MicroRNA 322 Aggravates Dexamethasone-Induced Muscle Atrophy by Targeting IGF1R and INSR


Abstract
1. Introduction
2. Results
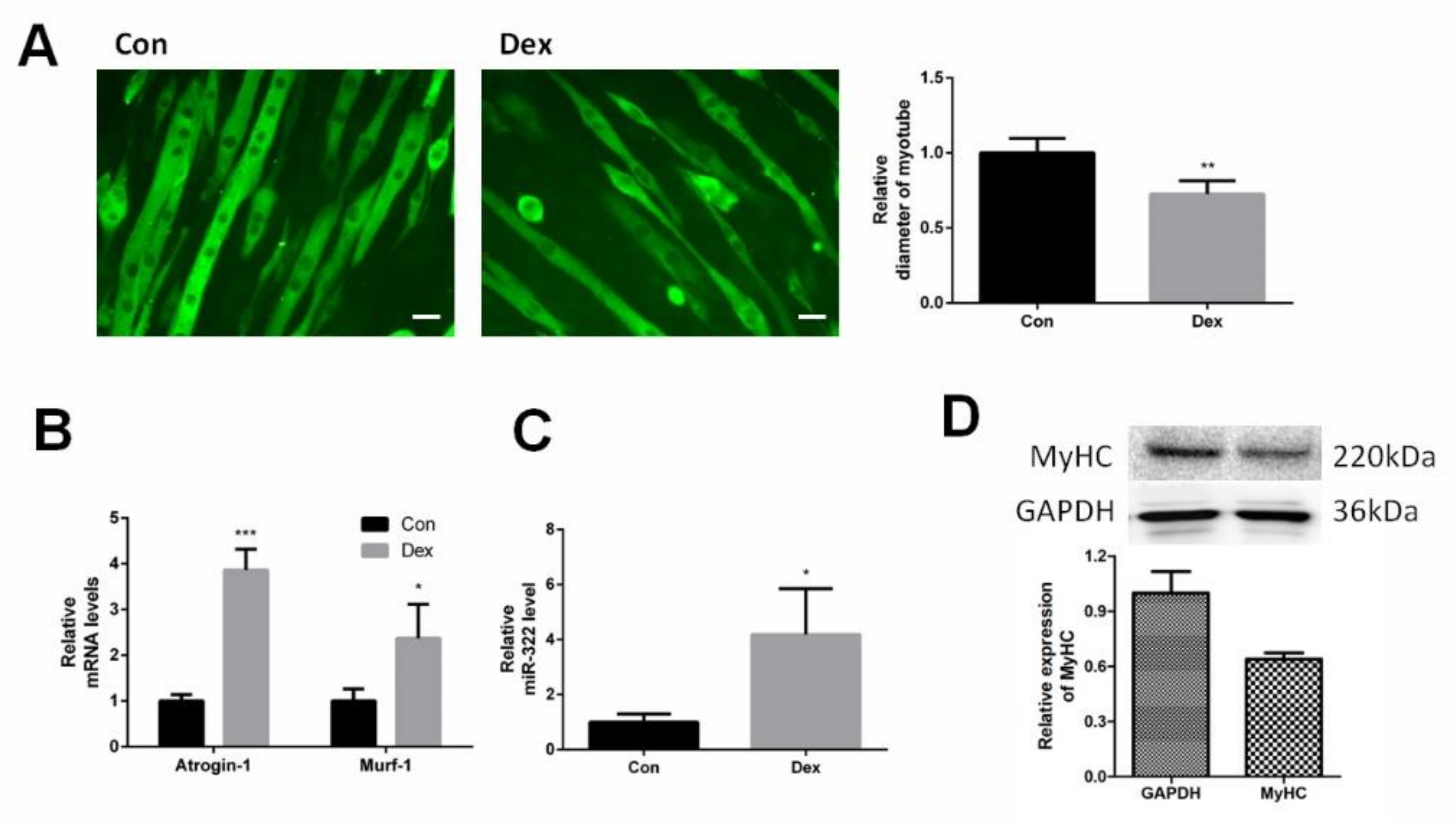
2.1. miR-322 Increased in Dex-Induced C2C12 Myotubes
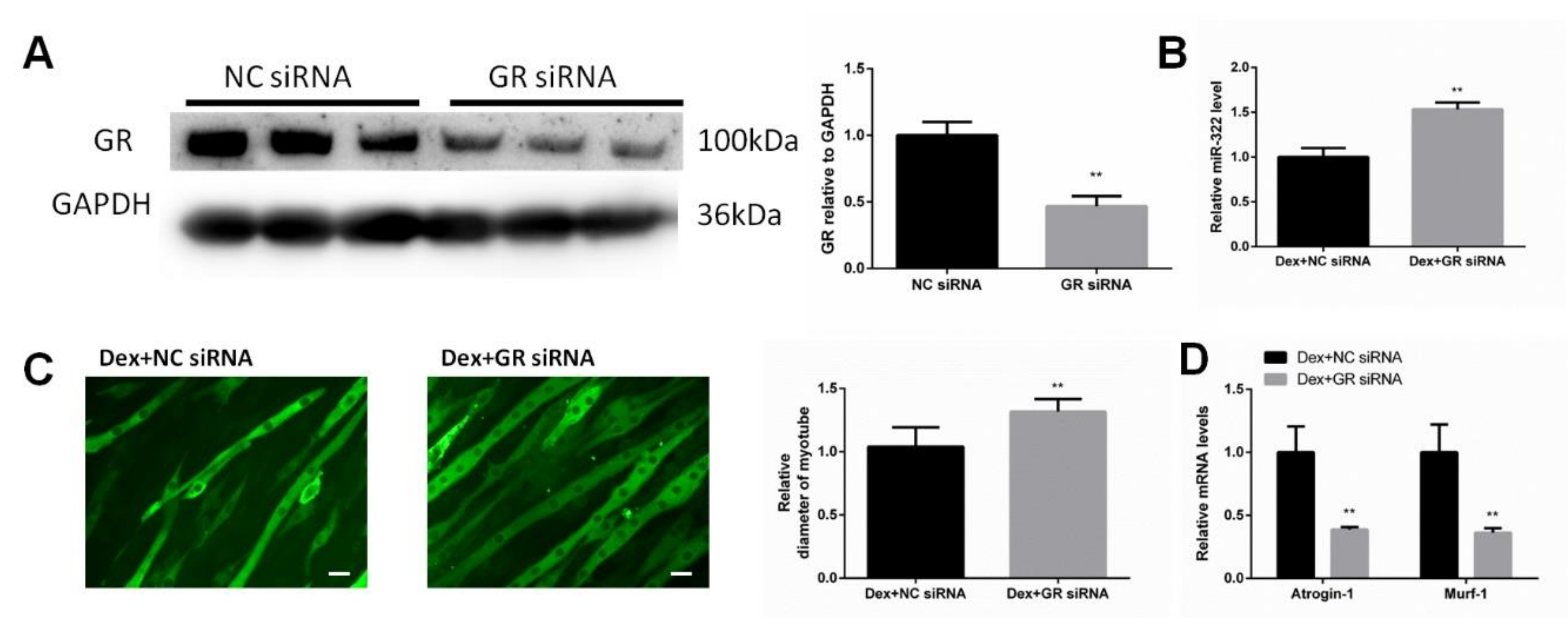
2.2. Dex Required GR to Increase the miR-322 Expression in C2C12 Myotubes
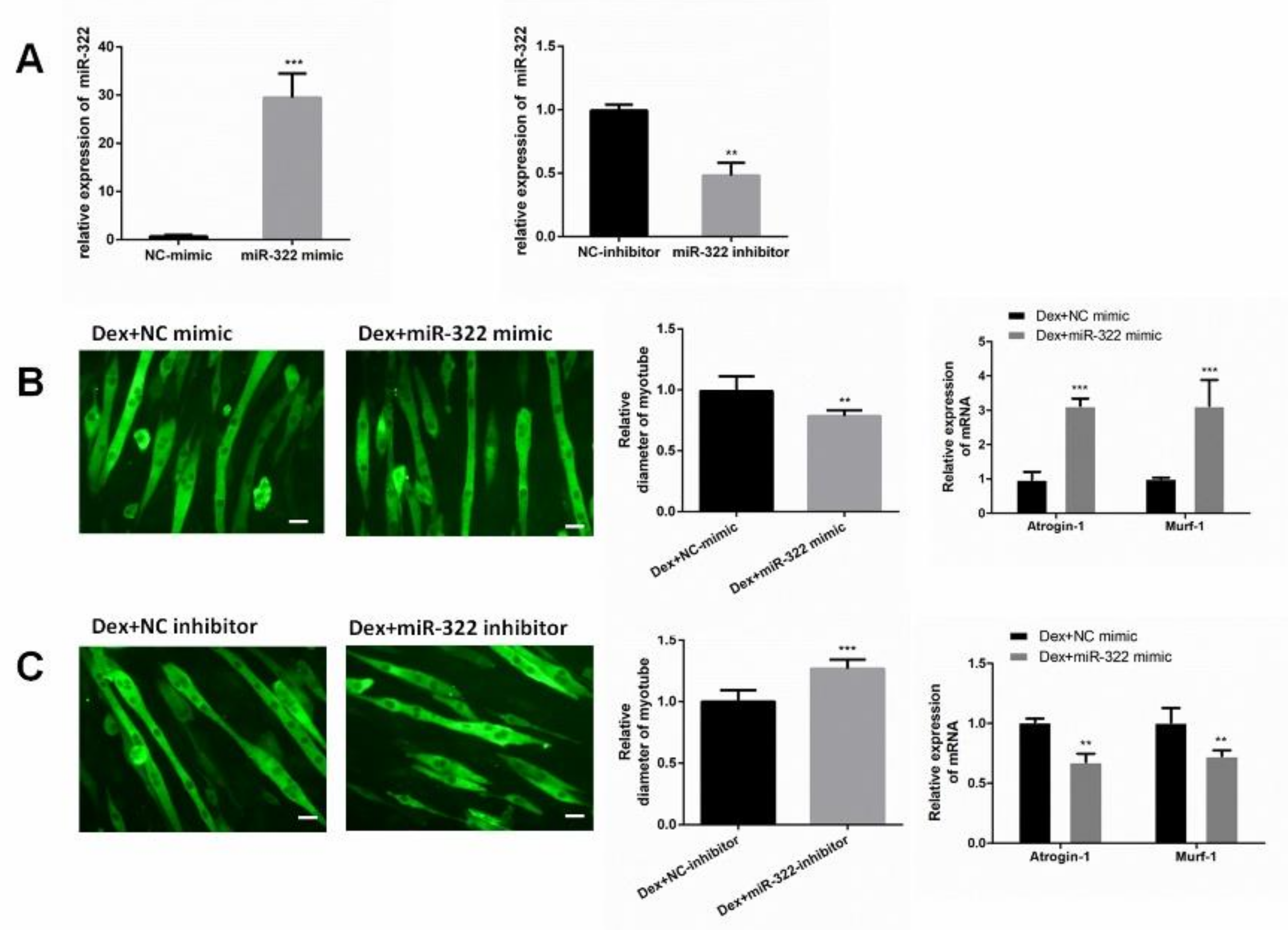
2.3. miR-322 Aggravated Dex-Induced Atrophy in C2C12 Myotubes
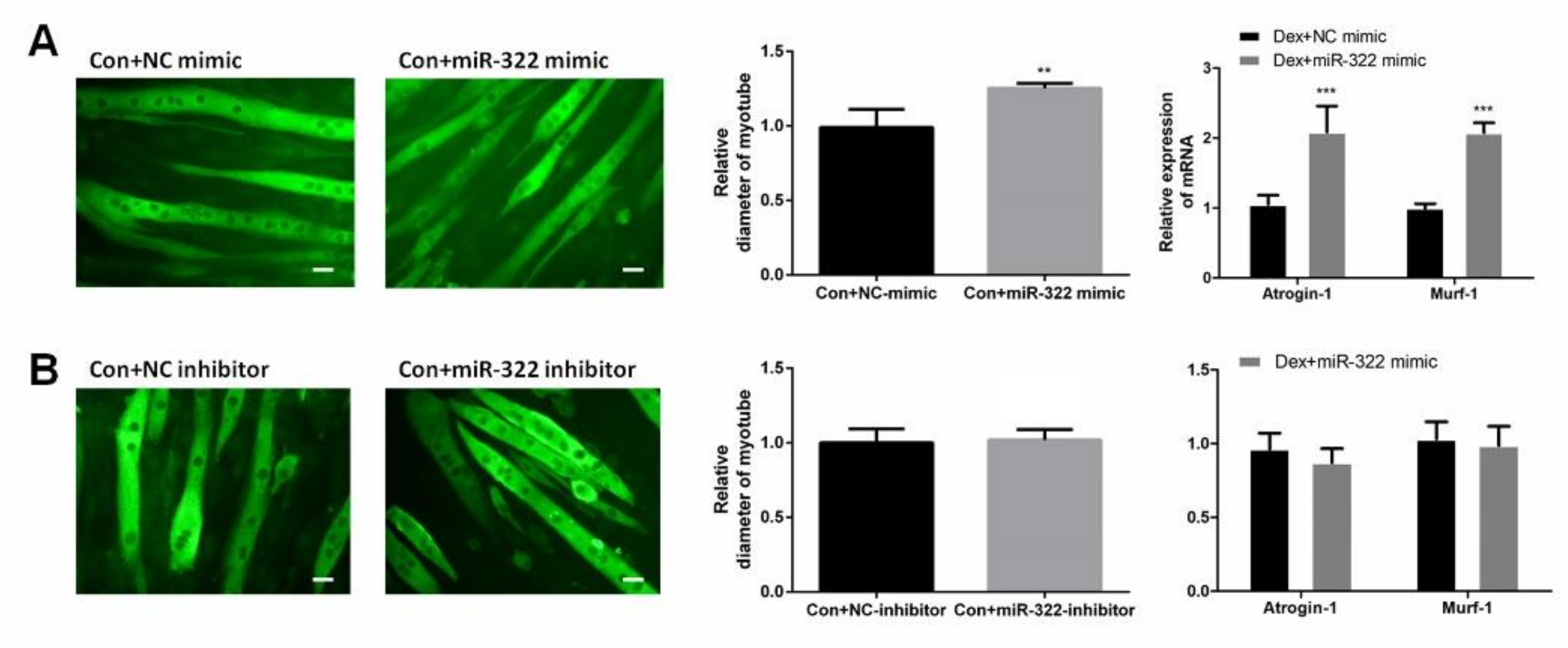
2.4. miR-322 Induced Muscle Atrophy In Vitro Without Dex-Treated
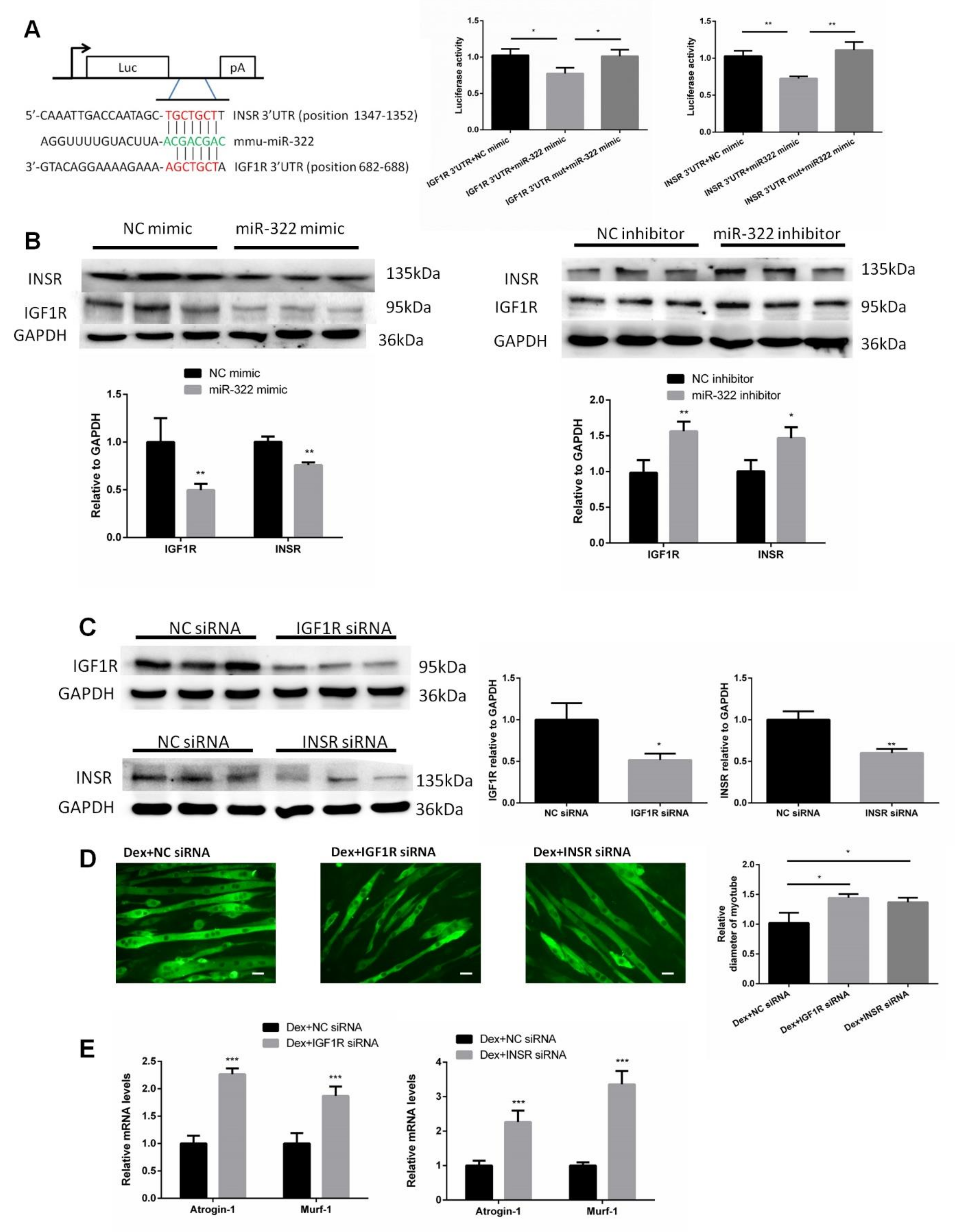
2.5. IGF1R and INSR Are Target Genes of miR-322
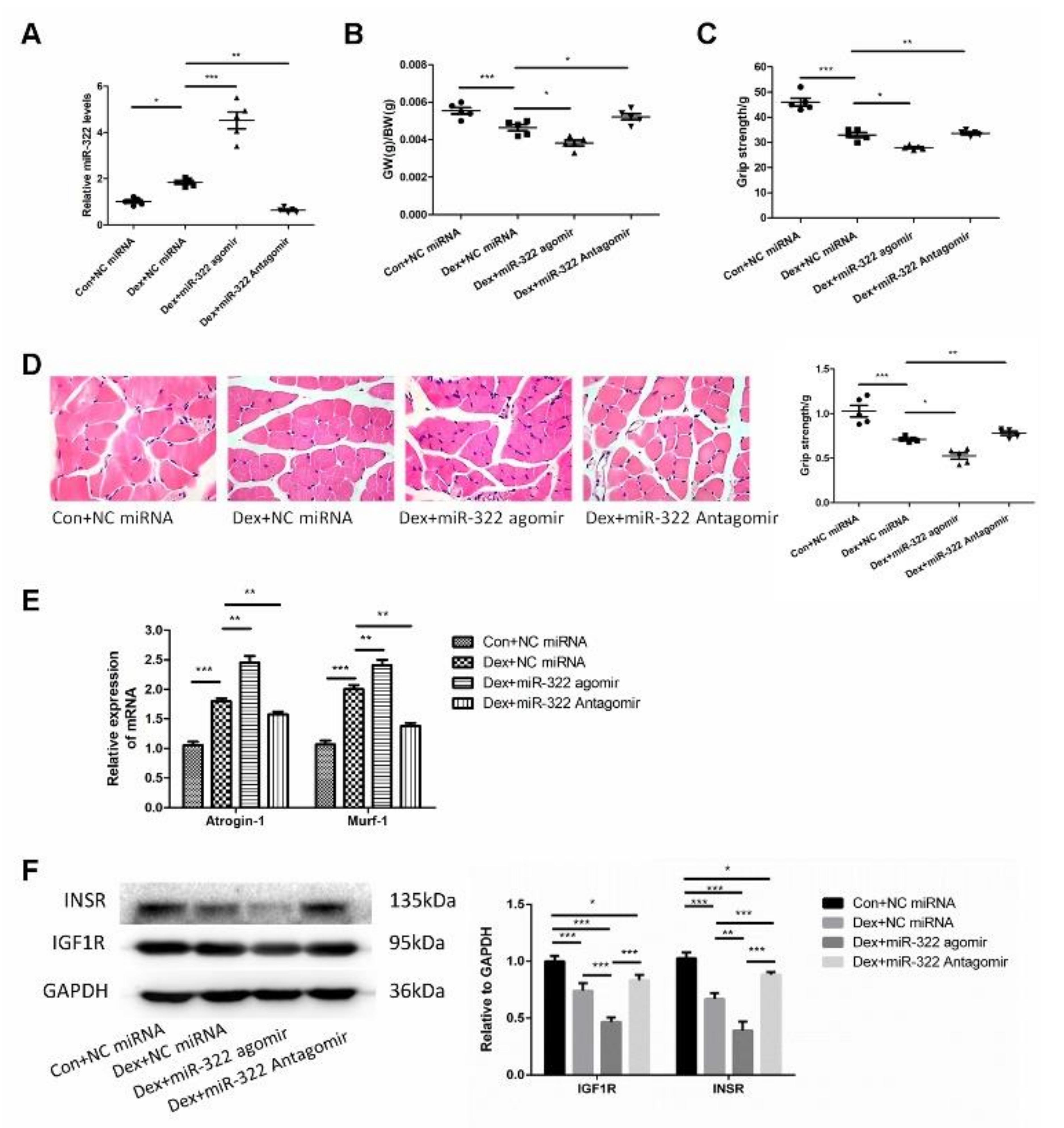
2.6. miR-322 Contributed to Dex-Induced Muscle Atrophy In Vivo
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Transfection
4.2. Luciferase Reporter Assays
4.3. Animal Experiments
4.4. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
4.5. Western Blot
4.6. Staining
4.7. Grip-Strength Test
4.8. Statistical Analysis
Supplementary Materials
Supplementary File 1Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Dex | Dexamethasone |
| IGF1R | Insulin-like growth factor 1 receptor |
| INSR | Insulin receptor |
| GR | Glucocorticoid receptor |
| HE | Hematoxylin eosin |
| WT | Wild-type |
| MyHC | Myosin heavy chain |
References
- Zelena, D.; Makara, B.G. Steroids: The physiologic and pharmacologic effects of glucocorticoids. Orv. Hetil. 2015, 156, 1415–1425. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Behrend, E.N.; Kemppainen, R.J. Glucocorticoid therapy: Pharmacology, indications, and complications. Vet. Clin. N. Am. Small Anim. Pract. 1997, 27, 187. [Google Scholar] [CrossRef]
- Malkawi, A.K.; Alzoubi, K.H.; Jacob, M.; Matic, G.; Ali, A.; Al Faraj, A.; Almuhanna, F.; Dasouki, M.; Abdel Rahman, A.M. Metabolomics Based Profiling of Dexamethasone Side Effects in Rats. Front. Pharmacol. 2018, 9, 46. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, M.A.; Zunszain, P.A.; Anacker, C.; Musaelyan, K.; Pariante, C.M. Glucocorticoids and inflammation: A double-headed sword in depression? How do neuroendocrine and inflammatory pathways interact during stress to contribute to the pathogenesis of depression? Mod. Trends Pharm. 2013, 28, 127. [Google Scholar]
- Ebert, S.M.; Dyle, M.C.; Kunkel, S.D.; Bullard, S.A.; Bongers, K.S.; Fox, D.K.; Dierdorff, J.M.; Foster, E.D.; Adams, C.M. Stress-induced skeletal muscle Gadd45a expression reprograms myonuclei and causes muscle atrophy. J. Biol. Chem. 2012, 287, 27290–27301. [Google Scholar] [CrossRef]
- Dutt, V.; Gupta, S.; Dabur, R.; Injeti, E.; Mittal, A. Skeletal muscle atrophy: Potential therapeutic agents and their mechanisms of action. Pharmacol. Res. 2015, 99, 86–100. [Google Scholar] [CrossRef]
- Löfberg, E.; Gutierrez, A.; Wernerman, J.; Anderstam, B.; Mitch, W.E.; Price, S.R.; Bergström, J.; Alvestrand, A. Effects of high doses of glucocorticoids on free amino acids, ribosomes and protein turnover in human muscle. Eur. J. Clin. Investig. 2015, 32, 345–353. [Google Scholar] [CrossRef]
- Thissen, J.P.; Schakman, O.; Gilson, H. Mechanisms of glucocorticoid-induced myopathy. Bone 2009, 45, S123–S124. [Google Scholar] [CrossRef]
- Sun, H.; Gong, Y.; Qiu, J.; Chen, Y.; Ding, F.; Zhao, Q. TRAF6 inhibition rescues dexamethasone-induced muscle atrophy. Int. J. Mol. Sci. 2014, 15, 11126–11141. [Google Scholar] [CrossRef]
- Li, J.; Chan, M.C.; Yu, Y.; Bei, Y.; Chen, P.; Zhou, Q.; Cheng, L.; Chen, L.; Ziegler, O.; Rowe, G.C.; et al. miR-29b contributes to multiple types of muscle atrophy. Nat. Commun. 2017, 8, 15201. [Google Scholar] [CrossRef]
- Wang, X.H.; Mitch, W.E. Muscle wasting from kidney failure—A model for catabolic conditions. Int. J. Biochem. Cell Biol. 2013, 45, 2230–2238. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Mitch, W.E. Mechanisms of muscle wasting in chronic kidney disease. Nat. Rev. Nephrol. 2014, 10, 504. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.; Dai, G.; Hu, Z.; Wang, X.; Du, J.; Mitch, W.E. Regulation of muscle protein degradation: Coordinated control of apoptotic and ubiquitin-proteasome systems by phosphatidylinositol 3 kinase. J. Am. Soc. Nephrol. 2004, 15, 1537–1545. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, X.H.; Wang, H.; Du, J.; Mitch, W.E. Satellite cell dysfunction and impaired IGF-1 signaling cause CKD-induced muscle atrophy. J. Am. Soc. Nephrol. 2010, 21, 419. [Google Scholar] [CrossRef]
- Sandri, M.; Sandri, C.; Gilbert, A.; Skurk, C.; Calabria, E.; Picard, A.; Walsh, K.; Schiaffino, S.; Lecker, S.H.; Goldberg, A.L. Foxo transcription factors induce the atrophy-related ubiquitin ligase atrogin-1 and cause skeletal muscle atrophy. Cell 2004, 117, 399–412. [Google Scholar] [CrossRef]
- Ivey, K.N.; Srivastava, D. microRNAs as Developmental Regulators. Cold Spring Harb. Perspect. Biol. 2015, 7, a008144. [Google Scholar] [CrossRef] [PubMed]
- Horak, M.; Novak, J.; Bienertova-Vasku, J. Muscle-specific microRNAs in skeletal muscle development. Dev. Biol. 2016, 410, 1–13. [Google Scholar] [CrossRef]
- Moresi, V.; Marroncelli, N.; Coletti, D.; Adamo, S. Regulation of skeletal muscle development and homeostasis by gene imprinting, histone acetylation and microRNA. Biochim. Biophys. Acta 2015, 1849, 309–316. [Google Scholar] [CrossRef]
- Anja, K.; Tadeja, R.E.; Boris, R. MicroRNA in skeletal muscle development, growth, atrophy, and disease. Wiley Interdiscip. Rev. RNA 1900, 5, 509–525. [Google Scholar]
- Shogo, W.; Yoshio, K.; Mitsuharu, O.; Shigeru, M.; Katsuhiko, S.; Zhen, Y.; Stefano, S.; Hiroshi, A.; Takashi, U.; Takayuki, A. Translational suppression of atrophic regulators by microRNA-23a integrates resistance to skeletal muscle atrophy. J. Biol. Chem. 2011, 286, 38456. [Google Scholar]
- Matthew, B.H.; Myra, E.W.-H.; Bin, Z.; Jill, A.R.; Mitsi, A.B.; Jennifer, L.G.; Charles, D.S.; SRuss, P. miR-23a is decreased during muscle atrophy by a mechanism that includes calcineurin signaling and exosome-mediated export. Am. J. Physiol. Cell Physiol. 2014, 306, C551. [Google Scholar]
- He, S.; Teng, L.; Liangliang, F.; Shuhong, Z.; Bin, F.; Jianhua, C.; Xinyun, L. Identification of microRNAs involved in dexamethasone-induced muscle atrophy. Mol. Cell. Biochem. 2013, 381, 105–113. [Google Scholar]
- Fan, W.; Rui, L.; Neha, T.; Elizabeth, R.M.; Shreesti, S.; Jiao, Y.; Benjamin, S.; Jin, Y.; Yu, L. H19X-encoded miR-424(322)/-503 cluster: Emerging roles in cell differentiation, proliferation, plasticity and metabolism. Cell. Mol. Life Sci. 2019, 76, 903–920. [Google Scholar]
- Alexandre, M.; Fabrice, A.; Nathalie, M.; Michel, C.; Veronica, C.; Jeremy, B.; Carole, P.; David-Alexandre, T.; Jean-Sébastien, H.; Anne-Marie, L. miR-322 regulates insulin signaling pathway and protects against metabolic syndrome-induced cardiac dysfunction in mice. Biochim. Biophys. Acta 2016, 1862, 611–621. [Google Scholar]
- Sarkar, S.; Dey, B.K.; Dutta, A. MiR-322/424 and -503 are induced during muscle differentiation and promote cell cycle quiescence and differentiation by down-regulation of Cdc25A. Mol. Biol. Cell 2010, 21, 2138–2149. [Google Scholar] [CrossRef]
- Young Hoon, S.; Seok-Jin, L.; Ki-Baek, L.; Jin-Haeng, L.; Eui Man, J.; Sun Gun, C.; Sang-Chul, P.; In-Gyu, K. Dexamethasone downregulates caveolin-1 causing muscle atrophy via inhibited insulin signaling. J. Endocrinol. 2015, 225, 27–37. [Google Scholar]
- Crist, C.G.; Buckingham, M. Megarole for MicroRNA in Muscle Disease. Cell Metab. 2010, 12, 341–351. [Google Scholar] [CrossRef]
- Aaron, P.R.; Severine, L.; Hanneke, B.; Shogo, W.; Isabelle, G.; Erin, L.B.; Alexander, V.C.; Juleen, R.Z.; Rod, J.S.; Nigel, S.; et al. Regulation of miRNAs in human skeletal muscle following acute endurance exercise and short-term endurance training. J. Physiol. 2013, 591, 4637–4653. [Google Scholar]
- Aaron, P.R.; Shogo, W.; Lodovica, V.; MBenjamin, H.; Séverine, L.; Bertrand, L.; Takashi, U.; Romain, C.; Glenn, D.; Peter, H.; et al. Disruption of skeletal muscle mitochondrial network genes and miRNAs in amyotrophic lateral sclerosis. Neurobiol. Dis. 2013, 49, 107–117. [Google Scholar]
- Himani, K.; Kottaiswamy, A.; Arigela, H.; Sushmitha, S.; Peng Zhao, F.; Rengaraj, A.; Suan Liang Kelvin, T.; Sudarsanareddy, L.; Sabeera, B.; Sandhya, S.; et al. Muscle-specific microRNA1 (miR1) targets heat shock protein 70 (HSP70) during dexamethasone-mediated atrophy. J. Biol. Chem. 2013, 288, 6663–6678. [Google Scholar]
- Ricardo José, S.; Stefano, C.; Francesco, C.; Matteo, S.; Antonio, M.; Cristiano De, P.; Gerolamo Lanfranchi Marco, S. Involvement of microRNAs in the regulation of muscle wasting during catabolic conditions. J. Biol. Chem. 2014, 289, 21909. [Google Scholar]
- Cheng-Shyuan, R.; Jonathan Chris, J.; Seng-Feng, J.; Tsu-Hsiang, L.; Yi-Chun, C.; Po-Chou, L.; Chia-Jung, W.; Chia-Jung, L.; Ching-Hua, H. Entrapment neuropathy results in different microRNA expression patterns from denervation injury in rats. BMC Musculoskelet. Disord. 2010, 11, 181. [Google Scholar]
- Russell, A.; Wallace, M.; Kalanon, M.; Zacharewicz, E.; Della Gatta, P.; Garnham, A.; Lamon, S. STARS is reduced in ageing human skeletal muscle and targeted by miR-628-5p. Acta Physiologica 2017, 220, 263–274. [Google Scholar] [CrossRef]
- Lee, K.P.; Shin, Y.J.; Kwon, K.S. microRNA for determining the age-related myogenic capabilities of skeletal muscle. BMB Rep. 2015, 48, 595–596. [Google Scholar] [CrossRef]
- Bin, W.; Wen, D.; Dong, L.; Yan, X.; Zengqiang, Y.; Guangju, J. MST1, a key player, in enhancing fast skeletal muscle atrophy. BMC Biol. 2013, 11, 12. [Google Scholar]
- Paolo, E.P.; Nicoletta, F.; Simone, R.; Michele, F.; Elia, A.; Viola, F.G.; Flavia, P.; Giulia, R.; Sharmila, F.; Michele, F.; et al. Acylated and unacylated ghrelin impair skeletal muscle atrophy in mice. J. Clin. Investig. 2013, 123, 611–622. [Google Scholar]
- Brian, T.O.N.; Kevin, Y.L.; Katherine, K.; Samir, S.; Megan, T.K.; Joachim, F.; Kristin, I.S.; Matthew, M.R.; Weikang, C.; Andre, K.; et al. Insulin and IGF-1 receptors regulate FoxO-mediated signaling in muscle proteostasis. J. Clin. Investig. 2016, 126, 3433–3446. [Google Scholar]
- Stefano, S.; Kenneth, A.D.; Stefano, C.; Bert, B.; Marco, S. Mechanisms regulating skeletal muscle growth and atrophy. FEBS J. 2013, 280, 4294–4314. [Google Scholar]
- Zong-Kang, Z.; Jie, L.; Jin, L.; Baosheng, G.; Albert, L.; Ge, Z.; Bao-Ting, Z. Icaritin requires phosphatidylinositol 3 kinase/Akt signaling to counteract skeletal muscle atrophy following mechanical unloading. Sci. Rep. 2015, 6, 20300. [Google Scholar]
- Kim, S.; Kang, H. miR-15b induced by platelet-derived growth factor signaling is required for vascular smooth muscle cell proliferation. BMB Rep. 2013, 46, 550–554. [Google Scholar] [CrossRef]
- Martin, C.; Richard, P.; Roser, F.; Samantha, A.N.; Susannah, B.; Jen, L.; Jose, P.L.; Harnish, P.; Cyrus, C.; Avan, A.S.; et al. miR-424-5p reduces ribosomal RNA and protein synthesis in muscle wasting. J. Cachexia Sarcopenia Muscle 2017, 9, 400–416. [Google Scholar]
- Weidong, Z.; Weiping, Q.; Jiangping, P.; Yong, W.; William, A.B.; Christopher, C. Dependence of dexamethasone-induced Akt/FOXO1 signaling, upregulation of MAFbx, and protein catabolism upon the glucocorticoid receptor. Biochem. Biophys. Res. Commun. 2009, 378, 668–672. [Google Scholar]
- Monica, L.W.; Leslie, M.B.; Holger, M.R.; Jan, P.T.; Sue, C.B.; JDavid, F. A cell-autonomous role for the glucocorticoid receptor in skeletal muscle atrophy induced by systemic glucocorticoid exposure. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E1210. [Google Scholar]
- Gross, K.L.; Cidlowski, J.A. Tissue-specific glucocorticoid action: A family affair. Trends Endocrinol. Metabol. 2008, 19, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Caifu, C.; Dana, A.R.; Adam, J.B.; Zhaohui, Z.; Danny, H.L.; Julie, T.N.; Maura, B.; Nan Lan, X.; Vikram, R.M.; Mark, R.A.; et al. Real-time quantification of microRNAs by stem–loop RT–PCR. Nucl. Acids Res. 2005, 33, e179. [Google Scholar]
- Kyle, R.B.; Yann, S.G.; Shuichi, S.; Guangyan, X.; Sajedah, M.H.; Ashok, K. Inhibition of ER stress and unfolding protein response pathways causes skeletal muscle wasting during cancer cachexia. FASEB J. 2016, 30, 3053–3068. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| psiCHECK2-IGF1R-WT | CTCGAGATCTATACATCTGTACAGGAAAAGAAAA-GCTGCT-ATTTTTTTTTTGTTCTTTATCTTTGTGGATTTAATCTATGAA GCGGCCGC |
| psiCHECK2-IGF1R-Mut | CTCGAGATCTATACATCTGTACAGGAAAAGAAAA-GGTCCA-ATTTTTTTTTTGTTCTTTATCTTTGTGGATTTAATCTATGAA GCGGCCGC |
| psiCHECK2-INSR-WT | CTCGAGACAAAATCAGTTCCTCAAATTGACCAATAGC-TGCTGC-TTTCATATTTTATTTTGGGAAAGGGTGTGTATTCCTAAG GCGGCCGC |
| psiCHECK2-INSR-Mut | CTCGAGACAAAATCAGTTCCTCAAATTGACCAATAGC-TCCAGG-TTTCATATTTTATTTTGGGAAAGGGTGTGTATTCCTAAG GCGGCCGC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geng, H.; Song, Q.; Cheng, Y.; Li, H.; Yang, R.; Liu, S.; Hao, L. MicroRNA 322 Aggravates Dexamethasone-Induced Muscle Atrophy by Targeting IGF1R and INSR. Int. J. Mol. Sci. 2020, 21, 1111. https://doi.org/10.3390/ijms21031111
Geng H, Song Q, Cheng Y, Li H, Yang R, Liu S, Hao L. MicroRNA 322 Aggravates Dexamethasone-Induced Muscle Atrophy by Targeting IGF1R and INSR. International Journal of Molecular Sciences. 2020; 21(3):1111. https://doi.org/10.3390/ijms21031111
Chicago/Turabian StyleGeng, Hongwei, Qinglong Song, Yunyun Cheng, Haoyang Li, Rui Yang, Songcai Liu, and Linlin Hao. 2020. "MicroRNA 322 Aggravates Dexamethasone-Induced Muscle Atrophy by Targeting IGF1R and INSR" International Journal of Molecular Sciences 21, no. 3: 1111. https://doi.org/10.3390/ijms21031111
APA StyleGeng, H., Song, Q., Cheng, Y., Li, H., Yang, R., Liu, S., & Hao, L. (2020). MicroRNA 322 Aggravates Dexamethasone-Induced Muscle Atrophy by Targeting IGF1R and INSR. International Journal of Molecular Sciences, 21(3), 1111. https://doi.org/10.3390/ijms21031111