Crocetin Extracted from Saffron Shows Antitumor Effects in Models of Human Glioblastoma


 ,
,  ,
, 
 ,
,  and
and
Abstract
1. Introduction
2. Results
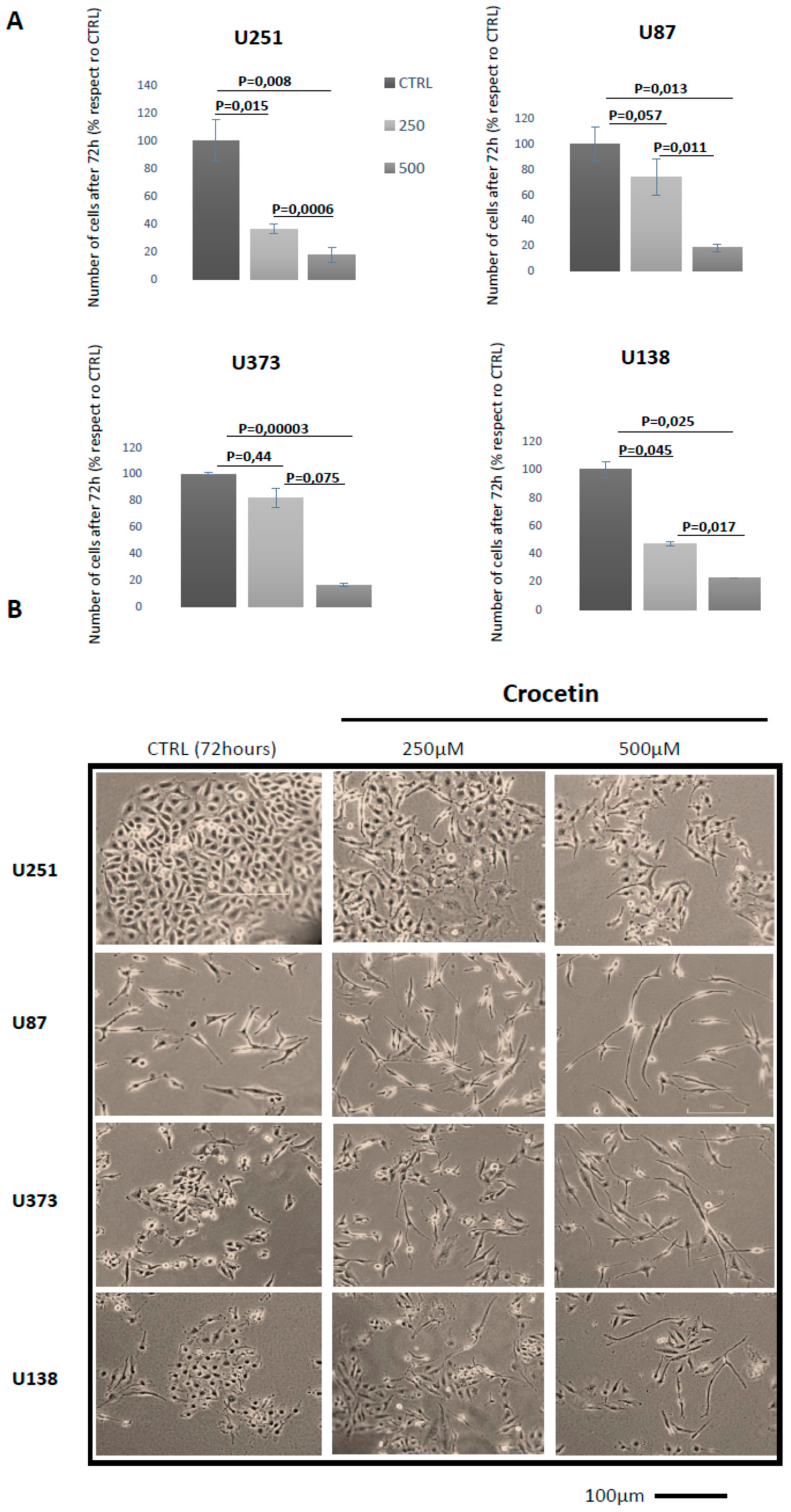
2.1. Crocetin (CCT) Reduces Proliferation and Induces Morphology Changes in Glioma Cells
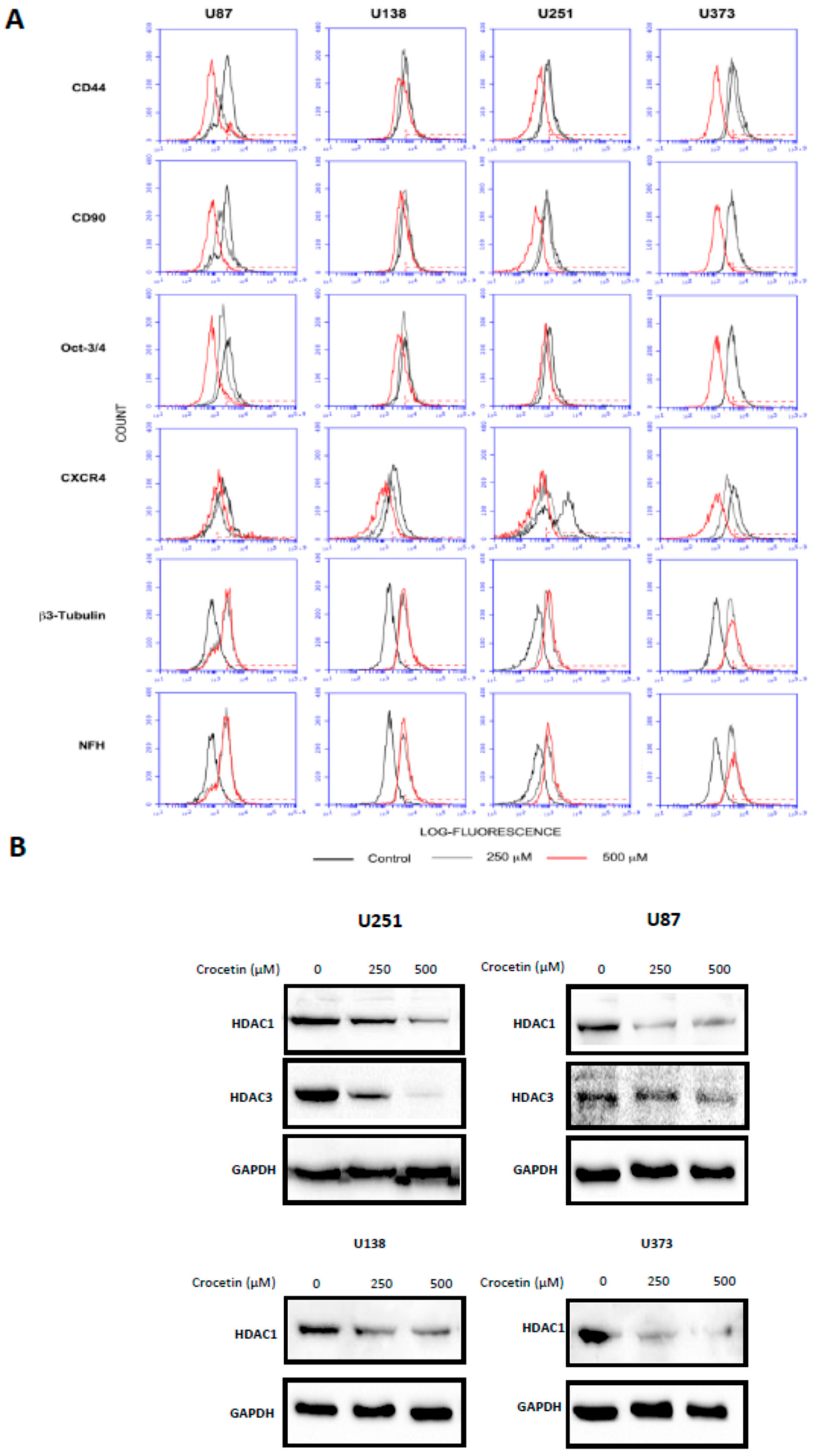
2.2. Crocetin Reduces the Levels of Mesenchymal Markers and Induces the Increase of Neuronal Markers in Glioma Cells
2.3. Crocetin Downmodulates HDACs in GBM Cancer Cells
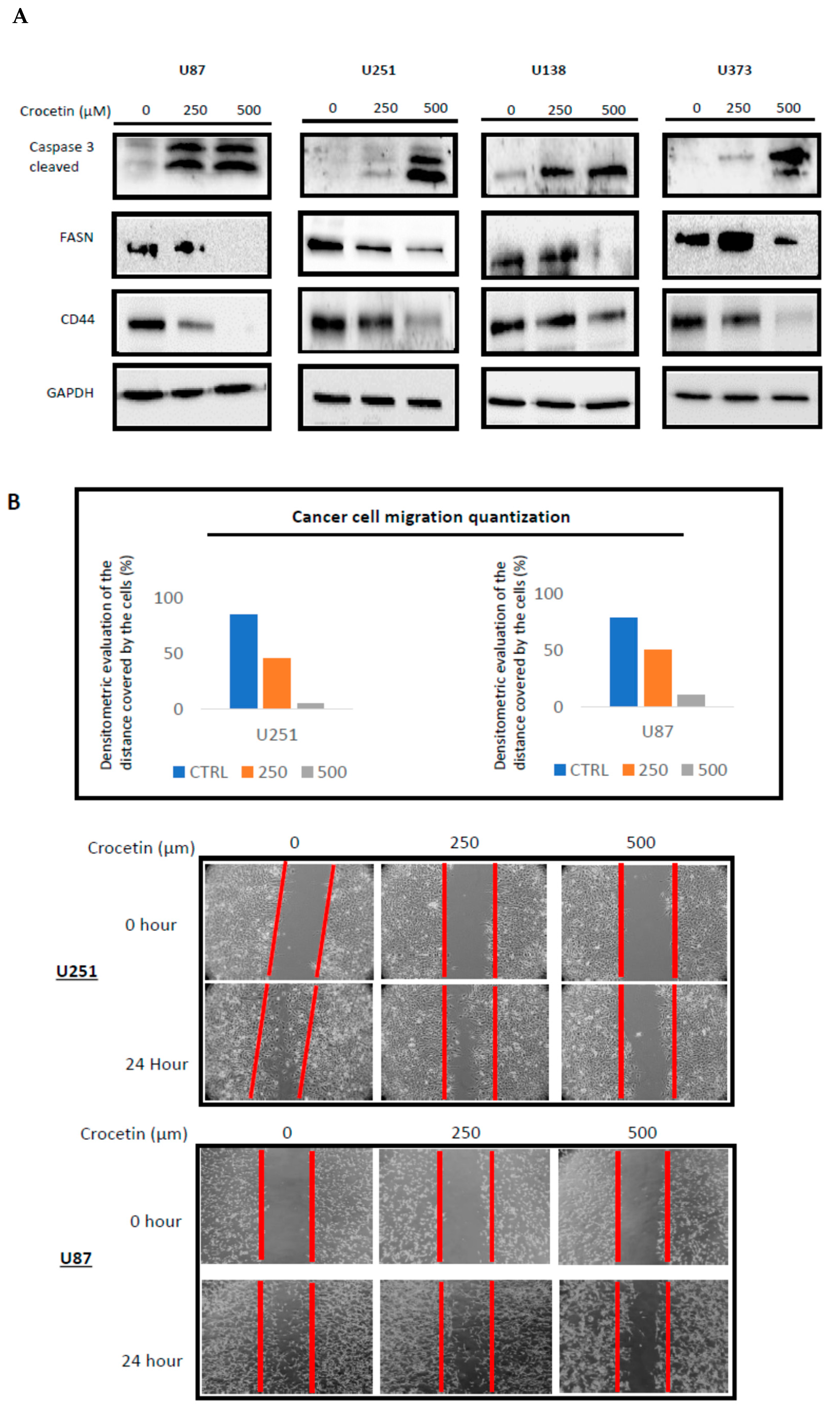
2.4. CCT Inhibits the Expression of FASN and CD44 Proteins, Inducing Cell Apoptosis and Reducing Migratory Capacity
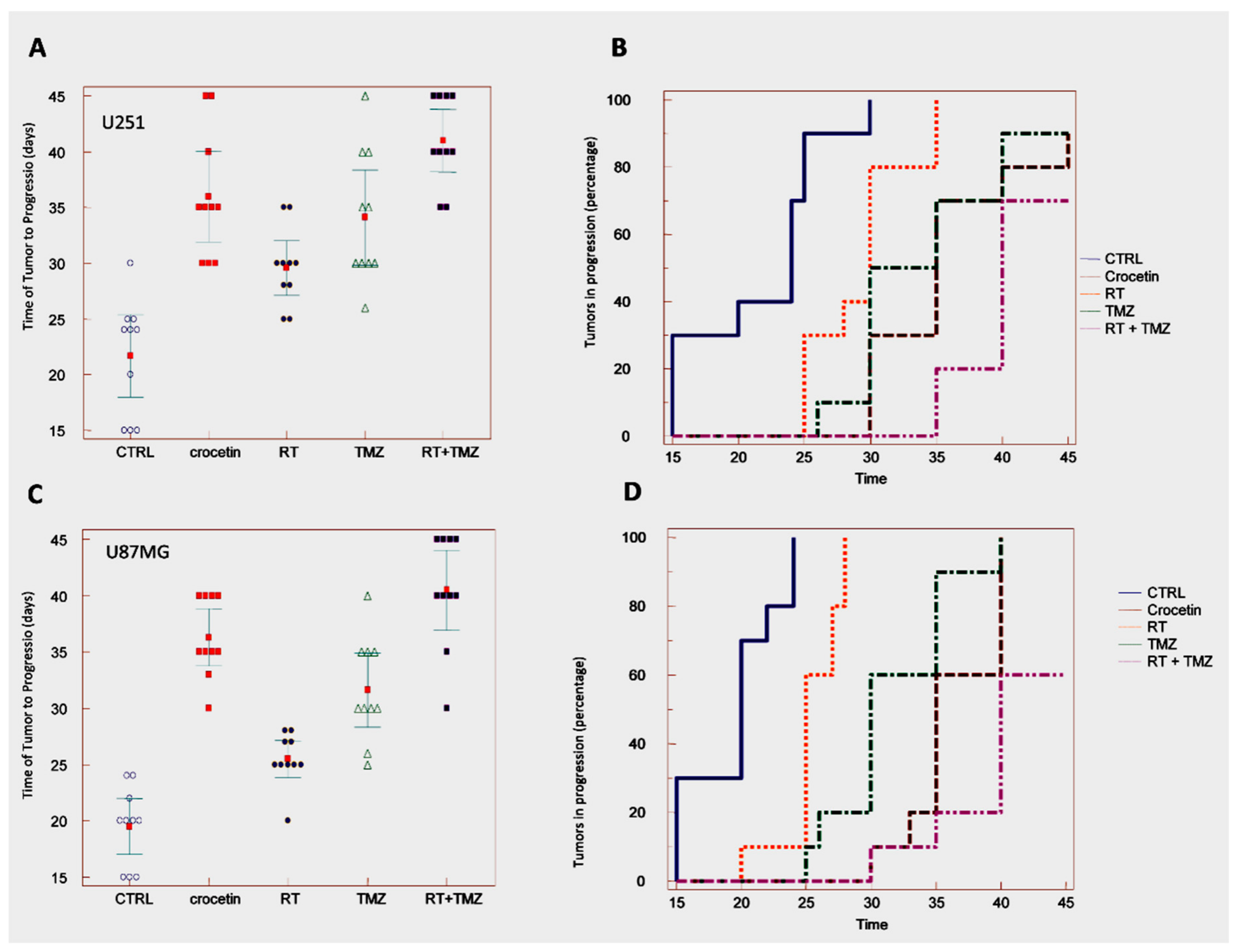
2.5. In Vivo Studies: Crocetin Modifies Tumour Growth of GBM Cells Subcutaneously Injected into Female nu/nu Mice (Subcutaneous Xenograft Model)
2.6. Crocetin Increased Disease-Free and Overall Survival in Orthotopic Intra-Brain Tumours as Determined by Using Differentiated Luc-U251MG Cell Model
3. Discussion
4. Materials and Methods
4.1. Crocetin Extraction
4.2. Cell Cultures and Cell Culture Materials
4.3. Western Blot
4.4. Fluorescence-Activated Cell Sorter Analysis
4.5. Wound Healing Repair Assay
4.6. Animal Experiments
4.6.1. Subcutaneous Xenograft Model
4.6.2. Evaluation of Treatment Response In Vivo
4.6.3. Orthotopic Intra-Brain Model
4.6.4. Evaluation of Treatment Response In Vivo
4.7. Histochemical and Immunohistochemical Analyses
4.8. Statistics
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gittleman, H.; Boscia, A.; Ostrom, Q.T.; Truitt, G.; Fritz, Y.; Kruchko, C.; Barnholtz-Sloan, J.S. Survivorship in Adults with Malignant Brain and other Central Nervous System Tumor from 2000–2014. Neuro Oncol. 2018, 20, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Sanai, N.; Polley, M.Y.; McDermott, M.W.; Parsa, A.T.; Berger, M.S. An extent of resection threshold for newly diagnosed glioblastoma. J. Neurosurg. 2011, 115, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Binda, E.; Visioli, A.; Reynolds, B.; Vescovi, A.L. Heterogeneity of cancer-initiating cells within glioblastoma. Front. Biosci. (Schol. Ed.) 2012, 4, 1235–1248. [Google Scholar] [PubMed]
- Nishikawa, M.; Inoue, A.; Ohnishi, T.; Kohno, S.; Ohue, S.; Matsumoto, S.; Suehiro, S.; Yamashita, D.; Ozaki, S.; Watanabe, H.; et al. Significance of Glioma Stem-Like Cells in the Tumor Periphery That Express High Levels of CD44 in Tumor Invasion, Early Progression, and Poor Prognosis in Glioblastoma. Stem Cells Int. 2018, 2018, 5387041. [Google Scholar] [CrossRef]
- Xu, H.; Tian, Y.; Yuan, X.; Wu, H.; Liu, Q.; Pestell, R.G.; Wu, K. The role of CD44 in epithelial-mesenchymal transition and cancer development. Onco Targets Ther. 2015, 8, 3783–3792. [Google Scholar]
- Wang, L.; Zuo, X.; Xie, K.; Wei, D. The Role of CD44 and Cancer Stem Cells. Methods Mol. Biol. 2018, 1692, 31–42. [Google Scholar]
- Avril, T.; Etcheverry, A.; Pineau, R.; Obacz, J.; Jegou, G.; Jouan, F.; Le Reste, P.J.; Hatami, M.; Colen, R.R.; Carlson, B.L.; et al. CD90 Expression Controls Migration and Predicts Dasatinib Response in Glioblastoma. Clin. Cancer Res. 2017, 23, 7360–7374. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. A decade of transcription factor-mediated reprogramming to pluripotency. Nat. Rev. Mol. Cell Biol. 2016, 17, 183–193. [Google Scholar] [CrossRef]
- Gravina, G.L.; Mancini, A.; Colapietro, A.; Delle Monache, S.; Sferra, R.; Vitale, F.; Cristiano, L.; Martellucci, S.; Marampon, F.; Mattei, V.; et al. The Small Molecule Ephrin Receptor Inhibitor, GLPG1790, Reduces Renewal Capabilities of Cancer Stem Cells, Showing Anti-Tumour Efficacy on Preclinical Glioblastoma Models. Cancers 2019, 11, 359. [Google Scholar] [CrossRef]
- Richardson, P.J. CXCR4 and Glioblastoma. Anticancer Agents Med. Chem. 2016, 16, 59–74. [Google Scholar] [CrossRef]
- Katsetos, C.D.; Draber, P.; Kavallaris, M. Targeting βIII-tubulin in glioblastoma multiforme: From cell biology and histopathology to cancer therapeutics. Anticancer Agents Med. Chem. 2011, 11, 719–728. [Google Scholar] [CrossRef] [PubMed]
- Miconi, G.; Palumbo, P.; Dehcordi, S.R.; La Torre, C.; Lombardi, F.; Evtoski, Z.; Cimini, A.M.; Galzio, R.; Cifone, M.G.; Cinque, B. Immunophenotypic characterization of human glioblastoma stem cells: Correlation with clinical outcome. J. Cell. Biochem. 2015, 116, 864–876. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, F.; Naderian, M.; Kadivar, M.; Nilipour, Y.; Gheytanchi, E. Expression of neuronal markers, NFP and GFAP, in malignant astrocytoma. Asian Pac. J. Cancer Prev. 2014, 15, 6315–6319. [Google Scholar] [CrossRef] [PubMed]
- Schcolnik-Cabrera, A.; Chávez-Blanco, A.; Domínguez-Gómez, G.; Taja-Chayeb, L.; Morales-Barcenas, R.; Trejo-Becerril, C.; Perez-Cardenas, E.; Gonzalez-Fierro, A.; Dueñas-González, A. Orlistat as a FASN inhibitor and multitargeted agent for cancer therapy. Expert Opin. Investig. Drugs 2018, 27, 475–489. [Google Scholar] [CrossRef]
- Grube, S.; Dünisch, P.; Freitag, D.; Klausnitzer, M.; Sakr, Y.; Walter, J.; Kalff, R.; Ewald, C. Overexpression of fatty acid synthase in human gliomas correlates with the WHO tumor grade and inhibition with Orlistat reduces cell viability and triggers apoptosis. J. Neurooncol. 2014, 118, 277–287. [Google Scholar] [CrossRef]
- Yasumoto, Y.; Miyazaki, H.; Vaidyan, L.K.; Kagawa, Y.; Ebrahimi, M.; Yamamoto, Y.; Ogata, M.; Katsuyama, Y.; Sadahiro, H.; Suzuki, M.; et al. Inhibition of fatty acid synthase decreases expression of stemness markers in glioma stem cells. PLoS ONE 2016, 11, e0147717. [Google Scholar] [CrossRef]
- Yamini, B. NF-κB, Mesenchymal Differentiation and Glioblastoma. Cells 2018, 7, 125. [Google Scholar] [CrossRef]
- Kawamura, Y.; Takouda, J.; Yoshimoto, K.; Nakashima, K. New aspects of glioblastoma multiforme revealed by similarities between neural and glioblastoma stem cells. Cell Biol. Toxicol. 2018, 34, 425–440. [Google Scholar] [CrossRef]
- Ramaiah, M.J.; Divyapriya, K.; Kumar, S.K.; Rajesh, Y.B.R.D. Drug-induced modifications and modulations of microRNAs and long non-coding RNAs for future therapy against Glioblastoma Multiforme. Gene 2020, 723, 144126. [Google Scholar] [CrossRef]
- Lee, D.H.; Ryu, H.W.; Won, H.R.; Kwon, S.H. Advances in epigenetic glioblastoma therapy. Oncotarget 2017, 8, 18577–18589. [Google Scholar] [CrossRef]
- Marampon, F.; Megiorni, F.; Camero, S.; Crescioli, C.; McDowell, H.P.; Sferra, R.; Vetuschi, A.; Pompili, S.; Ventura, L.; De Felice, F.; et al. HDAC4 and HDAC6 sustain DNA double strand break repair and stem-like phenotype by promoting radioresistance in glioblastoma cells. Cancer Lett. 2017, 397, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, S.; Selvaduray, K.R.; Radhakrishnan, A.K. Bioactive Compounds: Natural Defense against Cancer? Biomolecules 2019, 9, 758. [Google Scholar] [CrossRef] [PubMed]
- Vengoji, R.; Macha, M.A.; Batra, S.K.; Shonka, N.A. Natural products: A hope for glioblastoma patients. Oncotarget 2018, 9, 22194–22219. [Google Scholar] [CrossRef] [PubMed]
- Colapietro, A.; Mancini, A.; D’Alessandro, A.M.; Festuccia, C. Crocetin and Crocin from Saffron in Cancer Chemotherapy and Chemoprevention. Anti-Cancer Agents Med. Chem. 2019, 19, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Desai, V.; Bhushan, A. Natural Bioactive Compounds: Alternative Approach to the Treatment of Glioblastoma Multiforme. BioMed Res. Int. 2017, 2017, 9363040. [Google Scholar] [CrossRef] [PubMed]
- Festuccia, C.; Mancini, A.; Gravina, G.L.; Scarsella, L.; Llorens, S.; Alonso, G.L.; Tatone, C.; Di Cesare, E.; Jannini, E.A.; Lenzi, A.; et al. Antitumor effects of saffron-derived carotenoids in prostate cancer cell models. Biomed. Res. Int. 2014, 2014, 135048. [Google Scholar] [CrossRef] [PubMed]
- Suparmi, S.; de Haan, L.; Spenkelink, A.; Louisse, J.; Beekmann, K.; Rietjens, I.M.C.M. Combining in vitro data and physiologically based kinetic modeling facilitated reverse dosimetry to define in vivo dose-response curves for bixin- and crocetin-induced activation of PPARγ in humans. Mol. Nutr. Food Res. 2019, e1900880. [Google Scholar] [CrossRef]
- Deng, L.; Li, J.; Lu, S.; Su, Y. Crocin inhibits proliferation and induces apoptosis through suppressing MYCN expression in retinoblastoma. J. Biochem. Mol. Toxicol. 2019, 33, e22292. [Google Scholar] [CrossRef]
- Kim, S.H.; Lee, J.M.; Kim, S.C.; Park, C.B.; Lee, P.C. Proposed cytotoxic mechanisms of the saffron carotenoids crocin and crocetin on cancer cell lines. Biochem. Cell Biol. 2014, 92, 105–111. [Google Scholar] [CrossRef]
- Gezici, S. Comparative anticancer activity analysis of saffron extracts and a principle component, crocetin for prevention and treatment of human malignancies. J. Food Sci. Technol. 2019, 56, 5435–5443. [Google Scholar] [CrossRef]
- Neyshaburinezhad, N.; Kalalinia, F.; Hashemi, M. Encapsulation of crocetin into poly (lactic-co-glycolic acid) nanoparticles overcomes drug resistance in human ovarian cisplatin-resistant carcinoma cell line (A2780-RCIS). Mol. Biol. Rep. 2019, 46, 6525–6532. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Qu, Y.; Shen, X.Y.; Ouyang, T.; Fu, W.B.; Luo, T.; Wang, H.Q. Multiple Signal Pathways Involved in Crocetin-Induced Apoptosis in KYSE-150 Cells. Pharmacology 2019, 103, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Moradzadeh, M.; Ghorbani, A.; Erfanian, S.; Mohaddes, S.T.; Rahimi, H.; Ghayoor karimiani, S.; Mashkani, B.; Chiang, S.C.; El-Khamishi, S.F.; Tabarraei, A.; et al. Study of the mechanisms of crocetin-induced differentiation and apoptosis in human acute promyelocytic leukemia cells. J. Cell. Biochem. 2018, 120, 1943–1957. [Google Scholar] [CrossRef] [PubMed]
- Costea, T.; Hudiță, A.; Ciolac, O.A.; Gălățeanu, B.; Ginghină, O.; Costache, M.; Ganea, C.; Mocanu, M.M. Chemoprevention of Colorectal Cancer by Dietary Compounds. Int. J. Mol. Sci. 2018, 19, 3787. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Kridel, S.; Thorburn, A.; Kooshki, M.; Little, J.; Hebbar, S.; Robbins, M. Fatty acid synthase: A novel target for antiglioma therapy. Br. J. Cancer 2006, 95, 869–878. [Google Scholar] [CrossRef]
- Wakil, S.J. Fatty acid synthase, a proficient multifunctional enzyme. Biochemistry 1989, 28, 4523–4530. [Google Scholar] [CrossRef]
- Harabin-Slowinska, M.; Slowinski, J.; Konecki, J.; Mrowka, R. Expression of adhesion molecule CD44 in metastatic brain tumors. Folia neuropathologica/Association of Polish Neuropathologists and Medical Research centre. Polish Acad. Sci. 1998, 36, 179–184. [Google Scholar]
- Lim, S.; Kim, D.; Ju, S.; Shin, S.; Cho, I.; Park, S.-H.; Grailhe, R.; Lee, C.; Kim, Y.K. Glioblastoma-secreted soluble CD44 activates tau pathology in the brain. Exp. Mol. Med. 2018, 50, 5. [Google Scholar] [CrossRef]
- Wei, K.C.; Huang, C.Y.; Chen, P.Y.; Feng, L.Y.; Wu, T.W.E.; Chen, S.M.; Tsai, H.C.; Lu, Y.J.; Tsang, N.M.; Tseng, C.K.; et al. Evaluation of the prognostic value of CD44 in glioblastoma multiforme. Anticancer Res. 2010, 30, 253–259. [Google Scholar]
- Brown, D.V.; Daniel, P.M.; Giovanna, M.D.; Gogos, A.; Ng, W.; Morokoff, A.P.; Mantamadiotis, T. Coexpression analysis of CD133 and CD44 identifies Proneural and Mesenchymal subtypes of glioblastoma multiforme. Oncotarget 2015, 6, 6267–6280. [Google Scholar] [CrossRef]
- Karkoula, E.; Lemonakis, N.; Kokras, N.; Dalla, C.; Gikas, E.; Skaltsounis, A.L.; Tsarbopoulos, A. Trans-crocin 4 is not hydrolyzed to crocetin following i.p. administration in mice, while it shows penetration through the blood brain barrier. Fitoterapia 2018, 129, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Llorens, S.; Mancini, A.; Serrano-Díaz, J.; D’Alessandro, A.M.; Nava, E.; Alonso, G.L.; Carmona, M. Effects of Crocetin Esters and Crocetin from Crocus sativus L. on Aortic Contractility in Rat Genetic Hypertension. Molecules 2015, 20, 17570–17584. [Google Scholar] [CrossRef] [PubMed]
- Gravina, G.L.; Mancini, A.; Colapietro, A.; Delle Monache, S.; Sferra, R.; Pompili, S.; Vitale, F.; Martellucci, S.; Marampon, F.; Mattei, V.; et al. The Brain Penetrating and Dual TORC1/TORC2 Inhibitor, RES529, Elicits Anti-Glioma Activity and Enhances the Therapeutic Effects of Anti-Angiogenetic Compounds in Preclinical Murine Models. Cancers 2019, 11, 1604. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
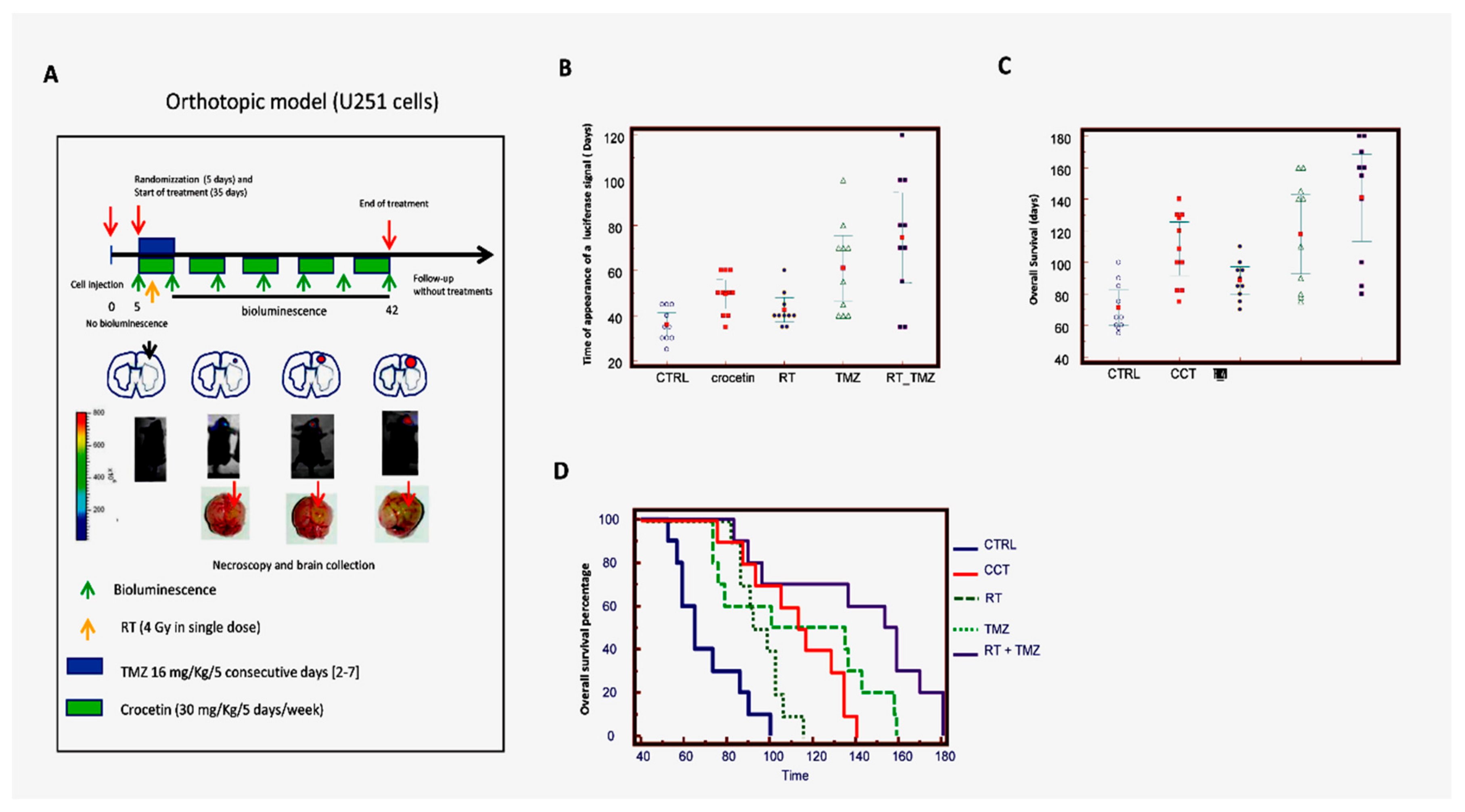{kind=link}
| % U87 | CD44 | CD90 | OCT3/4 | CXCR4 | β3 Tubulin. | NFH |
|---|---|---|---|---|---|---|
| Controls (CTRL) | 48.2 | 44.1 | 57.3 | 74.7 | 3.5 | 3.4 |
| CCT 250 μM | 16.6 | 24.7 | 21.0 | 55.8 | 37.5 | 39.1 |
| CCT 500 μM | 11.6 | 3.5 | 6.2 | 56.1 | 42.3 | 40.3 |
| % U138 | ||||||
| Control | 53.4 | 54 | 50.5 | 69.8 | 1.5 | 1.5 |
| CCT 250 μM | 43.4 | 46.5 | 46.9 | 35.5 | 40.9 | 42 |
| CCT 500 μM | 30.1 | 30.5 | 22.8 | 15 | 44.1 | 44.9 |
| % U251 | ||||||
| Control | 42.1 | 48.5 | 53.2 | 61.6 | 2.0 | 2.9 |
| CCT 250 μM | 32.7 | 27.9 | 27.2 | 34.5 | 35.6 | 32.3 |
| CCT 500 μM | 6.4 | 2.3 | 27.9 | 12.5 | 52.9 | 49.7 |
| % U373 | ||||||
| ControlL | 72 | 50.3 | 48.2 | 50.3 | 1.9 | 1.7 |
| CCT 250 μM | 47.8 | 48.6 | 47 | 14.9 | 42.6 | 47.3 |
| CCT 500 μM | 1.8 | 2.2 | 1.8 | 1.2 | 50.5 | 58.6 |
| U251 | Mean ± SD | Statistics |
| Control | 21.7 ± 5.2 | |
| Crocetin | 36.0 ± 5.7 | p < 0.0001 vs. CTRL |
| RT | 29.6 ± 3.4 | p = 0.0010 vs. CTRL p = 0.0081 vs. CCT |
| TMZ | 34.1 ± 6.0 | p < 0.0001 vs. CTRL p = 0.4759 vs. CCT (Not Significant, NS) |
| RT + TMZ | 41.0 ± 3.9 | p < 0.0001 vs. CTRL p = 0.0361 vs. CCT |
| U87MG | Mean ± SD | Statistics |
| Control | 19.5 ± 3.5 | |
| Crocetin | 36.3 ± 3.5 | p < 0.0001 vs. CTRL |
| RT | 25.5 ± 2.3 | p = 0.0003 vs. CTRL p < 0.0001 vs. CCT |
| TMZ | 31.6 ± 4.6 | p < 0.0001 vs. CTRL p = 0.0201 vs. CCT |
| RT + TMZ | 40.5 ± 5.0 | p < 0.0001 vs. CTRL p = 0.0447 vs. CCT |
| U251 | Hazard Ratio | CI 95% | Statistics |
| CTRL vs. CCT | 4.58 | 1.49 to 14.10 | p < 0.0001 |
| CTRL vs. RT | 3.08 | 1.11 to 8.54 | p < 0.0011 |
| CTRL vs. TMZ | 4.05 | 1.36 to 12.10 | p < 0.0001 |
| CTRL vs. RT + TMZ | 5.34 | 1.66 to 17.11 | p < 0.0001 |
| RT vs. CCT | 2.75 | 1.01 to 7.45 | p = 0.0044 |
| TMZ vs. CCT | 1.35 | 0.52 to 3.50 | p = 0.4545 (NS) |
| CCT vs. RT + TMZ | 2.36 | 0.83 to 6.37 | p = 0.0465 |
| U87MG | Hazard Ratio | CI 95% | statistics |
| CTRL vs. CCT | 4.69 | 1.46 to 13.95 | p < 0.0001 |
| CTRL vs. RT | 2.79 | 1.19 to 8.15 | p = 0.0004 |
| CTRL vs. TMZ | 4.39 | 1.42 to 13.55 | p < 0.0001 |
| CTRL vs. RT + TMZ | 5.52 | 1.73 to 17.62 | p < 0.0001 |
| RT vs. CCT | 3.30 | 1.40 to 13.18 | p < 0.0001 |
| TMZ vs. CCT | 1.67 | 0.67 to 4.12 | p = 0.1059 (NS) |
| CCT vs. RT + TMZ | 2.42 | 0.89 to 6.57 | p = 0.0219 |
| U251 | Mean ± SD | Statistics |
|---|---|---|
| Control | 36.0 ± 7.4 | |
| Crocetin | 49.5 ± 9.9 | p = 0.0019 vs. CTRL |
| RT | 42.5 ± 7.5 | p = 0.0472 vs. CTRL p < 0.0760 vs. CCT (NS) |
| TMZ | 61.0 ± 20.4 | p = 0.0038 vs. CTRL p = 0.1284 vs. CCT (NS) |
| RT + TMZ | 64.5 ± 27.8 | p < 0.0001 vs. CTRL p = 0.0205 vs. CCT |
| U251 | Mean ± SD | Statistics |
|---|---|---|
| Control | 71.3 ± 15.5 | |
| Crocetin | 108.7 ± 23.8 | p = 0.0008 vs. CTRL |
| RT | 88.5 ± 12.0 | p = 0.0129 vs. CTRL p = 0.0324 vs. CCT |
| TMZ | 117.8 ± 34.9 | p = 0.0023 vs. CTRL p = 0.5055 vs. CCT (NS) |
| RT + TMZ | 141.0 ± 38.5 | p = 0.0002 vs. CTRLP = 0.0394 vs. CCT |
| U251 | Hazard Ratio | CI 95% | Statistics |
|---|---|---|---|
| CTRL vs. CCT | CTRL vs. CCT | 3.97 | 1.69 to 10.55 |
| CTRL vs. RT | CTRL vs. RT | 2.26 | 0.84 to 6.85 |
| CTRL vs. TMZ | RT vs. CCT | 2.67 | 0.93 to 8.49 |
| CTRL vs. RT + TMZ | CTRL vs. TMZ | 3.35 | 1.18 to 9.51 |
| RT vs. CCT | CCT vs. TMZ | 1.71 | 0.69 to 4.24 |
| TMZ vs. CCT | RT + TMZ vs. CTRL | 4.63 | 2.12 to 12.87 |
| CCT vs. RT + TMZ | CCT vs. RT + TMZ | 2.47 | 0.99 to 6.87 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Colapietro, A.; Mancini, A.; Vitale, F.; Martellucci, S.; Angelucci, A.; Llorens, S.; Mattei, V.; Gravina, G.L.; Alonso, G.L.; Festuccia, C. Crocetin Extracted from Saffron Shows Antitumor Effects in Models of Human Glioblastoma. Int. J. Mol. Sci. 2020, 21, 423. https://doi.org/10.3390/ijms21020423
Colapietro A, Mancini A, Vitale F, Martellucci S, Angelucci A, Llorens S, Mattei V, Gravina GL, Alonso GL, Festuccia C. Crocetin Extracted from Saffron Shows Antitumor Effects in Models of Human Glioblastoma. International Journal of Molecular Sciences. 2020; 21(2):423. https://doi.org/10.3390/ijms21020423
Chicago/Turabian StyleColapietro, Alessandro, Andrea Mancini, Flora Vitale, Stefano Martellucci, Adriano Angelucci, Silvia Llorens, Vincenzo Mattei, Giovanni Luca Gravina, Gonzalo Luis Alonso, and Claudio Festuccia. 2020. "Crocetin Extracted from Saffron Shows Antitumor Effects in Models of Human Glioblastoma" International Journal of Molecular Sciences 21, no. 2: 423. https://doi.org/10.3390/ijms21020423
APA StyleColapietro, A., Mancini, A., Vitale, F., Martellucci, S., Angelucci, A., Llorens, S., Mattei, V., Gravina, G. L., Alonso, G. L., & Festuccia, C. (2020). Crocetin Extracted from Saffron Shows Antitumor Effects in Models of Human Glioblastoma. International Journal of Molecular Sciences, 21(2), 423. https://doi.org/10.3390/ijms21020423