A New and Integral Approach to the Etiopathogenesis and Treatment of Breast Cancer Based upon Its Hydrogen Ion Dynamics

 , , ,
, , , 

Abstract
1. Introduction
2. Cancer as an Acid-Base Disease
- (a)
- (b)
- (c)
- pH and cancer molecular biology, biochemistry and metabolism: Recent experimental data have clearly shown that the pH-related paradigm can reinterpret the molecular biology, biochemistry and intermediary metabolism of cancer cells and tissues from an integral and Unitarian dynamic perspective [8,37,38,39,40,41,42,45,46].
- (d)
- (e)
- pH and microenvironmental-intratumoral acidosis in cancer growth and dissemination: Through different mechanisms, from biochemical to immunological, the high extrusion of lactic acid and other metabolites from cancer cells creates a highly acidified extracellular media that stimulates different and coordinated mechanisms strategically organized to favor local growth, invasion and dissemination [8,22,49,50,51,52,53,54].
- (f)
- (g)
- pH, proton transporters (PTs), proton pumps (PPs) and their inhibitors (PTIs and PPIs): During the last decade the increasing interest and knowledge of the different membrane-bound PTs and PPs in cancer pathogenesis, as well as their inhibitors in cancer treatment, has made it possible to approach, from a different and non-mainstream perspective, the latest therapeutic efforts in cancer treatment, either in BC or in other human malignant tumors [15,36,40,42,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70].
- (h)
- pHi acidification and reverting cancer proton reversal (CPR) in cancer treatment: Attempts to revert CPR in cancer are the fundamental therapeutic issue in the entire paradigm of the H+-related dynamics of cancer, BC included, from the initiation stage to angiogenesis and to the treatment of metastatic disease [8,16,23,55,63,71].
- (i)
- pH and voltage-gated sodium channels (VGSC): The expression of Na+ channels synergically associated with Na+/H+ antiporter activity and over-expression is important in BC pathogenesis by stimulating local invasion and the metastatic process, while its suppression is a valuable complementary therapeutic option [72,73,74,75,76,77].
- (j)
- pH and environmental carcinogenesis: An integral explanation of human environmental carcinogenesis has been recently advanced, proposing that the oncogenic activity of many different carcinogens induce the same cancer-specific effects on cellular H+ dynamics (CPR). This recent and highly important integration into the H+-related paradigm strongly suggests the possibility of the existence of a universal mechanism responsible for environmental carcinogenesis [18,19].
- (k)
- pH and cancer immunity: The acidic pHe of tumors has been considered to be the ultimate mechanism allowing them to escape from the anti-tumor immunity of the parasitized human organism. The final result is that this microenvironmental-intratumoral-extracellular (EC) low pHe creates a protective shield around cancer tissues with the onset of a state of anergy and immunosuppression mediated by the EC acidification-induced loss of function of T and NK cells. It also helps to explain the limitations and failures of checkpoint blockade in immunotherapy. Contrariwise, counteracting microenvironmental tumor acidity improves the antitumor responses to immunotherapy [20,21,47,78,79].
- (l)
- (m)
- (n)
- (o)
- (p)
- pH and autophagy: No matter that autophagy still appears to be a controversial issue, is considered a defensive survival mechanism of cancer cells in order to overcome drug-induced cellular stress and cytotoxicity. Acidic conditions increase autophagy in cancer cells suggesting that autophagy is a protective mechanism for tumor cells to survive under the microenvironmental acidic stress. Inhibiting autophagy may lead to a full cytotoxic effect [96,97,98,99,100] (for further details see the text).
- (q)
- (r)
- (s)
- (t)
3. All Phases of Breast Cancer Are Weaved into Each Other to Conform a Single, All-Comprehensive and Progressive, Multistage Unity
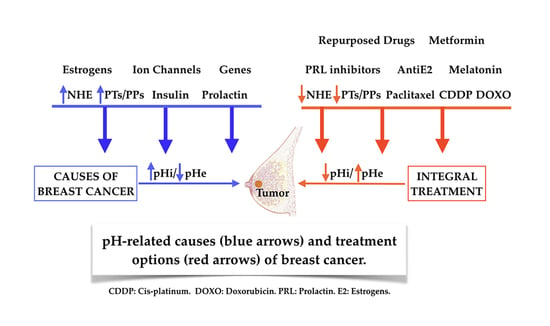
4. pH/NHE, Microenvironmental Acidosis and Immunity, Insulin, Prolactin, Estrogens, Genetic Abnormalities and Growth Factors in the Promotion of Breast Cancer
4.1. pH/NHE, H+ Extrusion and/or Intracellular Alkalization in the Etiology and Pathogenesis of Breast Cancer
4.2. A Universal Mechanism as a Final Mediating Cause of Breast Cancer
4.3. Tumor Microenvironmental (TME) Acidosis and Immunity
4.4. Insulin (INS) and Breast Cancer
4.5. Prolactin and Breast Cancer
4.6. Estrogens and Breast Cancer
- (a)
- NHE1 activity.
- (b)
- Hypoxia-inducible Factor activity (HIF).
- (c)
- Carbonic Anhydrases (CAs) activity: CA-XII expression relies on estradiol activity [156]. Therefore, ER+ is more likely to be associated with CA-XII rather than with CA-IX, while CA-IX is more frequently associated with ER- cells [212]. It has been shown that the selective inhibition of CA-XI improves the prognosis of the disease [200]. Although estrogens are growth factors, their effects or relations with the H+ dynamics of BC cells have not been well established. However, ER- BC cells seem to be associated with NHE1 activity [157] (Table 1). ERs show a high degree of heterogeneity in BC [156], as first reported by Puddefoot et al. in 1993 [258], and further confirmed by Leclercq some years later [259]. This heterogeneity implies, among other aspects, that at least four isoforms of ER alpha may exist, migrating to different isoelectric points in isoelectric focusing gels. Whether one or more of these different isoelectric points may be related or even contribute to changes in the pHi of mammary cells leading to BC remains to be established.
4.7. Ion Channels
4.8. PTs, PPs and Growth Factors
4.9. NHE1-Related Genetics of Breast Cancer
5. Hydrogen Ion Dynamics in Multiple Drug Resistance (MDR) in Breast Cancer and Other Malignant Tumors: An Integral Approach to Its Etiopathogenesis and Mediating Mechanisms
6. pH-Related Armamentarium in the Treatment of Breast Cancer
6.1. NHE Inhibitors and/or Intracellular Acidifiers
6.2. Proton Pumps (PPs) and Their Inhibitors (PPIs) in Cancer and Breast Cancer
6.3. Melatonin (MT) in Breast Cancer
6.4. Cisplatin (CDDP) and pH/NHE in Breast Cancer
6.5. pH and MDR in Breast Cancer: An Integrated Approach to Treatment
6.6. Repurposed Drugs in Breast Cancer Treatment
6.7. Metformin (MET) in Breast Cancer
- (i)
- It reduces circulating insulin and insulin/IGF-1 receptor-mediated activation of the PI3K pathway [245].
- (ii)
- MET inhibits the expression of the Hypoxia Inducible Factor 1 alpha (HIF-1α) gene expression, increases Pyruvate Dehydrogenase (PDH) gene expression [249] and decreases Warburg metabolism [35]. Additionally, HIF-1α is fundamental in tumoral angiogenesis and induces the expression of VEGF in BC [134,252,260,295]. Through this and other mechanisms MET also inhibits cancer growth, including triple-negative BC (TNBC) [254]. VC REMOVE:
- (iii)
- (iv)
- (v)
- (vi)
6.8. Autophagy and Cannibalism in Breast Cancer
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Evans, D.G.; Howell, A. Breast cancer risk-assessment models. Breast Cancer Res. 2007, 9, 213. [Google Scholar] [CrossRef] [PubMed]
- Redig, A.J.; McAllister, S.S. Breast cancer as a systemic disease: A view of metastasis. J. Intern. Med. 2013, 274, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Koedoot, E.; Fokkelman, M.; Rogkoti, V.M.; Smid, M.; van de Sandt, I.; de Bont, H.; Pont, C.; Klip, J.E.; Wink, S.; Timmermans, M.A.; et al. Uncovering the signaling landscape controlling breast cancer cell migration identifies novel metastasis driver genes. Nat. Commun. 2019, 10, 2983. [Google Scholar] [CrossRef] [PubMed]
- Samavat, H.; Kurzer, M.S. Estrogen metabolism and breast cancer. Cancer Lett. 2015, 356, 231–243. [Google Scholar] [CrossRef]
- Early Breast Cancer Trialists’ Collaborative Group. Effects of chemotherapy and hormonal therapy for early breast cancer on recurrence and 15-year survival: An overview of the randomised trials. Lancet 2005, 365, 1687–1717. [Google Scholar] [CrossRef]
- Egger, S.J.; Willson, M.L.; Morgan, J.; Walker, H.S.; Carrick, S.; Ghersi, D.; Wilcken, N. Platinum-containing regimens for metastatic breast cancer. Cochrane Database Syst. Rev. 2017, 6, CD003374. [Google Scholar] [CrossRef]
- Casey, J.R.; Grinstein, S.; Orlowski, J. Sensors and regulators of intracellular pH. Nat. Rev. Mol. Cell Biol. 2010, 11, 50–61. [Google Scholar] [CrossRef]
- Harguindey, S.; Stanciu, D.; Devesa, J.; Alfarouk, K.; Cardone, R.A.; Polo Orozco, J.D.; Devesa, P.; Rauch, C.; Orive, G.; Anitua, E.; et al. Cellular acidification as a new approach to cancer treatment and to the understanding and therapeutics of neurodegenerative diseases. Semin. Cancer Biol. 2017, 43, 157–179. [Google Scholar] [CrossRef]
- Harguindey, S. Hydrogen ion dynamics and cancer: An appraisal. Med. Pediatr. Oncol. 1982, 10, 217–236. [Google Scholar] [CrossRef]
- Pouyssegur, J.; Sardet, C.; Franchi, A.; L’Allemain, G.; Paris, S. A specific mutation abolishing Na+/H+ antiport activity in hamster fibroblasts precludes growth at neutral and acidic pH. Proc. Natl. Acad. Sci. USA 1984, 81, 4833–4837. [Google Scholar] [CrossRef]
- Moolenaar, W.H. Effects of growth factors on intracellular pH regulation. Annu. Rev. Physiol. 1986, 48, 363–376. [Google Scholar] [CrossRef]
- L’Allemain, G.; Paris, S.; Pouyssegur, J. Growth factor action and intracellular pH regulation in fibroblasts. Evidence for a major role of the Na+/H+ antiport. J. Biol. Chem. 1984, 259, 5809–5815. [Google Scholar] [PubMed]
- Harguindey, S. The Hydrogen Ion (pH) and its Implications in Cancer. Ph.D. Thesis, University of The Basque Country, Bilbao, Spain, 1989. [Google Scholar]
- Lagadic-Gossmann, D.; Hardonniere, K.; Mograbi, B.; Sergent, O.; Huc, L. Disturbances in H(+) dynamics during environmental carcinogenesis. Biochimie 2019, 163, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Spugnini, E.P.; Sonveaux, P.; Stock, C.; Perez-Sayans, M.; De Milito, A.; Avnet, S.; Garcia, A.G.; Harguindey, S.; Fais, S. Proton channels and exchangers in cancer. Biochim. Biophys. Acta 2015, 1848, 2715–2726. [Google Scholar] [CrossRef] [PubMed]
- Orive, G.; Reshkin, S.J.; Harguindey, S.; Pedraz, J.L. Hydrogen ion dynamics and the Na+/H+ exchanger in cancer angiogenesis and antiangiogenesis. Br. J. Cancer 2003, 89, 1395–1399. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Deng, C.; Zhang, M.; Zou, D.; Xu, M. Reduction of intracellular pH inhibits the expression of VEGF in K562 cells after targeted inhibition of the Na+/H+ exchanger. Leuk. Res. 2007, 31, 507–514. [Google Scholar] [CrossRef]
- Hardonniere, K.; Huc, L.; Sergent, O.; Holme, J.A.; Lagadic-Gossmann, D. Environmental carcinogenesis and pH homeostasis: Not only a matter of dysregulated metabolism. Semin. Cancer Biol. 2017, 43, 49–65. [Google Scholar] [CrossRef]
- Aravena, C.; Beltran, A.R.; Cornejo, M.; Torres, V.; Diaz, E.S.; Guzman-Gutierrez, E.; Pardo, F.; Leiva, A.; Sobrevia, L.; Ramirez, M.A. Potential role of sodium-proton exchangers in the low concentration arsenic trioxide-increased intracellular pH and cell proliferation. PLoS ONE 2012, 7, e51451. [Google Scholar] [CrossRef]
- Huber, V.; Camisaschi, C.; Berzi, A.; Ferro, S.; Lugini, L.; Triulzi, T.; Tuccitto, A.; Tagliabue, E.; Castelli, C.; Rivoltini, L. Cancer acidity: An ultimate frontier of tumor immune escape and a novel target of immunomodulation. Semin. Cancer Biol. 2017, 43, 74–89. [Google Scholar] [CrossRef]
- Lacroix, R.; Rozeman, E.A.; Kreutz, M.; Renner, K.; Blank, C.U. Targeting tumor-associated acidity in cancer immunotherapy. Cancer Immunol. Immunother. 2018, 67, 1331–1348. [Google Scholar] [CrossRef]
- Pillai, S.R.; Damaghi, M.; Marunaka, Y.; Spugnini, E.P.; Fais, S.; Gillies, R.J. Causes, consequences, and therapy of tumors acidosis. Cancer Metastasis Rev. 2019, 38, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Cardone, R.A.; Alfarouk, K.O.; Elliott, R.L.; Alqahtani, S.S.; Ahmed, S.B.M.; Aljarbou, A.N.; Greco, M.R.; Cannone, S.; Reshkin, S.J. The Role of Sodium Hydrogen Exchanger 1 in Dysregulation of Proton Dynamics and Reprogramming of Cancer Metabolism as a Sequela. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Harguindey, S.; Reshkin, S.J. “The new pH-centric anticancer paradigm in Oncology and Medicine”; SCB, 2017. Semin. Cancer Biol. 2017, 43, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Harguindey, S.; Polo Orozco, J.; Alfarouk, K.O.; Devesa, J. Hydrogen Ion Dynamics of Cancer and a New Molecular, Biochemical and Metabolic Approach to the Etiopathogenesis and Treatment of Brain Malignancies. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef]
- Rich, I.N.; Worthington-White, D.; Garden, O.A.; Musk, P. Apoptosis of leukemic cells accompanies reduction in intracellular pH after targeted inhibition of the Na(+)/H(+) exchanger. Blood 2000, 95, 1427–1434. [Google Scholar] [CrossRef]
- Perona, R.; Serrano, R. Increased pH and tumorigenicity of fibroblasts expressing a yeast proton pump. Nature 1988, 334, 438–440. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Fouad, Y.A.; Aanei, C. Revisiting the hallmarks of cancer. Am. J. Cancer Res. 2017, 7, 1016–1036. [Google Scholar]
- Warburg, O. On the origin of cancer cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef]
- Warburg, O.H. The Prime Cause and Prevention of Cancer; Nobel Laureates: Lindau, Germany, 1969. [Google Scholar]
- Nagata, H.; Che, X.F.; Miyazawa, K.; Tomoda, A.; Konishi, M.; Ubukata, H.; Tabuchi, T. Rapid decrease of intracellular pH associated with inhibition of Na+/H+ exchanger precedes apoptotic events in the MNK45 and MNK74 gastric cancer cell lines treated with 2-aminophenoxazine-3-one. Oncol. Rep. 2011, 25, 341–346. [Google Scholar] [CrossRef]
- Persi, E.; Duran-Frigola, M.; Damaghi, M.; Roush, W.R.; Aloy, P.; Cleveland, J.L.; Gillies, R.J.; Ruppin, E. Systems analysis of intracellular pH vulnerabilities for cancer therapy. Nat. Commun. 2018, 9, 2997. [Google Scholar] [CrossRef] [PubMed]
- Quach, C.H.; Jung, K.H.; Lee, J.H.; Park, J.W.; Moon, S.H.; Cho, Y.S.; Choe, Y.S.; Lee, K.H. Mild Alkalization Acutely Triggers the Warburg Effect by Enhancing Hexokinase Activity via Voltage-Dependent Anion Channel Binding. PLoS ONE 2016, 11, e0159529. [Google Scholar] [CrossRef] [PubMed]
- Alfarouk, K.O.; Verduzco, D.; Rauch, C.; Muddathir, A.K.; Adil, H.H.; Elhassan, G.O.; Ibrahim, M.E.; David Polo Orozco, J.; Cardone, R.A.; Reshkin, S.J.; et al. Glycolysis, tumor metabolism, cancer growth and dissemination. A new pH-based etiopathogenic perspective and therapeutic approach to an old cancer question. Oncoscience 2014, 1, 777–802. [Google Scholar] [CrossRef] [PubMed]
- Witkiewicz, A.K.; Whitaker-Menezes, D.; Dasgupta, A.; Philp, N.J.; Lin, Z.; Gandara, R.; Sneddon, S.; Martinez-Outschoorn, U.E.; Sotgia, F.; Lisanti, M.P. Using the “reverse Warburg effect” to identify high-risk breast cancer patients: Stromal MCT4 predicts poor clinical outcome in triple-negative breast cancers. Cell Cycle 2012, 11, 1108–1117. [Google Scholar] [CrossRef] [PubMed]
- Reshkin, S.J.; Bellizzi, A.; Caldeira, S.; Albarani, V.; Malanchi, I.; Poignee, M.; Alunni-Fabbroni, M.; Casavola, V.; Tommasino, M. Na+/H+ exchanger-dependent intracellular alkalinization is an early event in malignant transformation and plays an essential role in the development of subsequent transformation-associated phenotypes. FASEB J. 2000, 14, 2185–2197. [Google Scholar] [CrossRef] [PubMed]
- Grillo-Hill, B.K.; Choi, C.; Jimenez-Vidal, M.; Barber, D.L. Increased H(+) efflux is sufficient to induce dysplasia and necessary for viability with oncogene expression. Elife 2015, 4. [Google Scholar] [CrossRef]
- Harguindey, S.; Orive, G.; Luis Pedraz, J.; Paradiso, A.; Reshkin, S.J. The role of pH dynamics and the Na+/H+ antiporter in the etiopathogenesis and treatment of cancer. Two faces of the same coin—One single nature. Biochim. Biophys. Acta 2005, 1756, 1–24. [Google Scholar] [CrossRef]
- Amith, S.R.; Fliegel, L. Na(+)/H(+) exchanger-mediated hydrogen ion extrusion as a carcinogenic signal in triple-negative breast cancer etiopathogenesis and prospects for its inhibition in therapeutics. Semin. Cancer Biol. 2017, 43, 35–41. [Google Scholar] [CrossRef]
- Amith, S.R.; Fong, S.; Baksh, S.; Fliegel, L. Na (+)/H (+)exchange in the tumour microenvironment: Does NHE1 drive breast cancer carcinogenesis? Int. J. Dev. Biol. 2015, 59, 367–377. [Google Scholar] [CrossRef]
- Amith, S.R.; Fliegel, L. Regulation of the Na+/H+ Exchanger (NHE1) in Breast Cancer Metastasis. Cancer Res. 2013, 73, 1259–1264. [Google Scholar] [CrossRef]
- Amith, S.R.; Wilkinson, J.M.; Fliegel, L. Assessing Na(+)/H(+) exchange and cell effector functionality in metastatic breast cancer. Biochim. Open 2016, 2, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Amith, S.R.; Wilkinson, J.M.; Baksh, S.; Fliegel, L. The Na(+)/H(+) exchanger (NHE1) as a novel co-adjuvant target in paclitaxel therapy of triple-negative breast cancer cells. Oncotarget 2015, 6, 1262–1275. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Kim, S.S.; Lee, J. Cancer cell metabolism: Implications for therapeutic targets. Exp. Mol. Med. 2013, 45, e45. [Google Scholar] [CrossRef] [PubMed]
- Parks, S.K.; Chiche, J.; Pouyssegur, J. Disrupting proton dynamics and energy metabolism for cancer therapy. Nat. Rev. Cancer 2013, 13, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Estrella, V.; Enriquez-Navas, P.; El-Kenawi, A.; Russell, S.; Abrahams, D.; Ibrahim-Hashim, A.; Longo, D.; Reshetnyak, Y.; Luddy, K. Lymph Nodes Inhibit T-cell Effector Functions Locally by Establishing Acidic Niches. bioRxiv 2019. [Google Scholar] [CrossRef]
- Cardone, R.A.; Casavola, V.; Reshkin, S.J. The role of disturbed pH dynamics and the Na+/H+ exchanger in metastasis. Nat. Rev. Cancer 2005, 5, 786–795. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.; Spugnini, E.P.; Assaraf, Y.G.; Azzarito, T.; Rauch, C.; Fais, S. Microenvironment acidity as a major determinant of tumor chemoresistance: Proton pump inhibitors (PPIs) as a novel therapeutic approach. Drug Resist. Updat. 2015, 23, 69–78. [Google Scholar] [CrossRef]
- Corbet, C.; Feron, O. Tumour acidosis: From the passenger to the driver’s seat. Nat. Rev. Cancer 2017, 17, 577–593. [Google Scholar] [CrossRef]
- Robey, I.F.; Baggett, B.K.; Kirkpatrick, N.D.; Roe, D.J.; Dosescu, J.; Sloane, B.F.; Hashim, A.I.; Morse, D.L.; Raghunand, N.; Gatenby, R.A.; et al. Bicarbonate increases tumor pH and inhibits spontaneous metastases. Cancer Res. 2009, 69, 2260–2268. [Google Scholar] [CrossRef]
- Melnik, S.; Dvornikov, D.; Muller-Decker, K.; Depner, S.; Stannek, P.; Meister, M.; Warth, A.; Thomas, M.; Muley, T.; Risch, A.; et al. Cancer cell specific inhibition of Wnt/beta-catenin signaling by forced intracellular acidification. Cell Discov. 2018, 4, 37. [Google Scholar] [CrossRef]
- Lee, Z.W.; Teo, X.Y.; Song, Z.J.; Nin, D.S.; Novera, W.; Choo, B.A.; Dymock, B.W.; Moore, P.K.; Huang, R.Y.; Deng, L.W. Intracellular Hyper-Acidification Potentiated by Hydrogen Sulfide Mediates Invasive and Therapy Resistant Cancer Cell Death. Front. Pharmacol. 2017, 8, 763. [Google Scholar] [CrossRef] [PubMed]
- Harguindey, S.; Arranz, J.L.; Wahl, M.L.; Orive, G.; Reshkin, S.J. Proton transport inhibitors as potentially selective anticancer drugs. Anticancer Res. 2009, 29, 2127–2136. [Google Scholar] [PubMed]
- Reshkin, S.J.; Cardone, R.A.; Harguindey, S. Na+-H+ exchanger, pH regulation and cancer. Recent Pat. Anticancer Drug Discov. 2013, 8, 85–99. [Google Scholar] [CrossRef] [PubMed]
- Andersen, A.P.; Samsoe-Petersen, J.; Oernbo, E.K.; Boedtkjer, E.; Moreira, J.M.A.; Kveiborg, M.; Pedersen, S.F. The net acid extruders NHE1, NBCn1 and MCT4 promote mammary tumor growth through distinct but overlapping mechanisms. Int. J. Cancer 2018, 142, 2529–2542. [Google Scholar] [CrossRef]
- Boedtkjer, E.; Moreira, J.M.; Mele, M.; Vahl, P.; Wielenga, V.T.; Christiansen, P.M.; Jensen, V.E.; Pedersen, S.F.; Aalkjaer, C. Contribution of Na+,HCO3(-)-cotransport to cellular pH control in human breast cancer: A role for the breast cancer susceptibility locus NBCn1 (SLC4A7). Int. J. Cancer 2013, 132, 1288–1299. [Google Scholar] [CrossRef]
- Lee, S.; Axelsen, T.V.; Andersen, A.P.; Vahl, P.; Pedersen, S.F.; Boedtkjer, E. Disrupting Na(+), HCO(3)(-)-cotransporter NBCn1 (Slc4a7) delays murine breast cancer development. Oncogene 2016, 35, 2112–2122. [Google Scholar] [CrossRef]
- Baenke, F.; Dubuis, S.; Brault, C.; Weigelt, B.; Dankworth, B.; Griffiths, B.; Jiang, M.; Mackay, A.; Saunders, B.; Spencer-Dene, B.; et al. Functional screening identifies MCT4 as a key regulator of breast cancer cell metabolism and survival. J. Pathol. 2015, 237, 152–165. [Google Scholar] [CrossRef]
- Loo, S.Y.; Chang, M.K.; Chua, C.S.; Kumar, A.P.; Pervaiz, S.; Clement, M.V. NHE-1: A promising target for novel anti-cancer therapeutics. Curr. Pharm. Des. 2012, 18, 1372–1382. [Google Scholar] [CrossRef]
- Pinheiro, C.; Sousa, B.; Albergaria, A.; Paredes, J.; Dufloth, R.; Vieira, D.; Schmitt, F.; Baltazar, F. GLUT1 and CAIX expression profiles in breast cancer correlate with adverse prognostic factors and MCT1 overexpression. Histol. Histopathol. 2011, 26, 1279–1286. [Google Scholar] [CrossRef]
- Hsieh, M.J.; Chen, K.S.; Chiou, H.L.; Hsieh, Y.S. Carbonic anhydrase XII promotes invasion and migration ability of MDA-MB-231 breast cancer cells through the p38 MAPK signaling pathway. Eur. J. Cell Biol. 2010, 89, 598–606. [Google Scholar] [CrossRef]
- Neri, D.; Supuran, C.T. Interfering with pH regulation in tumours as a therapeutic strategy. Nat. Rev. Drug Discov. 2011, 10, 767–777. [Google Scholar] [CrossRef]
- Mboge, M.Y.; Mahon, B.P.; McKenna, R.; Frost, S.C. Carbonic Anhydrases: Role in pH Control and Cancer. Metabolites 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Berrino, E.; Supuran, C.T. Novel approaches for designing drugs that interfere with pH regulation. Expert Opin. Drug Discov. 2019, 14, 231–248. [Google Scholar] [CrossRef] [PubMed]
- Lauritzen, G.; Stock, C.M.; Lemaire, J.; Lund, S.F.; Jensen, M.F.; Damsgaard, B.; Petersen, K.S.; Wiwel, M.; Ronnov-Jessen, L.; Schwab, A.; et al. The Na+/H+ exchanger NHE1, but not the Na+, HCO3(-) cotransporter NBCn1, regulates motility of MCF7 breast cancer cells expressing constitutively active ErbB2. Cancer Lett. 2012, 317, 172–183. [Google Scholar] [CrossRef]
- Chen, C.H.; Lee, C.Z.; Lin, Y.C.; Kao, L.T.; Lin, H.C. Negative Association of Proton Pump Inhibitors With Subsequent Development of Breast Cancer: A Nationwide Population-Based Study. J. Clin. Pharmacol. 2019, 59, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Mihaila, R.G. A minireview on NHE1 inhibitors. A rediscovered hope in oncohematology. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech. Repub. 2015, 159, 519–526. [Google Scholar] [CrossRef]
- Goh, W.; Sleptsova-Freidrich, I.; Petrovic, N. Use of proton pump inhibitors as adjunct treatment for triple-negative breast cancers. An introductory study. J. Pharm. Pharm. Sci. 2014, 17, 439–446. [Google Scholar] [CrossRef]
- Flinck, M.; Kramer, S.H.; Schnipper, J.; Andersen, A.P.; Pedersen, S.F. The acid-base transport proteins NHE1 and NBCn1 regulate cell cycle progression in human breast cancer cells. Cell Cycle 2018, 17, 1056–1067. [Google Scholar] [CrossRef]
- White, K.A.; Grillo-Hill, B.K.; Barber, D.L. Cancer cell behaviors mediated by dysregulated pH dynamics at a glance. J. Cell Sci. 2017, 130, 663–669. [Google Scholar] [CrossRef]
- Brisson, L.; Gillet, L.; Calaghan, S.; Besson, P.; Le Guennec, J.; Roger, S.; Gore, J. NaV1. 5 enhances breast cancer cell invasiveness by increasing NHE1-dependent H+ efflux in caveolae. Oncogene 2011, 30, 2070–2076. [Google Scholar] [CrossRef]
- Wang, Y.; Li, S.J.; Wu, X.; Che, Y.; Li, Q. Clinicopathological and biological significance of human voltage-gated proton channel Hv1 protein overexpression in breast cancer. J. Biol. Chem. 2012, 287, 13877–13888. [Google Scholar] [CrossRef]
- Brisson, L.; Driffort, V.; Benoist, L.; Poet, M.; Counillon, L.; Antelmi, E.; Rubino, R.; Besson, P.; Labbal, F.; Chevalier, S.; et al. NaV1.5 Na(+) channels allosterically regulate the NHE-1 exchanger and promote the activity of breast cancer cell invadopodia. J. Cell Sci. 2013, 126, 4835–4842. [Google Scholar] [CrossRef] [PubMed]
- Schwab, A.; Stock, C. Ion channels and transporters in tumour cell migration and invasion. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130102. [Google Scholar] [CrossRef] [PubMed]
- Fraser, S.P.; Diss, J.K.; Chioni, A.M.; Mycielska, M.E.; Pan, H.; Yamaci, R.F.; Pani, F.; Siwy, Z.; Krasowska, M.; Grzywna, Z.; et al. Voltage-gated sodium channel expression and potentiation of human breast cancer metastasis. Clin. Cancer Res. 2005, 11, 5381–5389. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Kozminski, D.J.; Wold, L.A.; Modak, R.; Calhoun, J.D.; Isom, L.L.; Brackenbury, W.J. Therapeutic potential for phenytoin: Targeting Na(v)1.5 sodium channels to reduce migration and invasion in metastatic breast cancer. Breast Cancer Res. Treat. 2012, 134, 603–615. [Google Scholar] [CrossRef]
- Pilon-Thomas, S.; Kodumudi, K.N.; El-Kenawi, A.E.; Russell, S.; Weber, A.M.; Luddy, K.; Damaghi, M.; Wojtkowiak, J.W.; Mule, J.J.; Ibrahim-Hashim, A.; et al. Neutralization of Tumor Acidity Improves Antitumor Responses to Immunotherapy. Cancer Res. 2016, 76, 1381–1390. [Google Scholar] [CrossRef]
- Marches, R.; Vitetta, E.S.; Uhr, J.W. A role for intracellular pH in membrane IgM-mediated cell death of human B lymphomas. Proc. Natl. Acad. Sci. USA 2001, 98, 3434–3439. [Google Scholar] [CrossRef]
- Harguindey, S.; Pedraz, J.L.; Canero, R.G.; Katin, M. Edelfosine, apoptosis, MDR and Na+/H+ exchanger: Induction mechanisms and treatment implications. Apoptosis 2000, 5, 87–89. [Google Scholar] [CrossRef]
- Perez-Sala, D.; Collado-Escobar, D.; Mollinedo, F. Intracellular alkalinization suppresses lovastatin-induced apoptosis in HL-60 cells through the inactivation of a pH-dependent endonuclease. J. Biol. Chem. 1995, 270, 6235–6242. [Google Scholar] [CrossRef]
- Lagadic-Gossmann, D.; Huc, L.; Lecureur, V. Alterations of intracellular pH homeostasis in apoptosis: Origins and roles. Cell Death Differ. 2004, 11, 953–961. [Google Scholar] [CrossRef]
- Sergeeva, T.F.; Shirmanova, M.V.; Zlobovskaya, O.A.; Gavrina, A.I.; Dudenkova, V.V.; Lukina, M.M.; Lukyanov, K.A.; Zagaynova, E.V. Relationship between intracellular pH, metabolic co-factors and caspase-3 activation in cancer cells during apoptosis. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Daniel, C.; Bell, C.; Burton, C.; Harguindey, S.; Reshkin, S.J.; Rauch, C. The role of proton dynamics in the development and maintenance of multidrug resistance in cancer. Biochim. Biophys. Acta 2013, 1832, 606–617. [Google Scholar] [CrossRef] [PubMed]
- Walsh, M.; Fais, S.; Spugnini, E.P.; Harguindey, S.; Abu Izneid, T.; Scacco, L.; Williams, P.; Allegrucci, C.; Rauch, C.; Omran, Z. Proton pump inhibitors for the treatment of cancer in companion animals. J. Exp. Clin. Cancer Res. 2015, 34, 93. [Google Scholar] [CrossRef] [PubMed]
- Rauch, C. On the relationship between drug’s size, cell membrane mechanical properties and high levels of multi drug resistance: A comparison to published data. Eur. Biophys. J. 2009, 38, 537–546. [Google Scholar] [CrossRef]
- Weisburg, J.H.; Roepe, P.D.; Dzekunov, S.; Scheinberg, D.A. Intracellular pH and multidrug resistance regulate complement-mediated cytotoxicity of nucleated human cells. J. Biol. Chem. 1999, 274, 10877–10888. [Google Scholar] [CrossRef]
- Roepe, P.D. pH and multidrug resistance. Novartis Found. Symp. 2001, 240, 232–247. [Google Scholar] [CrossRef]
- Rauch, C.; Blanchard, A.; Wood, E.; Dillon, E.; Wahl, M.L.; Harguindey, S. Cell Membranes, Cytosolic pH and Drug Transport in Cancer and MDR: Physics, Biochemistry and Molecular Biology. Mult. Drug Resist. 2009, 1, 24. [Google Scholar]
- Roepe, P.D.; Wei, L.Y.; Cruz, J.; Carlson, D. Lower electrical membrane potential and altered pHi homeostasis in multidrug-resistant (MDR) cells: Further characterization of a series of MDR cell lines expressing different levels of P-glycoprotein. Biochemistry 1993, 32, 11042–11056. [Google Scholar] [CrossRef]
- Ho, C.C.; Ding, S.J. The pH-controlled nanoparticles size of polydopamine for anti-cancer drug delivery. J. Mater. Sci. Mater. Med. 2013, 24, 2381–2390. [Google Scholar] [CrossRef]
- Karanth, H.; Murthy, R.S. pH-sensitive liposomes--principle and application in cancer therapy. J. Pharm. Pharmacol. 2007, 59, 469–483. [Google Scholar] [CrossRef]
- Bariwal, J.; Kumar, V.; Chen, H.; Bhattarai, R.S.; Peng, Y.; Li, W.; Mahato, R.I. Nanoparticulate delivery of potent microtubule inhibitor for metastatic melanoma treatment. J. Control. Release 2019, 309, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Nemeth-Cahalan, K.L.; Kalman, K.; Hall, J.E. Molecular basis of pH and Ca2+ regulation of aquaporin water permeability. J. Gen. Physiol. 2004, 123, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Mosca, A.F.; de Almeida, A.; Wragg, D.; Martins, A.P.; Sabir, F.; Leoni, S.; Moura, T.F.; Prista, C.; Casini, A.; Soveral, G. Molecular Basis of Aquaporin-7 Permeability Regulation by pH. Cells 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Marino, M.L.; Pellegrini, P.; Di Lernia, G.; Djavaheri-Mergny, M.; Brnjic, S.; Zhang, X.; Hagg, M.; Linder, S.; Fais, S.; Codogno, P.; et al. Autophagy is a protective mechanism for human melanoma cells under acidic stress. J. Biol. Chem. 2012, 287, 30664–30676. [Google Scholar] [CrossRef] [PubMed]
- Wojtkowiak, J.W.; Rothberg, J.M.; Kumar, V.; Schramm, K.J.; Haller, E.; Proemsey, J.B.; Lloyd, M.C.; Sloane, B.F.; Gillies, R.J. Chronic autophagy is a cellular adaptation to tumor acidic pH microenvironments. Cancer Res. 2012, 72, 3938–3947. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, P.; Dyczynski, M.; Sbrana, F.V.; Karlgren, M.; Buoncervello, M.; Hagg-Olofsson, M.; Ma, R.; Hartman, J.; Bajalica-Lagercrantz, S.; Grander, D.; et al. Tumor acidosis enhances cytotoxic effects and autophagy inhibition by salinomycin on cancer cell lines and cancer stem cells. Oncotarget 2016. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, P.; Strambi, A.; Zipoli, C.; Hagg-Olofsson, M.; Buoncervello, M.; Linder, S.; De Milito, A. Acidic extracellular pH neutralizes the autophagy-inhibiting activity of chloroquine: Implications for cancer therapies. Autophagy 2014, 10, 562–571. [Google Scholar] [CrossRef]
- Matsuo, T.; Sadzuka, Y. Extracellular acidification by lactic acid suppresses glucose deprivation-induced cell death and autophagy in B16 melanoma cells. Biochem. Biophys. Res. Commun. 2018, 496, 1357–1361. [Google Scholar] [CrossRef]
- Gieling, R.G.; Parker, C.A.; De Costa, L.A.; Robertson, N.; Harris, A.L.; Stratford, I.J.; Williams, K.J. Inhibition of carbonic anhydrase activity modifies the toxicity of doxorubicin and melphalan in tumour cells in vitro. J. Enzyme Inhib. Med. Chem. 2013, 28, 360–369. [Google Scholar] [CrossRef]
- Morais-Santos, F.; Granja, S.; Miranda-Goncalves, V.; Moreira, A.H.; Queiros, S.; Vilaca, J.L.; Schmitt, F.C.; Longatto-Filho, A.; Paredes, J.; Baltazar, F.; et al. Targeting lactate transport suppresses in vivo breast tumour growth. Oncotarget 2015, 6, 19177–19189. [Google Scholar] [CrossRef]
- Bai, F.; Yu, Z.; Gao, X.; Gong, J.; Fan, L.; Liu, F. Simvastatin induces breast cancer cell death through oxidative stress up-regulating miR-140-5p. Aging 2019, 11, 3198–3219. [Google Scholar] [CrossRef]
- Iessi, E.; Logozzi, M.; Lugini, L.; Azzarito, T.; Federici, C.; Spugnini, E.P.; Mizzoni, D.; Di Raimo, R.; Angelini, D.F.; Battistini, L.; et al. Acridine Orange/exosomes increase the delivery and the effectiveness of Acridine Orange in human melanoma cells: A new prototype for theranostics of tumors. J. Enzyme Inhib. Med. Chem. 2017, 32, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Gdovin, M.J.; Kadri, N.; Rios, L.; Holliday, S.; Jordan, Z. Focal photodynamic intracellular acidification as a cancer therapeutic. Semin. Cancer Biol. 2017, 43, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Cole, W.H.; Everson, T.C. Spontaneous regression of cancer: Preliminary report. Ann. Surg. 1956, 144, 366–383. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, E.M. Complete regression of vesical carcinoma following urinary diversion. Am. J. Surg. 1960, 100, 133–136. [Google Scholar] [CrossRef]
- Papac, R.J. Spontaneous regression of cancer: Possible mechanisms. In Vivo 1998, 12, 571–578. [Google Scholar] [PubMed]
- Goldberg, L.G. Complete regression of carcinoma of the bladder following ureterosigmoidostomy. J. Urol. 1950, 63, 116–123. [Google Scholar] [CrossRef]
- Harguindey, S.; Cragoe, E.J., Jr. The Na+/H+ antiporter in oncology in the light of the spontaneous regression of cancer and cell metabolism. Med. Hypotheses 1992, 39, 229–237. [Google Scholar] [CrossRef]
- Oleksyszyn, J. Spontaneous Regression of Cancer, an Ever Actual Inspiration for the New Cancer Treatment. Cell Biol. 2016, 5. [Google Scholar] [CrossRef]
- Harguindey, S.S.; Kolbeck, R.C.; Bransome, E.D., Jr. Letter: Ureterosigmoidostomy and cancer: New observations. Ann. Intern. Med. 1975, 83, 833. [Google Scholar] [CrossRef]
- Gillies, R.J.; Gatenby, R.A. Metabolism and its sequelae in cancer evolution and therapy. Cancer J. 2015, 21, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Harguindey, S.; Gillis, M. Evolution and cancer: Possible relationships to changes in environmental hydrogen ion concentration. J. Theor. Biol. 1980, 86, 487–492. [Google Scholar] [CrossRef]
- Alfarouk, K.O.; Shayoub, M.E.; Muddathir, A.K.; Elhassan, G.O.; Bashir, A.H. Evolution of Tumor Metabolism might Reflect Carcinogenesis as a Reverse Evolution process (Dismantling of Multicellularity). Cancers (Basel) 2011, 3, 3002–3017. [Google Scholar] [CrossRef] [PubMed]
- Alfarouk, K.O.; Muddathir, A.K.; Shayoub, M.E. Tumor acidity as evolutionary spite. Cancers (Basel) 2011, 3, 408–414. [Google Scholar] [CrossRef]
- Gatenby, R.A.; Vincent, T.L. An evolutionary model of carcinogenesis. Cancer Res. 2003, 63, 6212–6220. [Google Scholar]
- Harguindey, S.; Pedraz, J.L.; Garcia Canero, R.; Perez de Diego, J.; Cragoe, E.J., Jr. Hydrogen ion-dependent oncogenesis and parallel new avenues to cancer prevention and treatment using a H(+)-mediated unifying approach: pH-related and pH-unrelated mechanisms. Crit. Rev. Oncog. 1995, 6, 1–33. [Google Scholar] [CrossRef]
- Saponaro, C.; Vagheggini, A.; Scarpi, E.; Centonze, M.; Catacchio, I.; Popescu, O.; Pastena, M.I.; Giotta, F.; Silvestris, N.; Mangia, A. NHERF1 and tumor microenvironment: A new scene in invasive breast carcinoma. J. Exp. Clin. Cancer Res. 2018, 37, 96. [Google Scholar] [CrossRef]
- Di Pompo, G.; Lemma, S.; Canti, L.; Rucci, N.; Ponzetti, M.; Errani, C.; Donati, D.M.; Russell, S.; Gillies, R.; Chano, T.; et al. Intratumoral acidosis fosters cancer-induced bone pain through the activation of the mesenchymal tumor-associated stroma in bone metastasis from breast carcinoma. Oncotarget 2017, 8, 54478–54496. [Google Scholar] [CrossRef]
- Gonzalez, J.F.; Madamba, A.; Antón, L.A.; Jurado, M.C.; Harguindey, S. Cervical mosaic and an integrated pathophysiological approach to early cervical neoplasia. J. Biol. Response Mod. 1987, 6, 473–483. [Google Scholar]
- Lobo, R.C.; Hubbard, N.E.; Damonte, P.; Mori, H.; Penzvalto, Z.; Pham, C.; Koehne, A.L.; Go, A.C.; Anderson, S.E.; Cala, P.M.; et al. Glucose Uptake and Intracellular pH in a Mouse Model of Ductal Carcinoma In situ (DCIS) Suggests Metabolic Heterogeneity. Front. Cell Dev. Biol. 2016, 4, 93. [Google Scholar] [CrossRef]
- Cong, D.; Zhu, W.; Shi, Y.; Pointer, K.B.; Clark, P.A.; Shen, H.; Kuo, J.S.; Hu, S.; Sun, D. Upregulation of NHE1 protein expression enables glioblastoma cells to escape TMZ-mediated toxicity via increased H(+) extrusion, cell migration and survival. Carcinogenesis 2014, 35, 2014–2024. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, S.; Eguchi, Y.; Kamiike, W.; Funahashi, Y.; Mignon, A.; Lacronique, V.; Matsuda, H.; Tsujimoto, Y. Bcl-2 prevents apoptotic mitochondrial dysfunction by regulating proton flux. Proc. Natl. Acad. Sci. USA 1998, 95, 1455–1459. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Shibuya, I.; Ise, T.; Chen, Z.S.; Akiyama, S.; Nakagawa, M.; Izumi, H.; Nakamura, T.; Matsuo, K.; Yamada, Y.; et al. Elevated expression of vacuolar proton pump genes and cellular PH in cisplatin resistance. Int. J. Cancer 2001, 93, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Dumas, J.F.; Brisson, L.; Chevalier, S.; Maheo, K.; Fromont, G.; Moussata, D.; Besson, P.; Roger, S. Metabolic reprogramming in cancer cells, consequences on pH and tumour progression: Integrated therapeutic perspectives with dietary lipids as adjuvant to anticancer treatment. Semin. Cancer Biol. 2017, 43, 90–110. [Google Scholar] [CrossRef] [PubMed]
- Roma-Rodrigues, C.; Mendes, R.; Baptista, P.V.; Fernandes, A.R. Targeting Tumor Microenvironment for Cancer Therapy. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Fais, S.; Venturi, G.; Gatenby, B. Microenvironmental acidosis in carcinogenesis and metastases: New strategies in prevention and therapy. Cancer Metastasis Rev. 2014, 33, 1095–1108. [Google Scholar] [CrossRef]
- Calcinotto, A.; Filipazzi, P.; Grioni, M.; Iero, M.; De Milito, A.; Ricupito, A.; Cova, A.; Canese, R.; Jachetti, E.; Rossetti, M.; et al. Modulation of microenvironment acidity reverses anergy in human and murine tumor-infiltrating T lymphocytes. Cancer Res. 2012, 72, 2746–2756. [Google Scholar] [CrossRef]
- Thews, O.; Gassner, B.; Kelleher, D.K.; Schwerdt, G.; Gekle, M. Impact of extracellular acidity on the activity of P-glycoprotein and the cytotoxicity of chemotherapeutic drugs. Neoplasia 2006, 8, 143–152. [Google Scholar] [CrossRef]
- Thiebaut, F.; Currier, S.J.; Whitaker, J.; Haugland, R.P.; Gottesman, M.M.; Pastan, I.; Willingham, M.C. Activity of the multidrug transporter results in alkalinization of the cytosol: Measurement of cytosolic pH by microinjection of a pH-sensitive dye. J. Histochem. Cytochem. 1990, 38, 685–690. [Google Scholar] [CrossRef]
- Fais, S. Proton pump inhibitor-induced tumour cell death by inhibition of a detoxification mechanism. J. Intern. Med. 2010, 267, 515–525. [Google Scholar] [CrossRef]
- Alfarouk, K.O. Tumor metabolism, cancer cell transporters, and microenvironmental resistance. J. Enzyme Inhib. Med. Chem. 2016, 31, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Thews, O.; Nowak, M.; Sauvant, C.; Gekle, M. Hypoxia-induced extracellular acidosis increases p-glycoprotein activity and chemoresistance in tumors in vivo via p38 signaling pathway. In Oxygen Transport to Tissue XXXII; Springer: Boston, MA, USA, 2011; pp. 115–122. [Google Scholar] [CrossRef]
- Damaghi, M.; Gillies, R. Phenotypic changes of acid-adapted cancer cells push them toward aggressiveness in their evolution in the tumor microenvironment. Cell Cycle 2017, 16, 1739–1743. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Singh, R.; Pochampally, R.; Watabe, K.; Mo, Y.Y. Acidosis promotes invasiveness of breast cancer cells through ROS-AKT-NF-kappaB pathway. Oncotarget 2014, 5, 12070–12082. [Google Scholar] [CrossRef] [PubMed]
- Montcourrier, P.; Silver, I.; Farnoud, R.; Bird, I.; Rochefort, H. Breast cancer cells have a high capacity to acidify extracellular milieu by a dual mechanism. Clin. Exp. Metastasis 1997, 15, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Rothberg, J.M.; Bailey, K.M.; Wojtkowiak, J.W.; Ben-Nun, Y.; Bogyo, M.; Weber, E.; Moin, K.; Blum, G.; Mattingly, R.R.; Gillies, R.J.; et al. Acid-mediated tumor proteolysis: Contribution of cysteine cathepsins. Neoplasia 2013, 15, 1125–1137. [Google Scholar] [CrossRef]
- Wani, B.; Aziz, S.A.; Ganaie, M.A.; Mir, M.H. Metabolic Syndrome and Breast Cancer Risk. Indian J. Med. Paediatr. Oncol. 2017, 38, 434–439. [Google Scholar] [CrossRef]
- Tsujimoto, T.; Kajio, H.; Sugiyama, T. Association between hyperinsulinemia and increased risk of cancer death in nonobese and obese people: A population-based observational study. Int. J. Cancer 2017, 141, 102–111. [Google Scholar] [CrossRef]
- Evans, J.L.; Maddux, B.A.; Goldfine, I.D. The molecular basis for oxidative stress-induced insulin resistance. Antioxid. Redox Signal. 2005, 7, 1040–1052. [Google Scholar] [CrossRef]
- Ramirez, M.A.; Beltran, A.R.; Araya, J.E.; Cornejo, M.; Toledo, F.; Fuentes, G.; Sobrevia, L. Involvement of Intracellular pH in Vascular Insulin Resistance. Curr. Vasc. Pharmacol. 2019, 17, 440–446. [Google Scholar] [CrossRef]
- Boyd, D.B. Insulin and cancer. Integr. Cancer Ther. 2003, 2, 315–329. [Google Scholar] [CrossRef]
- Moore, R.D.; Gupta, R.K. Effect of insulin on intracellular pH as observed by 31P NMR spectroscopy. Int. J. Quantum Chem. 1980, 18, 83–92. [Google Scholar] [CrossRef]
- Kaloyianni, M.; Bourikas, D.; Koliakos, G. The effect of insulin on Na+-H+ antiport activity of obese and normal subjects erythrocytes. Cell. Physiol. Biochem. 2001, 11, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Arcidiacono, B.; Iiritano, S.; Nocera, A.; Possidente, K.; Nevolo, M.T.; Ventura, V.; Foti, D.; Chiefari, E.; Brunetti, A. Insulin resistance and cancer risk: An overview of the pathogenetic mechanisms. Exp. Diabetes Res. 2012, 2012, 789174. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Narvaez, E.A.; Lunetta, K.L.; Hong, C.C.; Haddad, S.; Yao, S.; Cheng, T.D.; Bensen, J.T.; Bandera, E.V.; Haiman, C.A.; Troester, M.A.; et al. Genetic variation in the insulin, insulin-like growth factor, growth hormone, and leptin pathways in relation to breast cancer in African-American women: The AMBER consortium. NPJ Breast Cancer 2016, 2. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Gao, B.; Xiong, Q.J.; Wang, Y.C.; Huang, D.K.; Wu, W.N. Acid-sensing ion channels contribute to the effect of extracellular acidosis on proliferation and migration of A549 cells. Tumor Biol. 2017, 39. [Google Scholar] [CrossRef] [PubMed]
- Bellizzi, A.; Greco, M.R.; Rubino, R.; Paradiso, A.; Forciniti, S.; Zeeberg, K.; Cardone, R.A.; Reshkin, S.J. The scaffolding protein NHERF1 sensitizes EGFR-dependent tumor growth, motility and invadopodia function to gefitinib treatment in breast cancer cells. Int. J. Oncol. 2015, 46, 1214–1224. [Google Scholar] [CrossRef]
- Pedraz-Cuesta, E.; Fredsted, J.; Jensen, H.H.; Bornebusch, A.; Nejsum, L.N.; Kragelund, B.B.; Pedersen, S.F. Prolactin Signaling Stimulates Invasion via Na(+)/H(+) Exchanger NHE1 in T47D Human Breast Cancer Cells. Mol. Endocrinol. 2016, 30, 693–708. [Google Scholar] [CrossRef][Green Version]
- Clevenger, C.V.; Furth, P.A.; Hankinson, S.E.; Schuler, L.A. The role of prolactin in mammary carcinoma. Endocr. Rev. 2003, 24, 1–27. [Google Scholar] [CrossRef]
- Xu, J.; Sun, D.; Jiang, J.; Deng, L.; Zhang, Y.; Yu, H.; Bahl, D.; Langenheim, J.F.; Chen, W.Y.; Fuchs, S.Y.; et al. The role of prolactin receptor in GH signaling in breast cancer cells. Mol. Endocrinol. 2013, 27, 266–279. [Google Scholar] [CrossRef]
- Seo, E.J.; Sugimoto, Y.; Greten, H.J.; Efferth, T. Repurposing of Bromocriptine for Cancer Therapy. Front. Pharmacol. 2018, 9, 1030. [Google Scholar] [CrossRef]
- Yager, J.D.; Davidson, N.E. Estrogen carcinogenesis in breast cancer. N. Engl. J. Med. 2006, 354, 270–282. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, M.C.; Alfarouk, K.O.; Verduzco, D.; Bui, M.M.; Gillies, R.J.; Ibrahim, M.E.; Brown, J.S.; Gatenby, R.A. Vascular measurements correlate with estrogen receptor status. BMC Cancer 2014, 14, 279. [Google Scholar] [CrossRef] [PubMed]
- Gruvberger, S.; Ringner, M.; Chen, Y.; Panavally, S.; Saal, L.H.; Borg, A.; Ferno, M.; Peterson, C.; Meltzer, P.S. Estrogen receptor status in breast cancer is associated with remarkably distinct gene expression patterns. Cancer Res. 2001, 61, 5979–5984. [Google Scholar] [PubMed]
- Mouridsen, H.T. Letrozole in advanced breast cancer: The PO25 trial. Breast Cancer Res. Treat. 2007, 105, 19–29. [Google Scholar] [CrossRef]
- Ibrahim-Hashim, A.; Wojtkowiak, J.W.; de Lourdes Coelho Ribeiro, M.; Estrella, V.; Bailey, K.M.; Cornnell, H.H.; Gatenby, R.A.; Gillies, R.J. Free Base Lysine Increases Survival and Reduces Metastasis in Prostate Cancer Model; OMICS Publishing Group: Hyderabad, India, 2011. [Google Scholar]
- Harguindey, S.; DeCastro, L.; Barcos, M.; Getaz, E.P.; Henderson, E.S.; Freeman, A. Hypercalcemia complicating childhood malignancies: A report of seven cases with some pathophysiological considerations. Cancer 1979, 44, 2280–2290. [Google Scholar] [CrossRef]
- Della Guardia, L.; Thomas, M.A.; Cena, H. Insulin Sensitivity and Glucose Homeostasis Can Be Influenced by Metabolic Acid Load. Nutrients 2018, 10. [Google Scholar] [CrossRef]
- Harguindey, S. Homeostasis, diabetes and cancer. An. Med. Interna 1993, 10, 611–616. [Google Scholar]
- Munoz, J.; Mohd, K.; Amr, H.; Stoltenberg, M. Severe lactic acidosis in a patient with metastatic prostate cancer. J. Cancer Res. Ther. 2011, 7, 201–202. [Google Scholar] [CrossRef]
- Van der Mijn, J.C.; Kuiper, M.J.; Siegert, C.E.H.; Wassenaar, A.E.; van Noesel, C.J.M.; Ogilvie, A.C. Lactic Acidosis in Prostate Cancer: Consider the Warburg Effect. Case Rep. Oncol. 2017, 10, 1085–1091. [Google Scholar] [CrossRef]
- Tuccori, M.; Wu, J.W.; Yin, H.; Majdan, A.; Azoulay, L. The use of glyburide compared with other sulfonylureas and the risk of cancer in patients with type 2 diabetes. Diabetes Care 2015, 38, 2083–2089. [Google Scholar] [CrossRef]
- Chen, Q.; Liu, Y.; Zhu, X.L.; Feng, F.; Yang, H.; Xu, W. Increased NHE1 expression is targeted by specific inhibitor cariporide to sensitize resistant breast cancer cells to doxorubicin in vitro and in vivo. BMC Cancer 2019, 19, 211. [Google Scholar] [CrossRef] [PubMed]
- Pasello, G.; Urso, L.; Conte, P.; Favaretto, A. Effects of sulfonylureas on tumor growth: A review of the literature. Oncologist 2013, 18, 1118–1125. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Yang, T.; Xu, W. Enemies or weapons in hands: Investigational anti-diabetic drug glibenclamide and cancer risk. Expert Opin. Investig. Drugs 2017, 26, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; Lord, J.M.; Norman, R.J.; Yasmin, E.; Balen, A.H. Insulin-sensitising drugs (metformin, rosiglitazone, pioglitazone, D-chiro-inositol) for women with polycystic ovary syndrome, oligo amenorrhoea and subfertility. Cochrane Database Syst. Rev. 2012. [Google Scholar] [CrossRef] [PubMed]
- Belli, S.H.; Graffigna, M.N.; Oneto, A.; Otero, P.; Schurman, L.; Levalle, O.A. Effect of rosiglitazone on insulin resistance, growth factors, and reproductive disturbances in women with polycystic ovary syndrome. Fertil. Steril. 2004, 81, 624–629. [Google Scholar] [CrossRef]
- Harguindey, S.; Arranz, J.L.; Polo Orozco, J.D.; Rauch, C.; Fais, S.; Cardone, R.A.; Reshkin, S.J. Cariporide and other new and powerful NHE1 inhibitors as potentially selective anticancer drugs--an integral molecular/biochemical/metabolic/clinical approach after one hundred years of cancer research. J. Transl. Med. 2013, 11, 282. [Google Scholar] [CrossRef]
- Harguindey, S.; Orive, G.; Pedraz, J.L.; Bello, G.; Arranz, J.L.; Samaniego, J.M. Apparent cure of a case of metastatic ovarian carcinoma after the chronic treatment with Na+H+ antiport inhibitors. Oncologia 2002, 25, 62–66. [Google Scholar]
- He, B.; Zhang, M.; Zhu, R. Na+/H+ exchanger blockade inhibits the expression of vascular endothelial growth factor in SGC7901 cells. Oncol. Rep. 2010, 23, 79–87. [Google Scholar] [CrossRef]
- Evans, D.M.; Sloan-Stakleff, K.; Arvan, M.; Guyton, D.P. Time and dose dependency of the suppression of pulmonary metastases of rat mammary cancer by amiloride. Clin. Exp. Metastasis 1998, 16, 353–357. [Google Scholar] [CrossRef]
- Di Sario, A.; Bendia, E.; Omenetti, A.; De Minicis, S.; Marzioni, M.; Kleemann, H.W.; Candelaresi, C.; Saccomanno, S.; Alpini, G.; Benedetti, A. Selective inhibition of ion transport mechanisms regulating intracellular pH reduces proliferation and induces apoptosis in cholangiocarcinoma cells. Dig. Liver Dis. 2007, 39, 60–69. [Google Scholar] [CrossRef]
- Kellen, J.A.; Mirakian, A.; Kolin, A. Antimetastatic effect of amiloride in an animal tumour model. Anticancer Res. 1988, 8, 1373–1376. [Google Scholar]
- Porporato, P.E.; Dhup, S.; Dadhich, R.K.; Copetti, T.; Sonveaux, P. Anticancer targets in the glycolytic metabolism of tumors: A comprehensive review. Front. Pharmacol. 2011, 2, 49. [Google Scholar] [CrossRef] [PubMed]
- Luciani, F.; Spada, M.; De Milito, A.; Molinari, A.; Rivoltini, L.; Montinaro, A.; Marra, M.; Lugini, L.; Logozzi, M.; Lozupone, F.; et al. Effect of proton pump inhibitor pretreatment on resistance of solid tumors to cytotoxic drugs. J. Natl. Cancer Inst. 2004, 96, 1702–1713. [Google Scholar] [CrossRef] [PubMed]
- Atwal, K.S.; O’Neil, S.V.; Ahmad, S.; Doweyko, L.; Kirby, M.; Dorso, C.R.; Chandrasena, G.; Chen, B.C.; Zhao, R.; Zahler, R. Synthesis and biological activity of 5-aryl-4-(4-(5-methyl-1H-imidazol-4-yl)piperidin-1-yl)pyrimidine analogs as potent, highly selective, and orally bioavailable NHE-1 inhibitors. Bioorg. Med. Chem. Lett. 2006, 16, 4796–4799. [Google Scholar] [CrossRef] [PubMed]
- Tavares-Valente, D.; Baltazar, F.; Moreira, R.; Queiros, O. Cancer cell bioenergetics and pH regulation influence breast cancer cell resistance to paclitaxel and doxorubicin. J. Bioenerg. Biomembr. 2013, 45, 467–475. [Google Scholar] [CrossRef]
- Reshkin, S.J.; Bellizzi, A.; Cardone, R.A.; Tommasino, M.; Casavola, V.; Paradiso, A. Paclitaxel induces apoptosis via protein kinase A- and p38 mitogen-activated protein-dependent inhibition of the Na+/H+ exchanger (NHE) NHE isoform 1 in human breast cancer cells. Clin. Cancer Res. 2003, 9, 2366–2373. [Google Scholar]
- Granja, S.; Tavares-Valente, D.; Queiros, O.; Baltazar, F. Value of pH regulators in the diagnosis, prognosis and treatment of cancer. Semin. Cancer Biol. 2017, 43, 17–34. [Google Scholar] [CrossRef]
- Xiao-Fang, C.; Zheng, C.-L.; Akiyama, S.-I.; Tomoda, A. 2-Aminophenoxazine-3-one and 2-amino-4, 4α-dihydro-4α, 7-dimethyl-3H-phenoxazine-3-one cause cellular apoptosis by reducing higher intracellular pH in cancer cells. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2011, 87, 199. [Google Scholar] [CrossRef]
- Supuran, C.T. Carbonic Anhydrases and Metabolism. Metabolites 2018, 8. [Google Scholar] [CrossRef]
- Spugnini, E.P.; Buglioni, S.; Carocci, F.; Francesco, M.; Vincenzi, B.; Fanciulli, M.; Fais, S. High dose lansoprazole combined with metronomic chemotherapy: A phase I/II study in companion animals with spontaneously occurring tumors. J. Transl. Med. 2014, 12, 225. [Google Scholar] [CrossRef]
- Spugnini, E.; Fais, S. Proton pump inhibition and cancer therapeutics: A specific tumor targeting or it is a phenomenon secondary to a systemic buffering? Semin. Cancer Biol. 2017, 43, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.N.; Tian, B.; Guo, X.L. Repositioning of proton pump inhibitors in cancer therapy. Cancer Chemother. Pharmacol. 2017, 80, 925–937. [Google Scholar] [CrossRef]
- Wang, B.Y.; Zhang, J.; Wang, J.L.; Sun, S.; Wang, Z.H.; Wang, L.P.; Zhang, Q.L.; Lv, F.F.; Cao, E.Y.; Shao, Z.M.; et al. Intermittent high dose proton pump inhibitor enhances the antitumor effects of chemotherapy in metastatic breast cancer. J. Exp. Clin. Cancer Res. 2015, 34, 85. [Google Scholar] [CrossRef] [PubMed]
- Azzarito, T.; Venturi, G.; Cesolini, A.; Fais, S. Lansoprazole induces sensitivity to suboptimal doses of paclitaxel in human melanoma. Cancer Lett. 2015, 356, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Niikura, K. Effect of a V-ATPase inhibitor, FR202126, in syngeneic mouse model of experimental bone metastasis. Cancer Chemother. Pharmacol. 2007, 60, 555–562. [Google Scholar] [CrossRef] [PubMed]
- De Milito, A.; Iessi, E.; Logozzi, M.; Lozupone, F.; Spada, M.; Marino, M.L.; Federici, C.; Perdicchio, M.; Matarrese, P.; Lugini, L.; et al. Proton pump inhibitors induce apoptosis of human B-cell tumors through a caspase-independent mechanism involving reactive oxygen species. Cancer Res. 2007, 67, 5408–5417. [Google Scholar] [CrossRef] [PubMed]
- De Milito, A.; Canese, R.; Marino, M.L.; Borghi, M.; Iero, M.; Villa, A.; Venturi, G.; Lozupone, F.; Iessi, E.; Logozzi, M.; et al. pH-dependent antitumor activity of proton pump inhibitors against human melanoma is mediated by inhibition of tumor acidity. Int. J. Cancer 2010, 127, 207–219. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, Y.; Li, S.J. Lansoprazole induces apoptosis of breast cancer cells through inhibition of intracellular proton extrusion. Biochem. Biophys. Res. Commun. 2014, 448, 424–429. [Google Scholar] [CrossRef]
- Spugnini, E.P.; Baldi, A.; Buglioni, S.; Carocci, F.; de Bazzichini, G.M.; Betti, G.; Pantaleo, I.; Menicagli, F.; Citro, G.; Fais, S. Lansoprazole as a rescue agent in chemoresistant tumors: A phase I/II study in companion animals with spontaneously occurring tumors. J. Transl. Med. 2011, 9, 221. [Google Scholar] [CrossRef]
- Ding, D.C.; Sung, F.C.; Chen, W.; Wang, J.H.; Lin, S.Z. Proton pump inhibitors reduce breast cancer risk in gastric ulcer patients: A population-based cohort study. Breast J. 2019. [Google Scholar] [CrossRef]
- Lee, Y.Y.; Jeon, H.K.; Hong, J.E.; Cho, Y.J.; Ryu, J.Y.; Choi, J.J.; Lee, S.H.; Yoon, G.; Kim, W.Y.; Do, I.G.; et al. Proton pump inhibitors enhance the effects of cytotoxic agents in chemoresistant epithelial ovarian carcinoma. Oncotarget 2015, 6, 35040–35050. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Perut, F.; Fagioli, F.; Brach Del Prever, A.; Meazza, C.; Parafioriti, A.; Picci, P.; Gambarotti, M.; Avnet, S.; Baldini, N.; et al. Proton pump inhibitor chemosensitization in human osteosarcoma: From the bench to the patients’ bed. J. Transl. Med. 2013, 11, 268. [Google Scholar] [CrossRef] [PubMed]
- Payen, V.L.; Mina, E.; Van Hee, V.F.; Porporato, P.E.; Sonveaux, P. Monocarboxylate transporters in cancer. Mol. Metab. 2019. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Gao, Z.; Hu, X.; Xiang, F.; Wu, Z.; Zhang, J.; Han, X.; Yin, L.; Qin, J.; Lan, L. Downregulation of MCT 4 for lactate exchange promotes the cytotoxicity of NK cells in breast carcinoma. Cancer Med. 2018, 7, 4690–4700. [Google Scholar] [CrossRef]
- Nath, K.; Guo, L.; Nancolas, B.; Nelson, D.S.; Shestov, A.A.; Lee, S.C.; Roman, J.; Zhou, R.; Leeper, D.B.; Halestrap, A.P.; et al. Mechanism of antineoplastic activity of lonidamine. Biochim. Biophys. Acta 2016, 1866, 151–162. [Google Scholar] [CrossRef]
- Mboge, M.Y.; Chen, Z.; Wolff, A.; Mathias, J.V.; Tu, C.; Brown, K.D.; Bozdag, M.; Carta, F.; Supuran, C.T.; McKenna, R.; et al. Selective inhibition of carbonic anhydrase IX over carbonic anhydrase XII in breast cancer cells using benzene sulfonamides: Disconnect between activity and growth inhibition. PLoS ONE 2018, 13, e0207417. [Google Scholar] [CrossRef]
- Bartosova, M.; Parkkila, S.; Pohlodek, K.; Karttunen, T.J.; Galbavy, S.; Mucha, V.; Harris, A.L.; Pastorek, J.; Pastorekova, S. Expression of carbonic anhydrase IX in breast is associated with malignant tissues and is related to overexpression of c-erbB2. J. Pathol. 2002, 197, 314–321. [Google Scholar] [CrossRef]
- Lock, F.E.; McDonald, P.C.; Lou, Y.; Serrano, I.; Chafe, S.C.; Ostlund, C.; Aparicio, S.; Winum, J.Y.; Supuran, C.T.; Dedhar, S. Targeting carbonic anhydrase IX depletes breast cancer stem cells within the hypoxic niche. Oncogene 2013, 32, 5210–5219. [Google Scholar] [CrossRef]
- Meehan, J.; Ward, C.; Turnbull, A.; Bukowski-Wills, J.; Finch, A.J.; Jarman, E.J.; Xintaropoulou, C.; Martinez-Perez, C.; Gray, M.; Pearson, M.; et al. Inhibition of pH regulation as a therapeutic strategy in hypoxic human breast cancer cells. Oncotarget 2017, 8, 42857–42875. [Google Scholar] [CrossRef]
- Keizer, H.G.; Joenje, H. Increased cytosolic pH in multidrug-resistant human lung tumor cells: Effect of verapamil. J. Natl. Cancer Inst. 1989, 81, 706–709. [Google Scholar] [CrossRef]
- Sohn, J.H.; Kim, Y.T.; Rha, S.Y.; Yoo, N.C.; Roh, J.K.; Kim, B.S.; Suh, C.O.; Kim, G.E.; Jang, W.I.; Chung, H.C. Paclitaxel and Cisplatin Combination Chemotherapy in Pretreated Breast Cancer. Cancer Res. Treat. 2003, 35, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Elserafi, M.M.; Zeeneldin, A.A.; Abdelsalam, I.M.; Nassar, H.R.; Moneer, M.M.; Buhoush, W.H. First-line paclitaxel and cisplatin used sequentially or in combination in metastatic breast cancer: A phase II randomized study. J. Egypt. Natl. Cancer Inst. 2018, 30, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Shetti, D.; Zhang, B.; Fan, C.; Mo, C.; Lee, B.H.; Wei, K. Low Dose of Paclitaxel Combined with XAV939 Attenuates Metastasis, Angiogenesis and Growth in Breast Cancer by Suppressing Wnt Signaling. Cells 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Cardone, R.A.; Greco, M.R.; Zeeberg, K.; Zaccagnino, A.; Saccomano, M.; Bellizzi, A.; Bruns, P.; Menga, M.; Pilarsky, C.; Schwab, A.; et al. A novel NHE1-centered signaling cassette drives epidermal growth factor receptor-dependent pancreatic tumor metastasis and is a target for combination therapy. Neoplasia 2015, 17, 155–166. [Google Scholar] [CrossRef]
- Raudenska, M.; Balvan, J.; Fojtu, M.; Gumulec, J.; Masarik, M. Unexpected therapeutic effects of cisplatin. Metallomics 2019, 11, 1182–1199. [Google Scholar] [CrossRef]
- Rosenberg, B. Cisplatin: Its history and possible mechanisms of action. In Cisplatin; Elsevier: Cambridge, MA, USA, 1980; pp. 9–20. [Google Scholar] [CrossRef]
- Makovec, T. Cisplatin and beyond: Molecular mechanisms of action and drug resistance development in cancer chemotherapy. Radiol. Oncol. 2019, 53, 148–158. [Google Scholar] [CrossRef]
- Chen, Z.; Ai, L.; Mboge, M.Y.; Tu, C.; McKenna, R.; Brown, K.D.; Heldermon, C.D.; Frost, S.C. Differential expression and function of CAIX and CAXII in breast cancer: A comparison between tumorgraft models and cells. PLoS ONE 2018, 13, e0199476. [Google Scholar] [CrossRef]
- Dauchy, R.T.; Xiang, S.; Mao, L.; Brimer, S.; Wren, M.A.; Yuan, L.; Anbalagan, M.; Hauch, A.; Frasch, T.; Rowan, B.G.; et al. Circadian and melatonin disruption by exposure to light at night drives intrinsic resistance to tamoxifen therapy in breast cancer. Cancer Res. 2014, 74, 4099–4110. [Google Scholar] [CrossRef]
- Lacerda, J.Z.; Ferreira, L.C.; Lopes, B.C.; Aristizabal-Pachon, A.F.; Bajgelman, M.C.; Borin, T.F.; Zuccari, D. Therapeutic Potential of Melatonin in the Regulation of MiR-148a-3p and Angiogenic Factors in Breast Cancer. Microrna 2019, 8, 237–247. [Google Scholar] [CrossRef]
- Hill, S.M.; Belancio, V.P.; Dauchy, R.T.; Xiang, S.; Brimer, S.; Mao, L.; Hauch, A.; Lundberg, P.W.; Summers, W.; Yuan, L.; et al. Melatonin: An inhibitor of breast cancer. Endocr. Relat. Cancer 2015, 22, R183–R204. [Google Scholar] [CrossRef]
- Mediavilla, M.D.; Sanchez-Barcelo, E.J.; Tan, D.X.; Manchester, L.; Reiter, R.J. Basic mechanisms involved in the anti-cancer effects of melatonin. Curr. Med. Chem. 2010, 17, 4462–4481. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.; Marzouk, M.A.; Adhikari, S.; Wright, T.D.; Miller, B.P.; Matossian, M.D.; Elliott, S.; Wright, M.; Alzoubi, M.; Collins-Burow, B.M.; et al. Pharmacological, Mechanistic, and Pharmacokinetic Assessment of Novel Melatonin-Tamoxifen Drug Conjugates as Breast Cancer Drugs. Mol. Pharmacol. 2019, 96, 272–296. [Google Scholar] [CrossRef] [PubMed]
- De Castro, T.B.; Bordin-Junior, N.A.; de Almeida, E.A.; de Campos Zuccari, D.A.P. Evaluation of melatonin and AFMK levels in women with breast cancer. Endocrine 2018, 62, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Nooshinfar, E.; Safaroghli-Azar, A.; Bashash, D.; Akbari, M.E. Melatonin, an inhibitory agent in breast cancer. Breast Cancer 2017, 24, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Sonehara, N.M.; Lacerda, J.Z.; Jardim-Perassi, B.V.; de Paula, R., Jr.; Moschetta-Pinheiro, M.G.; Souza, Y.S.T.; de Andrade, J.C.J.; De Campos Zuccari, D.A.P. Melatonin regulates tumor aggressiveness under acidosis condition in breast cancer cell lines. Oncol. Lett. 2019, 17, 1635–1645. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-García, V.; González, A.; Alonso-González, C.; Martínez-Campa, C.; Cos, S. Regulation of vascular endothelial growth factor by melatonin in human breast cancer cells. J. Pineal Res. 2013, 54, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Rosales-Corral, S.A.; Tan, D.X.; Acuna-Castroviejo, D.; Qin, L.; Yang, S.F.; Xu, K. Melatonin, a Full Service Anti-Cancer Agent: Inhibition of Initiation, Progression and Metastasis. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef]
- Menéndez-Menéndez, J.; Hermida-Prado, F.; Granda-Díaz, R.; González, A.; García-Pedrero, J.M.; Del-Río-Ibisate, N.; González-González, A.; Cos, S.; Alonso-González, C.; Martínez-Campa, C. Deciphering the molecular basis of melatonin protective effects on breast cells treated with doxorubicin: TWIST1 a transcription factor involved in EMT and metastasis, a novel target of melatonin. Cancers (Basel) 2019, 11, 1011. [Google Scholar] [CrossRef]
- Borin, T.F.; Arbab, A.S.; Gelaleti, G.B.; Ferreira, L.C.; Moschetta, M.G.; Jardim-Perassi, B.V.; Iskander, A.S.; Varma, N.R.; Shankar, A.; Coimbra, V.B.; et al. Melatonin decreases breast cancer metastasis by modulating Rho-associated kinase protein-1 expression. J. Pineal Res. 2016, 60, 3–15. [Google Scholar] [CrossRef]
- Jardim-Perassi, B.V.; Lourenco, M.R.; Doho, G.M.; Grigolo, I.H.; Gelaleti, G.B.; Ferreira, L.C.; Borin, T.F.; Moschetta, M.G.; Pires de Campos Zuccari, D.A. Melatonin Regulates Angiogenic Factors under Hypoxia in Breast Cancer Cell Lines. Anticancer Agents Med. Chem. 2016, 16, 347–358. [Google Scholar] [CrossRef]
- Chottanapund, S.; Van Duursen, M.B.; Navasumrit, P.; Hunsonti, P.; Timtavorn, S.; Ruchirawat, M.; Van den Berg, M. Anti-aromatase effect of resveratrol and melatonin on hormonal positive breast cancer cells co-cultured with breast adipose fibroblasts. Toxicol. In Vitro 2014, 28, 1215–1221. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.H.; Ho, C.T.; Chen, Z.F.; Chen, L.C.; Whang-Peng, J.; Lin, T.N.; Ho, Y.S. The apple polyphenol phloretin inhibits breast cancer cell migration and proliferation via inhibition of signals by type 2 glucose transporter. J. Food Drug Anal. 2018, 26, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Meiners, C. Clinical Response of Metastatic Breast Cancer to Multi-targeted Therapeutic Approach: A Single Case Report. Cancers (Basel) 2011, 3, 1454–1466. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; Chao, T.K.; Chang, C.C.; Yo, Y.T.; Yu, M.H.; Lai, H.C. Drug screening identifies niclosamide as an inhibitor of breast cancer stem-like cells. PLoS ONE 2013, 8, e74538. [Google Scholar] [CrossRef]
- Pronzato, P.; Amoroso, D.; Bertelli, G.; Conte, P.F.; Cusimano, M.P.; Ciottoli, G.B.; Gulisano, M.; Lionetto, R.; Rosso, R. Phase II study of lonidamine in metastatic breast cancer. Br. J. Cancer 1989, 59, 251–253. [Google Scholar] [CrossRef]
- Bougnoux, P.; Hajjaji, N.; Ferrasson, M.N.; Giraudeau, B.; Couet, C.; Le Floch, O. Improving outcome of chemotherapy of metastatic breast cancer by docosahexaenoic acid: A phase II trial. Br. J. Cancer 2009, 101, 1978–1985. [Google Scholar] [CrossRef]
- Wannous, R.; Bon, E.; Gillet, L.; Chamouton, J.; Weber, G.; Brisson, L.; Goré, J.; Bougnoux, P.; Besson, P.; Roger, S. Suppression of PPARβ, and DHA treatment, inhibit NaV1. 5 and NHE-1 pro-invasive activities. Pflug. Arch. 2014, 467, 1249–1259. [Google Scholar] [CrossRef]
- Li, Y.; Li, P.K.; Roberts, M.J.; Arend, R.C.; Samant, R.S.; Buchsbaum, D.J. Multi-targeted therapy of cancer by niclosamide: A new application for an old drug. Cancer Lett. 2014, 349, 8–14. [Google Scholar] [CrossRef]
- Naujokat, C.; Steinhart, R. Salinomycin as a drug for targeting human cancer stem cells. J. Biomed. Biotechnol. 2012, 2012, 950658. [Google Scholar] [CrossRef]
- Lega, I.C.; Austin, P.C.; Gruneir, A.; Goodwin, P.J.; Rochon, P.A.; Lipscombe, L.L. Association between metformin therapy and mortality after breast cancer: A population-based study. Diabetes Care 2013, 36, 3018–3026. [Google Scholar] [CrossRef]
- Roshan, M.H.; Shing, Y.K.; Pace, N.P. Metformin as an adjuvant in breast cancer treatment. SAGE Open Med. 2019, 7, 2050312119865114. [Google Scholar] [CrossRef]
- Grover-McKay, M.; Walsh, S.A.; Seftor, E.A.; Thomas, P.A.; Hendrix, M.J. Role for glucose transporter 1 protein in human breast cancer. Pathol. Oncol. Res. 1998, 4, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Garrido, P.; Osorio, F.G.; Moran, J.; Cabello, E.; Alonso, A.; Freije, J.M.; Gonzalez, C. Loss of GLUT4 induces metabolic reprogramming and impairs viability of breast cancer cells. J. Cell. Physiol. 2015, 230, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, G.; Wang, Y.; Tang, S.; Sun, X.; Feng, X.; Li, Y.; Bao, G.; Li, P.; Mao, X. Suppression of tumor angiogenesis by metformin treatment via a mechanism linked to targeting of HER2/HIF-1α/VEGF secretion axis. Oncotarget 2015, 6, 44579. [Google Scholar] [CrossRef] [PubMed]
- Schexnayder, C.; Broussard, K.; Onuaguluchi, D.; Poche, A.; Ismail, M.; McAtee, L.; Llopis, S.; Keizerweerd, A.; McFerrin, H.; Williams, C. Metformin Inhibits Migration and Invasion by Suppressing ROS Production and COX2 Expression in MDA-MB-231 Breast Cancer Cells. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Wang, Y.; Liu, Z.; Sun, Y.; Wang, X.; Wei, G.; Wei, J. Metformin exerts anticancer effects through the inhibition of the Sonic hedgehog signaling pathway in breast cancer. Int. J. Mol. Med. 2015, 36, 204–214. [Google Scholar] [CrossRef]
- Riobo-Del Galdo, N.A.; Lara Montero, A.; Wertheimer, E.V. Role of Hedgehog Signaling in Breast Cancer: Pathogenesis and Therapeutics. Cells 2019, 8. [Google Scholar] [CrossRef]
- Iliopoulos, D.; Hirsch, H.A.; Struhl, K. Metformin decreases the dose of chemotherapy for prolonging tumor remission in mouse xenografts involving multiple cancer cell types. Cancer Res. 2011, 71, 3196–3201. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Martin, A.; Oliveras-Ferraros, C.; Del Barco, S.; Martin-Castillo, B.; Menendez, J.A. The anti-diabetic drug metformin suppresses self-renewal and proliferation of trastuzumab-resistant tumor-initiating breast cancer stem cells. Breast Cancer Res. Treat. 2011, 126, 355–364. [Google Scholar] [CrossRef]
- Leone, A.; Di Gennaro, E.; Bruzzese, F.; Avallone, A.; Budillon, A. New perspective for an old antidiabetic drug: Metformin as anticancer agent. Cancer Treat. Res. 2014, 159, 355–376. [Google Scholar] [CrossRef]
- De, A.; Kuppusamy, G. Metformin in breast cancer: Preclinical and clinical evidence. Curr. Probl. Cancer 2019. [Google Scholar] [CrossRef] [PubMed]
- Bayraktar, S.; Hernadez-Aya, L.F.; Lei, X.; Meric-Bernstam, F.; Litton, J.K.; Hsu, L.; Hortobagyi, G.N.; Gonzalez-Angulo, A.M. Effect of metformin on survival outcomes in diabetic patients with triple receptor-negative breast cancer. Cancer 2012, 118, 1202–1211. [Google Scholar] [CrossRef] [PubMed]
- Qu, H.; Yang, X. Metformin inhibits angiogenesis induced by interaction of hepatocellular carcinoma with hepatic stellate cells. Cell Biochem. Biophys. 2015, 71, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes, T.A.; Farias, L.C.; Santos, E.S.; de Carvalho Fraga, C.A.; Orsini, L.A.; de Freitas Teles, L.; Feltenberger, J.D.; de Jesus, S.F.; de Souza, M.G.; Santos, S.H.; et al. Metformin increases PDH and suppresses HIF-1alpha under hypoxic conditions and induces cell death in oral squamous cell carcinoma. Oncotarget 2016, 7, 55057–55068. [Google Scholar] [CrossRef]
- Han, J.; Li, Y.; Liu, X.; Zhou, T.; Sun, H.; Edwards, P.; Gao, H.; Yu, F.S.; Qiao, X. Metformin suppresses retinal angiogenesis and inflammation in vitro and in vivo. PLoS ONE 2018, 13, e0193031. [Google Scholar] [CrossRef]
- Ni, H.Z.; Liu, Z.; Sun, L.L.; Zhou, M.; Liu, C.; Li, W.D.; Li, X.Q. Metformin inhibits angiogenesis of endothelial progenitor cells via miR-221-mediated p27 expression and autophagy. Future Med. Chem. 2019, 11, 2263–2272. [Google Scholar] [CrossRef]
- Blagosklonny, M.V. Hypoxia-inducible factor: Achilles’ heel of antiangiogenic cancer therapy (review). Int. J. Oncol. 2001, 19, 257–262. [Google Scholar] [CrossRef]
- De Francesco, E.M.; Lappano, R.; Santolla, M.F.; Marsico, S.; Caruso, A.; Maggiolini, M. HIF-1alpha/GPER signaling mediates the expression of VEGF induced by hypoxia in breast cancer associated fibroblasts (CAFs). Breast Cancer Res. 2013, 15, R64. [Google Scholar] [CrossRef]
- Briggs, K.J.; Koivunen, P.; Cao, S.; Backus, K.M.; Olenchock, B.A.; Patel, H.; Zhang, Q.; Signoretti, S.; Gerfen, G.J.; Richardson, A.L.; et al. Paracrine Induction of HIF by Glutamate in Breast Cancer: EglN1 Senses Cysteine. Cell 2016, 166, 126–139. [Google Scholar] [CrossRef]
- Song, C.W.; Lee, H.; Dings, R.P.; Williams, B.; Powers, J.; Santos, T.D.; Choi, B.H.; Park, H.J. Metformin kills and radiosensitizes cancer cells and preferentially kills cancer stem cells. Sci. Rep. 2012, 2, 362. [Google Scholar] [CrossRef]
- Turashvili, G.; Brogi, E. Tumor Heterogeneity in Breast Cancer. Front. Med. 2017, 4, 227. [Google Scholar] [CrossRef] [PubMed]
- Waks, A.G.; Winer, E.P. Breast Cancer Treatment: A Review. JAMA 2019, 321, 288–300. [Google Scholar] [CrossRef]
- Puddefoot, J.; Baker, V.; Bakkers, B.; Marsigliante, S.; Barker, S.; Panahy, C.; Goode, A.; Carpenter, R.; Vinson, G. The nature and significance of multiple isoforms of the oestrogen receptor in breast tumours. J. Mol. Endocrinol. 1993, 11, 83–90. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Leclercq, G. Molecular forms of the estrogen receptor in breast cancer. J. Steroid Biochem. Mol. Biol. 2002, 80, 259–272. [Google Scholar] [CrossRef]
- Brahimi-Horn, C.; Pouyssegur, J. The role of the hypoxia-inducible factor in tumor metabolism growth and invasion. Bull. Cancer 2006, 93, E73–E80. [Google Scholar]
- Williams, B.; Howard, R.L. Glucose-induced changes in Na+/H+ antiport activity and gene expression in cultured vascular smooth muscle cells. Role of protein kinase C. J. Clin. Invest. 1994, 93, 2623–2631. [Google Scholar] [CrossRef]
- Hawsawi, Y.M.; Al-Numair, N.S.; Sobahy, T.M.; Al-Ajmi, A.M.; Al-Harbi, R.M.; Baghdadi, M.A.; Oyouni, A.A.; Alamer, O.M. The role of BRCA1/2 in hereditary and familial breast and ovarian cancers. Mol. Genet. Genomic Med. 2019, 7, e879. [Google Scholar] [CrossRef]
- Taherian-Esfahani, Z.; Taheri, M.; Dashti, S.; Kholghi-Oskooei, V.; Geranpayeh, L.; Ghafouri-Fard, S. Assessment of the expression pattern of mTOR-associated lncRNAs and their genomic variants in the patients with breast cancer. J. Cell. Physiol. 2019, 234, 22044–22056. [Google Scholar] [CrossRef]
- Nijhout, H.F. Metaphors and the role of genes in development. Bioessays 1990, 12, 441–446. [Google Scholar] [CrossRef]
- Lim, B.; Woodward, W.A.; Wang, X.; Reuben, J.M.; Ueno, N.T. Inflammatory breast cancer biology: The tumour microenvironment is key. Nat. Rev. Cancer 2018, 18, 485. [Google Scholar] [CrossRef]
- Barriere, H.; Poujeol, C.; Tauc, M.; Blasi, J.M.; Counillon, L.; Poujeol, P. CFTR modulates programmed cell death by decreasing intracellular pH in Chinese hamster lung fibroblasts. Am. J. Physiol. Cell Physiol. 2001, 281, C810–C824. [Google Scholar] [CrossRef]
- Harguindey, S.; Gonzalez Molinillo, J.; Chinchilla, D.; Reshkin, S.; Tomoda, A. Further Along a Clinical Protocol Using a Cocktail of PTIs in Human Cancer. In Proceedings of the 2nd ISPD Meeting, Nice, France, 18–19 November 2011. [Google Scholar]
- Matthews, H.; Ranson, M.; Kelso, M.J. Anti-tumour/metastasis effects of the potassium-sparing diuretic amiloride: An orally active anti-cancer drug waiting for its call-of-duty? Int. J. Cancer 2011, 129, 2051–2061. [Google Scholar] [CrossRef] [PubMed]
- Harguindey, S. Use of Na+/H+ antiporter inhibitors as a novel approach to cancer treatment. In Amiloride and its Analogs: Unique Cation Transport Inhibitors; VCH Publishers Inc.: New York, NY, USA, 1992; pp. 317–334. [Google Scholar]
- Rath, S.; Liebl, J.; Furst, R.; Vollmar, A.M.; Zahler, S. Regulation of endothelial signaling and migration by v-ATPase. Angiogenesis 2014, 17, 587–601. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, C.; Wang, J.; Fan, Y.; Wang, Z.; Wang, Y. Proton pump inhibitors increase the chemosensitivity of patients with advanced colorectal cancer. Oncotarget 2017, 8, 58801–58808. [Google Scholar] [CrossRef] [PubMed]
- Avnet, S.; Lemma, S.; Cortini, M.; Pellegrini, P.; Perut, F.; Zini, N.; Kusuzaki, K.; Chano, T.; Grisendi, G.; Dominici, M.; et al. Altered pH gradient at the plasma membrane of osteosarcoma cells is a key mechanism of drug resistance. Oncotarget 2016, 7, 63408–63423. [Google Scholar] [CrossRef] [PubMed]
- Canitano, A.; Iessi, E.; Spugnini, E.P.; Federici, C.; Fais, S. Proton pump inhibitors induce a caspase-independent antitumor effect against human multiple myeloma. Cancer Lett. 2016, 376, 278–283. [Google Scholar] [CrossRef]
- Papagerakis, S.; Bellile, E.; Peterson, L.A.; Pliakas, M.; Balaskas, K.; Selman, S.; Hanauer, D.; Taylor, J.M.; Duffy, S.; Wolf, G. Proton pump inhibitors and histamine 2 blockers are associated with improved overall survival in patients with head and neck squamous carcinoma. Cancer Prev. Res. 2014, 7, 1258–1269. [Google Scholar] [CrossRef]
- Falcone, R.; Roberto, M.; D’Antonio, C.; Romiti, A.; Milano, A.; Onesti, C.E.; Marchetti, P.; Fais, S. High-doses of proton pump inhibitors in refractory gastro-intestinal cancer: A case series and the state of art. Dig. Liver Dis. 2016, 48, 1503–1505. [Google Scholar] [CrossRef]
- Ikemura, K.; Hiramatsu, S.; Okuda, M. Drug Repositioning of Proton Pump Inhibitors for Enhanced Efficacy and Safety of Cancer Chemotherapy. Front. Pharmacol. 2017, 8, 911. [Google Scholar] [CrossRef]
- Fako, V.E.; Wu, X.; Pflug, B.; Liu, J.Y.; Zhang, J.T. Repositioning proton pump inhibitors as anticancer drugs by targeting the thioesterase domain of human fatty acid synthase. J. Med. Chem. 2015, 58, 778–784. [Google Scholar] [CrossRef]
- Sanchez-Sanchez, A.M.; Antolin, I.; Puente-Moncada, N.; Suarez, S.; Gomez-Lobo, M.; Rodriguez, C.; Martin, V. Melatonin cytotoxicity is associated to warburg effect inhibition in ewing sarcoma cells. PLoS ONE 2015, 10, e0135420. [Google Scholar] [CrossRef] [PubMed]
- De Lima Mota, A.; Jardim-Perassi, B.V.; de Castro, T.B.; Colombo, J.; Sonehara, N.M.; Nishiyama, V.K.G.; Pierri, V.A.G.; de Campos Zuccari, D.A.P. Melatonin modifies tumor hypoxia and metabolism by inhibiting HIF-1α and energy metabolic pathway in the in vitro and in vivo models of breast cancer. Melatonin Res. 2019, 2, 83–98. [Google Scholar] [CrossRef]
- Turner, N.C.; Tutt, A.N. Platinum chemotherapy for BRCA1-related breast cancer: Do we need more evidence? Breast Cancer Res. 2012, 14, 115. [Google Scholar] [CrossRef]
- Dasari, S.; Tchounwou, P.B. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef]
- Shirmanova, M.V.; Druzhkova, I.N.; Lukina, M.M.; Dudenkova, V.V.; Ignatova, N.I.; Snopova, L.B.; Shcheslavskiy, V.I.; Belousov, V.V.; Zagaynova, E.V. Chemotherapy with cisplatin: Insights into intracellular pH and metabolic landscape of cancer cells in vitro and in vivo. Sci. Rep. 2017, 7, 8911. [Google Scholar] [CrossRef]
- Boscoboinik, D.; Gupta, R.S.; Epand, R.M. Investigation of the relationship between altered intracellular pH and multidrug resistance in mammalian cells. Br. J. Cancer 1990, 61, 568–572. [Google Scholar] [CrossRef]
- Epand, R.F.; Epand, R.M.; Gupta, R.S.; Cragoe, E.J., Jr. Reversal of intrinsic multidrug resistance in Chinese hamster ovary cells by amiloride analogs. Br. J. Cancer 1991, 63, 247–251. [Google Scholar] [CrossRef][Green Version]
- Omran, Z.; Scaife, P.; Stewart, S.; Rauch, C. Physical and biological characteristics of multi drug resistance (MDR): An integral approach considering pH and drug resistance in cancer. Semin. Cancer Biol. 2017, 43, 42–48. [Google Scholar] [CrossRef]
- Alfarouk, K.O.; Stock, C.M.; Taylor, S.; Walsh, M.; Muddathir, A.K.; Verduzco, D.; Bashir, A.H.; Mohammed, O.Y.; Elhassan, G.O.; Harguindey, S.; et al. Resistance to cancer chemotherapy: Failure in drug response from ADME to P-gp. Cancer Cell Int. 2015, 15, 71. [Google Scholar] [CrossRef]
- Wei, L.Y.; Roepe, P.D. Low external pH and osmotic shock increase the expression of human MDR protein. Biochemistry 1994, 33, 7229–7238. [Google Scholar] [CrossRef]
- Jin, W.; Li, Q.; Lin, Y.; Lu, Y.; Li, H.; Wang, L.; Hu, R.; Ma, L.; Wang, J.; Pang, T. Reversal of Imatinib resistance in BCR-ABL-positive leukemia after inhibition of the Na+/H+ exchanger. Cancer Lett. 2011, 308, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.H.; Lu, Y.; Jin, W.N.; Lin, Y.N.; Hu, R.H.; Zhu, X.F.; Wang, J.X.; Pang, T.X. Effect of intracellular acidification on drug resistance of leukemia cells with high P-glycoprotein expression. Zhonghua Xue Ye Xue Za Zhi 2009, 30, 605–609. [Google Scholar] [PubMed]
- Gallagher Ferdia, A.; Kettunen Mikko, I.; Day Sam, E. Magnetic resonance imaging of pH in vivo using hyperpolarized 13C labelled bicarbonate. Nature 2008, 453, 940–944. [Google Scholar] [CrossRef]
- Lemma, S.; Di Pompo, G.; Porporato, P.E.; Sboarina, M.; Russell, S.; Gillies, R.J.; Baldini, N.; Sonveaux, P.; Avnet, S. MDA-MB-231 breast cancer cells fuel osteoclast metabolism and activity: A new rationale for the pathogenesis of osteolytic bone metastases. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 3254–3264. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.O.; Lee, S.K.; Kim, J.H.; Kim, N.; You, G.Y.; Moon, J.W.; Kim, S.J.; Park, S.H.; Kim, H.S. Metformin regulates glucose transporter 4 (GLUT4) translocation through AMP-activated protein kinase (AMPK)-mediated Cbl/CAP signaling in 3T3-L1 preadipocyte cells. J. Biol. Chem. 2012, 287, 44121–44129. [Google Scholar] [CrossRef] [PubMed]
- Barron, C.; Tsiani, E.; Tsakiridis, T. Expression of the glucose transporters GLUT1, GLUT3, GLUT4 and GLUT12 in human cancer cells. BMC Proc. 2012. [Google Scholar] [CrossRef]
- Adekola, K.; Rosen, S.T.; Shanmugam, M. Glucose transporters in cancer metabolism. Curr. Opin. Oncol. 2012, 24, 650–654. [Google Scholar] [CrossRef]
- Chen, C.; Lou, T. Hypoxia inducible factors in hepatocellular carcinoma. Oncotarget 2017, 8, 46691–46703. [Google Scholar] [CrossRef]
- Dallaglio, K.; Bruno, A.; Cantelmo, A.R.; Esposito, A.I.; Ruggiero, L.; Orecchioni, S.; Calleri, A.; Bertolini, F.; Pfeffer, U.; Noonan, D.M.; et al. Paradoxic effects of metformin on endothelial cells and angiogenesis. Carcinogenesis 2014, 35, 1055–1066. [Google Scholar] [CrossRef]
- Zheng, L.; Yang, W.; Wu, F.; Wang, C.; Yu, L.; Tang, L.; Qiu, B.; Li, Y.; Guo, L.; Wu, M.; et al. Prognostic significance of AMPK activation and therapeutic effects of metformin in hepatocellular carcinoma. Clin. Cancer Res. 2013, 19, 5372–5380. [Google Scholar] [CrossRef]
- Incio, J.; Suboj, P.; Chin, S.M.; Vardam-Kaur, T.; Liu, H.; Hato, T.; Babykutty, S.; Chen, I.; Deshpande, V.; Jain, R.K.; et al. Metformin Reduces Desmoplasia in Pancreatic Cancer by Reprogramming Stellate Cells and Tumor-Associated Macrophages. PLoS ONE 2015, 10, e0141392. [Google Scholar] [CrossRef] [PubMed]
- Phoenix, K.N.; Vumbaca, F.; Claffey, K.P. Therapeutic metformin/AMPK activation promotes the angiogenic phenotype in the ERalpha negative MDA-MB-435 breast cancer model. Breast Cancer Res. Treat. 2009, 113, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.J.; Zhang, P.; Li, H.H.; Jiang, Z.W.; Jiang, C.C.; Liu, H. Cisplatin combined with metformin inhibits migration and invasion of human nasopharyngeal carcinoma cells by regulating E-cadherin and MMP-9. Asian Pac. J. Cancer Prev. 2014, 15, 4019–4023. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Cui, H.; Kang, L.; Zhang, X.; Jin, Z.; Lu, L.; Fan, Z. Metformin inhibits thyroid cancer cell growth, migration, and EMT through the mTOR pathway. Tumor Biol. 2015, 36, 6295–6304. [Google Scholar] [CrossRef]
- Son, T.X.; Huyen, N.T.B.; Saimuang, K.; Prachayasittikul, V.; On, W.C. Metformin inhibits migration and invasion of cholangiocarcinoma cells. Asian Pac. J. Cancer Prev. 2017, 18, 473. [Google Scholar] [CrossRef]
- Ferretti, A.C.; Hidalgo, F.; Tonucci, F.M.; Almada, E.; Pariani, A.; Larocca, M.C.; Favre, C. Metformin and glucose starvation decrease the migratory ability of hepatocellular carcinoma cells: Targeting AMPK activation to control migration. Sci. Rep. 2019, 9, 2815. [Google Scholar] [CrossRef]
- Biondo, L.A.; Batatinha, H.A.; Souza, C.O.; Teixeira, A.A.S.; Silveira, L.S.; Alonso-Vale, M.I.; Oyama, L.M.; Alves, M.J.; Seelaender, M.; Neto, J.C.R. Metformin Mitigates Fibrosis and Glucose Intolerance Induced by Doxorubicin in Subcutaneous Adipose Tissue. Front. Pharmacol. 2018, 9, 452. [Google Scholar] [CrossRef]
- Marino, M.L.; Fais, S.; Djavaheri-Mergny, M.; Villa, A.; Meschini, S.; Lozupone, F.; Venturi, G.; Della Mina, P.; Pattingre, S.; Rivoltini, L.; et al. Proton pump inhibition induces autophagy as a survival mechanism following oxidative stress in human melanoma cells. Cell Death Dis. 2010, 1, e87. [Google Scholar] [CrossRef]
- Cecconi, F.; Jaattela, M. Targeting ions-induced autophagy in cancer. Cancer Cell 2014, 26, 599–600. [Google Scholar] [CrossRef][Green Version]
- Lugini, L.; Matarrese, P.; Tinari, A.; Lozupone, F.; Federici, C.; Iessi, E.; Gentile, M.; Luciani, F.; Parmiani, G.; Rivoltini, L.; et al. Cannibalism of live lymphocytes by human metastatic but not primary melanoma cells. Cancer Res. 2006, 66, 3629–3638. [Google Scholar] [CrossRef]
- Fais, S.; Overholtzer, M. Cell-in-cell phenomena in cancer. Nat. Rev. Cancer 2018, 18, 758–766. [Google Scholar] [CrossRef] [PubMed]
- Haupt, S.; Keam, S.P.; Haupt, Y. Cannibalism in Breast Cancer: The Dangers of Overeating. Trends Cancer 2019, 5, 761–762. [Google Scholar] [CrossRef] [PubMed]


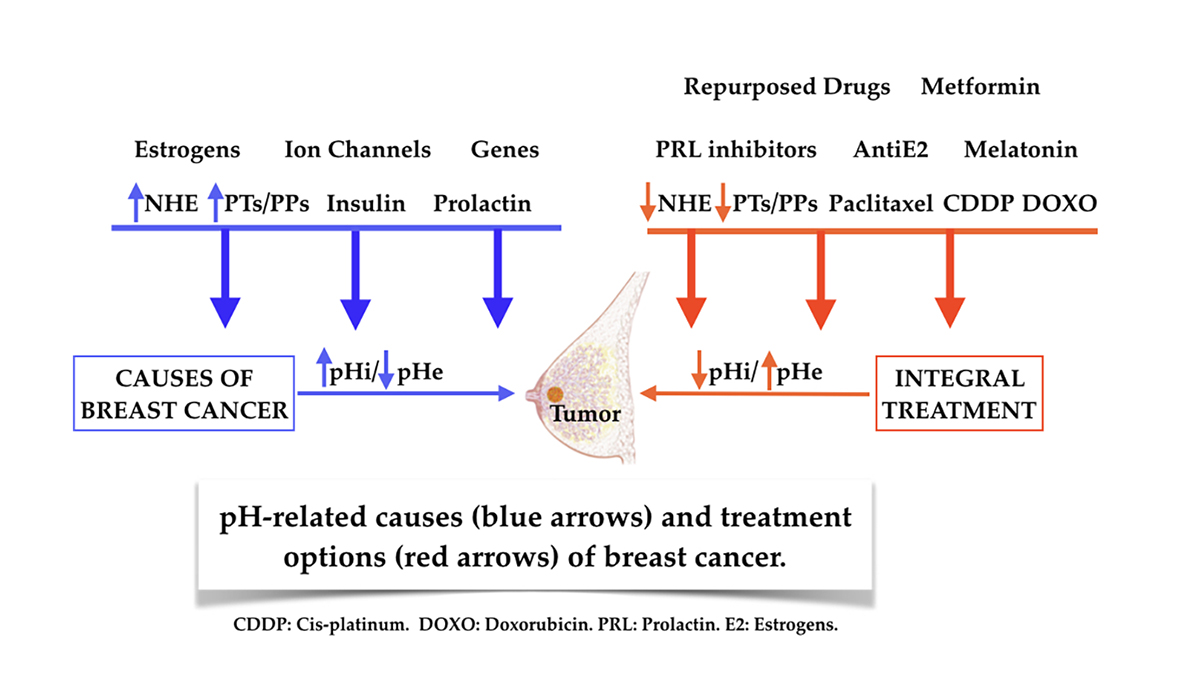{kind=link}
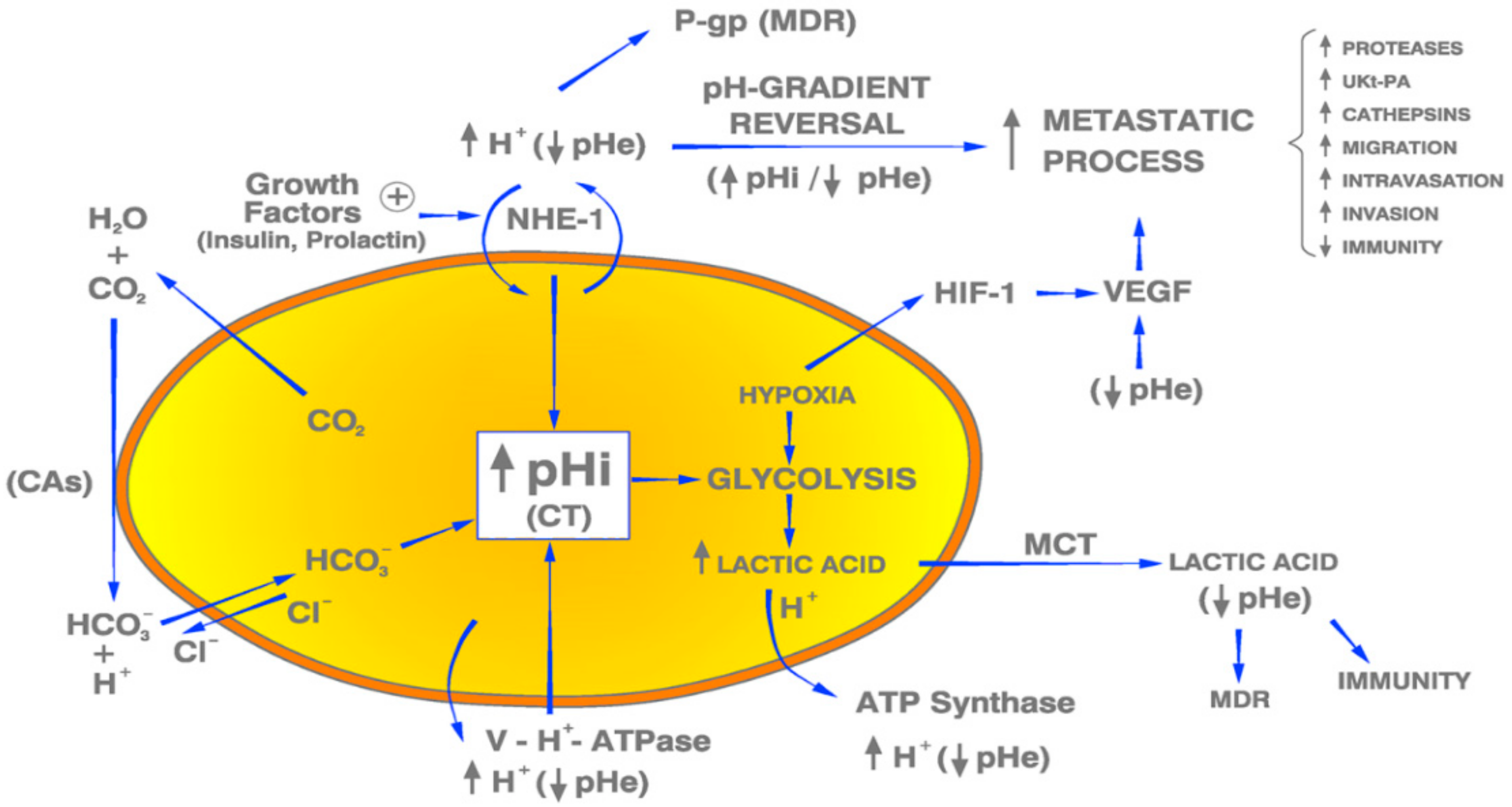{kind=link}
| Mechanisms | Summary | References |
|---|---|---|
| H+ extrusion and/or elevated pHi | H+ extrusion from cells is sufficient to induce transformation, growth and invasion in BC and other tumors. NHE-mediated H+ extrusion by itself has a carcinogenic effect on breast cells and increases MDR. | [8,23,25,37,38,39,40,41,42,43,44,48,54,59,122,123,124,125] |
| Tumor micro-environmental (TME) acidosis, immunity and MDR | Acidity of the TME disrupts the body immune defense mechanisms towards malignant tumors, locally and systemically. This allows a relentless and uncontrolled tumor progression. TME also has an essential role in the progression of inflammatory BC. Thus, TME is a novel therapeutic target in BC. TME acidity also induces MDR. | [20,21,22,47,49,50,51,78,79,84,87,116,120,126,127,128,129,130,131,132,133,134,135,136,137,138] |
| Insulin (INS) and insulin-like growth factor-1 (IGF-1) | INS and INS-resistance have a direct effect in raising pHi and are associated with breast cancer carcinogenicity and progression. Over-expression of insulin/insulin-like growth factor-1 is associated with a decrease in the life span of women with BC. | [3,35,39,139,140,141,142,143,144,145,146,147,148,149] |
| Prolactin (PRL) | PRL stimulates growth, motility and invasiveness of BC cells through NHE1 activation. In this way contributes to the metastatic process of human BC and becomes another therapeutic target. | [150,151,152,153] |
| Estrogens | Estrogens play a crucial role in breast tumorigenesis by promoting cell proliferation and decreasing apoptosis. ER-cells are considered to have a higher expression of NHE activity and are preferably associated with CA-IX over CA-XII. Inhibition of CA-IX improves the prognosis of the disease. | [4,154,155,156,157] |
| Ion channels | NaV1.5-Na+channels associated with NHE-1 are overexpressed in BC, stimulating the formation of invadopodia, facilitating local growth and the metastatic process. | [8,72,73,75,76,77,148] |
| PTs, PPs, and Growth factors | NHE1-overexpression is stimulated by a myriad of factors, which alone or in combination induce a carcinogenic elevation of pHi as the oncogenic response of normal cells of different origins and locations. Carbonic anhydrases (CAs) also have an important role in the pathogenesis of BC, mainly in hypoxic conditions. NHE1 levels are significantly higher in BC tissue than in adjacent normal tissue, and also in resistant BC cells when compared to sensitive cells. | [8,25,41,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71] |
| Drug | Summary | References |
|---|---|---|
| Amiloride (AM) (and/or liposomal amiloride), proton transport inhibitors (PTIs) and intracellular (IC) acidifiers | AM is a non-specific NHE inhibitor first introduced for human use as a K+ sparing diuretic. It works as an antiangiogenic agent and has proved to be most effective as an antimetastatic drug in transplanted breast tumors in animals. A positive clinical experience in an occasional patient has been reported with its chronic use when traditional chemotherapy had failed. Also, the many anti-cancer effects of AM have been fully described. However, its utilization has not entered clinical trials in bedside oncology. (For further details, see the text). | [8,16,25,26,32,53,54,60,63,64,67,68,69,79,82,88,101,110,118,129,158,165,170,171,172,173,174,175,176,177,178,179,180,181,182,183] |
| Proton pump inhibitors (PPIs) and TME alkalization | PPIs are useful in the prevention of BC. Besides, the clinical utilization of V-ATPase inhibitors is a novel therapeutic measure to counteract the abnormal proton dynamics of BC and other tumors. PPIs also benefit from the microenvironmental acidity of tumors. Preclinical and clinical studies also support a direct anti-tumor effect of PPIs in BC and other solid tumors. | [22,49,51,67,78,85,128,130,177,184,185,186,187,188,189,190,191,192,193,194,195,196] |
| Monocarboxylate transport (MCT) inhibitors | Quercetin is a pan-monocarboxylate transporter (MCT) inhibitor and intracellular acidifier. Liposomal quercetin is also available, since gastrointestinal absorption is very limited in the non-liposomal drug form. | [101,102,137,197,198,199] |
| Acetazolamide (AZM) | AZM is a carbonic anhydrase (CA) pan-inhibitor and cell acidifier. CAIX inhibition significantly reduces invasion of BC cells and represents a most promising drug in the treatment of BC, alone or in combination with different NHE inhibitors. | [63,65,165,183,200,201,202,203] |
| Doxorubicin (DOXO) | There is a progressive increase in resistance to DOXO by increasing elevations of pHi, resistance that is suppressed by P-gp inhibitors, while P-gp also increases pHi. MDR is characterized by a reversal of the pH gradient (cancer proton reversal or CPR) across cancer cell membranes. | [8,25,84,88,89,90,101,179,204] |
| Paclitaxel | The inhibition of NHE1, which is fundamental in the chemotherapy of triple-negative BC metastasis, improves the efficacy of Paclitaxel and mediates in Paclitaxel-induced apoptosis of BC cells. | [40,42,44,179,180,205,206,207,208] |
| Cis-platinum (CDDP) | The first effect of CDDP is the induction of intracellular acidification by inhibiting H+ extrusion secondary to downregulation of NHE-1. Contrariwise, the activity of NHE-1 and its effect on elevating pHi increases CDDP resistance to treatment. | [206,209,210,211] |
| Antiestrogens | ER- breast cancer cells have a higher expression of NHE activity and are preferably associated with CA-IX over CA-XII. Inhibition of CA-IX improves the prognosis of the disease. Although the role of Tamoxifen and Letrozole is well established, no further connections among pH dynamics and these antiestrogens have been described. | [4,5,154,155,156,157,200,212,213] |
| Melatonin (MT) | MT has an antiestrogenic effect and only for this reason it should be contemplated in BC therapy. Treatment with MT modulates tumor aggressiveness and increases apoptosis n BC cell lines. MT also suppresses tumor aerobic metabolism (the Warburg effect) and decreases breast cancer angiogenesis and metastasis. | [214,215,216,217,218,219,220,221,222,223,224,225] |
| Cariporide (CP) | CP (HOE-642) is a powerful NHE1 inhibitor that, unfortunately, is not available for clinical use in bedside oncology. It induces apoptotic cells death in different malignant tumors. | [8,25,35,170] |
| Compound 9t (C9t) (Unavailable) | C9t is 500-fold more potent against NHE1 than cariporide and has a 1400-fold greater selectivity for NHE1 over NHE2. Besides, C9t is orally bioavailable, has low side-effects in mice and it presents a significantly improved safety profile over other NHE1inhibitors. | [8,35,178] |
| Phx-3 | Phx-3 is a potent, selective and non-toxic NHE1 inhibitor that triggers apoptosis in a variety of cancer cell lines and is highly effective in some animal tumor models. | [8,32,182] |
| Repurposed drugs | Phloretin, Lonidamine, Niclosamide, Docosaexaenoic acid, Salinomycin and Simvastatin have been reported to be useful in the treatment of BC because of their pH-related effects. Resveratrol also has a role as an aromatase inhibitor. (For further details, see the text). | [8,103,226,227,228,229,230,231,232,233,234] |
| Metformin (MET) | MET has been introduced as an anticancer agent in BC. It induces intracellular hyperacidification in tumor xenograft models. MET has been reported to inhibit insulin and insulin/IGF-1, HIF-1α, Warburg metabolism, gene expression, angiogenesis, cancer migration, invasion and metastasis, apart from reducing the side effects of doxorubicin. MET has also been reported to act synergistically with chemotherapy and decrease its dosages, thus, its side-effects. It has also been used to target resistant cells in BC and has been considered a radio-sensitizer. | [235,236,237,238,239,240,241,242,243,244,245,246,247,248,249,250,251,252,253,254,255] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harguindey, S.; Alfarouk, K.; Polo Orozco, J.; Hardonnière, K.; Stanciu, D.; Fais, S.; Devesa, J. A New and Integral Approach to the Etiopathogenesis and Treatment of Breast Cancer Based upon Its Hydrogen Ion Dynamics. Int. J. Mol. Sci. 2020, 21, 1110. https://doi.org/10.3390/ijms21031110
Harguindey S, Alfarouk K, Polo Orozco J, Hardonnière K, Stanciu D, Fais S, Devesa J. A New and Integral Approach to the Etiopathogenesis and Treatment of Breast Cancer Based upon Its Hydrogen Ion Dynamics. International Journal of Molecular Sciences. 2020; 21(3):1110. https://doi.org/10.3390/ijms21031110
Chicago/Turabian StyleHarguindey, Salvador, Khalid Alfarouk, Julián Polo Orozco, Kévin Hardonnière, Daniel Stanciu, Stefano Fais, and Jesús Devesa. 2020. "A New and Integral Approach to the Etiopathogenesis and Treatment of Breast Cancer Based upon Its Hydrogen Ion Dynamics" International Journal of Molecular Sciences 21, no. 3: 1110. https://doi.org/10.3390/ijms21031110
APA StyleHarguindey, S., Alfarouk, K., Polo Orozco, J., Hardonnière, K., Stanciu, D., Fais, S., & Devesa, J. (2020). A New and Integral Approach to the Etiopathogenesis and Treatment of Breast Cancer Based upon Its Hydrogen Ion Dynamics. International Journal of Molecular Sciences, 21(3), 1110. https://doi.org/10.3390/ijms21031110