibition of Pancreatic Cancer Stem Cells. PLoS ONE 2011, 6, e24099. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.-H.; Dimri, M.; Dimri, G.P. MicroRNA-31 Is a Transcriptional Target of Histone Deacetylase Inhibitors and a Regulator of Cellular Senescence. J. Biol. Chem. 2015, 290, 10555–10567. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Cai, J.; Cai, J.; Wang, Z.; Cai, L. EZH2-miRNA Positive Feedback Promotes Tumor Growth in Ovarian Cancer. Front. Oncol. 2020, 10, 608393. [Google Scholar] [CrossRef] [PubMed]
- Enkhbaatar, Z.; Terashima, M.; Oktyabri, D.; Tange, S.; Ishimura, A.; Yano, S.; Suzuki, T. KDM5B histone demethylase controls epithelial-mesenchymal transition of cancer cells by regulating the expression of the microRNA-200 family. Cell Cycle 2013, 12, 2100–2112. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhao, H.; Zhang, Q.; Shi, Z.; Zhang, Y.; Zhao, L.; Ren, Y.; Ou, R.; Xu, Y. Human papillomavirus type 16 E7 oncoprotein-induced upregulation of lysine-specific demethylase 5A promotes cervical cancer progression by regulating the microRNA-424–5p/suppressor of zeste 12 pathway. Exp. Cell Res. 2020, 396, 112277. [Google Scholar] [CrossRef]
- Wang, X.; Lu, Z.; Gomez, A.; Hon, G.C.; Yue, Y.; Han, D.; Fu, Y.; Parisien, M.; Dai, Q.; Jia, G.; et al. N6-methyladenosine-dependent regulation of messenger RNA stability. Nature 2014, 505, 117–120. [Google Scholar] [CrossRef]
- Alarcón, C.R.; Lee, H.; Goodarzi, H.; Halberg, N.; Tavazoie, S.F. N6-methyladenosine marks primary microRNAs for processing. Nature 2015, 519, 482–485. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Hao, Y.-J.; Zhang, Y.; Li, M.-M.; Wang, M.; Han, W.; Wu, Y.; Lv, Y.; Hao, J.; Wang, L.; et al. m(6)A RNA Methylation Is Regulated by MicroRNAs and Promotes Reprogramming to Pluripotency. Cell Stem Cell 2015, 16, 289–301. [Google Scholar] [CrossRef] [Green Version]
- Ramassone, A.; Pagotto, S.; Veronese, A.; Visone, R. Epigenetics and MicroRNAs in Cancer. Int. J. Mol. Sci. 2018, 19, 459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majid, S.; Dar, A.A.; Saini, S.; Yamamura, S.; Hirata, H.; Tanaka, Y.; Deng, G.; Dahiya, R. MicroRNA-205-directed transcriptional activation of tumor suppressor genes in prostate cancer. Cancer 2010, 116, 5637–5649. [Google Scholar] [CrossRef]
- Zardo, G.; Ciolfi, A.; Vian, L.; Starnes, L.M.; Billi, M.; Racanicchi, S.; Maresca, C.; Fazi, F.; Travaglini, L.; Noguera, N.; et al. Polycombs and microRNA-223 regulate human granulopoiesis by transcriptional control of target gene expression. Blood 2012, 119, 4034–4046. [Google Scholar] [CrossRef] [Green Version]
- Esteller, M. Epigenetics in Cancer. N. Engl. J. Med. 2008, 358, 1148–1159. [Google Scholar] [CrossRef] [PubMed]
- Gama-Sosa, M.A.; Slagel, V.A.; Trewyn, R.W.; Oxenhandler, R.; Kuo, K.C.; Gehrke, C.W.; Ehrlich, M. The 5-methylcytosine content of DNA from human tumors. Nucleic Acids Res. 1983, 11, 6883–6894. [Google Scholar] [CrossRef]
- Feinberg, A.P.; Vogelstein, B. Hypomethylation distinguishes genes of some human cancers from their normal counterparts. Nature 1983, 301, 89–92. [Google Scholar] [CrossRef]
- Feinberg, A.P.; Vogelstein, B. Hypomethylation of ras oncogenes in primary human cancers. Biochem. Biophys. Res. Commun. 1983, 111, 47–54. [Google Scholar] [CrossRef]
- Ehrlich, M. DNA hypomethylation in cancer cells. Epigenomics 2009, 1, 239–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, J.; Vives, L.; Jordà, M.; Morales, C.; Muñoz, M.; Vendrell, E.; Peinado, M.A. Genome-wide tracking of unmethylated DNA Alu repeats in normal and cancer cells. Nucleic Acids Res. 2008, 36, 770–784. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.-J.; White-Cross, J.A.; Shen, L.; Issa, J.-P.J.; Rashid, A. Hypomethylation of long interspersed nuclear element-1 in hepatocellular carcinomas. Off. J. USA Can. Acad. Pathol. Inc. 2009, 22, 442–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauch, T.A.; Zhong, X.; Wu, X.; Wang, M.; Kernstine, K.H.; Wang, Z.; Riggs, A.D.; Pfeifer, G.P. High-resolution mapping of DNA hypermethylation and hypomethylation in lung cancer. Proc. Natl. Acad. Sci. USA 2008, 105, 252–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baylin, S.B.; Jones, P.A. A decade of exploring the cancer epigenome—Biological and translational implications. Nat. Rev. Cancer 2011, 11, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Kanwal, R.; Gupta, K.; Gupta, S. Cancer Epigenetics: An Introduction. In Methods in Molecular Biology; Verma, M., Ed.; Springer: New York, NY, USA, 2015; Volume 1238, pp. 3–25. ISBN 978-1-4939-1803-4. [Google Scholar]
- Esteller, M. Cancer epigenomics: DNA methylomes and histone-modification maps. Nat. Rev. Genet. 2007, 8, 286–298. [Google Scholar] [CrossRef]
- Fraga, M.F.; Ballestar, E.; Villar-Garea, A.; Boix-Chornet, M.; Espada, J.; Schotta, G.; Bonaldi, T.; Haydon, C.; Ropero, S.; Petrie, K.; et al. Loss of acetylation at Lys16 and trimethylation at Lys20 of histone H4 is a common hallmark of human cancer. Nat. Genet. 2005, 37, 391–400. [Google Scholar] [CrossRef]
- Nebbioso, A.; Tambaro, F.P.; Dell’Aversana, C.; Altucci, L. Cancer epigenetics: Moving forward. PLoS Genet. 2018, 14, e1007362. [Google Scholar] [CrossRef] [Green Version]
- Tam, W.L.; Weinberg, R.A. The epigenetics of epithelial-mesenchymal plasticity in cancer. Nat. Med. 2013, 19, 1438–1449. [Google Scholar] [CrossRef] [Green Version]
- Markouli, M.; Strepkos, D.; Basdra, E.K.; Papavassiliou, A.G.; Piperi, C. Prominent Role of Histone Modifications in the Regulation of Tumor Metastasis. Int. J. Mol. Sci. 2021, 22, 2778. [Google Scholar] [CrossRef]
- Di Cerbo, V.; Schneider, R. Cancers with wrong HATs: The impact of acetylation. Brief. Funct. Genom. 2013, 12, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Croce, C.M. Causes and consequences of microRNA dysregulation in cancer. Nat. Rev. Genet. 2009, 10, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, V.; Shah, J. Recent trends in targeting miRNAs for cancer therapy. J. Pharm. Pharmacol. 2020, 72, 1732–1749. [Google Scholar] [CrossRef]
- Calin, G.A.; Sevignani, C.; Dumitru, C.D.; Hyslop, T.; Noch, E.; Yendamuri, S.; Shimizu, M.; Rattan, S.; Bullrich, F.; Negrini, M.; et al. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc. Natl. Acad. Sci. USA 2004, 101, 2999–3004. [Google Scholar] [CrossRef] [Green Version]
- Jansson, M.D.; Lund, A.H. MicroRNA and cancer. Mol. Oncol. 2012, 6, 590–610. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Thomson, J.M.; Hemann, M.T.; Hernando-Monge, E.; Mu, D.; Goodson, S.; Powers, S.; Cordon-Cardo, C.; Lowe, S.W.; Hannon, G.J.; et al. A microRNA polycistron as a potential human oncogene. Nature 2005, 435, 828–833. [Google Scholar] [CrossRef] [PubMed]
- Woods, K.; Thomson, J.M.; Hammond, S.M. Direct Regulation of an Oncogenic Micro-RNA Cluster by E2F Transcription Factors. J. Biol. Chem. 2007, 282, 2130–2134. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Wang, C.; Wang, M.; Li, Z.; Casimiro, M.C.; Liu, M.; Wu, K.; Whittle, J.; Ju, X.; Hyslop, T.; et al. A cyclin D1/microRNA 17/20 regulatory feedback loop in control of breast cancer cell proliferation. J. Cell Biol. 2008, 182, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Lima, R.T.; Busacca, S.; Almeida, G.M.; Gaudino, G.; Fennell, D.A.; Vasconcelos, M.H. MicroRNA regulation of core apoptosis pathways in cancer. Eur. J. Cancer 2011, 47, 163–174. [Google Scholar] [CrossRef]
- Si, W.; Shen, J.; Zheng, H.; Fan, W. The role and mechanisms of action of microRNAs in cancer drug resistance. Clin. Epigenetics 2019, 11, 25. [Google Scholar] [CrossRef]
- Caravia, X.M.; López-Otín, C. Regulatory Roles of miRNAs in Aging. In microRNA: Basic Science; Santulli, G., Ed.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2015; Volume 887, pp. 213–230. ISBN 978-3-319-22379-7. [Google Scholar]
- Olivieri, F.; Rippo, M.R.; Monsurrò, V.; Salvioli, S.; Capri, M.; Procopio, A.D.; Franceschi, C. MicroRNAs linking inflamm-aging, cellular senescence and cancer. Ageing Res. Rev. 2013, 12, 1056–1068. [Google Scholar] [CrossRef]
- Hu, H.; Du, L.; Nagabayashi, G.; Seeger, R.C.; Gatti, R.A. ATM is down-regulated by N-Myc-regulated microRNA-421. Proc. Natl. Acad. Sci. USA 2010, 107, 1506–1511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Zhou, Y.; Sun, Q.; Zhou, J.; Pan, H.; Sui, X. Regulation of Autophagy by MiRNAs and Their Emerging Roles in Tumorigenesis and Cancer Treatment. Int. Rev. Cell Mol. Biol. 2017, 334, 1–26. [Google Scholar] [CrossRef]
- Fu, L.-L.; Wen, X.; Bao, J.-K.; Liu, B. MicroRNA-modulated autophagic signaling networks in cancer. Int. J. Biochem. Cell Biol. 2012, 44, 733–736. [Google Scholar] [CrossRef]
- Suzuki, H.; Maruyama, R.; Yamamoto, E.; Niinuma, T.; Kai, M. Relationship Between Noncoding RNA Dysregulation and Epigenetic Mechanisms in Cancer. Adv. Exp. Med. Biol. 2016, 927, 109–135. [Google Scholar] [CrossRef]
- Xia, W.; Chen, Q.; Wang, J.; Mao, Q.; Dong, G.; Shi, R.; Zheng, Y.; Xu, L.; Jiang, F. DNA methylation mediated silencing of microRNA-145 is a potential prognostic marker in patients with lung adenocarcinoma. Sci. Rep. 2015, 5, 16901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, W.; Gu, J.; Huang, M.; Wu, X.; Hildebrandt, M.A.T. Epigenetic analysis of microRNA genes in tumors from surgically resected lung cancer patients and association with survival. Mol. Carcinog. 2015, 54, E45–E51. [Google Scholar] [CrossRef] [Green Version]
- Daugaard, I.; Knudsen, A.; Kjeldsen, T.E.; Hager, H.; Hansen, L.L. The association between miR-34 dysregulation and distant metastases formation in lung adenocarcinoma. Exp. Mol. Pathol. 2017, 102, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Nadal, E.; Chen, G.; Gallegos, M.; Lin, L.; Ferrer-Torres, D.; Truini, A.; Wang, Z.; Lin, J.; Reddy, R.M.; Llatjos, R.; et al. Epigenetic Inactivation of microRNA-34b/c Predicts Poor Disease-Free Survival in Early-Stage Lung Adenocarcinoma. Clin. Cancer Res. 2013, 19, 6842–6852. [Google Scholar] [CrossRef] [Green Version]
- Brueckner, B.; Stresemann, C.; Kuner, R.; Mund, C.; Musch, T.; Meister, M.; Sültmann, H.; Lyko, F. The Human let-7a-3 Locus Contains an Epigenetically Regulated MicroRNA Gene with Oncogenic Function. Cancer Res. 2007, 67, 1419–1423. [Google Scholar] [CrossRef] [Green Version]
- Fabbri, M.; Garzon, R.; Cimmino, A.; Liu, Z.; Zanesi, N.; Callegari, E.; Liu, S.; Alder, H.; Costinean, S.; Fernandez-Cymering, C.; et al. MicroRNA-29 family reverts aberrant methylation in lung cancer by targeting DNA methyltransferases 3A and 3B. Proc. Natl. Acad. Sci. USA 2007, 104, 15805–15810. [Google Scholar] [CrossRef] [Green Version]
- Tan, M.; Wu, J.; Cai, Y. Suppression of Wnt signaling by the miR-29 family is mediated by demethylation of WIF-1 in non-small-cell lung cancer. Biochem. Biophys. Res. Commun. 2013, 438, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Incoronato, M.; Urso, L.; Portela, A.; Laukkanen, M.O.; Soini, Y.; Quintavalle, C.; Keller, S.; Esteller, M.; Condorelli, G. Epigenetic Regulation of miR-212 Expression in Lung Cancer. PLoS ONE 2011, 6, e27722. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Lv, D.; Li, M.; Zhang, X.; Sun, G.; Bai, Y.; Chang, D. Hypermethylation of miRNA-589 promoter leads to upregulation of HDAC5 which promotes malignancy in non-small cell lung cancer. Int. J. Oncol. 2017, 50, 2079–2090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, K.-W.; Hu, L.-Y.; Wu, C.-W.; Li, S.-C.; Lai, C.-H.; Kao, H.-W.; Fang, W.-L.; Lin, W.-C. Epigenetic regulation of miR-196b expression in gastric cancer. Genes Chromosom. Cancer 2010, 49, 969–980. [Google Scholar] [CrossRef]
- Li, N.; Wang, W.; Xu, B.; Gong, H. miR-196b regulates gastric cancer cell proliferation and invasion via PI3K/AKT/mTOR signaling pathway. Oncol. Lett. 2016, 11, 1745–1749. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Tang, H.; Liu, G.; Xiao, S.; Liang, D.; Ma, J.; Yang, Y.; Luo, H.; Zhu, Y.; Xie, F.; et al. MicroRNA-196b promotes gastric cancer progression by targeting ECRG4. Anticancer Drugs 2021, 32, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, Y.; Akiyama, Y.; Otsubo, T.; Shimada, S.; Yuasa, Y. Involvement of epigenetically silenced microRNA-181c in gastric carcinogenesis. Carcinogenesis 2010, 31, 777–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zabaglia, L.M.; Bartolomeu, N.C.; Dos Santos, M.P.; Peruquetti, R.L.; Chen, E.; de Arruda Cardoso Smith, M.; Payão, S.L.M.; Rasmussen, L.T. Decreased MicroRNA miR-181c Expression Associated with Gastric Cancer. J. Gastrointest. Cancer 2018, 49, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, N.; Asadi, M.; Shanehbandi, D.; Zafari, V.; Shomali, N.; Asvadi, T.; Sepehri, B. Evaluation of the Methylation of MIR129-2 Gene in Gastric Cancer. J. Gastrointest. Cancer 2020, 51, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.-Y.; Lin, K.-Y.; Tien, M.-T.; Wu, C.-T.; Uen, Y.-H.; Tseng, T.-L. Frequent DNA Methylation of MiR-129-2 and Its Potential Clinical Implication in Hepatocellular Carcinoma. Genes Chromosomes Cancer 2013, 52, 636–643. [Google Scholar] [CrossRef]
- Liu, Z.; Dou, C.; Yao, B.; Xu, M.; Ding, L.; Wang, Y.; Jia, Y.; Li, Q.; Zhang, H.; Tu, K.; et al. Methylation-Mediated Repression of MicroRNA-129-2 Suppresses Cell Aggressiveness by Inhibiting High Mobility Group Box 1 in Human Hepatocellular Carcinoma. Oncotarget 2016, 7, 36909–36923. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-W.; Liu, J.C.; Deatherage, D.E.; Luo, J.; Mutch, D.G.; Goodfellow, P.J.; Miller, D.S.; Huang, T.H.-M. Epigenetic Repression of MicroRNA-129-2 Leads to Overexpression of SOX4 Oncogene in Endometrial Cancer. Cancer Res. 2009, 69, 9038–9046. [Google Scholar] [CrossRef] [Green Version]
- Loginov, V.I.; Pronina, I.V.; Burdennyy, A.M.; Filippova, E.A.; Kazubskaya, T.P.; Kushlinsky, D.N.; Utkin, D.O.; Khodyrev, D.S.; Kushlinskii, N.E.; Dmitriev, A.A.; et al. Novel MiRNA Genes Deregulated by Aberrant Methylation in Ovarian Carcinoma Are Involved in Metastasis. Gene 2018, 662, 28–36. [Google Scholar] [CrossRef]
- Li, Y.; Nie, Y.; Tu, S.; Wang, H.; Zhou, Y.; Du, Y.; Cao, J.; Ye, M. Epigenetically Deregulated MiR-200c Is Involved in a Negative Feedback Loop with DNMT3a in Gastric Cancer Cells. Oncol. Rep. 2016, 36, 2108–2116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.; Liu, M.X.; Mak, C.S.-L.; Yung, M.M.-H.; Leung, T.H.-Y.; Xu, D.; Ngu, S.-F.; Chan, K.K.-L.; Yang, H.; Ngan, H.Y.-S.; et al. Methylation-Associated Silencing of MiR-193a-3p Promotes Ovarian Cancer Aggressiveness by Targeting GRB7 and MAPK/ERK Pathways. Theranostics 2018, 8, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Zhao, F.; Hui, L.; Li, X.; Zhang, D.; Lin, W.; Chen, Z.; Ning, Y. Suppressing MiR-199a-3p by Promoter Methylation Contributes to Tumor Aggressiveness and Cisplatin Resistance of Ovarian Cancer through Promoting DDR1 Expression. J. Ovarian Res. 2017, 10, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Zhen, S.; Ye, Z.; Lu, J.; Wang, L.; Li, P.; Li, J.; Zheng, X.; Li, H.; Chen, W.; et al. A Feedback Loop Between MiR-30a/c-5p and DNMT1 Mediates Cisplatin Resistance in Ovarian Cancer Cells. Cell. Physiol. Biochem. 2017, 41, 973–986. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Pei, M.; Li, Z.; Li, H.; Liu, Y.; Li, J. Double-Negative Feedback Interaction between DNA Methyltransferase 3A and MicroRNA-145 in the Warburg Effect of Ovarian Cancer Cells. Cancer Sci. 2018, 109, 2734–2745. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Tang, J.; Liu, X.; Zeng, L.; Cheng, C.; Luo, Y.; Li, L.; Qin, S.-L.; Sang, Y.; Deng, L.-M.; et al. Downregulation of MiR-129-2 by Promoter Hypermethylation Regulates Breast Cancer Cell Proliferation and Apoptosis. Oncol. Rep. 2016, 35, 2963–2969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben Gacem, R.; Ben Abdelkrim, O.; Ziadi, S.; Ben Dhiab, M.; Trimeche, M. Methylation of MiR-124a-1, MiR-124a-2, and MiR-124a-3 Genes Correlates with Aggressive and Advanced Breast Cancer Disease. Tumor Biol. J. Int. Soc. Oncodev. Biol. Med. 2014, 35, 4047–4056. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Li, S.; Cui, X.; Lv, X.; Jiao, Y.; Yu, F.; Yao, H.; Song, E.; Chen, Y.; Wang, M.; et al. The Overexpression of Hypomethylated MiR-663 Induces Chemotherapy Resistance in Human Breast Cancer Cells by Targeting Heparin Sulfate Proteoglycan 2 (HSPG2). J. Biol. Chem. 2013, 288, 10973–10985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Q.; Jiang, Y.; Yin, Y.; Li, Q.; He, J.; Jing, Y.; Qi, Y.-T.; Xu, Q.; Li, W.; Lu, B.; et al. A Regulatory Circuit of MiR-148a/152 and DNMT1 in Modulating Cell Transformation and Tumor Angiogenesis through IGF-IR and IRS1. J. Mol. Cell Biol. 2013, 5, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Bessa Garcia, S.A.; Araújo, M.; Pereira, T.; Mouta, J.; Freitas, R. HOX Genes Function in Breast Cancer Development. Biochim. Biophys. Acta Rev. Cancer 2020, 1873, 188358. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.; McDonnell, K.; Choi, H.; Gao, D.; Hahn, M.; Joshi, N.; Park, S.-M.; Catena, R.; Do, Y.; Brazin, J.; et al. Suppression of MiRNA-708 by Polycomb Group Promotes Metastases by Calcium-Induced Cell Migration. Cancer Cell 2013, 23, 63–76. [Google Scholar] [CrossRef] [Green Version]
- Tsai, K.-W.; Wu, C.-W.; Hu, L.-Y.; Li, S.-C.; Liao, Y.-L.; Lai, C.-H.; Kao, H.-W.; Fang, W.-L.; Huang, K.-H.; Chan, W.-C.; et al. Epigenetic Regulation of MiR-34b and MiR-129 Expression in Gastric Cancer. Int. J. Cancer 2011, 129, 2600–2610. [Google Scholar] [CrossRef]
- Toiyama, Y.; Okugawa, Y.; Tanaka, K.; Araki, T.; Uchida, K.; Hishida, A.; Uchino, M.; Ikeuchi, H.; Hirota, S.; Kusunoki, M.; et al. A Panel of methylated microrna biomarkers for identifying high-risk patients with ulcerative colitis-associated colorectal cancer. Gastroenterology 2017, 153, 1634–1646.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilting, S.M.; van Boerdonk, R.A.A.; Henken, F.E.; Meijer, C.J.L.M.; Diosdado, B.; Meijer, G.A.; le Sage, C.; Agami, R.; Snijders, P.J.F.; Steenbergen, R.D.M. Methylation-Mediated Silencing and Tumour Suppressive Function of Hsa-MiR-124 in Cervical Cancer. Mol. Cancer 2010, 9, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhoef, V.M.J.; Heideman, D.A.M.; van Kemenade, F.J.; Rozendaal, L.; Bosgraaf, R.P.; Hesselink, A.T.; Bekkers, R.L.M.; Massuger, L.F.A.G.; Steenbergen, R.D.M.; Snijders, P.J.F.; et al. Methylation Marker Analysis and HPV16/18 Genotyping in High-Risk HPV Positive Self-Sampled Specimens to Identify Women with High Grade CIN or Cervical Cancer. Gynecol. Oncol. 2014, 135, 58–63. [Google Scholar] [CrossRef] [PubMed]
- De Strooper, L.M.A.; Verhoef, V.M.J.; Berkhof, J.; Hesselink, A.T.; de Bruin, H.M.E.; van Kemenade, F.J.; Bosgraaf, R.P.; Bekkers, R.L.M.; Massuger, L.F.A.G.; Melchers, W.J.G.; et al. Validation of the FAM19A4/Mir124-2 DNA Methylation Test for Both Lavage- and Brush-Based Self-Samples to Detect Cervical (Pre)Cancer in HPV-Positive Women. Gynecol. Oncol. 2016, 141, 341–347. [Google Scholar] [CrossRef] [Green Version]
- Vink, F.J.; Dick, S.; Heideman, D.A.M.; De Strooper, L.M.A.; Steenbergen, R.D.M.; Lissenberg-Witte, B.L.W.; Floore, A.; Bonde, J.H.; Valenčak, A.O.; Poljak, M.; et al. Classification of High-Grade Cervical Intraepithelial Neoplasia by P16(Ink4a), Ki-67, HPV E4 and FAM19A4/MiR124-2 Methylation Status Demonstrates Considerable Heterogeneity with Potential Consequences for Management. Int. J. Cancer 2021. [Google Scholar] [CrossRef]
- Bonde, J.; Floore, A.; Ejegod, D.; Vink, F.J.; Hesselink, A.; van de Ven, P.M.; Valenčak, A.O.; Pedersen, H.; Doorn, S.; Quint, W.G.; et al. Methylation Markers FAM19A4 and MiR124-2 as Triage Strategy for Primary Human Papillomavirus Screen Positive Women: A Large European Multicenter Study. Int. J. Cancer 2021, 148, 396–405. [Google Scholar] [CrossRef]
- Torres-Ferreira, J.; Ramalho-Carvalho, J.; Gomez, A.; Menezes, F.D.; Freitas, R.; Oliveira, J.; Antunes, L.; Bento, M.J.; Esteller, M.; Henrique, R.; et al. MiR-193b Promoter Methylation Accurately Detects Prostate Cancer in Urine Sediments and MiR-34b/c or MiR-129-2 Promoter Methylation Define Subsets of Clinically Aggressive Tumors. Mol. Cancer 2017, 16, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Outeiro-Pinho, G.; Barros-Silva, D.; Aznar, E.; Sousa, A.-I.; Vieira-Coimbra, M.; Oliveira, J.; Gonçalves, C.S.; Costa, B.M.; Junker, K.; Henrique, R.; et al. MicroRNA-30a-5p(Me): A Novel Diagnostic and Prognostic Biomarker for Clear Cell Renal Cell Carcinoma in Tissue and Urine Samples. J. Exp. Clin. Cancer Res. 2020, 39, 98. [Google Scholar] [CrossRef] [PubMed]
- Padrão, N.A.; Monteiro-Reis, S.; Torres-Ferreira, J.; Antunes, L.; Leça, L.; Montezuma, D.; Ramalho-Carvalho, J.; Dias, P.C.; Monteiro, P.; Oliveira, J.; et al. MicroRNA Promoter Methylation: A New Tool for Accurate Detection of Urothelial Carcinoma. Br. J. Cancer 2017, 116, 634–639. [Google Scholar] [CrossRef] [Green Version]
- Konno, M.; Koseki, J.; Asai, A.; Yamagata, A.; Shimamura, T.; Motooka, D.; Okuzaki, D.; Kawamoto, K.; Mizushima, T.; Eguchi, H.; et al. Distinct Methylation Levels of Mature MicroRNAs in Gastrointestinal Cancers. Nat. Commun. 2019, 10, 3888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muraoka, T.; Soh, J.; Toyooka, S.; Aoe, K.; Fujimoto, N.; Hashida, S.; Maki, Y.; Tanaka, N.; Shien, K.; Furukawa, M.; et al. The Degree of MicroRNA-34b/c Methylation in Serum-Circulating DNA Is Associated with Malignant Pleural Mesothelioma. Lung Cancer 2013, 82, 485–490. [Google Scholar] [CrossRef] [Green Version]
- Sato, H.; Soh, J.; Aoe, K.; Fujimoto, N.; Tanaka, S.; Namba, K.; Torigoe, H.; Shien, K.; Yamamoto, H.; Tomida, S.; et al. Droplet Digital PCR as a Novel System for the Detection of MicroRNA-34b/c Methylation in Circulating DNA in Malignant Pleural Mesothelioma. Int. J. Oncol. 2019, 54, 2139–2148. [Google Scholar] [CrossRef]
- Kalimutho, M.; Di Cecilia, S.; Del Vecchio Blanco, G.; Roviello, F.; Sileri, P.; Cretella, M.; Formosa, A.; Corso, G.; Marrelli, D.; Pallone, F.; et al. Epigenetically Silenced MiR-34b/c as a Novel Faecal-Based Screening Marker for Colorectal Cancer. Br. J. Cancer 2011, 104, 1770–1778. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Song, Y.-C.; Cao, P.-L.; Zhang, H.; Guo, Q.; Yan, R.; Diao, D.-M.; Cheng, Y.; Dang, C.-X. Detection of MiR-34a and MiR-34b/c in Stool Sample as Potential Screening Biomarkers for Noninvasive Diagnosis of Colorectal Cancer. Med. Oncol. 2014, 31, 894. [Google Scholar] [CrossRef]
- Langevin, S.M.; Stone, R.A.; Bunker, C.H.; Grandis, J.R.; Sobol, R.W.; Taioli, E. MicroRNA-137 Promoter Methylation in Oral Rinses from Patients with Squamous Cell Carcinoma of the Head and Neck Is Associated with Gender and Body Mass Index. Carcinogenesis 2010, 31, 864–870. [Google Scholar] [CrossRef] [Green Version]
- Lujambio, A.; Calin, G.A.; Villanueva, A.; Ropero, S.; Sánchez-Céspedes, M.; Blanco, D.; Montuenga, L.M.; Rossi, S.; Nicoloso, M.S.; Faller, W.J.; et al. A MicroRNA DNA Methylation Signature for Human Cancer Metastasis. Proc. Natl. Acad. Sci. USA 2008, 105, 13556–13561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, K.; Emoto, N.; Hamano, E.; Sunohara, M.; Kawakami, M.; Kage, H.; Kitano, K.; Nakajima, J.; Goto, A.; Fukayama, M.; et al. Genome Structure-Based Screening Identified Epigenetically Silenced MicroRNA Associated with Invasiveness in Non-Small-Cell Lung Cancer. Int. J. Cancer 2012, 130, 2580–2590. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Min, L.; Zhang, X.; Hu, S.; Wang, B.; Liu, W.; Wang, R.; Gu, X.; Shen, W.; Lv, H.; et al. Decreased MiRNA-148a Is Associated with Lymph Node Metastasis and Poor Clinical Outcomes and Functions as a Suppressor of Tumor Metastasis in Non-Small Cell Lung Cancer. Oncol. Rep. 2013, 30, 1832–1840. [Google Scholar] [CrossRef] [Green Version]
- Siemens, H.; Neumann, J.; Jackstadt, R.; Mansmann, U.; Horst, D.; Kirchner, T.; Hermeking, H. Detection of MiR-34a Promoter Methylation in Combination with Elevated Expression of c-Met and β-Catenin Predicts Distant Metastasis of Colon Cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2013, 19, 710–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hildebrandt, M.A.T.; Gu, J.; Lin, J.; Ye, Y.; Tan, W.; Tamboli, P.; Wood, C.G.; Wu, X. Hsa-MiR-9 Methylation Status Is Associated with Cancer Development and Metastatic Recurrence in Patients with Clear Cell Renal Cell Carcinoma. Oncogene 2010, 29, 5724–5728. [Google Scholar] [CrossRef] [Green Version]
- Gebauer, K.; Peters, I.; Dubrowinskaja, N.; Hennenlotter, J.; Abbas, M.; Scherer, R.; Tezval, H.; Merseburger, A.S.; Stenzl, A.; Kuczyk, M.A.; et al. Hsa-Mir-124-3 CpG Island Methylation Is Associated with Advanced Tumours and Disease Recurrence of Patients with Clear Cell Renal Cell Carcinoma. Br. J. Cancer 2013, 108, 131–138. [Google Scholar] [CrossRef] [Green Version]
- Oltra, S.S.; Peña-Chilet, M.; Vidal-Tomas, V.; Flower, K.; Martinez, M.T.; Alonso, E.; Burgues, O.; Lluch, A.; Flanagan, J.M.; Ribas, G. Methylation Deregulation of MiRNA Promoters Identifies MiR124-2 as a Survival Biomarker in Breast Cancer in Very Young Women. Sci. Rep. 2018, 8, 14373. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, Z.; Gao, Y.; Li, N.; Li, B.; Tan, F.; Tan, X.; Lu, N.; Sun, Y.; Sun, J.; et al. DNA Hypermethylation of MicroRNA-34b/c Has Prognostic Value for Stage Ⅰ Non-Small Cell Lung Cancer. Cancer Biol. Ther. 2011, 11, 490–496. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.H.; Lee, W.K.; Lee, E.B.; Son, J.W.; Kim, D.S.; Park, J.Y. Combined Effect of Metastasis-Related MicroRNA, MiR-34 and MiR-124 Family, Methylation on Prognosis of Non-Small-Cell Lung Cancer. Clin. Lung Cancer 2017, 18, e13–e20. [Google Scholar] [CrossRef]
- Kitano, K.; Watanabe, K.; Emoto, N.; Kage, H.; Hamano, E.; Nagase, T.; Sano, A.; Murakawa, T.; Nakajima, J.; Goto, A.; et al. CpG Island Methylation of MicroRNAs Is Associated with Tumor Size and Recurrence of Non-Small-Cell Lung Cancer. Cancer Sci. 2011, 102, 2126–2131. [Google Scholar] [CrossRef]
- Langevin, S.M.; Stone, R.A.; Bunker, C.H.; Lyons-Weiler, M.A.; LaFramboise, W.A.; Kelly, L.; Seethala, R.R.; Grandis, J.R.; Sobol, R.W.; Taioli, E. MicroRNA-137 Promoter Methylation Is Associated with Poorer Overall Survival in Patients with Squamous Cell Carcinoma of the Head and Neck. Cancer 2011, 117, 1454–1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, M.; Cuatrecasas, M.; Balaguer, F.; Hur, K.; Toiyama, Y.; Castells, A.; Boland, C.R.; Goel, A. The Clinical Significance of MiR-148a as a Predictive Biomarker in Patients with Advanced Colorectal Cancer. PLoS ONE 2012, 7, e46684. [Google Scholar] [CrossRef]
- Agirre, X.; Vilas-Zornoza, A.; Jiménez-Velasco, A.; Martin-Subero, J.I.; Cordeu, L.; Gárate, L.; San José-Eneriz, E.; Abizanda, G.; Rodríguez-Otero, P.; Fortes, P.; et al. Epigenetic Silencing of the Tumor Suppressor MicroRNA Hsa-MiR-124a Regulates CDK6 Expression and Confers a Poor Prognosis in Acute Lymphoblastic Leukemia. Cancer Res. 2009, 69, 4443–4453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roman-Gomez, J.; Agirre, X.; Jiménez-Velasco, A.; Arqueros, V.; Vilas-Zornoza, A.; Rodriguez-Otero, P.; Martin-Subero, I.; Garate, L.; Cordeu, L.; San José-Eneriz, E.; et al. Epigenetic Regulation of MicroRNAs in Acute Lymphoblastic Leukemia. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2009, 27, 1316–1322. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.-Y.; Yim, R.L.-H.; Kwong, Y.-L.; Leung, C.-Y.; Hui, P.-K.; Cheung, F.; Liang, R.; Jin, D.-Y.; Chim, C.-S. Epigenetic Inactivation of the MIR129-2 in Hematological Malignancies. J. Hematol. Oncol. 2013, 6, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Wong, K.Y.; Calin, G.A.; Chng, W.-J.; Chan, G.C.-F.; Chim, C.S. Epigenetic Silencing of MiR-340-5p in Multiple Myeloma: Mechanisms and Prognostic Impact. Clin. Epigenetics 2019, 11, 71. [Google Scholar] [CrossRef]
- Qin, Y.; Zhang, S.; Deng, S.; An, G.; Qin, X.; Li, F.; Xu, Y.; Hao, M.; Yang, Y.; Zhou, W.; et al. Epigenetic Silencing of MiR-137 Induces Drug Resistance and Chromosomal Instability by Targeting AURKA in Multiple Myeloma. Leukemia 2017, 31, 1123–1135. [Google Scholar] [CrossRef]
- Toyota, M.; Suzuki, H.; Sasaki, Y.; Maruyama, R.; Imai, K.; Shinomura, Y.; Tokino, T. Epigenetic Silencing of MicroRNA-34b/c and B-Cell Translocation Gene 4 Is Associated with CpG Island Methylation in Colorectal Cancer. Cancer Res. 2008, 68, 4123–4132. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; He, C.; Wang, M.; Ma, X.; Mo, F.; Yang, S.; Han, J.; Wei, X. Targeting Epigenetic Regulators for Cancer Therapy: Mechanisms and Advances in Clinical Trials. Signal Transduct. Target. Ther. 2019, 4, 62. [Google Scholar] [CrossRef] [Green Version]
- Berg, J.L.; Perfler, B.; Hatzl, S.; Mayer, M.-C.; Wurm, S.; Uhl, B.; Reinisch, A.; Klymiuk, I.; Tierling, S.; Pregartner, G.; et al. Micro-RNA-125a Mediates the Effects of Hypomethylating Agents in Chronic Myelomonocytic Leukemia. Clin. Epigenetics 2021, 13, 1. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Type of Tumor | miRNAs Inactivation via DNA Hypermethylation | miRNAs Activation via DNA Hypomethylation | DNA Methylation Induced by miRNAs and Feedback Loop | miRNAs Dysregulation via Histone Modifications and Vice Versa | Targets/Pathways Affected by miRNAs Dysregulation with Potential Clinical Implications |
|---|---|---|---|---|---|
| Lung cancer | miR-145 [87]; miR-127, miR-9 [88]; miR-34b/c [89,90] | miR let-7a-3 [91] | miR-29 ⇒DNMT3A&DNMT3B [92]; miR-29a/b ⇔ DNMT3A & DNMT3B [93] | H3K9me2 & H3K27me3 ⇒ miR-212 [94]; miR 589 ⇒ HDAC5 [95] | miR-145 ⇒ c-Myc, AEG-1, EGFR, NUDT1 [87] |
| Gastric cancer | miR-181c [99] miR-129-2 [101,117] | miR-196b [96] | miR-200c ⇔ DNMT3A [106] | HDAC ⇒ miR-127 [33] | miR-196b ⇒PI3K/ AKT)/ mTOR pathway [97]; miR-196b ⇒ ECRG4 [98] |
| Ovarian cancer | mir-193a-3p [107], miR-199a-3p [108] | miR-191 [105] | miR-30a/c-5p ⇔ DNMT1 [109] | miR-145 ⇒ DNMT3A [110]; miR-137⇒ EZH2 [7]; miR-101-3p, let-7e-5p, miR-26a-5p, miR-98-5p, miR-141-3p ⇔ EZH2 [43] | miR-193a-3p ⇒ MAPK/ERK [107]; miR-199a-3p ⇒ DDR1 [108]; miR-30a/c-5p ⇒ SNAIL [109]; miR-145 ⇒ DNMT3A, HK2 [110] |
| Breast cancer | miR-129-2 [111]; miR-124a-1, miR-124a-2 & miR-124a-3 [112] | miR-663 [113] | miR-148a & miR-152 ⇔ DNMT1 [114] | H3K27me3 ⇒ miR-708 [116] | miR-129-2 ⇒ BCL2L2 [111]; miR-148a & miR-152 ⇒ IGF-1R, IRS1 [114] |
| Cancer Type | Type of Marker | miRNA | Source of miRNA | Sensibility | Specificity | Reference |
|---|---|---|---|---|---|---|
| CRC | Single | miR-1 | Tissue | 84.6% | 75.8% | [118] |
| miR-9 | 61.5% | 77.4% | ||||
| miR-124 | 76.9% | 67.7% | ||||
| miR-137 | 76.9% | 80.6% | ||||
| miR-34b/c | 100% | 56.5% | ||||
| Prostate carcinoma | Single | miR-193b | Urine | 91.6% | 95.7% | [124] |
| RCC | Single | miR30a-5p | Urine | 83% | 53% | [125] |
| Urothelial carcinomas | Signature | miR-663a | Urine | 87.7% | 84% | [126] |
| miR-129-2 | ||||||
| Pancreatic carcinoma | Single | miR-17-5p | Plasma | ND | ND | [127] |
| Pleural mesothelioma | Single | miR34b/c | Serum | 67% | 77% | [128,129] |
| 65.7% | 94.9% | |||||
| CRC | Single | miR-34b/c | Feces | ND | ND | [130,131] |
| 95% | 100% | |||||
| CRC | Single | miR-34a | Feces | 76.8% | 93.6% | [130,131] |
| HNSCC | Single | miR-137 | Oral rinses | 46.5% | 81.1% | [132] |
| Cancer Type | Type of Marker | miRNA | End Point | Univariate Analysis | Adjusted Analysis | Reference |
|---|---|---|---|---|---|---|
| ccRCC | Single | miR-9-1 | RFS | p = 0.034 | HR = 2.74 95% CI = 0.78–9.60 | [137,138] |
| Single | miR9-3 | RFS | p = 0.007 | HR = 5.85 95% CI = 1.30–26.35 | ||
| ccRCC | Single | miR-124-3 | RFS | p = 0.0005 | NA | [137,138] |
| Breast cancer | Single | miR-124-2 | OS | p = 0.0009 | HR = 3.23 p = 0.001 | [139] |
| NSCLC | Single | miR34b/c | RFS | p = 0.017 | HR = 2.60 95% CI = 1.34–5.06 p = 0.005 | [140] |
| OS | p = 0.010 | HR = 2.20 95% CI = 1.03–4.67 p = 0.027 | ||||
| NSCLC | Single | miR34b/c | RFS | p = 0.0003 | HR = 2.16 95% CI = 1.32–3.52 p = 0.002 | [90] |
| OS | p = 0.016 | HR = 1.79 95% CI = 1.07–3.02 p = 0.027 | ||||
| NSCLC | Signature | miR-34b/c | OS | p < 0.0001 | HR = 4.44 95% CI = 2.15–9.18 p < 0.0001 | [141] |
| miR-124-3 | ||||||
| NSCLC | Signature | miR-152 | RFS | p = 0.0177 | NA | [142] |
| miR-9-3 | ||||||
| miR-124-1 | ||||||
| miR-124-2 | ||||||
| miR-124-3 | ||||||
| NSCLC | Single | miR-127 | OS | p = 0.010 | HR = 1.97 95% CI = 1.15–3.40 p = 0.014 | [88] |
| HNSCC | Single | miR-137 | OS | p = 0.046 | HR = 3.68 95% CI = 1.01–13.38 p < 0.05 | [143] |
| CRC | Single | miR-148a | RFS | p = 0.020 | NA HR = 3.046 95% CI = 1.56–5.93 p = 0.0011 | [144] |
| OS | p = 0.0015 | |||||
| ALL | Single | miR-124a | RFS | p = 0.001 | p < 0.001 | [145] |
| OS | p < 0.001 | p = 0.005 | ||||
| ALL | Signature | miR-124a1 miR-124a2 miR-124a3 miR-34b/c miR-9-1 miR-9-2 miR-9-3 miR-10b miR-203 miR-196b miR-132/212 | RFS OS | p < 0.001 p < 0.001 | p < 0.001 p < 0.001 | [146] |
| Chronic lymphocytic leukemia | Single | miR-129-2 | OS | p = 0.004 | NA | [147] |
| Multiple myeloma | Single | miR-340 | OS | p < 0.001 | HR = 8.983 95% CI = 2.2–36.63 p = 0.002 | [148] |
| Multiple myeloma | Single | miR-137 | RFS | p = 0.043 | NA | [149] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pajares, M.J.; Alemany-Cosme, E.; Goñi, S.; Bandres, E.; Palanca-Ballester, C.; Sandoval, J. Epigenetic Regulation of microRNAs in Cancer: Shortening the Distance from Bench to Bedside. Int. J. Mol. Sci. 2021, 22, 7350. https://doi.org/10.3390/ijms22147350
Pajares MJ, Alemany-Cosme E, Goñi S, Bandres E, Palanca-Ballester C, Sandoval J. Epigenetic Regulation of microRNAs in Cancer: Shortening the Distance from Bench to Bedside. International Journal of Molecular Sciences. 2021; 22(14):7350. https://doi.org/10.3390/ijms22147350
Chicago/Turabian StylePajares, María J., Ester Alemany-Cosme, Saioa Goñi, Eva Bandres, Cora Palanca-Ballester, and Juan Sandoval. 2021. "Epigenetic Regulation of microRNAs in Cancer: Shortening the Distance from Bench to Bedside" International Journal of Molecular Sciences 22, no. 14: 7350. https://doi.org/10.3390/ijms22147350
APA StylePajares, M. J., Alemany-Cosme, E., Goñi, S., Bandres, E., Palanca-Ballester, C., & Sandoval, J. (2021). Epigenetic Regulation of microRNAs in Cancer: Shortening the Distance from Bench to Bedside. International Journal of Molecular Sciences, 22(14), 7350. https://doi.org/10.3390/ijms22147350