Expression, Purification, and Characterization of Anti-Zika virus Envelope Protein: Polyclonal and Chicken-Derived Single Chain Variable Fragment Antibodies

 , ,
, ,
Abstract
1. Introduction
2. Results
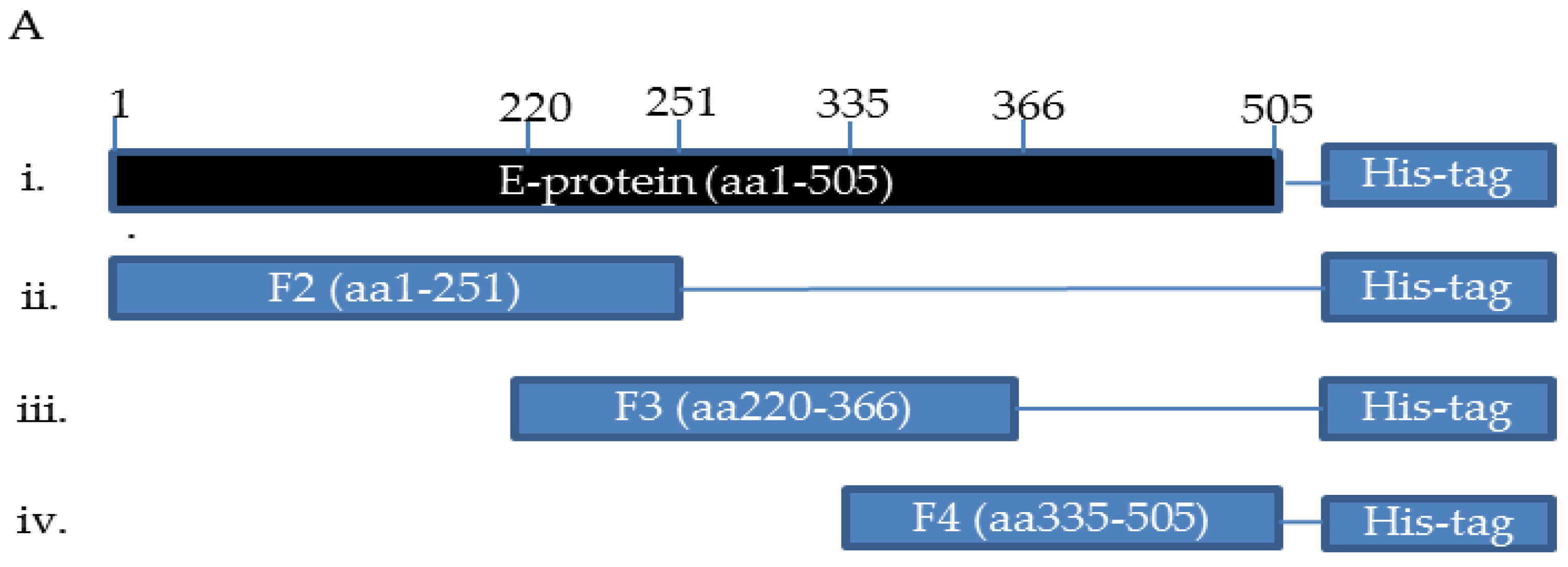
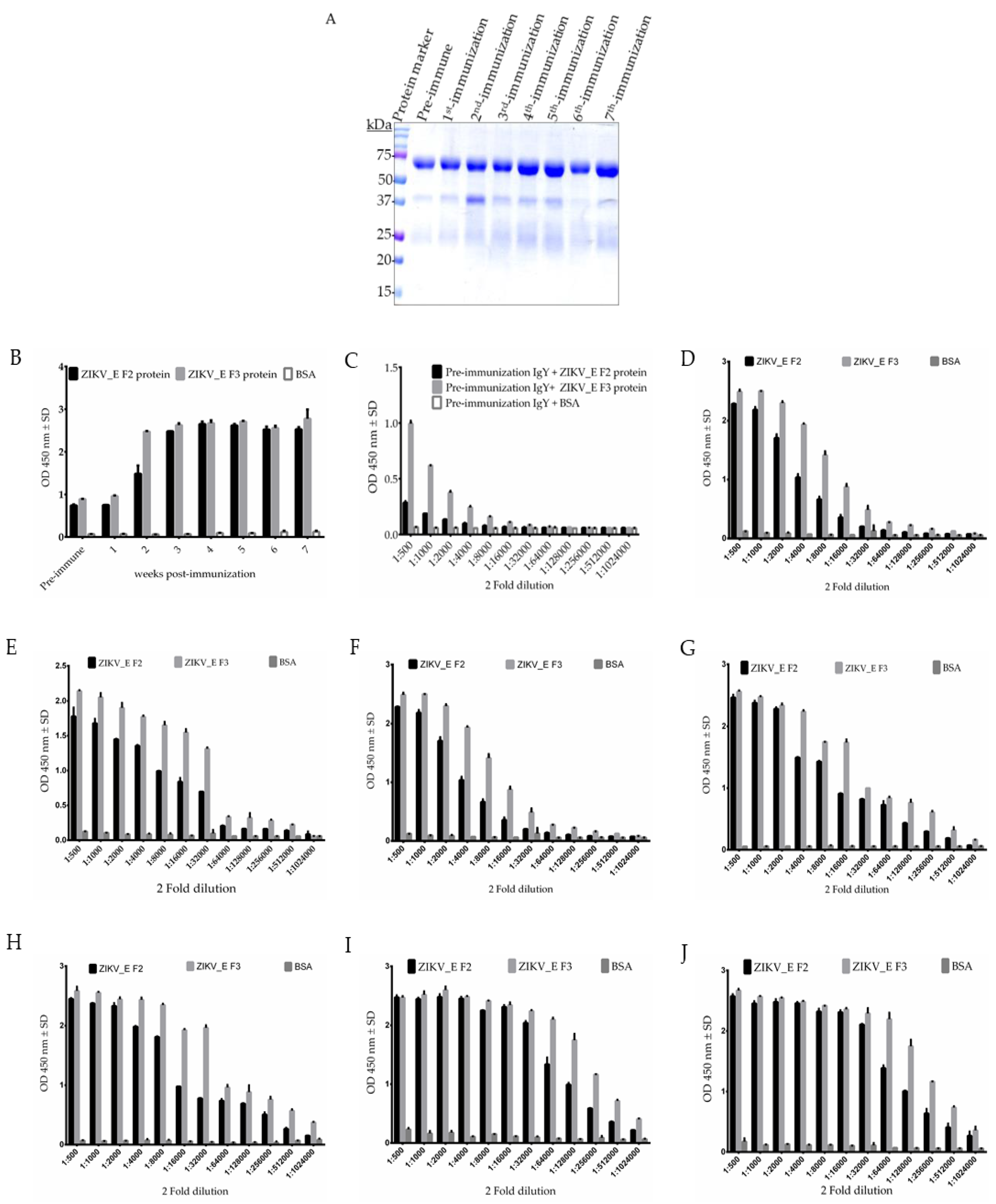
2.1. Expression, Purification, Immunization, and Characterization of ZIKV_E Recombinant Proteins
2.2. Purification and Characterization of Polyclonal Anti-ZIKV_E IgY Antibodies
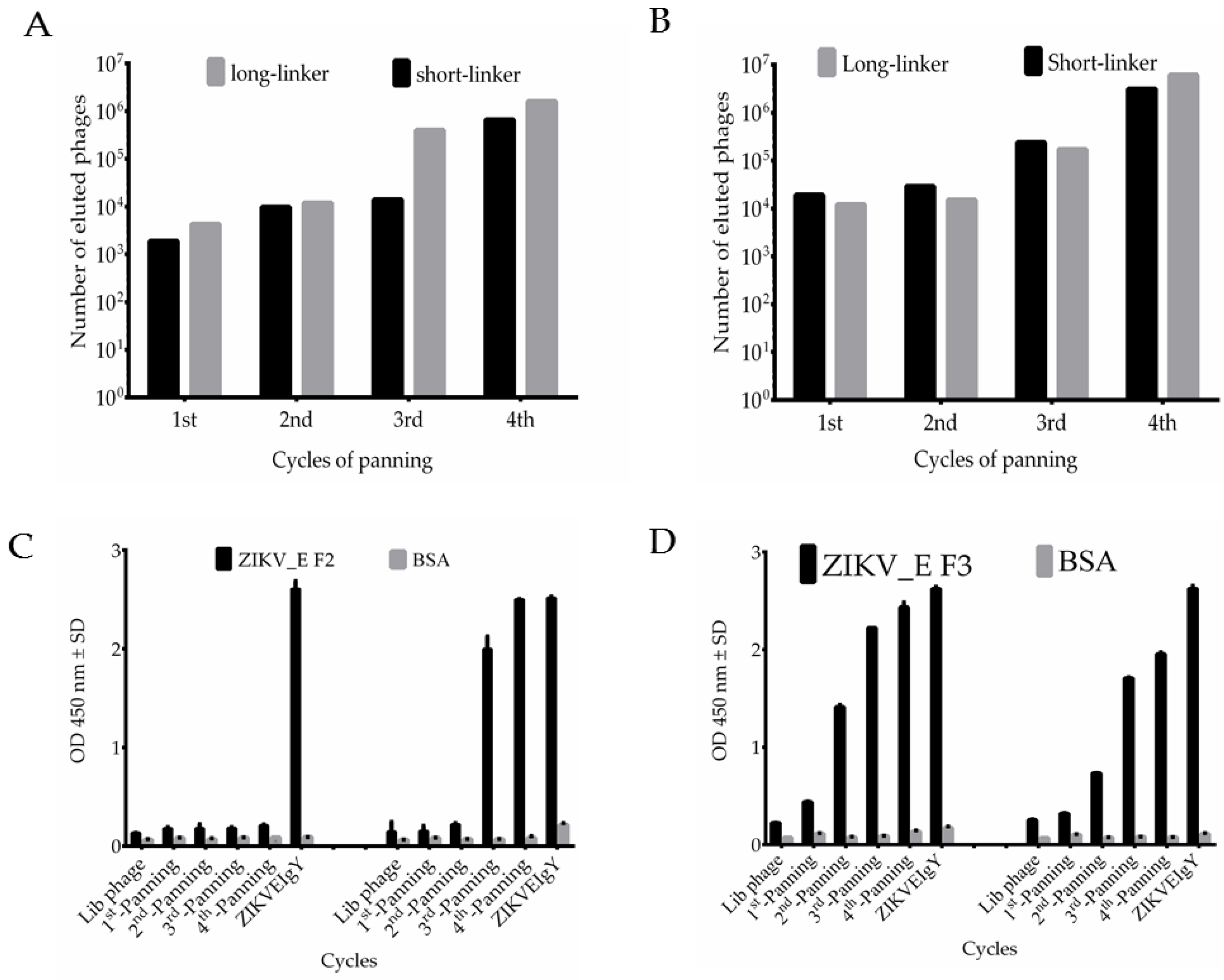
2.3. Bio-Panning Process and Characterization of Phage-Displayed Chicken Anti-ZIKV_E scFv Antibody Libraries
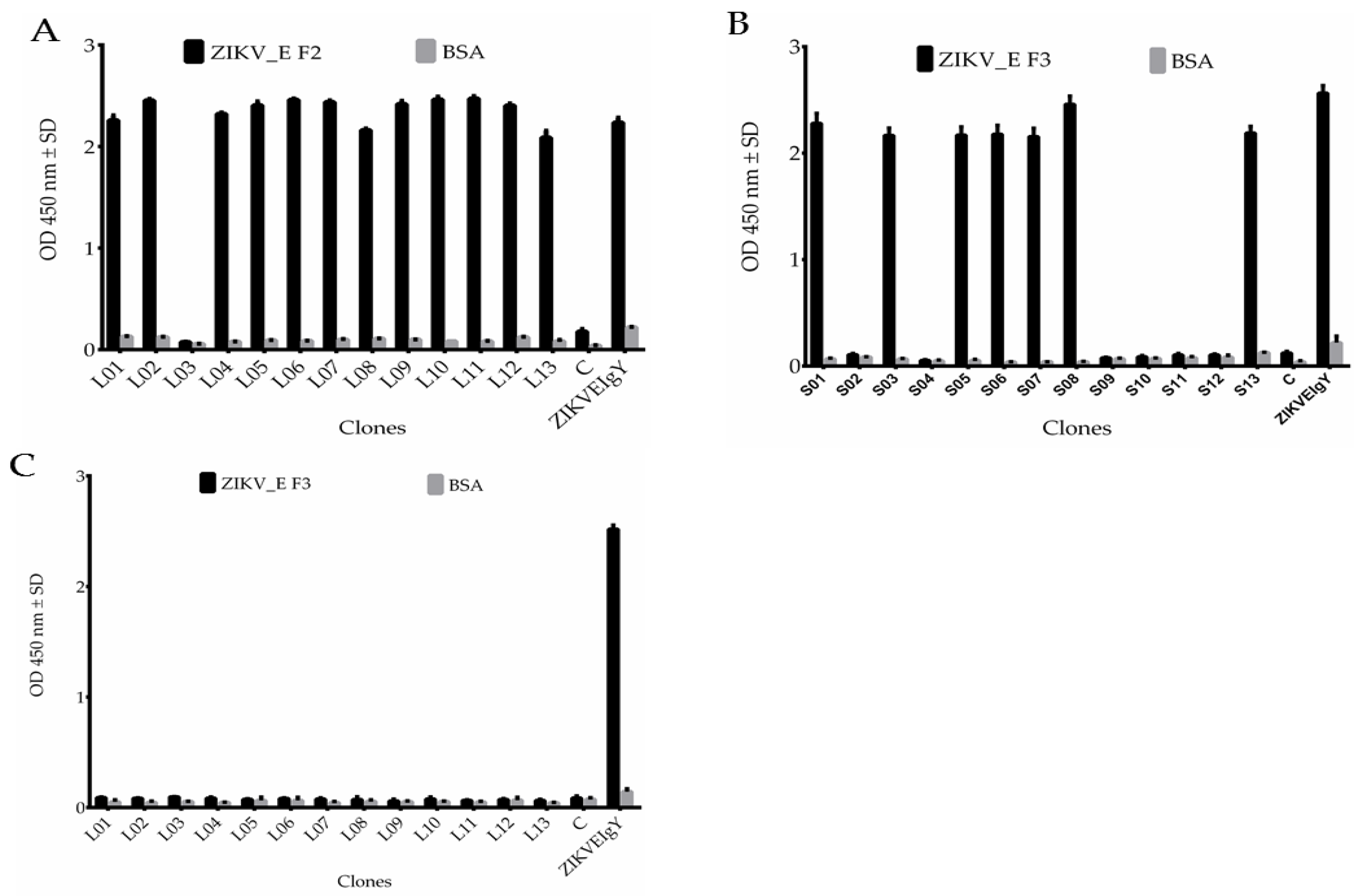
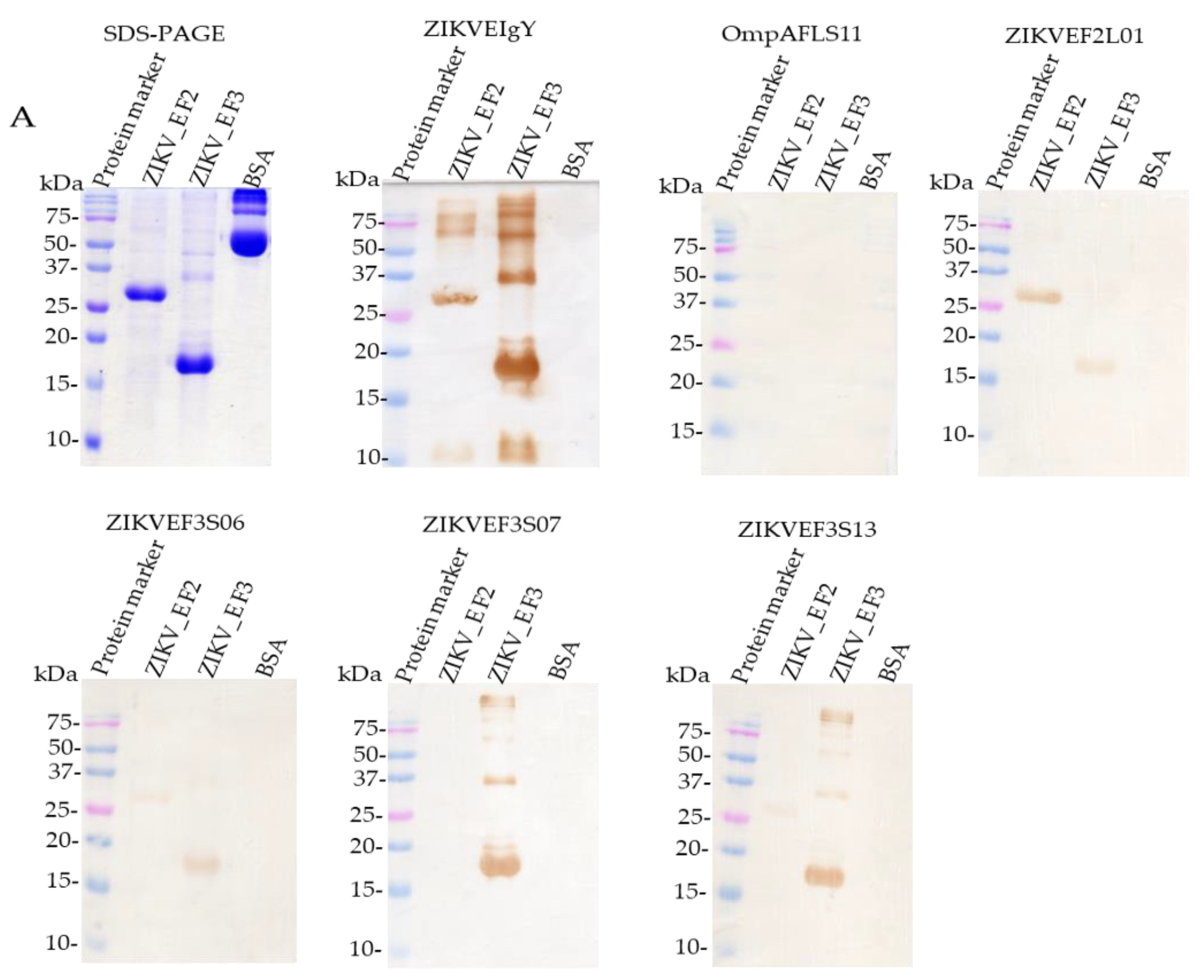
2.4. Sequence Analysis, Expression, Purification, and Characterization of Anti-ZIKV_E scFv Antibodies
2.5. Characterization of Binding Specificity of Anti-ZIKV_E scFv and Polyclonal Anti-ZIKV_E IgY Antibodies
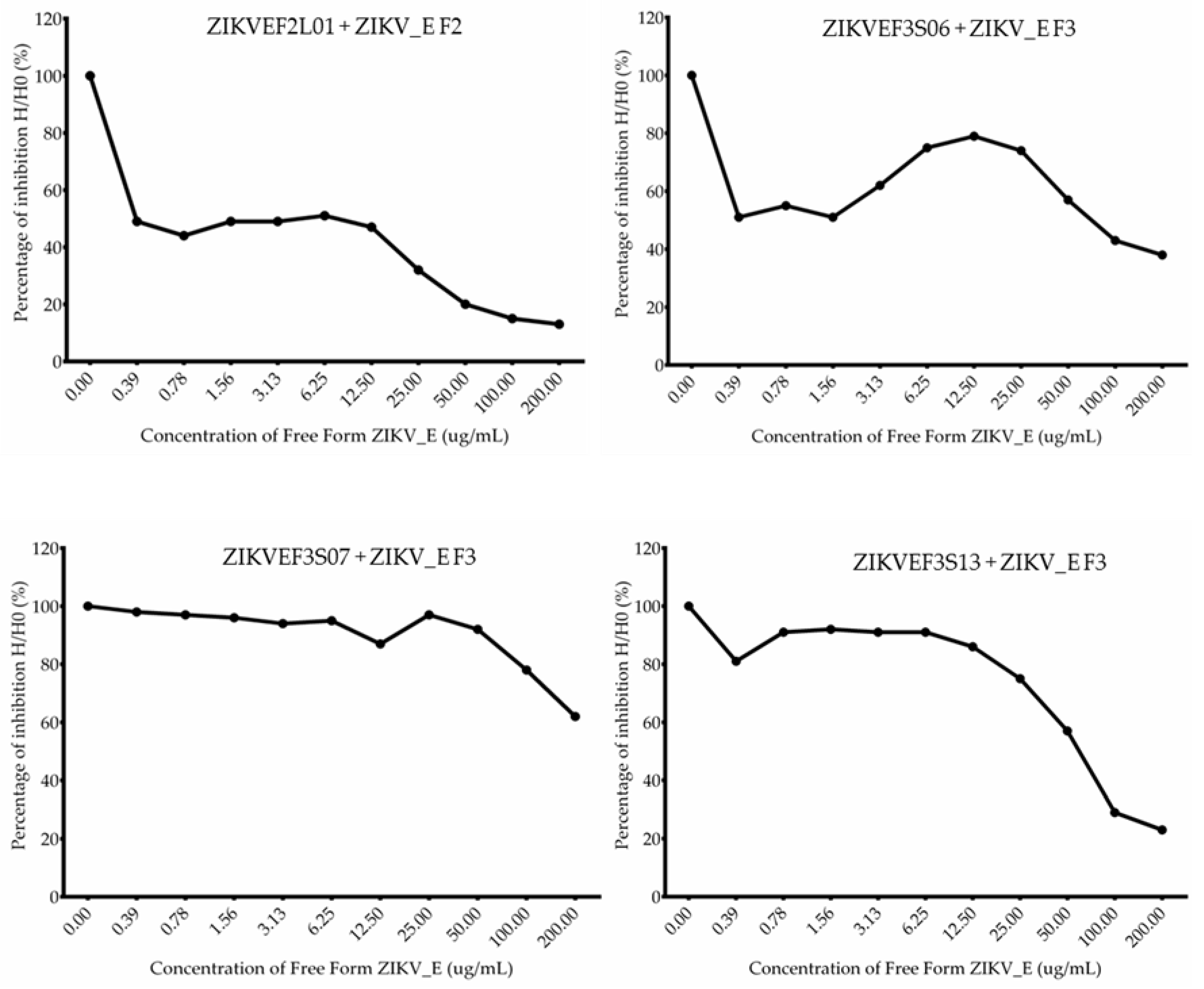
2.6. Competitive ELISA Assay of Anti-ZIKV_E scFv Antibodies
3. Discussion
4. Materials and Methods
4.1. Plasmid Vectors, Bacterial Strains, Media, Cell Lines, and Antibodies
4.2. Preparation of ZIKV_E Gene Fragments
4.3. Expression and Purification of ZIKV_E Recombinant Proteins
4.4. Immunization of White Leghorn Chicken
4.5. Extraction of Anti-ZIKV_E IgY from Egg Yolk
4.6. ELISA for Chicken Immune Response Analysis
4.7. Chicken Antibody Library Construction
4.8. Bio-Panning of Anti-ZIKV_E scFv from Chicken Antibody Library
4.9. Anti-ZIKV_E scFv Antibodies Selection, Expression, and Purification after the Fourth Round of Bio-Panning
4.10. SDS-PAGE and Western Blot Analysis
4.11. Enzyme-Linked Immunosorbent Assay (ELISA)
4.12. Expression of ZIKV_E Protein in Vero Cells
4.13. Competitive ELISA
4.14. Sequence Analysis of Anti-ZIKV_E scFv Antibodies Genes
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ZIKV_E | Zika virus envelope |
| TMB | 3,3′,5,5′-tetramethylbenzidine |
| IgY | Immunoglobulin yolk |
| DAB | 3,3′-diaminobenzedine |
| BD | Becton Dickinson |
| TBS | Tris-buffered saline |
| RPM | Revolution per minute |
| RT | Room temperature |
| MIN h | Minute hour |
| ScFv | Single chain variable fragment |
References
- Musso, D.; Gubler, D.J. Zika Virus. Clin. Microbiol. Rev. 2016, 29, 487–524. [Google Scholar] [CrossRef]
- Weaver, S.C.; Costa, F.; Garcia-Blanco, M.A.; Ko, A.I.; Ribeiro, G.S.; Saade, G.; Shi, P.Y.; Vasilakis, N. Zika Virus: History, Emergence, Biology, and Prospects for Control. Antiviral. Res. 2016, 130, 69–80. [Google Scholar] [CrossRef]
- Paul, R.E. The When and the Where of Zika Epidemic Potential in Europe-An Evidence Base for Public Health Preparedness. EBioMedicine 2016, 9, 17–18. [Google Scholar] [CrossRef][Green Version]
- Knipe, D.; Howley, P. Fields Virology (Knipe, Fields Virology)-2 Volume Set; LWW: Philadelphia, PA, USA, 2013. [Google Scholar]
- Al-Qahtani, A.A.; Nazir, N.; Al-Anazi, M.R.; Rubino, S.; Al-Ahdal, M.N. Zika Virus: A New Pandemic Threat. J. Infect. Dev. Ctries. 2016, 10, 201–207. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Carteaux, G.; Maquart, M.; Bedet, A.; Contou, D.; Brugieres, P.; Fourati, S.; Cleret de Langavant, L.; de Broucker, T.; Brun-Buisson, C.; Leparc-Goffart, I.; et al. Zika Virus Associated with Meningoencephalitis. N. Engl. J. Med. 2016, 374, 1595–1596. [Google Scholar] [CrossRef]
- Cao-Lormeau, V.M.; Blake, A.; Mons, S.; Lastere, S.; Roche, C.; Vanhomwegen, J.; Dub, T.; Baudouin, L.; Teissier, A.; Larre, P.; et al. Guillain-Barre Syndrome Outbreak Associated with Zika Virus Infection in French Polynesia: A Case-Control Study. Lancet 2016, 387, 1531–1539. [Google Scholar] [CrossRef]
- Oehler, E.; Watrin, L.; Larre, P.; Leparc-Goffart, I.; Lastere, S.; Valour, F.; Baudouin, L.; Mallet, H.; Musso, D.; Ghawche, F. Zika Virus Infection Complicated by Guillain-Barre Syndrome-Case Report, French Polynesia, December 2013. Euro Surveill. 2014, 19. [Google Scholar] [CrossRef] [PubMed]
- Mlakar, J.; Korva, M.; Tul, N.; Popovic, M.; Poljsak-Prijatelj, M.; Mraz, J.; Kolenc, M.; Resman Rus, K.; Vesnaver Vipotnik, T.; Fabjan Vodusek, V.; et al. Zika Virus Associated with Microcephaly. N. Engl. J. Med. 2016, 374, 951–958. [Google Scholar] [CrossRef]
- Dick, G.W.; Kitchen, S.F.; Haddow, A.J. Zika Virus. I. Isolations and Serological Specificity. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef]
- Xie, X.; Shan, C.; Shi, P.-Y. Restriction of Zika Virus by Host Innate Immunity. Cell Host Microbe 2016, 19, 566–567. [Google Scholar] [CrossRef]
- Lazear, H.M.; Diamond, M.S. Zika Virus: New Clinical Syndromes and Its Emergence in the Western Hemisphere. J. Virol. 2016, 90, 4864–4875. [Google Scholar] [CrossRef]
- Pierson, T.C.; Diamond, M. Flaviviruses. In Fields Virology: Sixth Edition, 6th ed.; Knipe, D.M., Howley, H.P., Eds.; Wolters Kluwer/Lippencott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 747–794. [Google Scholar]
- Gatherer, D.; Kohl, A. Zika Virus: A Previously Slow Pandemic Spreads Rapidly through the Americas. J. Gen. Virol. 2016, 97, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Sager, G.; Gabaglio, S.; Sztul, E.; Belov, G.A. Role of Host Cell Secretory Machinery in Zika Virus Life Cycle. Viruses 2018, 10, 559. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Thurmond, S.; Islas, L.; Hui, K.; Hai, R. Zika Virus Genome Biology and Molecular Pathogenesis. Emerg. Microbes. Infect. 2017, 6, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.H.; Yip, B.S.; Huang, L.M.; Wu, S.C. Zika Virus Structural Biology and Progress in Vaccine Development. Biotechnol. Adv. 2018, 36, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Modis, Y.; Ogata, S.; Clements, D.; Harrison, S.C. Structure of the Dengue Virus Envelope Protein after Membrane Fusion. Nature 2004, 427, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Azeredo, E.; Dos Santos, F.; Barbosa, L.; Souza, T.; Badolato-Correa, J.; Sanchez-Arcila, J. Clinical and Laboratory Profile of Zika and Dengue Infected Patients: Lessons Learned From the Co-circulation of Dengue, Zika and Chikungunya in Brazil. PLoS Curr. 2018, 10. [Google Scholar] [CrossRef]
- Jara-Acevedo, R.; Díez, P.; González-González, M.; Dégano, R.M.; Ibarrola, N.; Gongora, R.; Orfao, A.; Fuentes, M. Methods for Selecting Phage Display Antibody Libraries. Curr. Pharm. Des. 2016, 22, 6490–6499. [Google Scholar] [CrossRef]
- Du, X.-J.; Wu, Y.-N.; Zhang, W.-W.; Dong, F.; Wang, S. Construction and Quality Examination of Murine Naive T7 Phage Display Antibody Library. Food Agric. Immunol. 2010, 21, 81–90. [Google Scholar] [CrossRef]
- Huston, J.S.; Levinson, D.; Mudgett-Hunter, M.; Tai, M.-S.; Novotný, J.; Margolies, M.N.; Ridge, R.J.; Bruccoleri, R.E.; Haber, E.; Crea, R. Protein Engineering of Antibody Binding Sites: Recovery of Specific Activity in an Anti-Digoxin Single-Chain Fv Analogue Produced in Escherichia Coli. Proc. Natl. Acad. Sci. USA 1988, 85, 5879–5883. [Google Scholar] [CrossRef]
- Siegel, D.L. Recombinant Monoclonal Antibody Technology. Transfus. Clin. Boil. J. De La Soc. Fr. De Transfus. Sang. 2002, 9, 15–22. [Google Scholar] [CrossRef]
- Jara-Acevedo, R.; Diez, P.; Gonzalez-Gonzalez, M.; Degano, R.M.; Ibarrola, N.; Gongora, R.; Orfao, A.; Fuentes, M. Screening Phage-Display Antibody Libraries Using Protein Arrays. Methods Mol. Boil. (Clifton, N.J.) 2018, 1701, 365–380. [Google Scholar] [CrossRef]
- Rahman, S.; Van Nguyen, S.; Icatlo, F.C., Jr.; Umeda, K.; Kodama, Y. Oral Passive IgY-Based Immunotherapeutics: A Novel Solution for Prevention and Treatment of Alimentary Tract Diseases. Hum. Vaccine. Immunother. 2013, 9, 1039–1048. [Google Scholar] [CrossRef] [PubMed]
- Michael, A.; Meenatchisundaram, S.; Parameswari, G.; Subbraj, T.; Selvakumaran, R.; Ramalingam, S. Chicken Egg Yolk Antibodies (IgY) as an Alternative to Mammalian Antibodies. Indian J. Sci. Technol. 2010, 3, 468–474. Available online: http://www.indjst.org (accessed on 25 December 2019).
- Duffy, M.R.; Chen, T.H.; Hancock, W.T.; Powers, A.M.; Kool, J.L.; Lanciotti, R.S.; Pretrick, M.; Marfel, M.; Holzbauer, S.; Dubray, C.; et al. Zika Virus Outbreak on Yap Island, Federated States of Micronesia. N. Engl. J. Med. 2009, 360, 2536–2543. [Google Scholar] [CrossRef] [PubMed]
- Bird, C.R.; Thorpe, R. Purification of Immunoglobulin Y (IgY) from Chicken Eggs. In The Protein Protocols Handbook; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1779–1781. [Google Scholar] [CrossRef]
- Lee, C.H.; Leu, S.J.; Lee, Y.C.; Liu, C.I.; Lin, L.T.; Mwale, P.F.; Chiang, J.R.; Tsai, B.Y.; Chen, C.C.; Hung, C.S.; et al. Characterization of Chicken-Derived Single Chain Antibody Fragments against Venom of Naja Naja Atra. Toxins 2018, 10, 383. [Google Scholar] [CrossRef] [PubMed]
- Daugherty, P.S.; Chen, G.; Iverson, B.L.; Georgiou, G. Quantitative Analysis of the Effect of the Mutation Frequency on the Affinity Maturation of Single Chain Fv Antibodies. Proc. Natl. Acad. Sci. USA 2000, 97, 2029–2034. [Google Scholar] [CrossRef]
- Dai, L.; Song, J.; Lu, X.; Deng, Y.Q.; Musyoki, A.M.; Cheng, H.; Zhang, Y.; Yuan, Y.; Song, H.; Haywood, J.; et al. Structures of the Zika Virus Envelope Protein and Its Complex with a Flavivirus Broadly Protective Antibody. Cell Host Microbe 2016, 19, 696–704. [Google Scholar] [CrossRef]
- Deng, Y.Q.; Dai, J.X.; Ji, G.H.; Jiang, T.; Wang, H.J.; Yang, H.O.; Tan, W.L.; Liu, R.; Yu, M.; Ge, B.X.; et al. A Broadly Flavivirus Cross-Neutralizing Monoclonal Antibody that Recognizes a Novel Epitope within the Fusion Loop of E Protein. PLoS ONE 2011, 6, e16059. [Google Scholar] [CrossRef]
- Munhoz, L.S.; Vargas, G.D.Á.; Fischer, G.; Lima, M.D.; Esteves, P.A.; Hübner, S.d.O. Avian IgY Antibodies: Characteristics and Applications in Immunodiagnostic. Ciência Rural 2014, 44, 153–160. [Google Scholar] [CrossRef]
- Akita, E.; Nakai, S. Immunoglobulins from Egg Yolk: Isolation and Purification. J. Food Sci. 1992, 57, 629–634. [Google Scholar] [CrossRef]
- Kuno, G.; Chang, G.J. Full-Length Sequencing and Genomic Characterization of Bagaza, Kedougou, and Zika Viruses. Arch Virol. 2007, 152, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Han, J.F.; Qiu, Y.; Yu, J.Y.; Wang, H.J.; Deng, Y.Q.; Li, X.F.; Zhao, H.; Sun, H.X.; Qin, C.F. Immunization with Truncated Envelope Protein of Zika Virus Induces Protective Immune Response in Mice. Sci. Rep. 2017, 7, 10047. [Google Scholar] [CrossRef]
- Yang, M.; Dent, M.; Lai, H.; Sun, H.; Chen, Q. Immunization of Zika Virus Envelope Protein Domain III Induces Specific and Neutralizing Immune Responses Against Zika Virus. Vaccine 2017, 35, 4287–4294. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Yang, R.; Liu, Z.; Li, M.; Liu, H.; Jin, X. Recombinant Zika Virus Envelope Protein Elicited Protective Immunity Against Zika Virus in Immunocompetent Mice. PLoS ONE 2018, 13, e0194860. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, M.C.; Schaefer, R.; Gava, D.; Souza, C.K.; da Silva Vaz, I., Jr.; Bastos, A.P.; Venancio, E.J. Production and Application of Anti-Nucleoprotein IgY Antibodies for Influenza A Virus Detection in Swine. J. Immunol. Methods 2018, 461, 100–105. [Google Scholar] [CrossRef]
- Silva Ados, S.; Vasconcelos, G.A.; Kappel, L.A.; Pinto, M.A.; Paula, V.S. An Immunoenzymatic Assay for the Diagnosis of Hepatitis A Utilising Immunoglobulin Y. Mem. Inst. Oswaldo Cruz. 2012, 107, 960–963. [Google Scholar] [CrossRef][Green Version]
- Cabezas, S.; Rojas, G.; Pavon, A.; Alvarez, M.; Pupo, M.; Guillen, G.; Guzman, M.G. Selection of Phage-Displayed Human Antibody Fragments on Dengue Virus Particles Captured by a Monoclonal Antibody: Application to the Four Serotypes. J. Virol. Methods 2008, 147, 235–243. [Google Scholar] [CrossRef]
- Nilvebrant, J.; Sidhu, S.S. Construction of Synthetic Antibody Phage-Display Libraries. Methods Mol. Biol. (Clifton, N.J.) 2018, 1701, 45–60. [Google Scholar] [CrossRef]
- Li, T.W.; Cheng, S.F.; Tseng, Y.T.; Yang, Y.C.; Liu, W.C.; Wang, S.C.; Chou, M.J.; Lin, Y.J.; Wang, Y.; Hsiao, P.W.; et al. Development of Single-Chain Variable Fragments (scFv) Against Influenza Virus Targeting Hemagglutinin Subunit 2 (HA2). Arch Virol. 2016, 161, 19–31. [Google Scholar] [CrossRef]
- Barbas, C.F., 3rd; Kang, A.S.; Lerner, R.A.; Benkovic, S.J. Assembly of Combinatorial Antibody Libraries on Phage Surfaces: The Gene III Site. Proc. Natl. Acad. Sci. USA 1991, 88, 7978–7982. [Google Scholar] [CrossRef]
- Calvet, G.; Aguiar, R.S.; Melo, A.S.; Sampaio, S.A.; De Filippis, I.; Fabri, A.; Araujo, E.S.; de Sequeira, P.C.; de Mendonça, M.C.; de Oliveira, L. Detection and Sequencing of Zika Virus from Amniotic Fluid of Fetuses with Microcephaly in Brazil: A Case Study. Lancet Infect. Dis. 2016, 16, 653–660. [Google Scholar] [CrossRef]
- Terpe, K. Overview of Bacterial Expression Systems for Heterologous Protein Production: From Molecular and Biochemical Fundamentals to Commercial Systems. Appl. Microbial. Biotechnol. 2006, 72, 211. [Google Scholar] [CrossRef] [PubMed]
- Akita, E.M.; Nakai, S. Production and Purification of Fab’ Fragments from Chicken Egg Yolk Immunoglobulin Y (IgY). J. Immunol. Methods 1993, 162, 155–164. [Google Scholar] [CrossRef]
- Engvall, E.; Jonsson, K.; Perlmann, P. Enzyme-Linked Immunosorbent Assay. II. Quantitative Assay of Protein Antigen, Immunoglobulin G, by Means of Enzyme-Labelled Antigen and Antibody-Coated Tubes. Biochim. Biophys. Acta (BBA)-Protein Struct. 1971, 251, 427–434. [Google Scholar] [CrossRef]
- Palmieri, M.; Vagnini, M.; Pitzurra, L.; Brunetti, B.; Cartechini, L. Identification of Animal Glue and Hen-Egg Yolk in Paintings by Use of Enzyme-Linked Immunosorbent Assay (ELISA). Anal. Bioanal. Chem. 2013, 405, 6365–6371. [Google Scholar] [CrossRef]
- Van Coillie, E.; De Block, J.; Reybroeck, W. Development of an Indirect Competitive ELISA for Flumequine Residues in Raw Milk Using Chicken Egg Yolk Antibodies. J. Agric. Food Chem. 2004, 52, 4975–4978. [Google Scholar] [CrossRef] [PubMed]
- Andris-Widhopf, J.; Rader, C.; Steinberger, P.; Fuller, R.; Barbas Iii, C.F. Methods for the Generation of Chicken Monoclonal Antibody Fragments by Phage Display. J. Immunol. Methods 2000, 242, 159–181. [Google Scholar] [CrossRef]
- Scott, J. Phage-Display Vectors (2.1–2.19) in Phage Display: A Laboratory Manual. Edited by cf Barbas iii, dr Burton, jk Scott, and gj silverman; Cold Spring Harbor: New York, NY, USA, 2001. [Google Scholar]
- Lee, C.-H.; Lee, Y.-C.; Leu, S.-J.; Lin, L.-T.; Chiang, J.-R.; Hsu, W.-J.; Yang, Y.-Y. Production and Characterization of Neutralizing Antibodies Against Bungarus Multicinctus Snake Venom. Appl. Environ. Microbiol. 2016, 82, 6973–6982. [Google Scholar] [CrossRef]
- Friguet, B.; Chaffotte, A.F.; Djavadi-Ohaniance, L.; Goldberg, M.E. Measurements of the True Affinity Constant in Solution of Antigen-Antibody Complexes by Enzyme-Linked Immunosorbent Assay. J. Immunol. Methods 1985, 77, 305–319. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Oligonucleotide (5′–3′) |
|---|---|
| ZIKVEFLF | TAAAATGGATCCATGATCAGGTGCATAGGAGTCAGC |
| ZIKVEFLR | AATTTTCTCGAGAGCAGAGACGGCTGTGGATAAG |
| ZIKVEF3F | AATTAAGGATCCGACATTCCATTACCTTGGCACGCT |
| ZIKVEF3R | TTAATTCTCGAGGATTACGGGGTTAGCGGTTATCAAC |
| ZIKVEF2R | TAAATTCTCGAGGGCATGTGCGTCCTTGAACTCTAC |
| ZIKVEF4F | TTATAAGGATCCACAGATGGACCTTGCAAGGTTCCA |
| Region | FR1 | FR2 | FR3 | FR4 | Total FRs | CDR1 | CDR2 | CDR3 | Total CDRs |
|---|---|---|---|---|---|---|---|---|---|
| VL | 5–10% | 6–19% | 9–16% | 0–9% | 5–19% | 50–88% | 29–71% | 36–64% | 29–88% |
| VH | 10–15% | 0–7% | 9–19% | 0% | 0–19% | 20–100% | 29–47% | 63% | 20–100% |
| scFv Clones | Inhibition of 50% Binding of Total Protein (µg/mL) | KD Values (M) |
|---|---|---|
| ZIKVEF2L01 | 0.5515 | 2.04 × 10−8 |
| ZIKVEF3S06 | 77.425 | 4.55 × 10−6 |
| ZIKVEF3S07 | 254.14 | 1.49 × 10−5 |
| ZIKVEF3S13 | 110.885 | 6.52 × 10−6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mwale, P.F.; Lee, C.-H.; Lin, L.-T.; Leu, S.-J.; Huang, Y.-J.; Chiang, L.-C.; Mao, Y.-C.; Yang, Y.-Y. Expression, Purification, and Characterization of Anti-Zika virus Envelope Protein: Polyclonal and Chicken-Derived Single Chain Variable Fragment Antibodies. Int. J. Mol. Sci. 2020, 21, 492. https://doi.org/10.3390/ijms21020492
Mwale PF, Lee C-H, Lin L-T, Leu S-J, Huang Y-J, Chiang L-C, Mao Y-C, Yang Y-Y. Expression, Purification, and Characterization of Anti-Zika virus Envelope Protein: Polyclonal and Chicken-Derived Single Chain Variable Fragment Antibodies. International Journal of Molecular Sciences. 2020; 21(2):492. https://doi.org/10.3390/ijms21020492
Chicago/Turabian StyleMwale, Pharaoh Fellow, Chi-Hsin Lee, Liang-Tzung Lin, Sy-Jye Leu, Yun-Ju Huang, Liao-Chun Chiang, Yan-Chiao Mao, and Yi-Yuan Yang. 2020. "Expression, Purification, and Characterization of Anti-Zika virus Envelope Protein: Polyclonal and Chicken-Derived Single Chain Variable Fragment Antibodies" International Journal of Molecular Sciences 21, no. 2: 492. https://doi.org/10.3390/ijms21020492
APA StyleMwale, P. F., Lee, C.-H., Lin, L.-T., Leu, S.-J., Huang, Y.-J., Chiang, L.-C., Mao, Y.-C., & Yang, Y.-Y. (2020). Expression, Purification, and Characterization of Anti-Zika virus Envelope Protein: Polyclonal and Chicken-Derived Single Chain Variable Fragment Antibodies. International Journal of Molecular Sciences, 21(2), 492. https://doi.org/10.3390/ijms21020492