Chromosome Missegregation in Single Human Oocytes Is Related to the Age and Gene Expression Profile

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
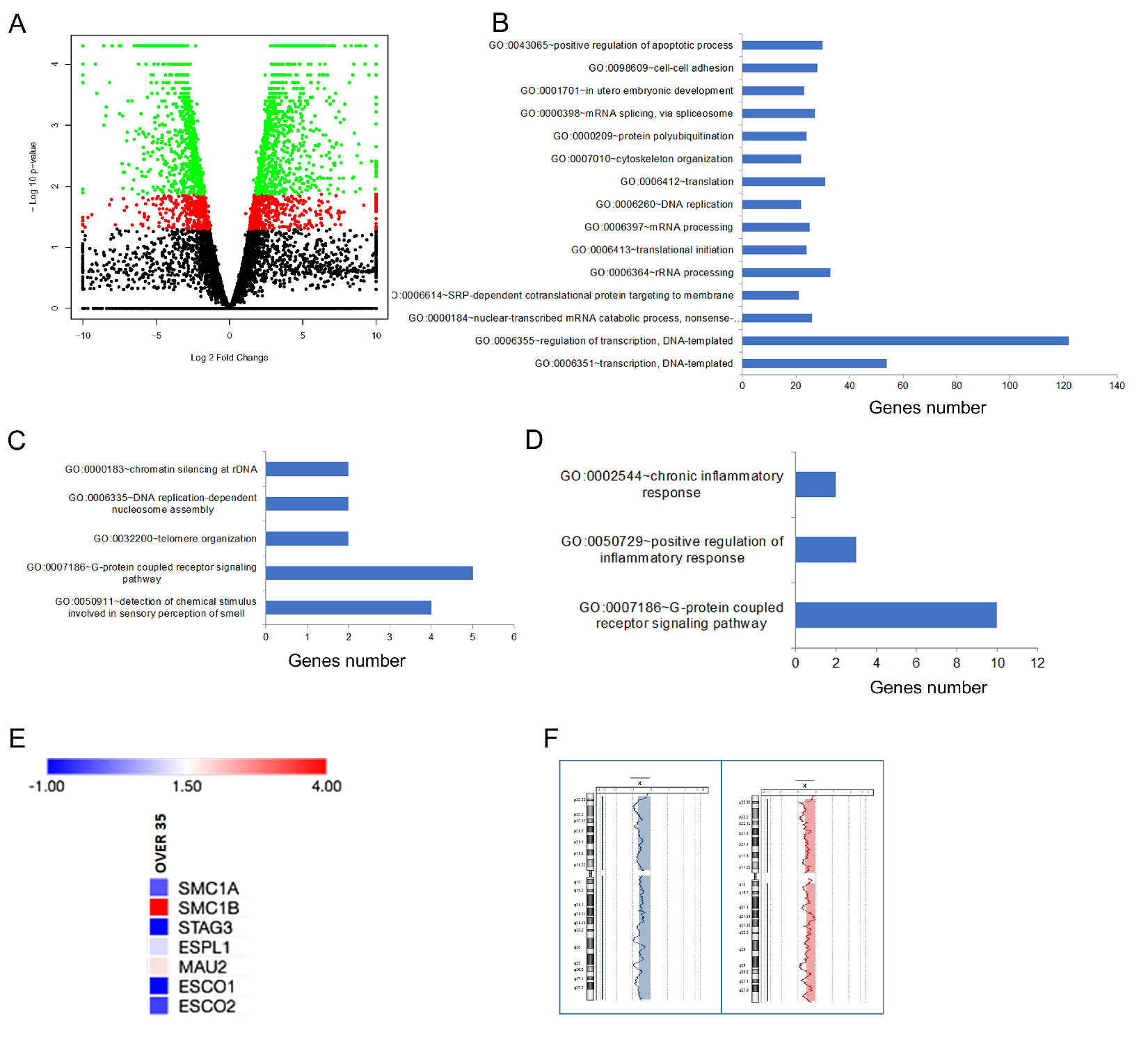
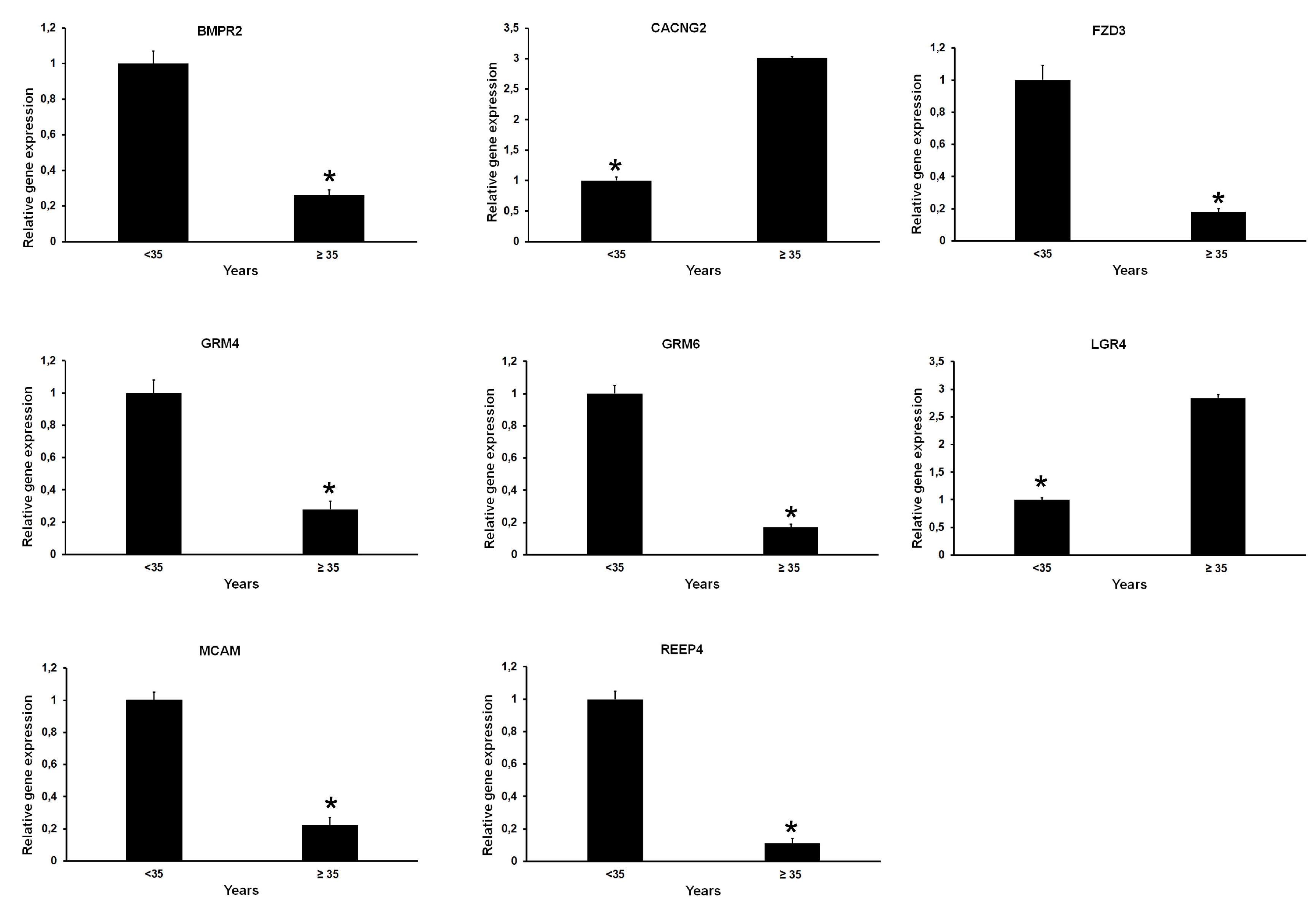
2.1. Differentially Expressed Genes in Relation to Age and Genome Integrity of Single Oocytes
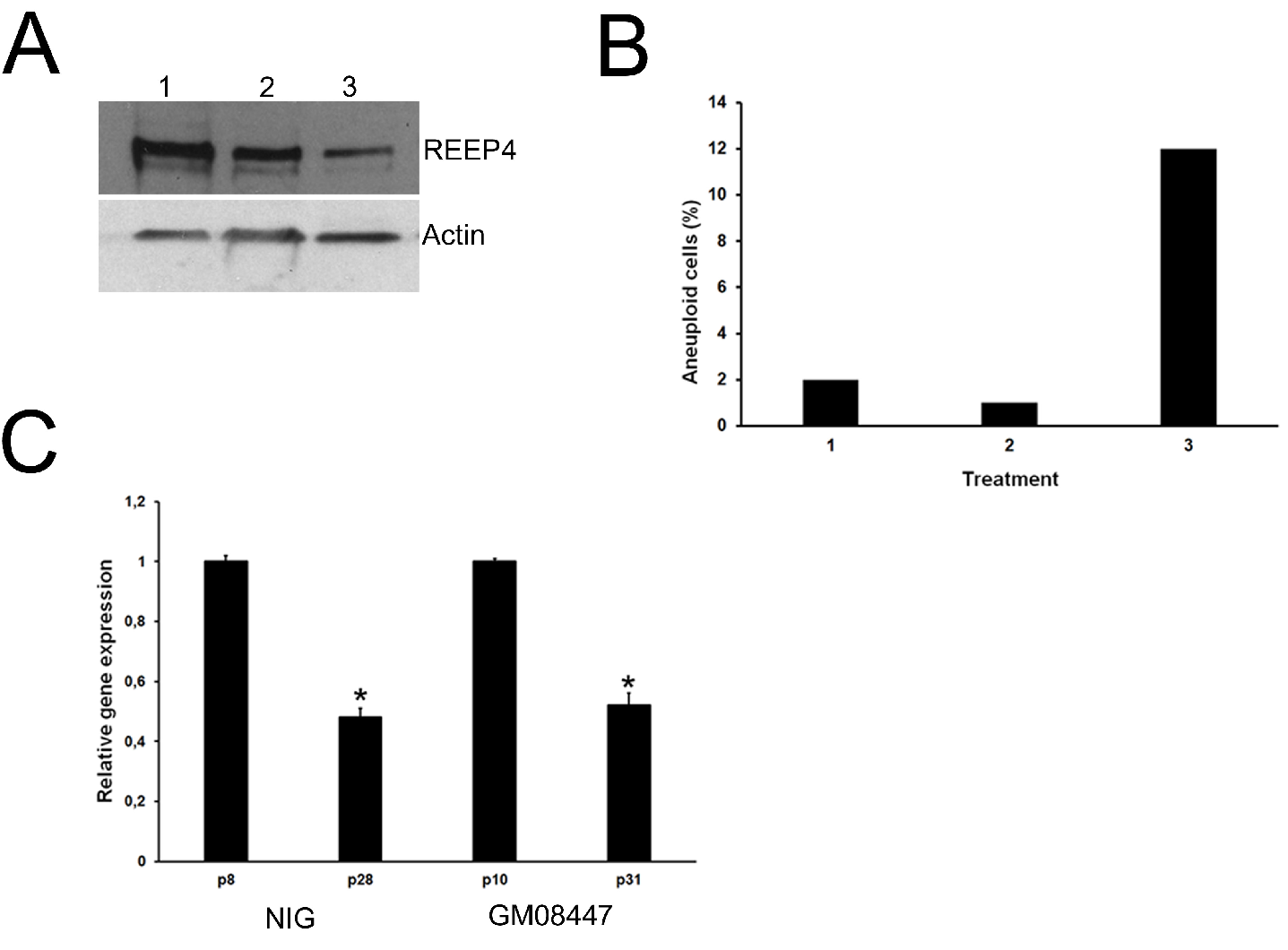
2.2. Interplay between Oocytes and Surrounding CCs
3. Discussion
4. Materials and Methods
4.1. Study Population and Ethical Approval
4.2. Cell Culture
4.3. siRNA Treatment and Cytogenetic Analysis
4.4. Western Blotting
4.5. Isolation of both DNA and RNA from Oocytes
4.6. RNA Sequencing
4.7. Pathway Analysis and Function
4.8. cDNA Synthesis and Quantitative Real-Time PCR (RT-qPCR)
4.9. Array Comparative Genomics Hybridization
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| aCGH | Array Comparative Genomics Hybridization |
| BMI | Body mass index |
| cAMP | cyclic Adenosine monophosphate |
| CCs | cumulus cells |
| COC | Cumulus oophorus cell complex |
| DAVID | Annotation, Visualization and Integrated Discovery |
| DEGs | Differentially expressed genes |
| ER | Endoplasmic reticulum |
| FDR | Fold discovery rate |
| GnRH | Gonadotrophin releasing hormone |
| HMG | Human menopausal gonadotrophin |
| IVF | In vitro fertilization |
| OSFs | Oocyte-secreted factors |
| rHCG | Recombinant human chorionic gonadotrophin |
| RPKM | Read per kilobase of exon per million fragments mapped |
| RT-qPCR | Quantitative real-time PCR |
| SMC | Structural Maintenance of Chromosomes |
References
- Braude, P.; Bolton, V.; Moore, S. Human gene expression first occurs between the four- and eight-cell stages of preimplantation development. Nature 1988, 332, 459–461. [Google Scholar] [CrossRef] [PubMed]
- Vassena, R.; Boue, S.; Gonzalez-Roca, E.; Aran, B.; Auer, H.; Veiga, A.; Izpisua Belmonte, J.C. Waves of early transcriptional activation and pluripotency program initiation during human preimplantation development. Development 2011, 138, 3699–3709. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; McKenzie, L.J.; Matzuk, M.M. Revisiting oocyte-somatic cell interactions: In search of novel intrafollicular predictors and regulators of oocyte developmental competence. Mol. Hum. Reprod. 2008, 14, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Robker, R.L.; Hennebold, J.D.; Russell, D.L. Co-ordination of ovulation and oocyte maturation: A good egg at the right time. Endocrinology 2018. [Google Scholar] [CrossRef]
- De La Fuente, R.; Eppig, J.J. Transcriptional activity of the mouse oocyte genome: Companion granulosa cells modulate transcription and chromatin remodeling. Dev. Biol. 2001, 229, 224–236. [Google Scholar] [CrossRef]
- Amireault, P.; Dube, F. Intracellular cAMP and calcium signaling by serotonin in mouse cumulus-oocyte complexes. Mol. Pharmacol. 2005, 68, 1678–1687. [Google Scholar] [CrossRef]
- Gilchrist, R.B.; Ritter, L.J.; Armstrong, D.T. Oocyte-somatic cell interactions during follicle development in mammals. Anim. Reprod. Sci. 2004, 82-83, 431–446. [Google Scholar] [CrossRef]
- Wigglesworth, K.; Lee, K.B.; O’Brien, M.J.; Peng, J.; Matzuk, M.M.; Eppig, J.J. Bidirectional communication between oocytes and ovarian follicular somatic cells is required for meiotic arrest of mammalian oocytes. Proc. Natl. Acad. Sci. USA 2013, 110, E3723–E3729. [Google Scholar] [CrossRef]
- Li, R.; Norman, R.J.; Armstrong, D.T.; Gilchrist, R.B. Oocyte-secreted factor(s) determine functional differences between bovine mural granulosa cells and cumulus cells. Biol. Reprod. 2000, 63, 839–845. [Google Scholar] [CrossRef]
- Capalbo, A.; Hoffmann, E.R.; Cimadomo, D.; Ubaldi, F.M.; Rienzi, L. Human female meiosis revised: New insights into the mechanisms of chromosome segregation and aneuploidies from advanced genomics and time-lapse imaging. Hum. Reprod. Update 2017, 23, 706–722. [Google Scholar] [CrossRef]
- Chiang, T.; Duncan, F.E.; Schindler, K.; Schultz, R.M.; Lampson, M.A. Evidence that weakened centromere cohesion is a leading cause of age-related aneuploidy in oocytes. Curr. Biol. 2010, 20, 1522–1528. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, S.I.; Hassold, T.J.; Hunt, P.A. Human aneuploidy: Mechanisms and new insights into an age-old problem. Nat. Rev. Genet. 2012, 13, 493–504. [Google Scholar] [CrossRef]
- Hassold, T.; Hunt, P. Maternal age and chromosomally abnormal pregnancies: What we know and what we wish we knew. Curr. Opin. Pediatr. 2009, 21, 703–708. [Google Scholar] [CrossRef]
- American College of Obstetricians and Gynecologists Committee on Gynecologic Practice and Practice Committee. Female age-related fertility decline. Committee Opinion No. 589. Fertil. Steril. 2014, 101, 633–634. [Google Scholar] [CrossRef] [PubMed]
- Hassold, T.; Hunt, P. To err (meiotically) is human: The genesis of human aneuploidy. Nat. Rev. Genet. 2001, 2, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Pellestor, F.; Andreo, B.; Arnal, F.; Humeau, C.; Demaille, J. Maternal aging and chromosomal abnormalities: New data drawn from in vitro unfertilized human oocytes. Hum. Genet 2003, 112, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Fragouli, E.; Escalona, A.; Gutierrez-Mateo, C.; Tormasi, S.; Alfarawati, S.; Sepulveda, S.; Noriega, L.; Garcia, J.; Wells, D.; Munne, S. Comparative genomic hybridization of oocytes and first polar bodies from young donors. Reprod. Biomed. Online 2009, 19, 228–237. [Google Scholar] [CrossRef]
- Fragouli, E.; Katz-Jaffe, M.; Alfarawati, S.; Stevens, J.; Colls, P.; Goodall, N.N.; Tormasi, S.; Gutierrez-Mateo, C.; Prates, R.; Schoolcraft, W.B.; et al. Comprehensive chromosome screening of polar bodies and blastocysts from couples experiencing repeated implantation failure. Fertil. Steril. 2010, 94, 875–887. [Google Scholar] [CrossRef]
- Hassold, T.; Hall, H.; Hunt, P. The origin of human aneuploidy: Where we have been, where we are going. Hum. Mol. Genet. 2007, 16, R203–R208. [Google Scholar] [CrossRef]
- Greaney, J.; Wei, Z.; Homer, H. Regulation of chromosome segregation in oocytes and the cellular basis for female meiotic errors. Hum. Reprod. Update 2017. [Google Scholar] [CrossRef]
- Tsutsumi, M.; Fujiwara, R.; Nishizawa, H.; Ito, M.; Kogo, H.; Inagaki, H.; Ohye, T.; Kato, T.; Fujii, T.; Kurahashi, H. Age-related decrease of meiotic cohesins in human oocytes. PLoS ONE 2014, 9, e96710. [Google Scholar] [CrossRef] [PubMed]
- Revenkova, E.; Herrmann, K.; Adelfalk, C.; Jessberger, R. Oocyte cohesin expression restricted to predictyate stages provides full fertility and prevents aneuploidy. Curr. Biol. 2010, 20, 1529–1533. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Ma, P.; Zhu, W.; Schultz, R.M. Age-associated increase in aneuploidy and changes in gene expression in mouse eggs. Dev. Biol. 2008, 316, 397–407. [Google Scholar] [CrossRef]
- Grondahl, M.L.; Yding Andersen, C.; Bogstad, J.; Nielsen, F.C.; Meinertz, H.; Borup, R. Gene expression profiles of single human mature oocytes in relation to age. Hum. Reprod. 2010, 25, 957–968. [Google Scholar] [CrossRef] [PubMed]
- Assidi, M.; Montag, M.; Sirard, M.A. Use of both cumulus cells’ transcriptomic markers and zona pellucida birefringence to select developmentally competent oocytes in human assisted reproductive technologies. BMC Genom. 2015, 16, S9. [Google Scholar] [CrossRef]
- Bunel, A.; Nivet, A.L.; Blondin, P.; Vigneault, C.; Richard, F.J.; Sirard, M.A. Cumulus cell gene expression associated with pre-ovulatory acquisition of developmental competence in bovine oocytes. Reprod. Fertil. Dev. 2014, 26, 855–865. [Google Scholar] [CrossRef] [PubMed]
- Dieci, C.; Lodde, V.; Labreque, R.; Dufort, I.; Tessaro, I.; Sirard, M.A.; Luciano, A.M. Differences in cumulus cell gene expression indicate the benefit of a pre-maturation step to improve in-vitro bovine embryo production. Mol. Hum. Reprod. 2016, 22, 882–897. [Google Scholar] [CrossRef][Green Version]
- Vigone, G.; Merico, V.; Prigione, A.; Mulas, F.; Sacchi, L.; Gabetta, M.; Bellazzi, R.; Redi, C.A.; Mazzini, G.; Adjaye, J.; et al. Transcriptome based identification of mouse cumulus cell markers that predict the developmental competence of their enclosed antral oocytes. BMC Genom. 2013, 14, 380. [Google Scholar] [CrossRef]
- Biase, F.H.; Kimble, K.M. Functional signaling and gene regulatory networks between the oocyte and the surrounding cumulus cells. BMC Genom. 2018, 19, 351. [Google Scholar] [CrossRef]
- O’Shea, L.C.; Mehta, J.; Lonergan, P.; Hensey, C.; Fair, T. Developmental competence in oocytes and cumulus cells: Candidate genes and networks. Syst. Biol. Reprod. Med. 2012, 58, 88–101. [Google Scholar] [CrossRef]
- Cecconi, S.; Colonna, R. Influence of granulosa cells and of different somatic cell types on mammalian oocyte development in vitro. Zygote 1996, 4, 305–307. [Google Scholar] [CrossRef]
- Kidder, G.M.; Vanderhyden, B.C. Bidirectional communication between oocytes and follicle cells: Ensuring oocyte developmental competence. Can. J. Physiol. Pharmacol. 2010, 88, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Labrecque, R.; Sirard, M.A. The study of mammalian oocyte competence by transcriptome analysis: Progress and challenges. Mol. Hum. Reprod. 2014, 20, 103–116. [Google Scholar] [CrossRef]
- Barragan, M.; Pons, J.; Ferrer-Vaquer, A.; Cornet-Bartolome, D.; Schweitzer, A.; Hubbard, J.; Auer, H.; Rodolosse, A.; Vassena, R. The transcriptome of human oocytes is related to age and ovarian reserve. Mol. Hum. Reprod. 2017, 23, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Lequarre, A.S.; Traverso, J.M.; Marchandise, J.; Donnay, I. Poly(A) RNA is reduced by half during bovine oocyte maturation but increases when meiotic arrest is maintained with CDK inhibitors. Biol. Reprod. 2004, 71, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Paynton, B.V.; Bachvarova, R. Polyadenylation and deadenylation of maternal mRNAs during oocyte growth and maturation in the mouse. Mol. Reprod. Dev. 1994, 37, 172–180. [Google Scholar] [CrossRef]
- Su, Y.Q.; Sugiura, K.; Woo, Y.; Wigglesworth, K.; Kamdar, S.; Affourtit, J.; Eppig, J.J. Selective degradation of transcripts during meiotic maturation of mouse oocytes. Dev. Biol. 2007, 302, 104–117. [Google Scholar] [CrossRef]
- Steuerwald, N.M.; Bermudez, M.G.; Wells, D.; Munne, S.; Cohen, J. Maternal age-related differential global expression profiles observed in human oocytes. Reprod. Biomed. Online 2007, 14, 700–708. [Google Scholar] [CrossRef]
- Hinckley, M.; Vaccari, S.; Horner, K.; Chen, R.; Conti, M. The G-protein-coupled receptors GPR3 and GPR12 are involved in cAMP signaling and maintenance of meiotic arrest in rodent oocytes. Dev. Biol. 2005, 287, 249–261. [Google Scholar] [CrossRef]
- Luan, X.; Liu, D.; Cao, Z.; Luo, L.; Liu, M.; Gao, M.; Zhang, X. Transcriptome profiling identifies differentially expressed genes in Huoyan goose ovaries between the laying period and ceased period. PLoS ONE 2014, 9, e113211. [Google Scholar] [CrossRef]
- DiLuigi, A.; Weitzman, V.N.; Pace, M.C.; Siano, L.J.; Maier, D.; Mehlmann, L.M. Meiotic arrest in human oocytes is maintained by a Gs signaling pathway. Biol. Reprod. 2008, 78, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Lang, S.; Wylie, C.; Hammes, S.R. The Xenopus laevis isoform of G protein-coupled receptor 3 (GPR3) is a constitutively active cell surface receptor that participates in maintaining meiotic arrest in X. laevis oocytes. Mol. Endocrinol. 2008, 22, 1853–1865. [Google Scholar] [CrossRef]
- Mehlmann, L.M.; Saeki, Y.; Tanaka, S.; Brennan, T.J.; Evsikov, A.V.; Pendola, F.L.; Knowles, B.B.; Eppig, J.J.; Jaffe, L.A. The Gs-linked receptor GPR3 maintains meiotic arrest in mammalian oocytes. Science 2004, 306, 1947–1950. [Google Scholar] [CrossRef] [PubMed]
- Ledent, C.; Demeestere, I.; Blum, D.; Petermans, J.; Hamalainen, T.; Smits, G.; Vassart, G. Premature ovarian aging in mice deficient for Gpr3. Proc. Natl. Acad. Sci. USA 2005, 102, 8922–8926. [Google Scholar] [CrossRef] [PubMed]
- Dumesic, D.A.; Meldrum, D.R.; Katz-Jaffe, M.G.; Krisher, R.L.; Schoolcraft, W.B. Oocyte environment: Follicular fluid and cumulus cells are critical for oocyte health. Fertil. Steril. 2015, 103, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, R.Q.; Ou, S.B.; Zhang, N.F.; Ren, L.; Wei, L.N.; Zhang, Q.X.; Yang, D.Z. Increased GDF9 and BMP15 mRNA levels in cumulus granulosa cells correlate with oocyte maturation, fertilization, and embryo quality in humans. Reprod. Biol. Endocrinol. 2014, 12, 81. [Google Scholar] [CrossRef]
- Hsieh, M.; Johnson, M.A.; Greenberg, N.M.; Richards, J.S. Regulated expression of Wnts and Frizzleds at specific stages of follicular development in the rodent ovary. Endocrinology 2002, 143, 898–908. [Google Scholar] [CrossRef]
- Kimura, T.; Nakamura, T.; Murayama, K.; Umehara, H.; Yamano, N.; Watanabe, S.; Taketo, M.M.; Nakano, T. The stabilization of beta-catenin leads to impaired primordial germ cell development via aberrant cell cycle progression. Dev. Biol. 2006, 300, 545–553. [Google Scholar] [CrossRef]
- Wang, S.B.; Liu, Y.P.; Zhang, Z.H.; Wang, Z.K.; Xu, Y.X.; Wang, Z.C. Temporal and spatial expression profiles of Frizzled 3 in the ovary during the estrous cycle. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef]
- Cheng, Y.; Kawamura, K.; Takae, S.; Deguchi, M.; Yang, Q.; Kuo, C.; Hsueh, A.J. Oocyte-derived R-spondin2 promotes ovarian follicle development. FASEB J. 2013, 27, 2175–2184. [Google Scholar] [CrossRef]
- Revenkova, E.; Eijpe, M.; Heyting, C.; Hodges, C.A.; Hunt, P.A.; Liebe, B.; Scherthan, H.; Jessberger, R. Cohesin SMC1 beta is required for meiotic chromosome dynamics, sister chromatid cohesion and DNA recombination. Nat. Cell Biol. 2004, 6, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Okada, K.; Ogushi, S.; Miyano, T.; Miyake, M.; Yamashita, M. Loss of Rec8 from chromosome arm and centromere region is required for homologous chromosome separation and sister chromatid separation, respectively, in mammalian meiosis. Cell Cycle 2006, 5, 1448–1455. [Google Scholar] [CrossRef]
- Wood, A.J.; Severson, A.F.; Meyer, B.J. Condensin and cohesin complexity: The expanding repertoire of functions. Nat. Rev. Genet. 2010, 11, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Schlaitz, A.L.; Thompson, J.; Wong, C.C.; Yates, J.R., 3rd; Heald, R. REEP3/4 ensure endoplasmic reticulum clearance from metaphase chromatin and proper nuclear envelope architecture. Dev. Cell 2013, 26, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Obradors, A.; Rius, M.; Daina, G.; Ramos, L.; Benet, J.; Navarro, J. Whole-chromosome aneuploidy analysis in human oocytes: Focus on comparative genomic hybridization. Cytogenet. Genome Res. 2011, 133, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Hardy, K.; Hardy, P.J.; Jacobs, P.A.; Lewallen, K.; Hassold, T.J. Temporal changes in chromosome abnormalities in human spontaneous abortions: Results of 40 years of analysis. Am. J. Med. Genet. A 2016, 170, 2671–2680. [Google Scholar] [CrossRef]
- Hassold, T.; Abruzzo, M.; Adkins, K.; Griffin, D.; Merrill, M.; Millie, E.; Saker, D.; Shen, J.; Zaragoza, M. Human aneuploidy: Incidence, origin, and etiology. Environ. Mol. Mutagen. 1996, 28, 167–175. [Google Scholar] [CrossRef]
- Rienzi, L.; Balaban, B.; Ebner, T.; Mandelbaum, J. The oocyte. Hum. Reprod. 2012, 27, i2–i21. [Google Scholar] [CrossRef]
- Musio, A.; Montagna, C.; Zambroni, D.; Indino, E.; Barbieri, O.; Citti, L.; Villa, A.; Ried, T.; Vezzoni, P. Inhibition of BUB1 results in genomic instability and anchorage-independent growth of normal human fibroblasts. Cancer Res. 2003, 63, 2855–2863. [Google Scholar]
- Mannini, L.; Cucco, F.; Quarantotti, V.; Amato, C.; Tinti, M.; Tana, L.; Frattini, A.; Delia, D.; Krantz, I.D.; Jessberger, R.; et al. SMC1B is present in mammalian somatic cells and interacts with mitotic cohesin proteins. Sci. Rep. 2015, 5, 18472. [Google Scholar] [CrossRef]
- Cucco, F.; Servadio, A.; Gatti, V.; Bianchi, P.; Mannini, L.; Prodosmo, A.; De Vitis, E.; Basso, G.; Friuli, A.; Laghi, L.; et al. Mutant cohesin drives chromosomal instability in early colorectal adenomas. Hum. Mol. Genet. 2014, 23, 6. [Google Scholar] [CrossRef] [PubMed]
- Cucco, F.; Palumbo, E.; Camerini, S.; D’Alessio, B.; Quarantotti, V.; Casella, M.L.; Rizzo, I.M.; Cukrov, D.; Delia, D.; Russo, A.; et al. Separase prevents genomic instability by controlling replication fork speed. Nucleic Acids Res. 2018, 46, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Mannini, L.; Lamaze, F.C.; Cucco, F.; Amato, C.; Quarantotti, V.; Rizzo, I.M.; Krantz, I.D.; Bilodeau, S.; Musio, A. Mutant cohesin affects RNA polymerase II regulation in Cornelia de Lange syndrome. Sci. Rep. 2015, 5, 16803. [Google Scholar] [CrossRef]
- Cukrov, D.; Newman, T.A.C.; Leask, M.; Leeke, B.; Sarogni, P.; Patimo, A.; Kline, A.D.; Krantz, I.D.; Horsfield, J.A.; Musio, A. Antioxidant treatment ameliorates phenotypic features of SMC1A-mutated Cornelia de Lange syndrome in vitro and in vivo. Hum. Mol. Genet. 2018, 27, 3002–3011. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef]
- Trapnell, C.; Salzberg, S.L. How to map billions of short reads onto genomes. Nat. Biotechnol. 2009, 27, 455–457. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate - a Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. B Met. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Valli, R.; Marletta, C.; Pressato, B.; Montalbano, G.; Lo Curto, F.; Pasquali, F.; Maserati, E. Comparative genomic hybridization on microarray (a-CGH) in constitutional and acquired mosaicism may detect as low as 8% abnormal cells. Mol. Cytogenet. 2011, 4, 13. [Google Scholar] [CrossRef]
- Valli, R.; Maserati, E.; Marletta, C.; Pressato, B.; Lo Curto, F.; Pasquali, F. Evaluating chromosomal mosaicism by array comparative genomic hybridization in hematological malignancies: The proposal of a formula. Cancer Genet. 2011, 204, 216–218. [Google Scholar] [CrossRef]
- Valli, R.; De Bernardi, F.; Frattini, A.; Volpi, L.; Bignami, M.; Facchetti, F.; Pasquali, F.; Castelnuovo, P.; Maserati, E. Comparative genomic hybridization on microarray (a-CGH) in olfactory neuroblastoma: Analysis of ten cases and review of the literature. Genes Chromosomes Cancer 2015, 54, 771–775. [Google Scholar] [CrossRef] [PubMed]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barone, S.; Sarogni, P.; Valli, R.; Pallotta, M.M.; Silvia, G.; Frattini, A.; Khan, A.W.; Rapalini, E.; Parri, C.; Musio, A. Chromosome Missegregation in Single Human Oocytes Is Related to the Age and Gene Expression Profile. Int. J. Mol. Sci. 2020, 21, 1934. https://doi.org/10.3390/ijms21061934
Barone S, Sarogni P, Valli R, Pallotta MM, Silvia G, Frattini A, Khan AW, Rapalini E, Parri C, Musio A. Chromosome Missegregation in Single Human Oocytes Is Related to the Age and Gene Expression Profile. International Journal of Molecular Sciences. 2020; 21(6):1934. https://doi.org/10.3390/ijms21061934
Chicago/Turabian StyleBarone, Stefano, Patrizia Sarogni, Roberto Valli, Maria Michela Pallotta, Gazzi Silvia, Annalisa Frattini, Abdul Waheed Khan, Erika Rapalini, Cristiana Parri, and Antonio Musio. 2020. "Chromosome Missegregation in Single Human Oocytes Is Related to the Age and Gene Expression Profile" International Journal of Molecular Sciences 21, no. 6: 1934. https://doi.org/10.3390/ijms21061934
APA StyleBarone, S., Sarogni, P., Valli, R., Pallotta, M. M., Silvia, G., Frattini, A., Khan, A. W., Rapalini, E., Parri, C., & Musio, A. (2020). Chromosome Missegregation in Single Human Oocytes Is Related to the Age and Gene Expression Profile. International Journal of Molecular Sciences, 21(6), 1934. https://doi.org/10.3390/ijms21061934