The Autophagy Regulatory Molecule CSRP3 Interacts with LC3 and Protects Against Muscular Dystrophy


Abstract
1. Introduction
2. Results
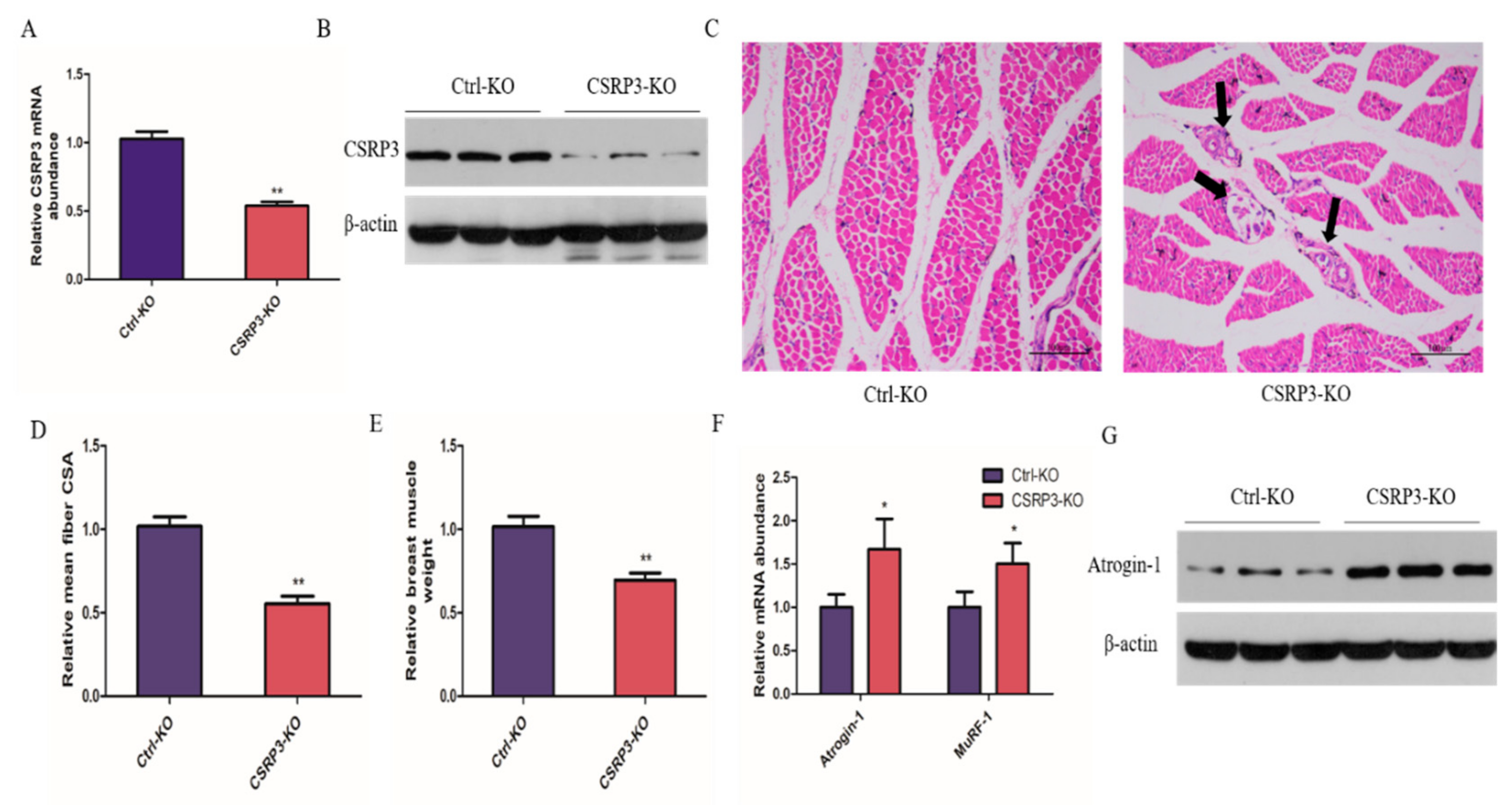
2.1. CSRP3 Silencing Results in Skeletal Muscle Atrophy
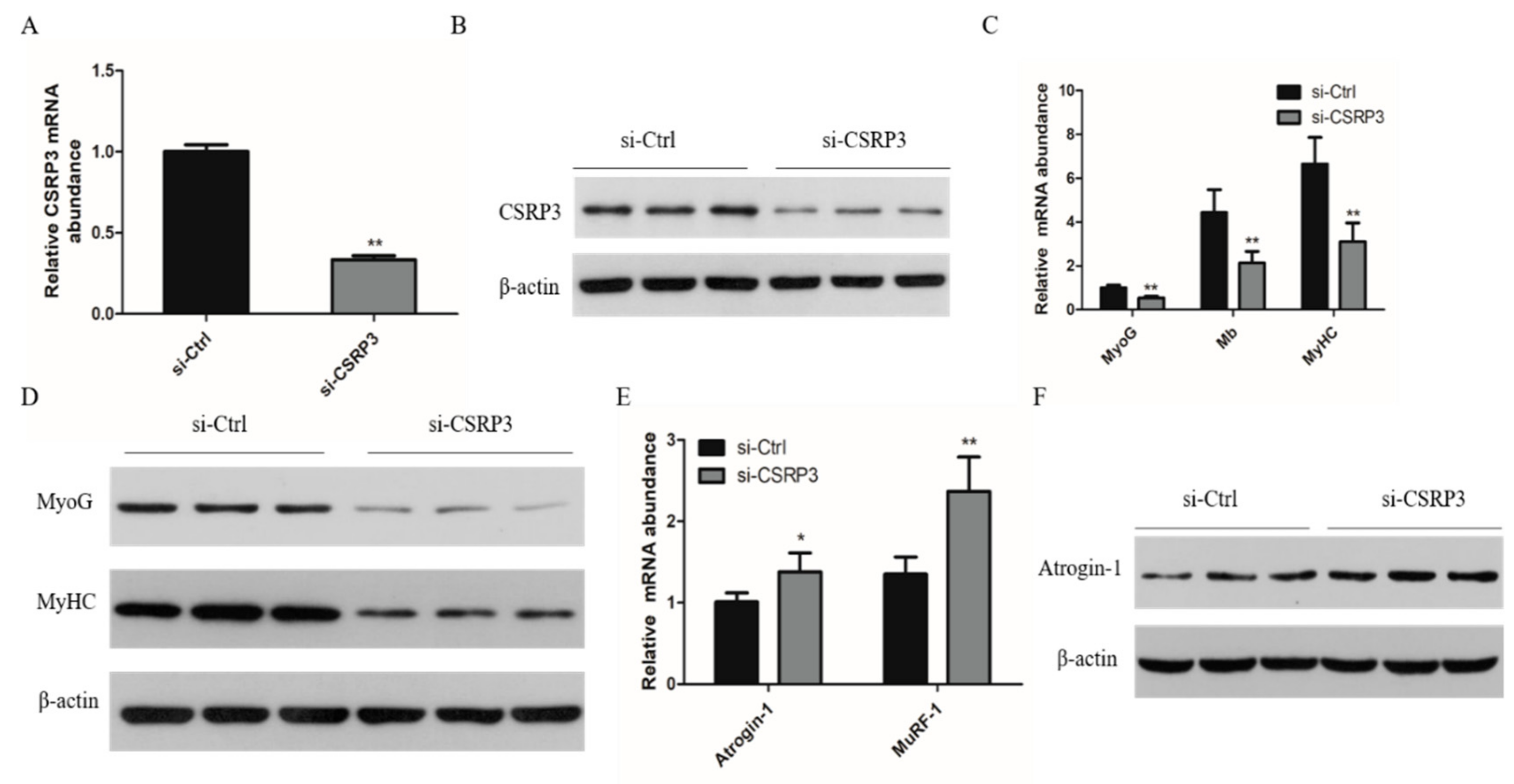
2.2. CSRP3 Regulates the Differentiation of Skeletal Muscle Cells
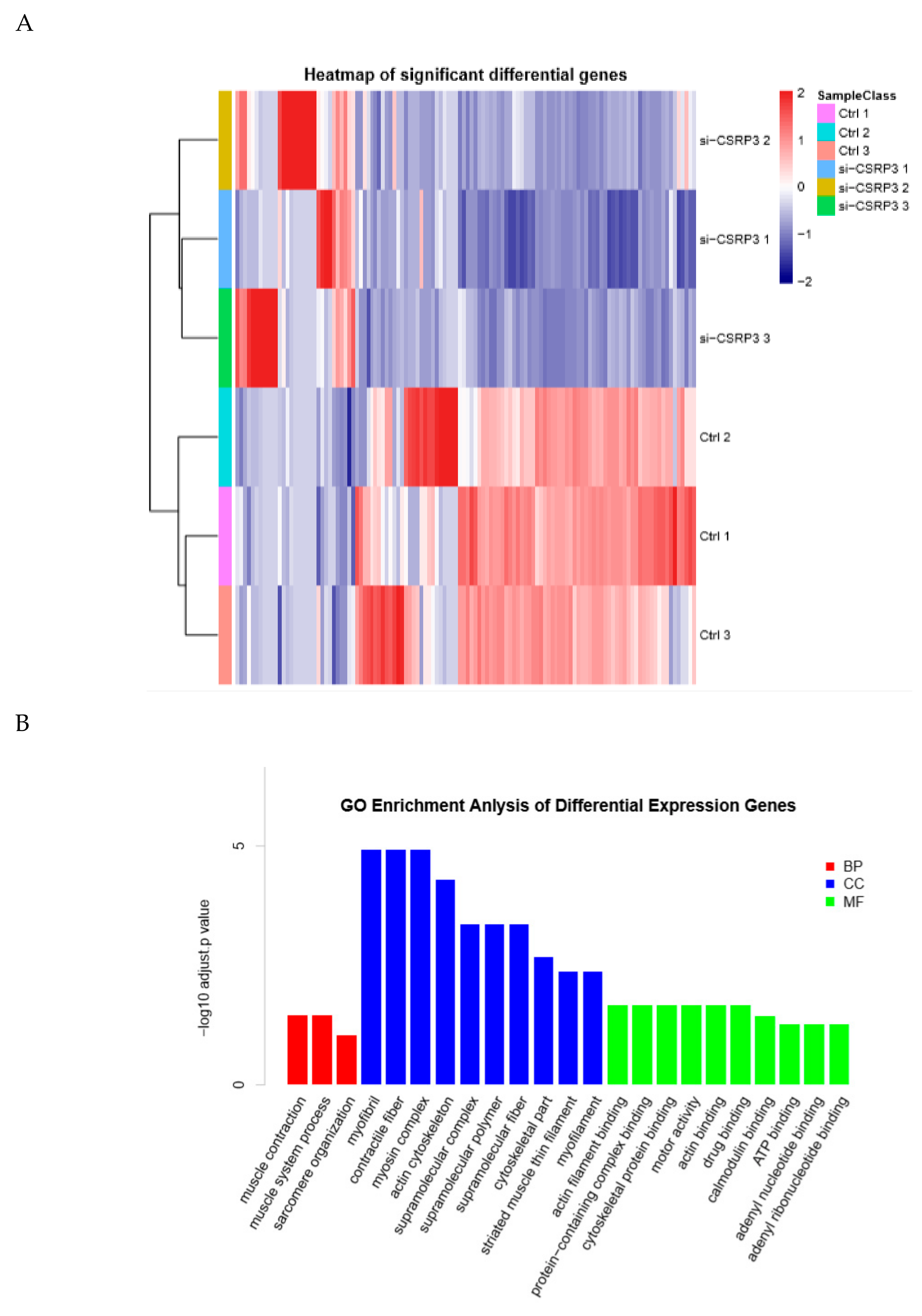
2.3. Gene Expression Analysis of CSRP3 Silenced Cells
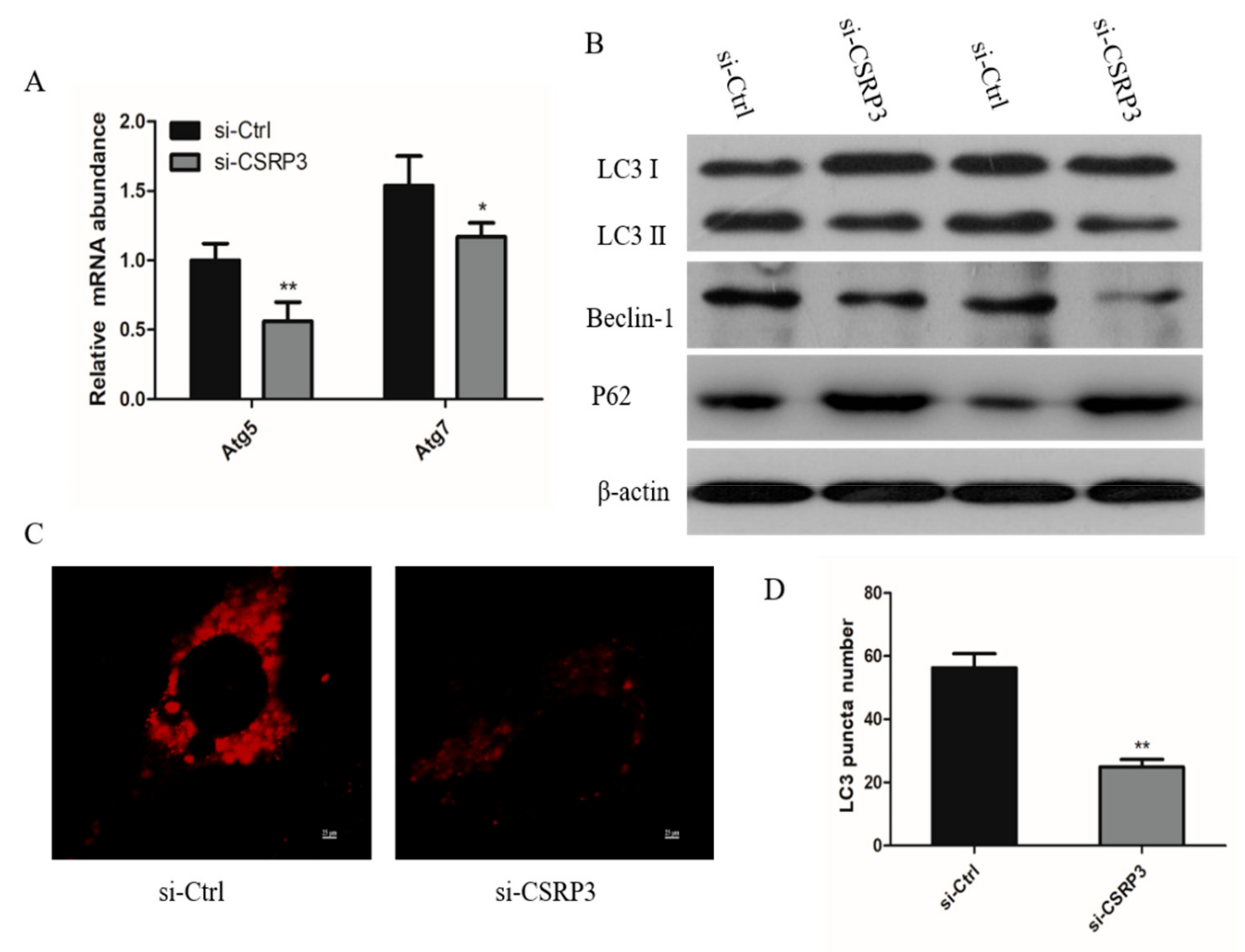
2.4. CSRP3 Silencing Results in Reduced Autophagy
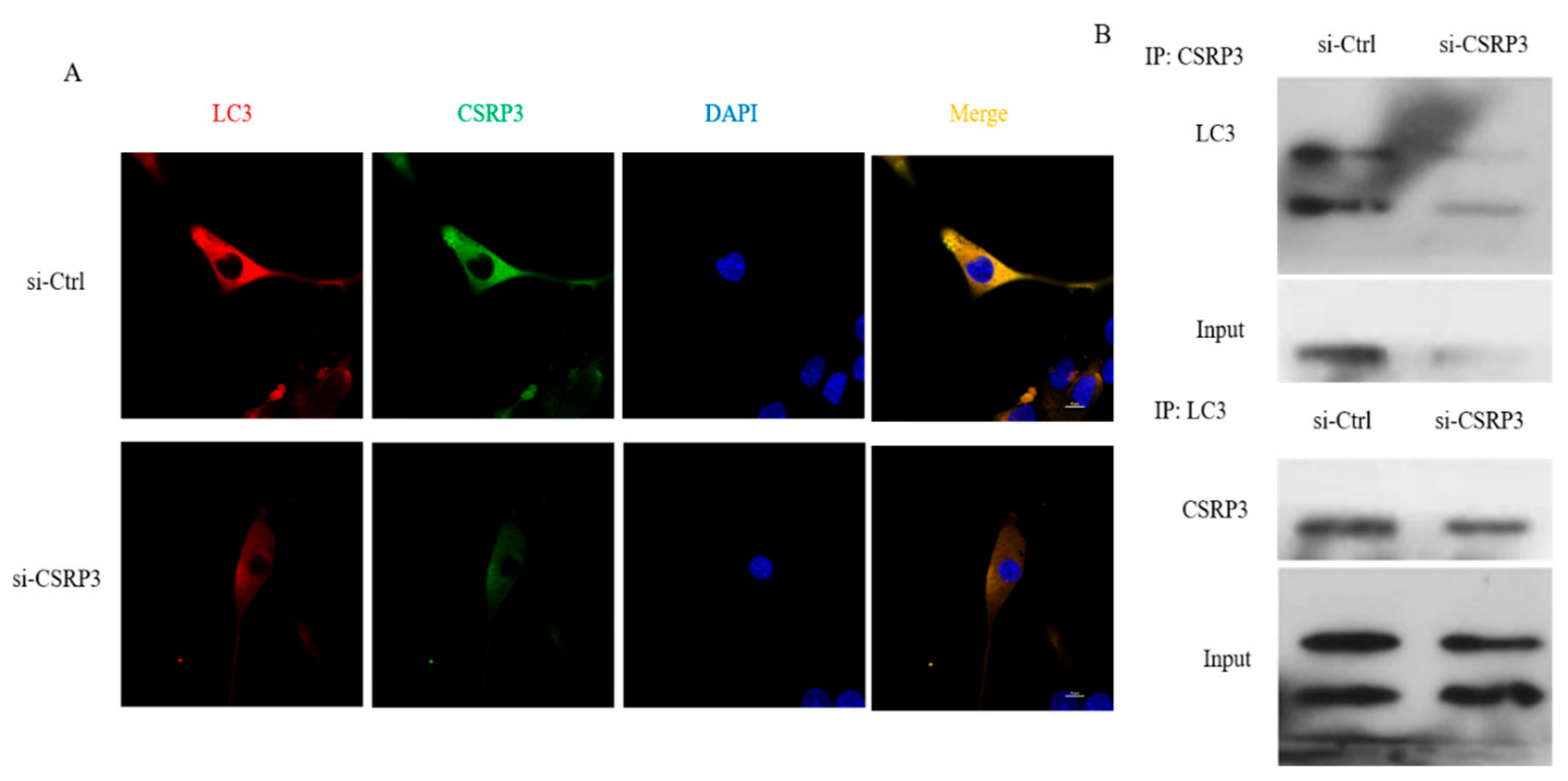
2.5. CSRP3 Interacts with LC3 to Regulate Autophagosome Formation
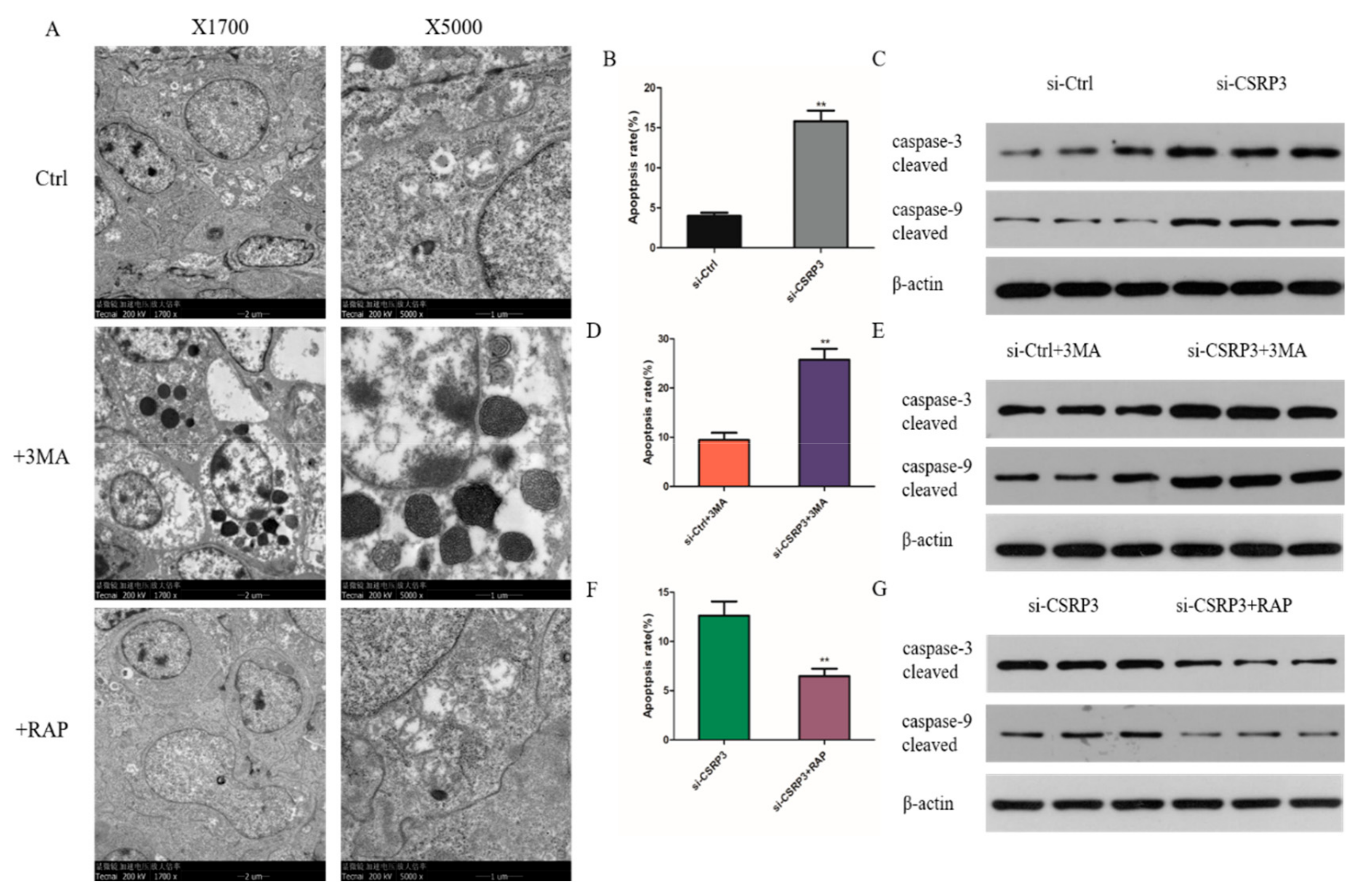
2.6. CSRP3 Protects Against Apoptosis by Regulating Autophagy
3. Discussion
4. Materials and Methods
4.1. Animal Procedures
4.2. Lentiviral Intramuscular Injections
4.3. Cell Cultures
4.4. CSRP3 Knockdown
4.5. Autophagy Treatment
4.6. RNA Extraction and Real-Time PCR (RT-PCR)
4.7. Western Blot and Immunoprecipitation Analysis
4.8. Histological and Morphometric Analysis
4.9. Immunofluorescence and Confocal Microscopy
4.10. Apoptosis Analysis
4.11. RNA Sequencing
4.12. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Lee, J.-K.; Cho, Y.-M.; Lee, J.-H. Association of Bovine CSRP3 and ACOX1 Genes with Carcass and Meat Quality Traits. Korean J. Agric. Sci. 2010, 37, 231–238. [Google Scholar]
- Oudot, C.; Gomes, A.; Nicolas, V.; Le Gall, M.; Chaffey, P.; Broussard, C.; Calamita, G.; Mastrodonato, M.; Gena, P.; Perfettini, J.-L. CSRP3 mediates polyphenols-induced cardioprotection in hypertension. J. Nutr. Biochem. 2019, 66, 29–42. [Google Scholar] [CrossRef]
- Gehmlich, K.; Geier, C.; Osterziel, K.J.; Van der Ven, P.F.; Fürst, D.O. Decreased interactions of mutant muscle LIM protein (MLP) with N-RAP and α-actinin and their implication for hypertrophic cardiomyopathy. Cell Tissue Res. 2004, 317, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Heineke, J.; Wollert, K.C.; Osinska, H.; Sargent, M.A.; York, A.J.; Robbins, J.; Molkentin, J.D. Calcineurin protects the heart in a murine model of dilated cardiomyopathy. J. Mol. Cell. Cardiol. 2010, 48, 1080–1087. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Papalouka, V.; Arvanitis, D.A.; Vafiadaki, E.; Mavroidis, M.; Papadodima, S.A.; Spiliopoulou, C.A.; Kremastinos, D.T.; Kranias, E.G.; Sanoudou, D. Muscle LIM protein interacts with cofilin 2 and regulates F-actin dynamics in cardiac and skeletal muscle. Mol. Cell. Biol. 2009, 29, 6046–6058. [Google Scholar] [CrossRef] [PubMed]
- Knöll, R.; Kostin, S.; Klede, S.; Savvatis, K.; Klinge, L.; Stehle, I.; Gunkel, S.; Kötter, S.; Babicz, K.; Sohns, M. A common MLP (muscle LIM protein) variant is associated with cardiomyopathy. Circ. Res. 2010, 106, 695. [Google Scholar] [CrossRef] [PubMed]
- Sanoudou, D.; Corbett, M.A.; Han, M.; Ghoddusi, M.; Nguyen, M.-A.T.; Vlahovich, N.; Hardeman, E.C.; Beggs, A.H. Skeletal muscle repair in a mouse model of nemaline myopathy. Hum. Mol. Genet. 2006, 15, 2603–2612. [Google Scholar] [CrossRef]
- Rabinowitz, J.D.; White, E. Autophagy and metabolism. Science 2010, 330, 1344–1348. [Google Scholar] [CrossRef]
- Sandri, M. Autophagy in skeletal muscle. FEBS Lett. 2010, 584, 1411–1416. [Google Scholar] [CrossRef]
- Sandri, M. Autophagy in health and disease. 3. Involvement of autophagy in muscle atrophy. Am. J. Physiol. Cell Physiol. 2010, 298, C1291–C1297. [Google Scholar] [CrossRef]
- Neel, B.A.; Lin, Y.; Pessin, J.E. Skeletal muscle autophagy: A new metabolic regulator. Trends Endocrinol. Metab. 2013, 24, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Bach, I. The LIM domain: Regulation by association. Mech. Dev. 2000, 91, 5–17. [Google Scholar] [CrossRef]
- Kadrmas, J.L.; Beckerle, M.C. The LIM domain: From the cytoskeleton to the nucleus. Nat. Rev. Mol. Cell Biol. 2004, 5, 920. [Google Scholar] [CrossRef] [PubMed]
- Tobias, E.; Hurlstone, A.; MacKenzie, E.; McFarlane, R.; Black, D. The TES gene at 7q31. 1 is methylated in tumours and encodes a novel growth-suppressing LIM domain protein. Oncogene 2001, 20, 2844. [Google Scholar] [CrossRef]
- Tao, Y.; Wang, J.; Tokusumi, T.; Gajewski, K.; Schulz, R.A. Requirement of the LIM homeodomain transcription factor tailup for normal heart and hematopoietic organ formation in Drosophila melanogaster. Mol. Cell. Biol. 2007, 27, 3962–3969. [Google Scholar] [CrossRef]
- Feuerstein, R.; Wang, X.; Song, D.; Cooke, N.E.; Liebhaber, S.A. The LIM/double zinc-finger motif functions as a protein dimerization domain. Proc. Natl. Acad. Sci. 1994, 91, 10655–10659. [Google Scholar] [CrossRef]
- Sabatelli, P.; Castagnaro, S.; Tagliavini, F.; Chrisam, M.; Sardone, F.; Demay, L.; Richard, P.; Santi, S.; Maraldi, N.M.; Merlini, L. Aggresome–autophagy involvement in a sarcopenic patient with rigid spine syndrome and a p. C150R mutation in FHL1 gene. Front. Aging Neurosci. 2014, 6, 215. [Google Scholar] [CrossRef]
- Xia, W.-R.; Fu, W.; Wang, Q.; Zhu, X.; Xing, W.-W.; Wang, M.; Xu, D.-Q.; Xu, D.-G. Autophagy Induced FHL2 Upregulation Promotes IL-6 Production by Activating the NF-κB Pathway in Mouse Aortic Endothelial Cells after Exposure to PM2.5. Int. J. Mol. Sci. 2017, 18, 1484. [Google Scholar] [CrossRef]
- Shang, Y.; Wang, H.; Jia, P.; Zhao, H.; Liu, C.; Liu, W.; Song, Z.; Xu, Z.; Yang, L.; Wang, Y. Autophagy regulates spermatid differentiation via degradation of PDLIM1. Autophagy 2016, 12, 1575–1592. [Google Scholar] [CrossRef]
- Rashid, M.; Runci, A.; Polletta, L.; Carnevale, I.; Morgante, E.; Foglio, E.; Arcangeli, T.; Sansone, L.; Russo, M.; Tafani, M. Muscle LIM protein/CSRP3: A mechanosensor with a role in autophagy. Cell Death Discov. 2015, 1, 15014. [Google Scholar] [CrossRef]
- Flick, M.J.; Konieczny, S.F. The muscle regulatory and structural protein MLP is a cytoskeletal binding partner of betaI-spectrin. J. Cell Sci. 2000, 113, 1553–1564. [Google Scholar] [PubMed]
- Arber, S.; Hunter, J.J.; Ross Jr, J.; Hongo, M.; Sansig, G.; Borg, J.; Perriard, J.-C.; Chien, K.R.; Caroni, P. MLP-deficient mice exhibit a disruption of cardiac cytoarchitectural organization, dilated cardiomyopathy, and heart failure. Cell 1997, 88, 393–403. [Google Scholar] [CrossRef]
- Kong, Y.; Flick, M.J.; Kudla, A.J.; Konieczny, S.F. Muscle LIM protein promotes myogenesis by enhancing the activity of MyoD. Mol. Cell Biol. 1997, 17, 4750–4760. [Google Scholar] [CrossRef] [PubMed]
- Buyandelger, B.; Ng, K.-E.; Miocic, S.; Piotrowska, I.; Gunkel, S.; Ku, C.-H.; Knöll, R. MLP (muscle LIM protein) as a stress sensor in the heart. Pflügers Arch. 2011, 462, 135–142. [Google Scholar] [CrossRef]
- Barash, I.A.; Mathew, L.; Lahey, M.; Greaser, M.L.; Lieber, R.L. Muscle LIM protein plays both structural and functional roles in skeletal muscle. Am. J. Physiol. Cell Physiol. 2005, 289, C1312–C1320. [Google Scholar] [CrossRef]
- Jokl, E.J.; Blanco, G. Disrupted autophagy undermines skeletal muscle adaptation and integrity. Mamm. Genome 2016, 27, 525–537. [Google Scholar] [CrossRef]
- Funderburk, S.F.; Wang, Q.J.; Yue, Z. The Beclin 1–VPS34 complex–at the crossroads of autophagy and beyond. Trends Cell Biol. 2010, 20, 355–362. [Google Scholar] [CrossRef]
- Hussain, S.N.; Sandri, M. Role of autophagy in COPD skeletal muscle dysfunction. J. Appl. Physiol. 2012, 114, 1273–1281. [Google Scholar] [CrossRef]
- Codogno, P.; Meijer, A.J. Atg5: More than an autophagy factor. Nat. Cell Biol. 2006, 8, 1045. [Google Scholar] [CrossRef]
- Komatsu, M.; Waguri, S.; Ueno, T.; Iwata, J.; Murata, S.; Tanida, I.; Ezaki, J.; Mizushima, N.; Ohsumi, Y.; Uchiyama, Y. Impairment of starvation-induced and constitutive autophagy in Atg7-deficient mice. J. Cell Biol. 2005, 169, 425–434. [Google Scholar] [CrossRef]
- Vicencio, J.M.; Galluzzi, L.; Tajeddine, N.; Ortiz, C.; Criollo, A.; Tasdemir, E.; Morselli, E.; Younes, A.B.; Maiuri, M.C.; Lavandero, S. Senescence, apoptosis or autophagy? Gerontology 2008, 54, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Brill, A.; Torchinsky, A.; Carp, H.; Toder, V. The role of apoptosis in normal and abnormal embryonic development. J. Assist. Reprod. Genet. 1999, 16, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, J.; Yeh, E.S. Autophagy and apoptotic crosstalk: Mechanism of therapeutic resistance in HER2-positive breast cancer. Breast Cancer: Basic Clin. Res. 2016, 10, S32791. [Google Scholar] [CrossRef] [PubMed]
- Thorburn, A. Apoptosis and autophagy: Regulatory connections between two supposedly different processes. Apoptosis 2008, 13, 1–9. [Google Scholar] [CrossRef]
- Luo, S.; Rubinsztein, D. Atg5 and Bcl-2 provide novel insights into the interplay between apoptosis and autophagy. Cell Death Differ. 2007, 14, 1247–1250. [Google Scholar] [CrossRef]
- McMillan, E.M.; Quadrilatero, J. Autophagy is required and protects against apoptosis during myoblast differentiation. Biochem. J. 2014, 462, 267–277. [Google Scholar] [CrossRef]
- Maiuri, M.C.; Zalckvar, E.; Kimchi, A.; Kroemer, G. Self-eating and self-killing: Crosstalk between autophagy and apoptosis. Nat. Rev. Mol. Cell Biol. 2007, 8, 741. [Google Scholar] [CrossRef]
- Lekli, I.; Haines, D.D.; Balla, G.; Tosaki, A. Autophagy: An adaptive physiological countermeasure to cellular senescence and ischaemia/reperfusion-associated cardiac arrhythmias. J. Cell. Mol. Med. 2017, 21, 1058–1072. [Google Scholar] [CrossRef]
- Czegledi, A.; Tosaki, A.; Gyongyosi, A.; Zilinyi, R.; Tosaki, A.; Lekli, I. Electrically-Induced Ventricular Fibrillation Alters Cardiovascular Function and Expression of Apoptotic and Autophagic Proteins in Rat Hearts. Int. J. Mol. Sci. 2019, 20, 1628. [Google Scholar] [CrossRef]
- Chang, K.; Kang, P.; Liu, Y.; Huang, K.; Miao, T.; Sagona, A.P.; Nezis, I.P.; Bodmer, R.; Ocorr, K.; Bai, H. TGFB-INHB/activin signaling regulates age-dependent autophagy and cardiac health through inhibition of MTORC2. Autophagy 2019. [Google Scholar] [CrossRef]
- Ullrich, M.; Aßmus, B.; Augustin, A.M.; Häbich, H.; Abeßer, M.; Machado, J.M.; Werner, F.; Erkens, R.; Arias-Loza, A.-P.; Umbenhauer, S. SPRED2 deficiency elicits cardiac arrhythmias and premature death via impaired autophagy. J. Mol. Cell. Cardiol. 2019, 129, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Meyer, G.; Czompa, A.; Reboul, C.; Csepanyi, E.; Czegledi, A.; Bak, I.; Balla, G.; Balla, J.; Tosaki, A.; Lekli, I. The cellular autophagy markers Beclin-1 and LC3B-II are increased during reperfusion in fibrillated mouse hearts. Curr. Pharm. Des. 2013, 19, 6912–6918. [Google Scholar] [CrossRef]
- Mocanu, M.; Baxter, G.; Yellon, D. Caspase inhibition and limitation of myocardial infarct size: Protection against lethal reperfusion injury. Br. J. Pharmacol. 2000, 130, 197–200. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, P.; Bak, I.; Szendrei, L.; Vecsernyes, M.; Varga, E.; Blasig, I.E.; Tosaki, A. Non-specific caspase inhibition reduces infarct size and improves post-ischaemic recovery in isolated ischaemic/reperfused rat hearts. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2001, 364, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Yablonka–Reuveni, Z.; Paterson, B.M. MyoD and myogenin expression patterns in cultures of fetal and adult chicken myoblasts. J. Histochem. Cytochem. 2001, 49, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Chen, J.; Li, L.; Ren, X.; Cheng, T.; Lu, S.; Lawal, R.A.; Nie, Q.; Zhang, X.; Hanotte, O. c-Myc inhibits myoblast differentiation and promotes myoblast proliferation and muscle fibre hypertrophy by regulating the expression of its target genes, miRNAs and lincRNAs. Cell Death Differ. 2019, 26, 426. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| CSRP3 | CCCTCCACACCAACTAACCC | TCTGCAGCGTACACCGATTT |
| MyoG | CGGAGGCTGAAGAAGGTGAA | CGGTCCTCTGCCTGGTCAT |
| Mb | CCCTGAGACTTTGGATCGCTT | CTGGGATTTTGTGCTTCGTGG |
| MyHC | CTCCTCACGCTTTGGTAA | TGATAGTCGTATGGGTTGGT |
| Atrogin-1 | TCAACGGGTCGGCAAGTCT | TCCCTCCCATCGCTCAGTC |
| MuRF-1 | GGCAGCAGCATCATCTCGG | CCTCGCAGGTGACGCAGTAG |
| Atg5 | GATGAAATAACTGAAAGGGAAGC | TGAAGATCAAAGAGCAAACCAA |
| Atg7 | TCAGATTCAAGCACTTCAGA | GAGGAGATACAACCACAGAG |
| β-actin | CCGCTCTATGAAGGCTACGC | CTCTCGGCTGTGGTGGTGAA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, C.; Han, S.; Tang, S.; He, H.; Shen, X.; Zhao, J.; Chen, Y.; Wei, Y.; Wang, Y.; Zhu, Q.; et al. The Autophagy Regulatory Molecule CSRP3 Interacts with LC3 and Protects Against Muscular Dystrophy. Int. J. Mol. Sci. 2020, 21, 749. https://doi.org/10.3390/ijms21030749
Cui C, Han S, Tang S, He H, Shen X, Zhao J, Chen Y, Wei Y, Wang Y, Zhu Q, et al. The Autophagy Regulatory Molecule CSRP3 Interacts with LC3 and Protects Against Muscular Dystrophy. International Journal of Molecular Sciences. 2020; 21(3):749. https://doi.org/10.3390/ijms21030749
Chicago/Turabian StyleCui, Can, Shunshun Han, Shuyue Tang, Haorong He, Xiaoxu Shen, Jing Zhao, Yuqi Chen, Yuanhang Wei, Yan Wang, Qing Zhu, and et al. 2020. "The Autophagy Regulatory Molecule CSRP3 Interacts with LC3 and Protects Against Muscular Dystrophy" International Journal of Molecular Sciences 21, no. 3: 749. https://doi.org/10.3390/ijms21030749
APA StyleCui, C., Han, S., Tang, S., He, H., Shen, X., Zhao, J., Chen, Y., Wei, Y., Wang, Y., Zhu, Q., Li, D., & Yin, H. (2020). The Autophagy Regulatory Molecule CSRP3 Interacts with LC3 and Protects Against Muscular Dystrophy. International Journal of Molecular Sciences, 21(3), 749. https://doi.org/10.3390/ijms21030749