Differential Effects of Dietary Supplementation of Krill Meal, Soybean Meal, Butyrate, and Bactocell® on the Gene Expression of Atlantic Salmon Head Kidney

 ,
,  ,
,  ,
, 
 , ,
, ,  ,
, 
Abstract
1. Introduction
2. Results and Discussion
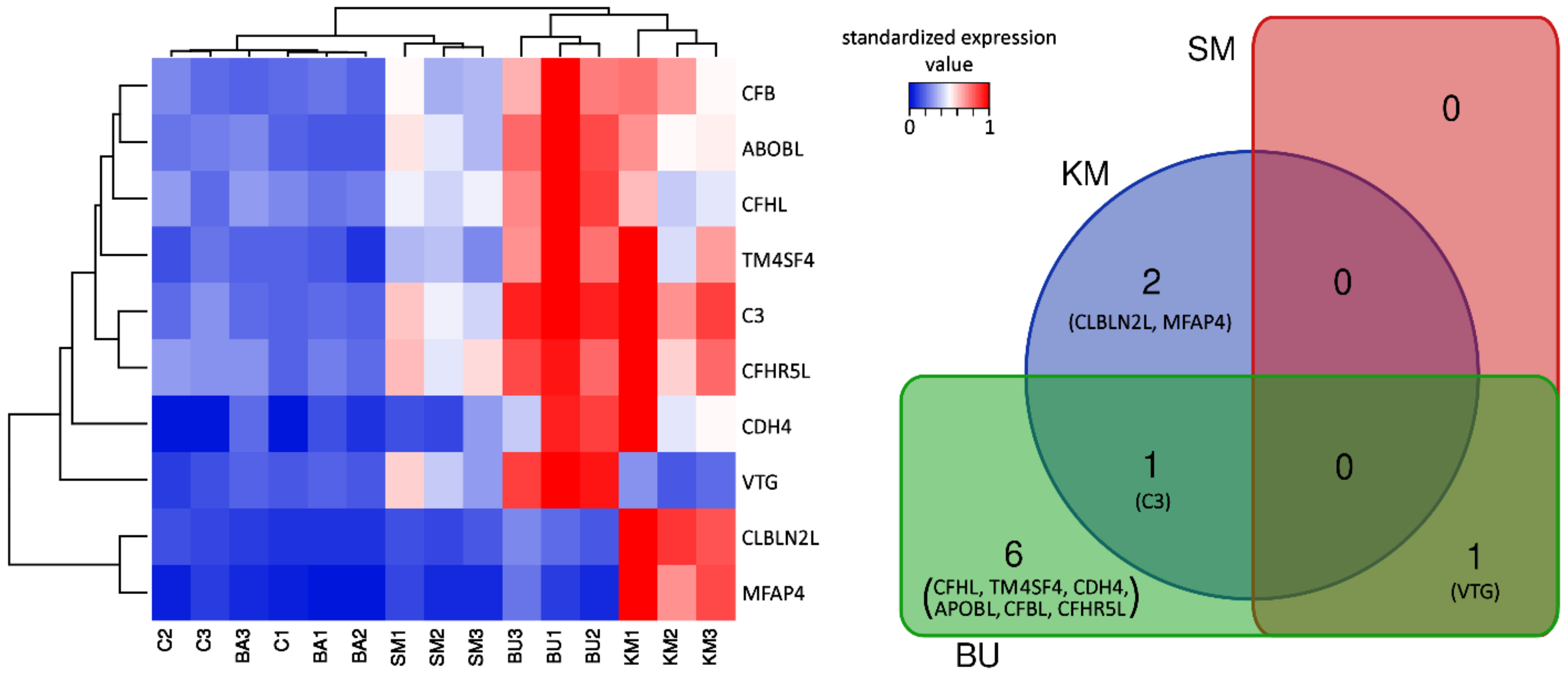
2.1. An Overview of the Effects of Different Feeds on Immune-Related Gene Expression in Head Kidney
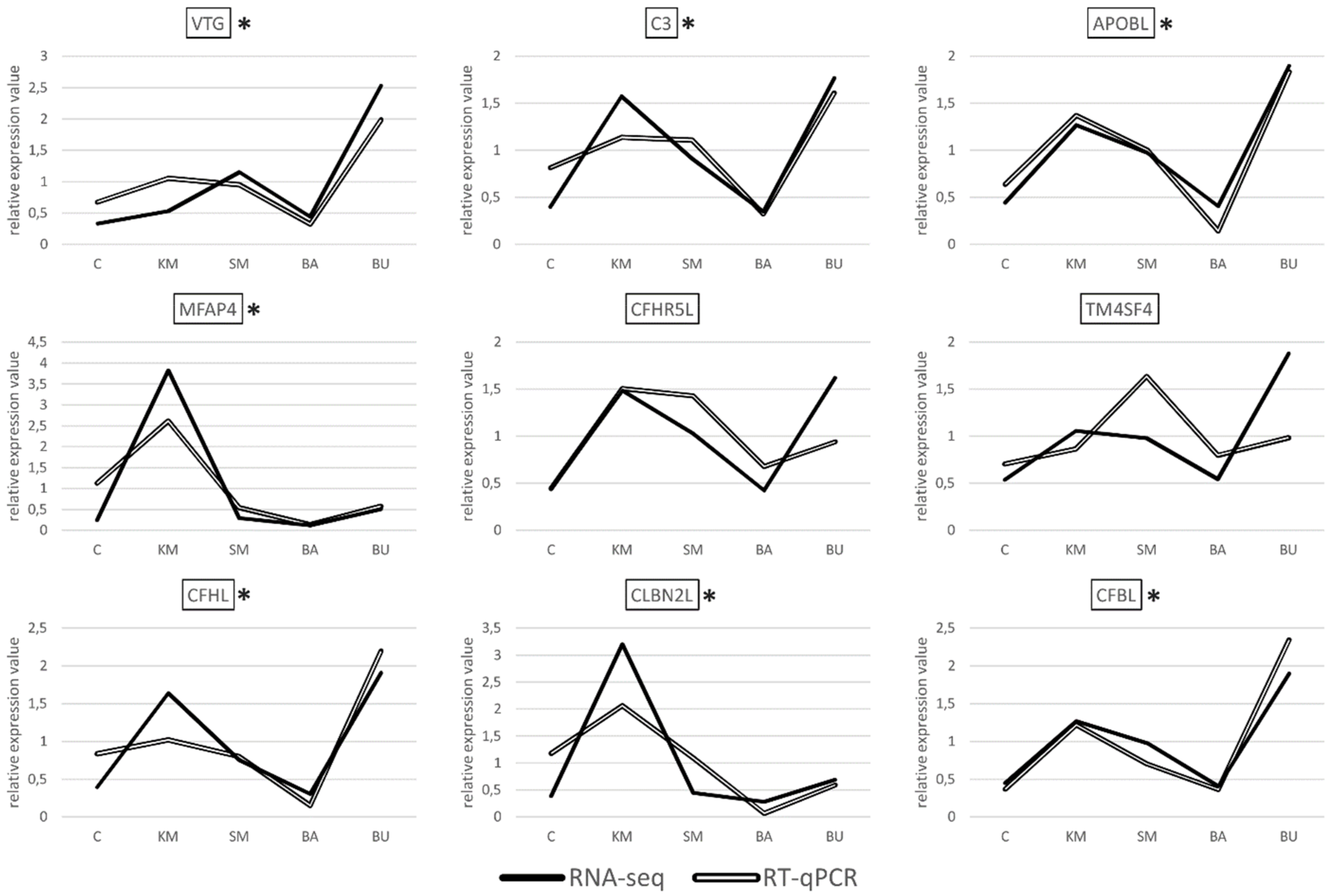
2.2. Validation of RNA-Seq Data by RT-qPCR
2.3. Modulation of Complement Proteins in Response to BU- and KM-Rich Diets
2.4. Vitellogenin Expression was Strongly Induced by the BU-Rich Diet
2.5. Involvement of Microfibril-Associated Protein 4 in the KM-Rich Diet Response
2.6. Possible Involvement of the Other DEGs and Immune Response
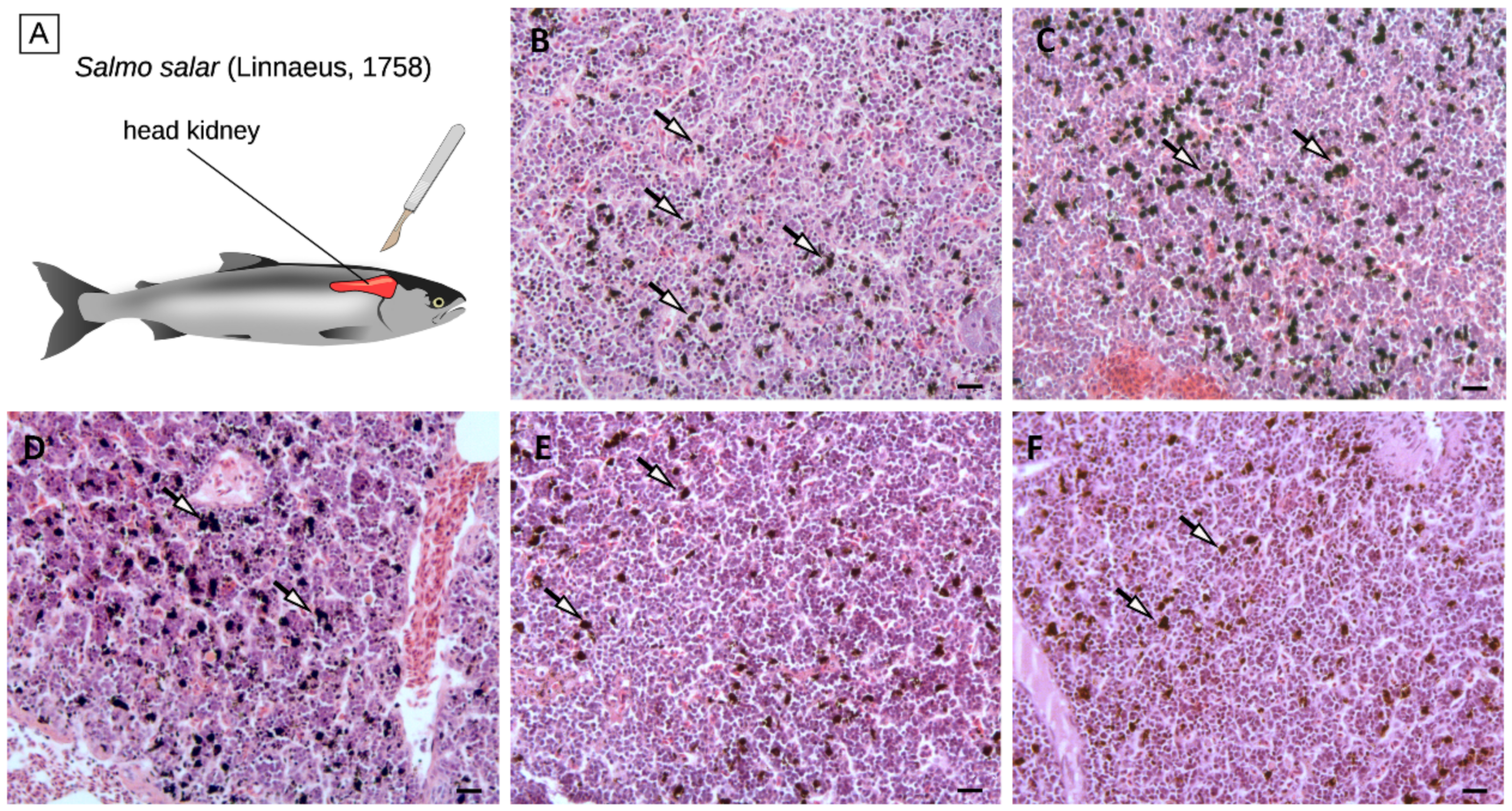
2.7. Histological Analyses
3. Conclusions
4. Methods
4.1. Experimental Setup
4.2. RNA Extraction and RNA-Sequencing Analysis
4.3. Gene Expression Analysis
4.4. Validation of Gene Expression Trends by RT-qPCR
4.5. Histological Image Analysis
4.6. Availability of Data and Materials
4.7. Ethics Approval
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| APOBL | apolipoprotein B-100-like |
| BA | Bactocell® |
| BU | butyrate |
| C | control |
| C3 | complement component C3 |
| CFBL | complement factor B-like |
| CBLN2L | cerebellin 2-like |
| CFHR5L | complement factor H-related protein 5-like |
| CDH4 | cadherin 4 |
| CFHL | complement factor H-like |
| DEGs | differentially expressed genes |
| FC | fold change |
| FHR | factor H-related proteins |
| FDR | false discovery rate |
| HISAT2 | hierarchical indexing for spliced alignment of transcripts |
| KM | krill meal |
| MFAP-4 | microfibril-associated protein 4 |
| MMCs | melanomacrophage centers |
| RT-qPCR | reverse transcriptase- quantitative polymerase chain reaction |
| SM | soybean meal |
| TM4SF4 | transmembrane 4L6 family member 4-like |
| TPM | Transcripts Per Million |
| VTG | vitellogenin |
References
- Joerink, M.; Ribeiro, C.M.; Stet, R.J.; Hermsen, T.; Savelkoul, H.F.; Wiegertjes, G.F. Head kidney-derived macrophages of common carp (Cyprinus carpio L.) show plasticity and functional polarization upon differential stimulation. J. Immunol. 2006, 177, 61–69. [Google Scholar]
- Wang, W.; Sun, J.; Liu, C.; Xue, Z. Application of immunostimulants in aquaculture: Current knowledge and future perspectives. Aquac. Res. 2017, 48, 1–23. [Google Scholar]
- Geven, E.J.; Klaren, P.H. The teleost head kidney: Integrating thyroid and immune signalling. Dev. Comp. Immunol. 2017, 66, 73–83. [Google Scholar]
- Harris, J.; Bird, D.J. Modulation of the fish immune system by hormones. Vet. Immunol. Immunopathol. 2000, 77, 163–176. [Google Scholar]
- Weyts, F.; Cohen, N.; Flik, G.; Verburg-van Kemenade, B. Interactions between the immune system and the hypothalamo-pituitary-interrenal axis in fish. Fish Shellfish Immunol. 1999, 9, 1–20. [Google Scholar]
- Deutsch, L. Evaluation of the effect of Neptune Krill Oil on chronic inflammation and arthritic symptoms. J. Am. Coll. Nutr. 2007, 26, 39–48. [Google Scholar]
- Batetta, B.; Griinari, M.; Carta, G.; Murru, E.; Ligresti, A.; Cordeddu, L.; Giordano, E.; Sanna, F.; Bisogno, T.; Uda, S.; et al. Endocannabinoids may mediate the ability of (n-3) fatty acids to reduce ectopic fat and inflammatory mediators in obese Zucker rats. J. Nutr. 2009, 139, 1495–1501. [Google Scholar]
- Amar, E.C.; Kiron, V.; Satoh, S.; Okamoto, N.; Watanabe, T.J. Effects of dietary βcarotene on the immune response of rainbow trout Oncorhynchus mykiss. Fish Sci. 2000, 66, 1068–1075. [Google Scholar]
- Howe, P.R.; Clifton, P.M.; James, M.J.J.L. Equal antithrombotic and triglyceride-lowering effectiveness of eicosapentaenoic acid-rich and docosahexaenoic acid-rich fish oil supplements. Lipids 1999, 34, S307–S308. [Google Scholar]
- Ringø, E.; Zhou, Z.; Olsen, R.; Song, S.J.A.N. Use of chitin and krill in aquaculture–the effect on gut microbiota and the immune system: A review. Aquac. Nutr. 2012, 18, 117–131. [Google Scholar]
- Ringø, E.; Sperstad, S.; Myklebust, R.; Mayhew, T.M.; Mjelde, A.; Melle, W.; Olsen, R.E.J.A.R. The effect of dietary krill supplementation on epithelium-associated bacteria in the hindgut of Atlantic salmon (Salmo salar L.): A microbial and electron microscopical study. Aquac. Res. 2006, 37, 1644–1653. [Google Scholar]
- Esteban, M.; Cuesta, A.; Ortuno, J.; Meseguer, J. Immunomodulatory effects of dietary intake of chitin on gilthead seabream (Sparus aurata L.) innate immune system. Fish Shellfish Immunol. 2001, 11, 303–315. [Google Scholar]
- Esteban, M.; Mulero, V.; Cuesta, A.; Ortuno, J.; Meseguer, J. Immunology Effects of injecting chitin particles on the innate immune response of gilthead seabream (Sparus aurataL). Fish Shellfish Immunol. 2000, 10, 543–554. [Google Scholar]
- Olsen, R.; Suontama, J.; Langmyhr, E.; Mundheim, H.; Ringø, E.; Melle, W.; Malde, M.; Hemre, G.I. The replacement of fish meal with Antarctic krill, Euphausia superba in diets for Atlantic salmon, Salmo salar. Aquac. Nutr. 2006, 12, 280–290. [Google Scholar]
- Cuesta, A.; Esteban, M.Á.; Meseguer, J. In vitro effect of chitin particles on the innate cellular immune system of gilthead seabream (Sparus aurata L). Fish Shellfish Immunol. 2003, 15, 1–11. [Google Scholar]
- Amar, E.C.; Kiron, V.; Satoh, S.; Watanabe, T. Enhancement of innate immunity in rainbow trout (Oncorhynchus mykiss Walbaum) associated with dietary intake of carotenoids from natural products. Fish Shellfish Immunol. 2004, 16, 527–537. [Google Scholar]
- Bui, H.T.D.; Khosravi, S.; Fournier, V.; Herault, M.; Lee, K.-J. Growth performance, feed utilization, innate immunity, digestibility and disease resistance of juvenile red seabream (Pagrus major) fed diets supplemented with protein hydrolysates. Aquaculture 2014, 418, 11–16. [Google Scholar]
- Khosravi, S.; Rahimnejad, S.; Herault, M.; Fournier, V.; Lee, C.-R.; Bui, H.T.D.; Jeong, J.-B.; Lee, K.-J. Effects of protein hydrolysates supplementation in low fish meal diets on growth performance, innate immunity and disease resistance of red sea bream Pagrus major. Fish Shellfish Immunol. 2015, 45, 858–868. [Google Scholar]
- Baeverfjord, G.; Krogdahl, Å. Development and regression of soybean meal induced enteritis in Atlantic salmon, Salmo salar L., distal intestine: A comparison with the intestines of fasted fish. J. Fish. Dis. 1996, 19, 375–387. [Google Scholar]
- Krogdahl, Å.; Bakke-McKellep, A.; Baeverfjord, G. Effects of graded levels of standard soybean meal on intestinal structure, mucosal enzyme activities, and pancreatic response in Atlantic salmon (Salmo salar L.). Aquac. Nutr. 2003, 9, 361–371. [Google Scholar]
- Krogdahl, A.; Bakke-McKellep, A.; Roed, K.; Baeverfjord, G. Feeding Atlantic salmon Salmo salar L. soybean products: Effects on disease resistance (furunculosis), and lysozyme and IgM levels in the intestinal mucosa. Aquac. Nutr. 2000, 6, 77–84. [Google Scholar]
- Rumsey, G.L.; Siwicki, A.K.; Anderson, D.P.; Bowser, P.R. Immunopathology. Effect of soybean protein on serological response, non-specific defense mechanisms, growth, and protein utilization in rainbow trout. Vet. Immunol. Immunopathol. 1994, 41, 323–339. [Google Scholar]
- Bakke-McKellep, A.; Koppang, E.; Gunnes, G.; Sanden, M.; Hemre, G.I.; Landsverk, T.; Krogdahl, Å.J. Histological, digestive, metabolic, hormonal and some immune factor responses in Atlantic salmon, Salmo salar L., fed genetically modified soybeans. J. Fish Dis. 2007, 30, 65–79. [Google Scholar]
- Lazado, C.C.; Caipang, C.M.A.; Estante, E.G. Prospects of host-associated microorganisms in fish and penaeids as probiotics with immunomodulatory functions. Fish Shellfish Immunol. 2015, 45, 2–12. [Google Scholar]
- Lazado, C.C.; Caipang, C.M.A.; Gallage, S.; Brinchmann, M.F.; Kiron, V. Expression profiles of genes associated with immune response and oxidative stress in Atlantic cod, Gadus morhua head kidney leukocytes modulated by live and heat-inactivated intestinal bacteria. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2010, 155, 249–255. [Google Scholar]
- Ferguson, R.; Merrifield, D.L.; Harper, G.M.; Rawling, M.D.; Mustafa, S.; Picchietti, S.; Balcàzar, J.L.; Davies, S.J. The effect of Pediococcus acidilactici on the gut microbiota and immune status of on-growing red tilapia (Oreochromis niloticus). J. Appl. Microbiol. 2010, 109, 851–862. [Google Scholar]
- Shelby, R.A.; Lim, C.; Yildirim-Aksoy, M.; Delaney, M.A. Effects of probiotic diet supplements on disease resistance and immune response of young Nile tilapia, Oreochromis niloticus. J. Appl. Microbiol. 2006, 18, 23–34. [Google Scholar]
- Standen, B.; Rawling, M.; Davies, S.; Castex, M.; Foey, A.; Gioacchini, G.; Carnevali, O.; Merrifield, D.J. Probiotic Pediococcus acidilactici modulates both localised intestinal-and peripheral-immunity in tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2013, 35, 1097–1104. [Google Scholar]
- Hoseinifar, S.H.; Esteban, M.Á.; Cuesta, A.; Sun, Y.-Z. Prebiotics and fish immune response: A review of current knowledge and future perspectives. Rev. Fisheries Sci. Aquac. 2015, 23, 315–328. [Google Scholar]
- Neissi, A.; Rafiee, G.; Nematollahi, M.; Safari, O. The effect of Pediococcus acidilactici bacteria used as probiotic supplement on the growth and non-specific immune responses of green terror, Aequidens rivulatus. Fish Shellfish Immunol. 2013, 35, 1976–1980. [Google Scholar]
- Knudsen, K.E.B.; Serena, A.; Canibe, N.; Juntunen, K.S. New insight into butyrate metabolism. Proc. Nutr. Soc. 2003, 62, 81–86. [Google Scholar]
- Corrêa-Oliveira, R.; Fachi, J.L.; Vieira, A.; Sato, F.T.; Vinolo, M.A.R. Regulation of immune cell function by short-chain fatty acids. Clin. Transl. Immunol. 2016, 5, e73. [Google Scholar]
- Tian, L.; Zhou, X.-Q.; Jiang, W.-D.; Liu, Y.; Wu, P.; Jiang, J.; Kuang, S.-Y.; Tang, L.; Tang, W.-N.; Zhang, Y.-A.; et al. Sodium butyrate improved intestinal immune function associated with NF-κB and p38MAPK signalling pathways in young grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2017, 66, 548–563. [Google Scholar]
- Bjerkeng, B.; Storebakken, T.; Wathne, E. Cholesterol and short-chain fatty acids in diets for Atlantic salmon Salmo salar (L.): Effects on growth, organ indices, macronutrient digestibility, and fatty acid composition. Aquac. Nutr. 1999, 5, 181–192. [Google Scholar]
- Meseguer, J.; Esteban, M.; Agulleiro, B. Stromal cells, macrophages and lymphoid cells in the head-kidney of sea bass (Dicentrarchus labrax L.). An ultrastructural study. Arch. Histol. Cytol. 1991, 54, 299–309. [Google Scholar]
- Meseguer, J.; López-Ruiz, A.; Garcí-Ayala, A. Reticulo-endothelial stroma of the head-kidney from the seawater teleost gilthead seabream (Sparus aurata L.): An ultrastructural and cytochemical study. Anat. Rec. 1995, 241, 303–309. [Google Scholar]
- Zapata, A. Ultrastructural study of the teleost fish kidney. Dev. Comp. Immunol. 1979, 3, 55–65. [Google Scholar]
- Dannevig, B.H.; Lauve, A.; Press, C.M.; Landsverk, T. Receptor-mediated endocytosis and phagocytosis by rainbow trout head kidney sinusoidal cells. Fish Shellfish Immunol. 1994, 4, 3–18. [Google Scholar]
- Steinel, N.C.; Bolnick, D.I. Melanomacrophage centers as a histological indicator of immune function in fish and other poikilotherms. Front. Immunol. 2017, 8, 827. [Google Scholar]
- Press, C.M.; Evensen, Ø.; Reitan, L.; Landsverk, T. Retention of furunculosis vaccine components in Atlantic salmon, Salmo salar L., following different routes of vaccine administration. J. Fish. Dis. 1996, 19, 215–224. [Google Scholar]
- Ellis, A. Antigen-trapping in the spleen and kidney of the plaice Pleuronectes platessa L. J. Fish. Dis. 1980, 3, 413–426. [Google Scholar]
- Jalili, M.; Jin, Y.; Bones, A.M.; Olsen, Y.; Vadstein, O.; Østensen, M.-A.; Buonocore, F.; Gerdol, M.; Pallavicini, A.; Scapigliati, G. Dietary fatty acid source has little effect on the development of the immune system in the pyloric caeca of Atlantic salmon fry. Sci. Rep. 2019, 9, 27. [Google Scholar]
- Su, Z.; Łabaj, P.P.; Li, S.; Thierry-Mieg, J.; Thierry-Mieg, D.; Shi, W.; Wang, C.; Schroth, G.P.; Setterquist, R.A.; Thompson, J.F.; et al. A comprehensive assessment of RNA-seq accuracy, reproducibility and information content by the Sequencing Quality Control Consortium. Nat. Biotechnol. 2014, 32, 903–914. [Google Scholar]
- Holland, M.C.H.; Lambris, J.D. The complement system in teleosts. Fish Shellfish Immunol. 2002, 12, 399–420. [Google Scholar]
- Boshra, H.; Li, J.; Sunyer, J. Recent advances on the complement system of teleost fish. Fish Shellfish Immunol. 2006, 20, 239–262. [Google Scholar]
- Magnadóttir, B.; Lange, S.; Gudmundsdottir, S.; Bøgwald, J.; Dalmo, R. Ontogeny of humoral immune parameters in fish. Fish Shellfish Immunol. 2005, 19, 429–439. [Google Scholar]
- Lange, S.; Bambir, S.; Dodds, A.W.; Magnadóttir, B.J.D. An immunohistochemical study on complement component C3 in juvenile Atlantic halibut (Hippoglossus hippoglossus L). Dev. Comp. Immunol. 2004, 28, 593–601. [Google Scholar]
- Morgan, B.P. The complement system: An overview. In Complement Methods and Protocols; Humana Press: Totowa, NJ, USA, 2000; pp. 1–13. [Google Scholar]
- Franchini, S.; Zarkadis, I.K.; Sfyroera, G.; Sahu, A.; Moore, W.T.; Mastellos, D.; LaPatra, S.E.; Lambris, J.D. Cloning and purification of the rainbow trout fifth component of complement (C5). Dev. Comp. Immunol. 2001, 25, 419–430. [Google Scholar]
- Zipfel, P.F.; Jokiranta, T.S.; Hellwage, J.; Koistinen, V.; Meri, S.J.I. The factor H protein family. Immunopharmacology 1999, 42, 53–60. [Google Scholar]
- Murphy, B.F.; D’apice, A.J. Identification of the components of glomerular immune deposits using monoclonal antibodies. Pathology 1988, 20, 130–136. [Google Scholar]
- McRae, J.L.; Duthy, T.G.; Griggs, K.M.; Ormsby, R.J.; Cowan, P.J.; Cromer, B.A.; McKinstry, W.J.; Parker, M.W.; Murphy, B.F.; Gordon, D.L. Human factor H-related protein 5 has cofactor activity, inhibits C3 convertase activity, binds heparin and C-reactive protein, and associates with lipoprotein. J. Immunol. 2005, 174, 6250–6256. [Google Scholar]
- Ren, G.; Doshi, M.; Hack, B.K.; Alexander, J.J.; Quigg, R.J. Isolation and characterization of a novel rat factor H-related protein that is up-regulated in glomeruli under complement attack. J. Biol. Chem. 2002, 277, 48351–48358. [Google Scholar]
- Markiewski, M.M.; Lambris, J.D. The role of complement in inflammatory diseases from behind the scenes into the spotlight. Am. J. Pathol. 2007, 171, 715–727. [Google Scholar]
- Skerka, C.; Hellwage, J.; Weber, W.; Tilkorn, A.; Buck, F.; Marti, T.; Kampen, E.; Beisiegel, U.; Zipfel, P.F. The human factor H-related protein 4 (FHR-4) a novel short consensus repeat-containing protein is associated with human triglyceride-rich lipoproteins. J. Biol. Chem. 1997, 272, 5627–5634. [Google Scholar]
- Burgos-Aceves, M.A.; Cohen, A.; Smith, Y.; Faggio, C. Estrogen regulation of gene expression in the teleost fish immune system. Fish Shellfish Immunol. 2016, 58, 42–49. [Google Scholar]
- Iwanowicz, L.R.; Stafford, J.L.; Patino, R.; Bengten, E.; Miller, N.W.; Blazer, V.S. Channel catfish (Ictalurus punctatus) leukocytes express estrogen receptor isoforms ERα and ERβ2 and are functionally modulated by estrogens. Fish Shellfish Immunol. 2014, 40, 109–119. [Google Scholar]
- Shi, X.; Zhang, S.; Pang, Q. Vitellogenin is a novel player in defense reactions. Fish Shellfish Immunol. 2006, 20, 769–772. [Google Scholar]
- Liu, Q.-H.; Zhang, S.-C.; Li, Z.-J.; Gao, C.-R. Characterization of a pattern recognition molecule vitellogenin from carp (Cyprinus carpio). Immunobiology 2009, 214, 257–267. [Google Scholar]
- Tong, Z.; Li, L.; Pawar, R.; Zhang, S. Vitellogenin is an acute phase protein with bacterial-binding and inhibiting activities. Immunobiology 2010, 215, 898–902. [Google Scholar]
- Garcia, J.; Munro, E.S.; Monte, M.M.; Fourrier, M.C.; Whitelaw, J.; Smail, D.A.; Ellis, A.E. Atlantic salmon (Salmo salar L.) serum vitellogenin neutralises infectivity of infectious pancreatic necrosis virus (IPNV). Fish Shellfish Immunol. 2010, 29, 293–297. [Google Scholar]
- Li, Z.; Zhang, S.; Liu, Q. Vitellogenin functions as a multivalent pattern recognition receptor with an opsonic activity. PLoS ONE 2008, 3, e1940. [Google Scholar]
- Liu, W.; Yang, Y.; Zhang, J.; Gatlin, D.M.; Ringø, E.; Zhou, Z. Effects of dietary microencapsulated sodium butyrate on growth, intestinal mucosal morphology, immune response and adhesive bacteria in juvenile common carp (Cyprinus carpio) pre-fed with or without oxidised oil. Br. J. Nutr. 2014, 112, 15–29. [Google Scholar]
- Pelissero, C.; Bennetau, B.; Babin, P.; Le Menn, F.; Dunogues, J. The estrogenic activity of certain phytoestrogens in the Siberian sturgeon Acipenser baeri. J. Steroid Biochem. Mol. Biol. 1991, 38, 293–299. [Google Scholar]
- Niu, D.; Peatman, E.; Liu, H.; Lu, J.; Kucuktas, H.; Liu, S.; Sun, F.; Zhang, H.; Feng, T.; Zhou, Z.; et al. Microfibrillar-associated protein 4 (MFAP4) genes in catfish play a novel role in innate immune responses. Dev. Comp. Immunol. 2011, 35, 568–579. [Google Scholar]
- Liu, H.; Peatman, E.; Wang, W.; Abernathy, J.; Liu, S.; Kucuktas, H.; Lu, J.; Xu, D.-H.; Klesius, P.; Waldbieser, G.; et al. Molecular responses of calreticulin genes to iron overload and bacterial challenge in channel catfish (Ictalurus punctatus). Dev. Comp. Immunol. 2011, 35, 267–272. [Google Scholar]
- Thomsen, T.; Schlosser, A.; Holmskov, U.; Sorensen, G.L. Ficolins and FIBCD1: Soluble and membrane bound pattern recognition molecules with acetyl group selectivity. Mol. Immunol. 2011, 48, 369–381. [Google Scholar]
- Schlosser, A.; Thomsen, T.; Shipley, J.M.; Hein, P.W.; Brasch, F.; Tornøe, I.; Nielsen, O.; Skjødt, K.; Palaniyar, N.; Steinhilber, W. Microfibril-associated protein 4 binds to surfactant protein A (SP-A) and colocalizes with SP-A in the extracellular matrix of the lung. Scand. J. Immunol. 2006, 64, 104–116. [Google Scholar]
- Lee, C.G.; Da Silva, C.A.; Lee, J.-Y.; Hartl, D.; Elias, J.A. Chitin regulation of immune responses: An old molecule with new roles. Curr. Opin. Immunol. 2008, 20, 684–689. [Google Scholar]
- Shibata, Y.; Metzger, W.J.; Myrvik, Q.N. Chitin particle-induced cell-mediated immunity is inhibited by soluble mannan: Mannose receptor-mediated phagocytosis initiates IL-12 production. J. Immunol. 1997, 159, 2462–2467. [Google Scholar]
- Gerwick, L.; Reynolds, W.S.; Bayne, C.J. A precerebellin-like protein is part of the acute phase response in rainbow trout, Oncorhynchus mykiss. Dev. Comp. Immunol. 2000, 24, 597–607. [Google Scholar] [CrossRef]
- Kumar, G.; Hummel, K.; Noebauer, K.; Welch, T.J.; Razzazi-Fazeli, E.; El-Matbouli, M. Proteome analysis reveals a role of rainbow trout lymphoid organs during Yersinia ruckeri infection process. Sci. Rep. 2018, 8, 13998. [Google Scholar] [CrossRef]
- Russell, S.; Hayes, M.A.; Simko, E.; Lumsden, J.S. Plasma proteomic analysis of the acute phase response of rainbow trout (Oncorhynchus mykiss) to intraperitoneal inflammation and LPS injection. Dev. Comp. Immunol. 2006, 30, 393–406. [Google Scholar] [CrossRef]
- Von Gersdorff Jørgensen, L.; Nemli, E.; Heinecke, R.D.; Raida, M.K.; Buchmann, K. Immune-relevant genes expressed in rainbow trout following immunisation with a live vaccine against Ichthyophthirius multifiliis. Dis. Aquat. Org. 2008, 80, 189–197. [Google Scholar]
- Tsoi, S.C.M.; Ewart, K.V.; Penny, S.; Melville, K.; Liebscher, R.S.; Brown, L.L.; Douglas, S.E. Identification of Immune-Relevant Genes from Atlantic Salmon Using Suppression Subtractive Hybridization. Mar. Biotechnol. 2004, 6, 199–214. [Google Scholar] [CrossRef]
- Park, K.C.; Osborne, J.A.; Tsoi, S.C.M.; Brown, L.L.; Johnson, S.C. Expressed sequence tags analysis of Atlantic halibut (Hippoglossus hippoglossus) liver, kidney and spleen tissues following vaccination against Vibrio anguillarum and Aeromonas salmonicida. Fish. Shellfish Immunol. 2005, 18, 393–415. [Google Scholar] [CrossRef]
- Wice, B.M.; Gordon, J.I. A tetraspan membrane glycoprotein produced in the human intestinal epithelium and liver that can regulate cell density-dependent proliferation. J. Biol. Chem. 1995, 270, 21907–21918. [Google Scholar]
- Hansen, J.D.; Woodson, J.C.; Hershberger, P.K.; Grady, C.; Gregg, J.L.; Purcell, M.K. Induction of anti-viral genes during acute infection with Viral hemorrhagic septicemia virus (VHSV) genogroup IVa in Pacific herring (Clupea pallasii). Fish Shellfish Immunol. 2012, 32, 259–267. [Google Scholar]
- Li, S.; Li, C.; Wang, X.; Wang, Y.; Liu, Z.; Zhai, T.; Zhang, Q.J. Molecular cloning and expression of PoIR2, a novel gene involved in immune response in Japanese flounder (Paralichthys olivaceus). J. Ocean Univ. China 2010, 9, 59–64. [Google Scholar]
- Van den Bossche, J.; Van Ginderachter, J.A. E-cadherin: From epithelial glue to immunological regulator. Eur. J. Immunol. 2013, 43, 34–37. [Google Scholar]
- Jiang, Y.; Feng, S.; Zhang, S.; Liu, H.; Feng, J.; Mu, X.; Sun, X.; Xu, P. Transcriptome signatures in common carp spleen in response to Aeromonas hydrophila infection. Fish Shellfish Immunol. 2016, 57, 41–48. [Google Scholar]
- Haugarvoll, E.; Thorsen, J.; Laane, M.; Huang, Q.; Koppang, E.O. Melanogenesis and evidence for melanosome transport to the plasma membrane in a CD83+ teleost leukocyte cell line. Pigment. Cell Res. 2006, 19, 214–225. [Google Scholar]
- Fishelson, L. Cytomorphological alterations of the thymus, spleen, head-kidney, and liver in cardinal fish (Apogonidae, Teleostei) as bioindicators of stress. J. Morphol. 2006, 267, 57–69. [Google Scholar]
- Reddy, S.J. Cadmium effect on histo-biomarkers and melano-macrophage centers in liver and kidney of Cyprinus carpio. World J. Fish. Mar. Sci. 2012, 4, 179–184. [Google Scholar]
- Agius, C.; Roberts, R. Effects of starvation on the melano-macrophage centres of fish. J. Fish. Biol. 1981, 19, 161–169. [Google Scholar]
- Mizuno, S.; Misaka, N.; Miyakoshi, Y.; Takeuchi, K.; Kasahara, N. Effects of starvation on melano-macrophages in the kidney of masu salmon (Oncorhynchus masou). Aquaculture 2002, 209, 247–255. [Google Scholar]
- Rosas, A.; MacGill, R.; Nosanchuk, J.D.; Kozel, T.; Casadevall, A. Activation of the alternative complement pathway by fungal melanins. Clin. Diagn. Lab. Immunol. 2002, 9, 144–148. [Google Scholar]
- Knudsen, D.; Jutfelt, F.; Sundh, H.; Sundell, K.; Koppe, W.; Frøkiær, H. Dietary soya saponins increase gut permeability and play a key role in the onset of soyabean-induced enteritis in Atlantic salmon (Salmo salar L.). Br. J. Nutr. 2008, 100, 120–129. [Google Scholar]
- Abbas, A.K.; Lichtman, A.H.; Pillai, S. Basic Immunology: Functions and Disorders of the Immune System; Elsevier Health Sciences: Philadelphia, PA, USA, 2015. [Google Scholar]
- Davidson, W.S.; Koop, B.F.; Jones, S.J.; Iturra, P.; Vidal, R.; Maass, A.; Jonassen, I.; Lien, S.; Omholt, S.W. Sequencing the genome of the Atlantic salmon (Salmo salar). Genome Biol. 2010, 11, 403. [Google Scholar]
- Kim, D.; Langmead, B.; Salzberg, S.A.-O. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar]
- Tekle, K.M.; Gundersen, S.; Klepper, K.; Bongo, L.A.-O.; Raknes, I.A.; Li, X.; Zhang, W.; Andreetta, C.; Mulugeta, T.D.; Kalas, M.A.-O.; et al. Norwegian e-Infrastructure for Life Sciences (NeLS). F1000 Res. 2018, 7, ELIXIR-968. [Google Scholar] [CrossRef]
- Wagner, G.P.; Kin, K.; Lynch, V.J. Measurement of mRNA abundance using RNA-seq data: RPKM measure is inconsistent among samples. Theory Biosci. 2012, 131, 281–285. [Google Scholar]
- Law, C.W.; Chen, Y.; Shi, W.; Smyth, G.K. Voom: Precision weights unlock linear model analysis tools for RNA-seq read counts. Genome Biol. 2014, 15, R29. [Google Scholar]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential expression analysis of multifactor RNA-Seq experiments with respect to biological variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar]
- Jorgensen, S.M.; Kleveland, E.J.; Grimholt, U.; Gjoen, T. Validation of reference genes for real-time polymerase chain reaction studies in Atlantic salmon. Mar. Biotechnol. 2006, 8, 398–408. [Google Scholar]
- Olsvik, P.A.; Lie, K.K.; Jordal, A.-E.O.; Nilsen, T.O.; Hordvik, I. Evaluation of potential reference genes in real-time RT-PCR studies of Atlantic salmon. BMC Mol. Biol. 2005, 6, 21. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| NCBI Gene ID | Gene Acronym | KM vs C | SM vs C | BA vs C | BU vs C | ||||
|---|---|---|---|---|---|---|---|---|---|
| LogFC | FDR | LogFC | FDR | LogFC | FDR | LogFC | FDR | ||
| 100136426 | VTG | ns | ns | 1.56 | 3.95 × 10−2 | ns | ns | 2.83 | 9.74 × 10−6 |
| 100196216 | MFAP4 | 3.76 | 1.66 × 10−2 | ns | ns | ns | ns | ns | ns |
| 106560553 | CFHL | ns | ns | ns | ns | ns | ns | 2.15 | 1.52 × 10−2 |
| 106560595 | TM4SF4 | ns | ns | ns | ns | ns | ns | 1.7 | 1.22 × 10−2 |
| 106566735 | CDH4 | ns | ns | ns | ns | ns | ns | 4.17 | 3.03 × 10−2 |
| 106571136 | CLBLN2L | 2.81 | 2.82 × 10−3 | ns | ns | ns | ns | ns | ns |
| 106571178 | APOBL | ns | ns | ns | ns | ns | ns | 1.99 | 5.52 × 10−4 |
| 106598656 | CFHR5L | ns | ns | ns | ns | ns | ns | 1.8 | 1.34 × 10−2 |
| 106602980 | CFBL | ns | ns | ns | ns | ns | ns | 1.81 | 1.34 × 10−2 |
| 106604987 | C3 | 1.73 | 5.04 × 10−3 | ns | ns | ns | ns | 2.08 | 4.08 × 10−4 |
| Composition | C | KM | SM | BA | BU |
|---|---|---|---|---|---|
| Fish Meal LT | 12.45 | 10.1 | 14.16 | 14.14 | 14.12 |
| Fish Meal SA Superprime | 12.5 | 7.5 | 11.84 | 11.86 | 11.88 |
| Krill meal | 7.4 | ||||
| Soya HP48, Non-GMO | 17.68 | 17.64 | 17.61 | ||
| Soya SPC, Non-GMO | 18.05 | 18 | 5 | 5 | 5 |
| Wheat Gluten, | 7.7 | 10 | 14.1 | 14.1 | 14.1 |
| Maize Gluten 60 | 5 | 5 | |||
| Pea Protein | 10 | 7.58 | 5 | 5 | 5 |
| Wheat | 12.5 | 12.5 | 11.2 | 11.2 | 11.2 |
| Fish Oil, 18 EPA+DHA | 9.71 | 8.71 | 9.82 | 9.82 | 9.82 |
| Rapeseed Oil, Crude | 9.44 | 9.82 | 9.91 | 9.91 | 9.91 |
| Vitamin and minerals | 2.14 | 2.41 | 2.00 | 2.00 | 2.00 |
| AA-mix | 0.82 | 1.04 | 0.97 | 0.97 | 0.97 |
| Bactocell PA10 | 0.03 | ||||
| Sodium Polyhydroxy Butyrate 30% coated | 0.05 | ||||
| YTTRIUM | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 |
| Lucantin Pink CWD 10% | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 |
| Water change | −0.4 | −0.15 | −1.76 | −1.76 | −1.75 |
| Total | 100 | 100 | 100 | 100 | 100 |
| NCBI Gene ID | Gene Acronym | Forward Primer (5′-> 3′) | Reverse Primer (5′-> 3′) |
|---|---|---|---|
| 100136426 | VTG | GTCTCTCTATGCCCCAAGCC | TCCACAGGTCTGTCCCTTCA |
| 100196216 | MFAP4 | GGCTAAAGTCCACGTCCAGT | CCGGCACCTCCATCTTTGAA |
| 106560553 | CFHL | ATGCCCAGTGATTCAAGCCA | CAGTAGCTACAGTTTACCTTCACA |
| 106560595 | TM4SF4 | CCATCCAGGTCATCAACGGT | AGCAAAAGGCCGTCAAGCTA |
| 106571136 | CLBLN2L | TTGGGAATTCAGGGAAGGCG | CCGGATTTTGGGTTTGCAGT |
| 106571178 | APOBL | TCCCCAGAAGATAGCCGACA | TGCAATGTTTTCTGCAGCCC |
| 106598656 | CFHR5L | TTGCCAATCTGGAGGATGGA | GACGACCCCAGTAATCCTTTTG |
| 106602980 | CFBL | AGAGGGAATCACCTGCAAGC | ACAGATTTACGGTGCCCCAG |
| 106604987 | C3 | TCGATTTGGTCGTCAAGCCA | GCAGGTCTTCAGACTTCCCC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jalili, M.; Gerdol, M.; Greco, S.; Pallavicini, A.; Buonocore, F.; Scapigliati, G.; Picchietti, S.; Esteban, M.A.; Rye, M.; Bones, A. Differential Effects of Dietary Supplementation of Krill Meal, Soybean Meal, Butyrate, and Bactocell® on the Gene Expression of Atlantic Salmon Head Kidney. Int. J. Mol. Sci. 2020, 21, 886. https://doi.org/10.3390/ijms21030886
Jalili M, Gerdol M, Greco S, Pallavicini A, Buonocore F, Scapigliati G, Picchietti S, Esteban MA, Rye M, Bones A. Differential Effects of Dietary Supplementation of Krill Meal, Soybean Meal, Butyrate, and Bactocell® on the Gene Expression of Atlantic Salmon Head Kidney. International Journal of Molecular Sciences. 2020; 21(3):886. https://doi.org/10.3390/ijms21030886
Chicago/Turabian StyleJalili, Mahsa, Marco Gerdol, Samuele Greco, Alberto Pallavicini, Francesco Buonocore, Giuseppe Scapigliati, Simona Picchietti, Maria Angeles Esteban, Morten Rye, and Atle Bones. 2020. "Differential Effects of Dietary Supplementation of Krill Meal, Soybean Meal, Butyrate, and Bactocell® on the Gene Expression of Atlantic Salmon Head Kidney" International Journal of Molecular Sciences 21, no. 3: 886. https://doi.org/10.3390/ijms21030886
APA StyleJalili, M., Gerdol, M., Greco, S., Pallavicini, A., Buonocore, F., Scapigliati, G., Picchietti, S., Esteban, M. A., Rye, M., & Bones, A. (2020). Differential Effects of Dietary Supplementation of Krill Meal, Soybean Meal, Butyrate, and Bactocell® on the Gene Expression of Atlantic Salmon Head Kidney. International Journal of Molecular Sciences, 21(3), 886. https://doi.org/10.3390/ijms21030886