Localization of Annexin A6 in Matrix Vesicles During Physiological Mineralization

 ,
, 
 ,
,  ,
,
Abstract

1. Introduction
2. Results and Discussion
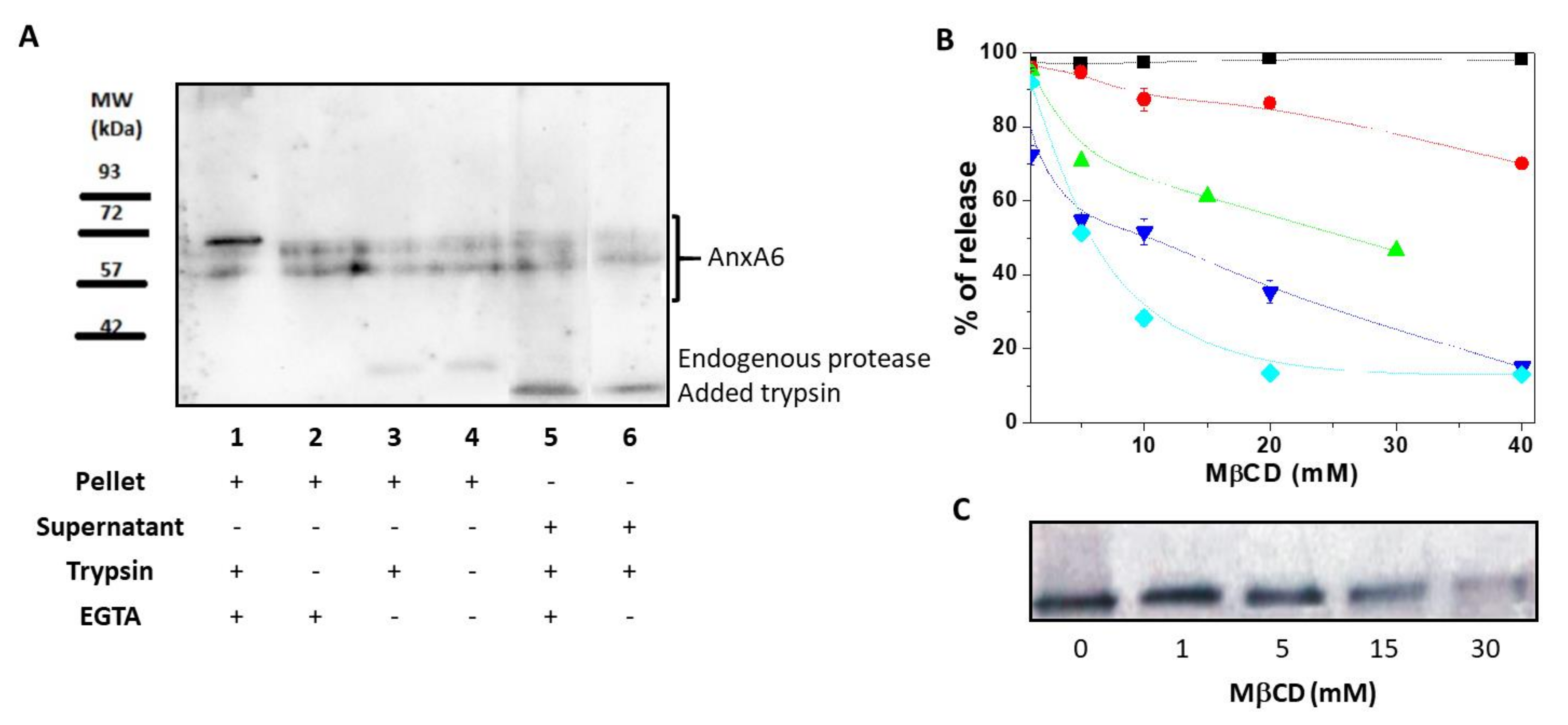
2.1. AnxA6 Associates With the MV Membrane as Both an Integral and a Peripheric Protein
2.2. AnxA6 Co-Localizes in Lipid Domains Enriched in Cholesterol
2.3. Interaction of AnxA6 With Lipid Monolayers
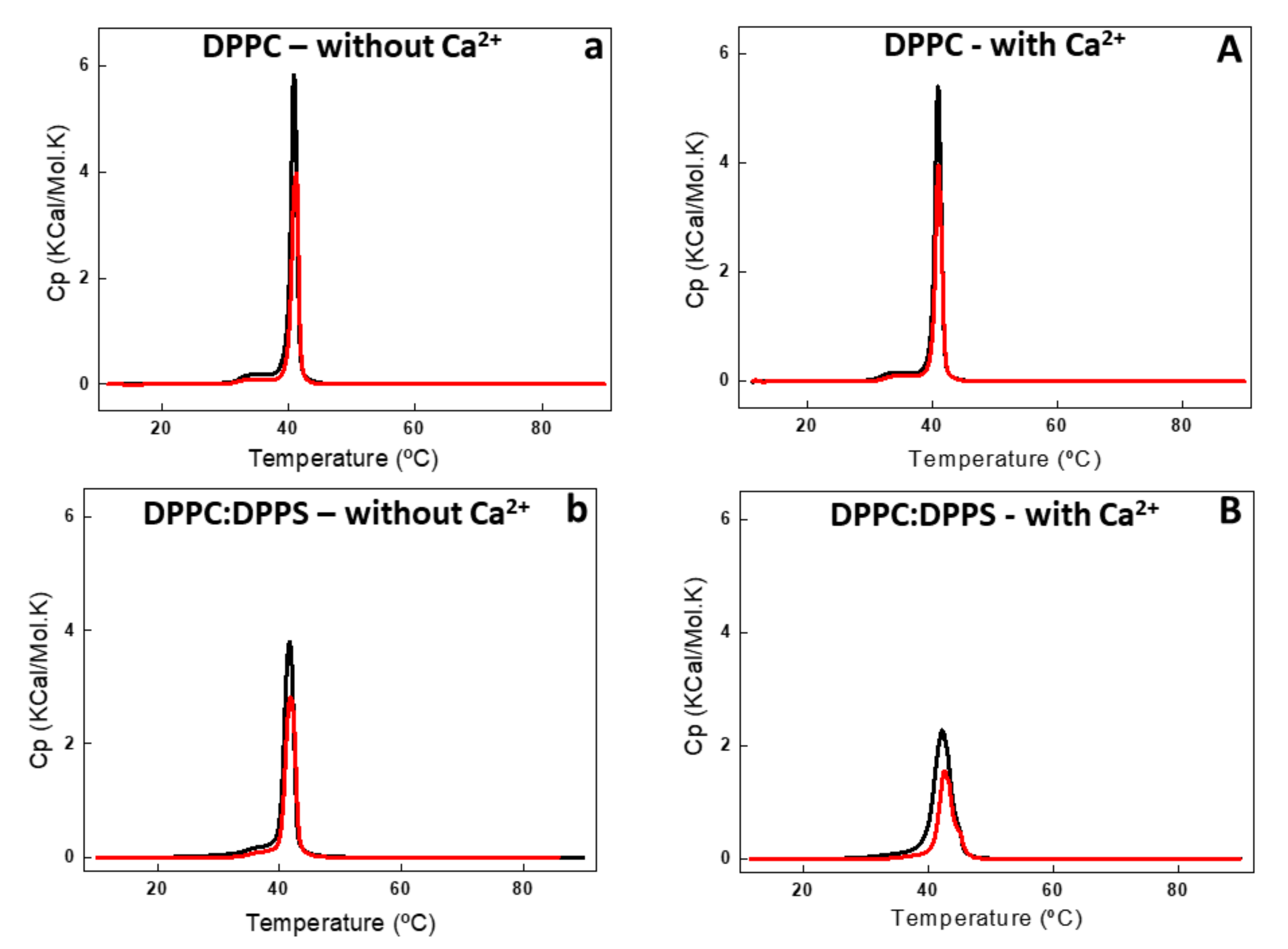
2.4. Interaction of AnxA6 With Lipid Bilayers
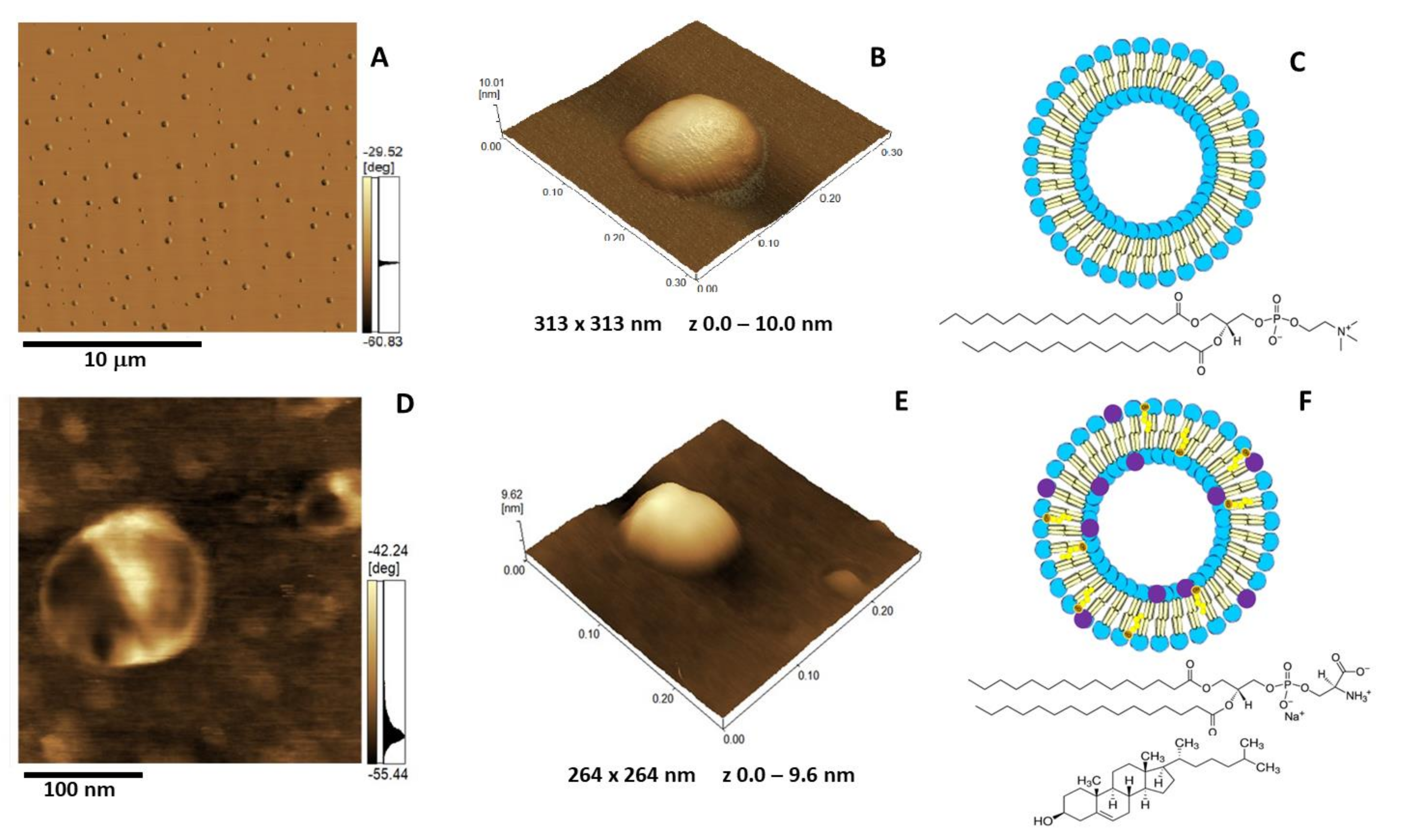
2.5. Atomic Force Microscopy Analysis
3. Materials and Methods
3.1. Materials
3.2. Expression of AnxA6
3.3. Isolation of Matrix Vesicles
3.4. Enzyme Activities, Total Protein, and Cholesterol Content
3.5. Treatment of Matrix Vesicles With Trypsin and Freeze-Thaw Cycles in the Absence and Presence of Calcium
3.6. Western Blotting
3.7. Treatment of Matrix Vesicles with Methyl-β-Cyclodextrin
3.8. Preparation of Liposomes
3.9. Preparation of Proteoliposomes
3.10. Dynamic Light Scattering (DLS)
3.11. Differential Scanning Calorimetry (DSC)
3.12. Exclusion Pressure (πexc)
3.13. Atomic Force Microscope (AFM) Imaging
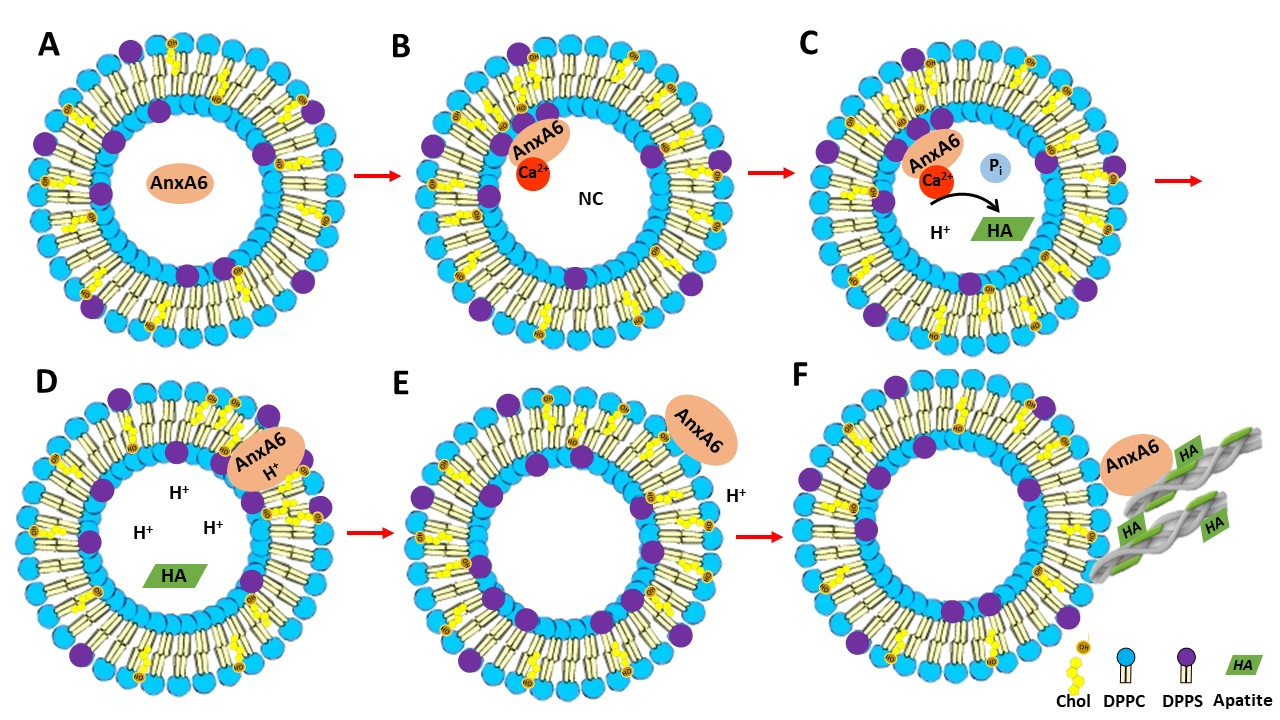
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Aghajanian, P.; Mohan, S. The art of building bone: Emerging role of chondrocyte-to-osteoblast transdifferentiation in endochondral ossification. Bone Res. 2018, 6, 19. [Google Scholar] [CrossRef]
- Wu, L.N.; Genge, B.R.; Lloyd, G.C.; Wuthier, R.E. Collagen-binding proteins in collagenase-released matrix vesicles from cartilage. Interaction between matrix vesicle proteins and different types of collagen. J. Biol. Chem. 1991, 266, 1195–1203. [Google Scholar] [PubMed]
- Kirsch, T.; Wuthier, R.E. Stimulation of calcification of growth plate cartilage matrix vesicles by binding to type II and X collagens. J. Biol. Chem. 1994, 269, 11462–11469. [Google Scholar] [PubMed]
- Anderson, H.C. Matrix vesicles and calcification. Role of extracellular membrane vesicles in the pathogenesis of various diseases, including cancer, renal diseases, atherosclerosis, and arthritis. Curr. Rheumatol. Rep. 2003, 5, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Anderson, H.C.; Mulhall, D.; Garimella, R. Role of extracellular membrane vesicles in the pathogenesis of various diseases, including cancer, renal diseases, atherosclerosis, and arthritis. Lab. Invest. 2010, 90, 1549–1557. [Google Scholar] [CrossRef]
- Golub, E.E. Role of matrix vesicles in biomineralization. Biochim. Biophys. Acta 2009, 1790, 1592–1598. [Google Scholar] [CrossRef]
- Wuthier, R.E.; Lipscomb, G.F. Matrix vesicles: Structure, composition, formation and function in calcification. Front. Biosci. 2011, 17, 2812–2902. [Google Scholar] [CrossRef]
- Bottini, M.; Mebarek, S.; Anderson, K.L.; Strzelecka-Kiliszek, A.; Bozycki, L.; Simão, A.M.S.; Bolean, M.; Ciancaglini, P.; Pikula, J.B.; Pikula, S.; et al. Matrix vesicles from chondrocytes and osteoblasts. Their biogenesis, properties, functions and biomimetic models. BBA Gen. Subj. 2018, 1862, 532–546. [Google Scholar]
- Balcerzak, M.; Malinowska, A.; Thouverey, C.; Sekrecka, A.; Dadlez, M.; Buchet, R.; Pikula, S. Proteome analysis of matrix vesicles isolated from femurs of chicken embryo. Proteomics 2008, 8, 192–205. [Google Scholar] [CrossRef]
- Thouverey, C.; Malinowska, A.; Balcerzak, M.; Strzelecka-Kiliszek, A.; Buchet, R.; Dadlez, M.; Pikula, S. Proteomic characterization of biogenesis and functions of matrix vesicles released from mineralizing human osteoblast-like cells. J. Proteom. 2011, 74, 1123–1134. [Google Scholar] [CrossRef]
- Ayala–Sanmartin, J. Cholesterol enhances phospholipid binding and aggregation of annexins by their core domain. Biochem. Biophys. Res. Commun. 2001, 283, 72–79. [Google Scholar] [CrossRef]
- Babiychuk, E.B.; Draeger, A. Annexins in cell membrane dynamics. Ca(2+)-regulated association of lipid microdomains. J. Cell. Biol. 2000, 150, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Golczak, M.; Kicinska, A.; Bandorowicz–Pikula, J.; Buchet, R.; Szewczyk, A.; Pikula, S. Acidic pH-induced folding of annexin VI is a prerequisite for its insertion into lipid bilayers and formation of ion channels by the protein molecules. Faseb J. 2001, 15, 1083–1085. [Google Scholar] [CrossRef]
- Strzelecka-Kiliszek, A.; Buszewska, M.E.; Podszywalow-Bartnicka, P.; Pikula, S.; Otulak, K.; Buchet, R.; Bandorowicz-Pikula, J. Calcium- and pH-dependent localization of annexin A6 isoforms in Balb/3T3 fibroblasts reflecting their potential participation in vesicular transport. J. Cell. Bioch. 2008, 104, 418–434. [Google Scholar] [CrossRef]
- Genge, B.R.; Cao, X.; Wu, L.N.; Buzzi, W.R.; Showman, R.W.; Arsenault, A.L.; Ishikawa, Y.; Wuthier, R.E. Establishment of the primary structure of the major lipid-dependent Ca2+ binding proteins of chicken growth plate cartilage matrix vesicles: Identity with anchorin CII (annexin V) and annexin II. J. Bone Min.. Res. 1992, 7, 807–819. [Google Scholar] [CrossRef]
- Von der Mark, K.; Mollenhauer, J. Annexin V interactions with collagen. Cell Mol. Life Sci. 1997, 53, 539–545. [Google Scholar] [CrossRef]
- Kirsch, T.; Harrison, G.; Golub, E.E.; Nah, H.D. The roles of annexins and types II and X collagen in matrix vesicle-mediated mineralization of growth plate cartilage. J. Biol. Chem. 2000, 275, 35577–35583. [Google Scholar] [CrossRef]
- Chen, N.X.; O’Neill, K.D.; Chen, X.; Moe, S.M. Annexin-mediated matrix vesicle calcification in vascular smooth muscle cells. J. Bone Min. Res. 2008, 23, 1798–1805. [Google Scholar] [CrossRef]
- Huber, R.; Romisch, J.; Pâques, E.P. The crystal and molecular structure of human annexin V, an anticoagulant protein that binds to calcium and membranes. Embo J. 1990, 9, 3867–3874. [Google Scholar] [CrossRef] [PubMed]
- Boye, T.L.; Jeppesen, J.C.; Maeda, K.; Pezesshkian, W.; Solovyeva, V.; Nylandsted, J.; Simonsen, A.C. Annexins induce curvature on free-edge membranes displaying distinct morphologies. Sci. Rep. 2018, 8, 10309–10322. [Google Scholar] [CrossRef] [PubMed]
- R.R.C. New. Liposomes: A practical approach; Oxford University Press: Oxford, UK, 1990; p. 58. [Google Scholar]
- Wu, L.N.; Yoshimori, T.; Genge, B.R.; Sauer, G.R.; Kirsch, T.; Ishikawa, Y.; Wuthier, R.E. Characterization of the nucleational core complex responsible for mineral induction by growth plate cartilage matrix vesicles. J. Biol. Chem. 1997, 268, 25084–25094. [Google Scholar]
- Mookhtiar, K.A.; Van Wart, H.E. Clostridium histolyticum collagenases: A new look at some old enzymes. Matrix Suppl. 1992, 1, 116–126. [Google Scholar]
- Chen, D.; Golub, E.E. Search for an endogenous collagenase in chicken endochondral bone matrix vesicles. Penn. Dent. J. Phila. 2001, 101, 12–33. [Google Scholar] [PubMed]
- Wuthier, R.E. Lipid composition of isolated epiphyseal cartilage cells, membranes and matrix vesicles. Biochim. Biophys. Acta 1975, 409, 128–143. [Google Scholar] [CrossRef]
- Abdallah, D.; Hamade, E.; Merhi, R.A.; Bassam, B.; Buchet, R.; Mebarek, S. Fatty acid composition in matrix vesicles and in microvilli from femurs of chicken embryos revealed selective recruitment of fatty acids. Fatty acid composition in matrix vesicles and in microvilli from femurs of chicken embryos revealed selective recruitment of fatty acids. Biochem. Biophys. Res. Commun. 2014, 446, 1161–1164. [Google Scholar] [PubMed]
- Kyrikou, I.; Georgopoulos, A.; Hatziantoniou, S.; Mavromoustakos, T.; Demetzos, C. A comparative study of the effects of cholesterol and sclareol, a bioactive labdane type diterpene, on phospholipid bilayers. Chem. Phys. Lipids 2005, 133, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Matsingou, C.; Demetzos, C. Calorimetric study on the induction of interdigitated phase in hydrated DPPC bilayers by bioactive labdanes and correlation to their liposome stability: The role of chemical structure. Chem. Phys. Lipids 2007, 145, 45–62. [Google Scholar] [CrossRef]
- Matsingou, C.; Hatziantoniou, S.; Georgopoulos, A.; Dimas, K.; Terzis, A.; Demetzos, C. Labdane-type diterpenes: Thermal effects on phospholipid bilayers, incorporation into liposomes and biological activity. Chem. Phys. Lipids 2006, 138, 1–11. [Google Scholar] [CrossRef]
- Benz, J.; Bergner, A.; Hofmann, A.; Demange, P.; Göttig, P.; Liemann, S.; Huber, R.; Voges, D. The structure of recombinant human annexin VI in crystals and membrane-bound. J. Mol. Biol. 1996, 260, 638–643. [Google Scholar] [CrossRef]
- Avila-Sakar, A.J.; Kretsinger, R.H.; Creutz, C.E. Membrane-bound 3D structures reveal the intrinsic flexibility of annexin VI. J. Struct. Biol. 2000, 30, 54–62. [Google Scholar] [CrossRef]
- Buzhynskyy, N.; Golczak, M.; Lai-Kee-Him, J.; Lambert, O.; Tessier, B.; Gounou, C.; Bérat, R.; Simon, A.; Granier, T.; Chevalier, J.M.; et al. Annexin-A6 presents two modes of association with phospholipid membranes. A combined QCM-D, AFM and cryo-TEM study. J. Struct. Biol. 2009, 168, 107–116. [Google Scholar] [CrossRef]
- Horcas, I.; Fernández, R.; Gómez-Rodríguez, J.M.; Colchero, J.; Gómez-Herrero, J.; Baró, A.M. WSXM: A software for scanning probe microscopy and a tool for nanotechnology. Rev. Sci. Instrum. 2007, 78, 013705. [Google Scholar] [CrossRef] [PubMed]
- Bandorowicz-Pikula, J.; Kirilenko, A.; Van Deursen, R.; Golczak, M.; Kühnel, M.; Lancelin, J.M.; Pikula, S.; Buchet, R. A Putative Consensus Sequence for the Nucleotide-Binding Site of Annexin A6. Biochemistry 2003, 42, 9137–9146. [Google Scholar] [CrossRef] [PubMed]
- Cyboron, G.W.; Wuthier, R.E. Purification and initial characterization of intrinsic membrane-bound alkaline phosphatase from chicken epiphyseal cartilage. J. Biol. Chem. 1981, 256, 7262–7268. [Google Scholar]
- Hosokawa, R.; Uchida, Y.; Fujiwara, S.; Noguchi, T. Lactate dehydrogenase isoenzymes are present in matrix vesicles. J. Biol. Chem. 1988, 263, 10045–10047. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Bolean, M.; Simão, A.M.S.; Favarin, B.Z.; Millán, J.L.; Ciancaglini, P. The effect of cholesterol on the reconstitution of alkaline phosphatase into liposomes. Biophys. Chem. 2010, 152, 74–79. [Google Scholar] [CrossRef]
- Bolean, M.; Simão, A.M.; Favarin, B.Z.; Millán, J.L.; Ciancaglini, P. Thermodynamic properties and characterization of proteoliposomes rich in microdomains carrying alkaline phosphatase. Biophys. Chem. 2011, 158, 111–118. [Google Scholar] [CrossRef]
- Bolean, M.; Simão, A.M.; Kiffer-Moreira, T.; Hoylaerts, M.F.; Millán, J.L.; Itri, R.; Ciancaglini, P. Proteoliposomes with the ability to transport Ca(2+) into the vesicles and hydrolyze phosphosubstrates on their surface. Arch. Biochem. Biophys. 2015, 584, 79–89. [Google Scholar] [CrossRef]
- Bolean, M.; Borin, I.A.; Simão, A.M.S.; Bottini, M.; Bagatolli, L.A.; Hoylaerts, M.F.; Millán, J.L.; Ciancaglini, P. Topographic analysis by atomic force microscopy of proteoliposomes matrix vesicle mimetics harboring TNAP and AnxA5. Biochim. Biophys. Acta Biomembr. 2017, 1859, 1911–1920. [Google Scholar] [CrossRef]
- Bolean, M.; Simão, A.M.S.; Barioni, M.B.; Favarin, B.Z.; Sebinelli, H.B.; Veschi, E.A.; Janku, T.A.B.; Bottini, M.; Hoylaerts, M.F.; Itri, R.; et al. Biophysical aspects of biomineralization. Biophys. Rev. 2017, 9, 747–760. [Google Scholar] [CrossRef]
- Hartree, E.F. Determination of protein: A modification of the Lowry method that gives a linear photometric response. Anal. Biochem. 1972, 48, 422–427. [Google Scholar] [CrossRef]
- Andrade, M.A.R.; Favarin, B.Z.; Derradi, R.; Bolean, M.; Simão, A.M.S.; Millán, J.L.; Ciancaglini, P.; Ramos, A.P. Pendant-drop method coupled to ultraviolet-visible spectroscopy: A useful tool to investigate interfacial phenomena. Coll. Surf. A. 2016, 504, 305–311. [Google Scholar] [CrossRef]
- Sebinelli, H.G.; Borin, I.A.; Ciancaglini, P.; Bolean, M. Topographical and mechanical properties of liposome surfaces harboring Na,K-ATPase by means of atomic force microscopy. Soft Matter. 2019, 15, 2737–2745. [Google Scholar] [CrossRef]
- Sztolsztener, M.E.; Strzelecka-Kiliszek, A.; Pikula, S.; Tylki-Szymanska, A.; Bandorowicz-Pikula, J. Cholesterol as a factor regulating intracellular localization of annexin A6 in Niemann-Pick type C human skin fibroblasts. Arch. Biochem. Biophys. 2010, 493, 221–233. [Google Scholar] [CrossRef]
- Wu, L.N.Y.; Genge, B.R.; Dunkelberger, D.G.; LeGeros, R.Z.; Concannon, B.; Wuthier, R.E. Physicochemical characterization of the nucleational core of matrix vesicles. J. Biol. Chem. 1997, 272, 4404–4411. [Google Scholar] [CrossRef]
- Cmoch, A.; Strzelecka-Kiliszek, A.; Palczewska, M.; Groves, P.; Pikula, S. Matrix vesicles isolated from mineralization-competent Saos-2 cells are selectively enriched with annexins and S100 proteins. Biochem. Biophys. Res. Commun. 2011, 412, 683–687. [Google Scholar] [CrossRef]
- Arispe, N.; Rojas, E.; Genge, B.R.; Wu, L.N.; Wuthier, R.E. Similarity in calcium channel activity of annexin V and matrix vesicles in planar lipid bilayers. Biophys. J. 1996, 71, 1764–1775. [Google Scholar] [CrossRef]
- Kaneko, N.; Ago, H.; Matsuda, R.; Inagaki, E.; Miyano, M. Crystal structure of annexin V with its ligand K-201 as a calcium channel activity inhibitor. J. Mol. Biol. 1997, 274, 16–20. [Google Scholar] [CrossRef]
- Kaneko, N.; Matsuda, R.; Toda, M.; Shimamoto, K. Inhibition of annexin V-dependent Ca2+ movement in large unilamellar vesicles by K201, a new 1,4-benzothiazepine derivative. Biochim. Biophys. Acta 1997, 1330, 1–7. [Google Scholar] [CrossRef]
- Hofmann, A.; Escherich, A.; Lewit-Bentley, A.; Benz, J.; Raguenes-Nicol, C.; Russo-Marie, F.; Gerke, V.; Moroder, L.; Huber, R. Interactions of benzodiazepine derivatives with annexins. J. Biol. Chem. 1998, 273, 2885–2894. [Google Scholar] [CrossRef]
- Wang, W.; Xu, J.; Kirsch, T. Annexin-mediated Ca2+ influx regulates growth plate chondrocyte maturation and apoptosis. J. Biol. Chem. 2003, 278, 3762–3769. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
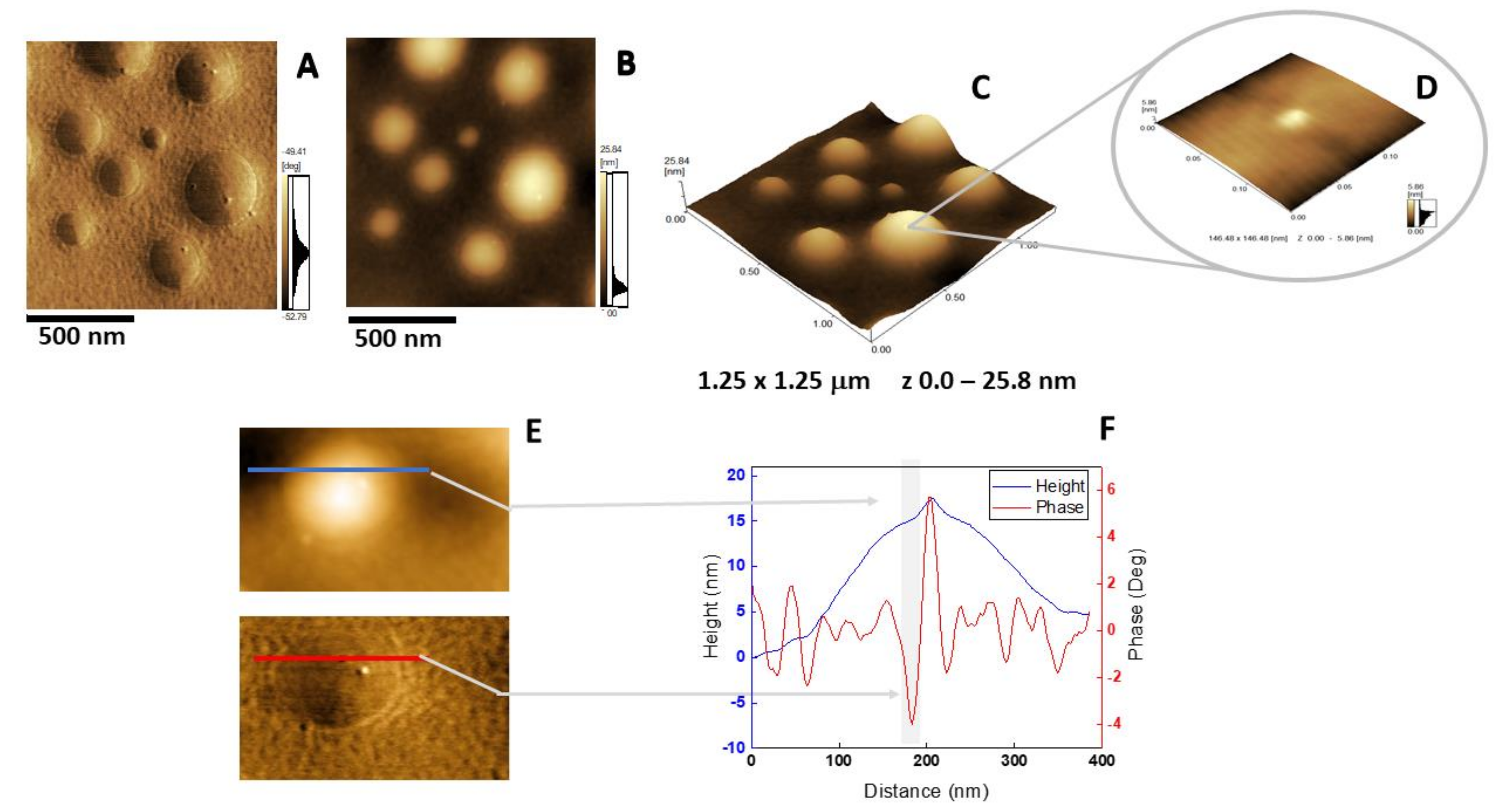{kind=link}
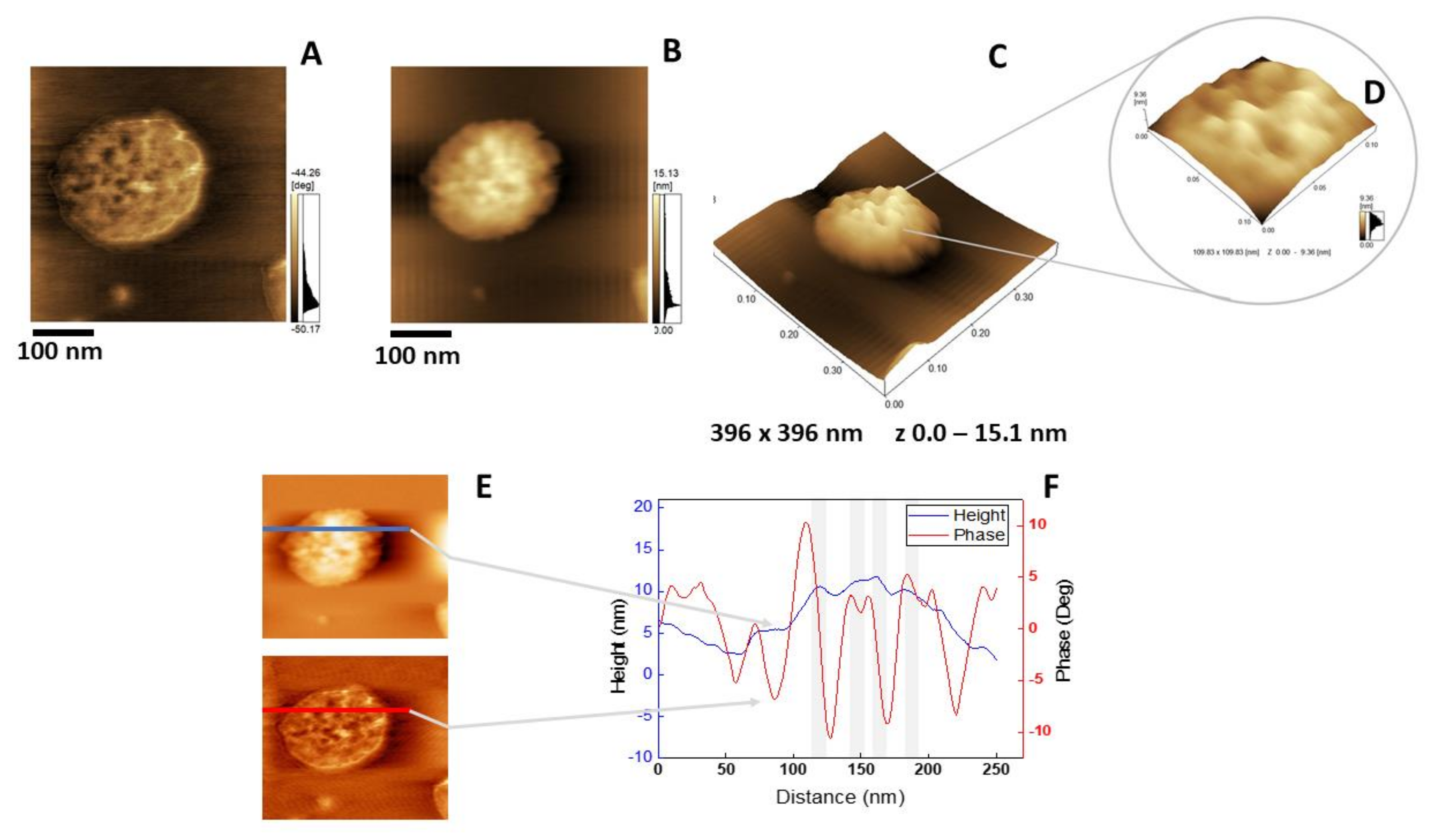{kind=link}
{kind=link}
| Lipid Composition (Molar Ratio) | Ca2+ (2mM) | πexc in Monolayers (mN/m) | [Protein] Incorporated in Liposomes (µg/mL) |
|---|---|---|---|
| DPPC | − | 22.2 ± 2.5 | 18.0 ± 0.4 |
| + | 25.1 ± 2.7 | 59.0 ± 1.1 | |
| DPPC:DPPS (9:1) | − | 23.7 ± 3.4 | 17.0 ± 0.3 |
| + | 32.9 ± 2.0 | 43.0 ± 0.9 | |
| DPPC:Chol (6:4) | − | ND* | 0.60 ± 0.01 |
| + | ND* | 13.0 ± 0.3 | |
| DPPC:Chol:DPPS (5:4:1) | − | 36.7 ± 1.0 | 19.0 ± 0.4 |
| + | 34.3 ± 1.3 | 54.0 ± 1.0 |
| Without Ca2+ | With Ca2+ | ||||||
|---|---|---|---|---|---|---|---|
| Lipid Composition (Molar Ratio) | AnxA6 |
ΔH (Kcal/mol) |
Tm (°C) | Δt1/2 |
ΔH (Kcal/mol) |
Tm (°C) | Δt1/2 |
| DPPC | − | 9.25 | 40.8 | 1.00 | 8.08 | 40.8 | 1.02 |
| + | 6.75 | 41.2 | 1.07 | 6.08 | 41.0 | 1.09 | |
| DPPC:DPPS (9:1) | − | 6.56 | 41.6 | 1.42 | 8.72 | 42.3 | 2.80 |
| + | 5.29 | 41.7 | 1.50 | 4.96 | 42.7 | 2.29 | |
| DPPC:Chol (6:4) | − | 1.24 | 46.7 | 29.59 | 0.57 | 44.3 | 22.86 |
| + | 0.70 | 46.2 | 26.97 | 0.36 | 49.5 | 24.66 | |
| DPPC:Chol:DPPS (5:4:1) | − | 5.36 | 43.6 | 14.54 | 1.14 | 46.3 | 19.37 |
| + | 0.06 | 42.4 | 1.63 | 0.43 | 46.2 | 13.89 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veschi, E.A.; Bolean, M.; Strzelecka-Kiliszek, A.; Bandorowicz-Pikula, J.; Pikula, S.; Granjon, T.; Mebarek, S.; Magne, D.; Ramos, A.P.; Rosato, N.; et al. Localization of Annexin A6 in Matrix Vesicles During Physiological Mineralization. Int. J. Mol. Sci. 2020, 21, 1367. https://doi.org/10.3390/ijms21041367
Veschi EA, Bolean M, Strzelecka-Kiliszek A, Bandorowicz-Pikula J, Pikula S, Granjon T, Mebarek S, Magne D, Ramos AP, Rosato N, et al. Localization of Annexin A6 in Matrix Vesicles During Physiological Mineralization. International Journal of Molecular Sciences. 2020; 21(4):1367. https://doi.org/10.3390/ijms21041367
Chicago/Turabian StyleVeschi, Ekeveliny Amabile, Maytê Bolean, Agnieszka Strzelecka-Kiliszek, Joanna Bandorowicz-Pikula, Slawomir Pikula, Thierry Granjon, Saida Mebarek, David Magne, Ana Paula Ramos, Nicola Rosato, and et al. 2020. "Localization of Annexin A6 in Matrix Vesicles During Physiological Mineralization" International Journal of Molecular Sciences 21, no. 4: 1367. https://doi.org/10.3390/ijms21041367
APA StyleVeschi, E. A., Bolean, M., Strzelecka-Kiliszek, A., Bandorowicz-Pikula, J., Pikula, S., Granjon, T., Mebarek, S., Magne, D., Ramos, A. P., Rosato, N., Millán, J. L., Buchet, R., Bottini, M., & Ciancaglini, P. (2020). Localization of Annexin A6 in Matrix Vesicles During Physiological Mineralization. International Journal of Molecular Sciences, 21(4), 1367. https://doi.org/10.3390/ijms21041367