Cannabidiol-Mediated Changes to the Phospholipid Profile of UVB-Irradiated Keratinocytes from Psoriatic Patients

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Comparison of the Phospholipid Profile of Healthy Keratinocytes and Psoriatic Keratinocytes Not Treated or Treated with CBD (Control vs. CBD vs. Ps vs. Ps + CBD)
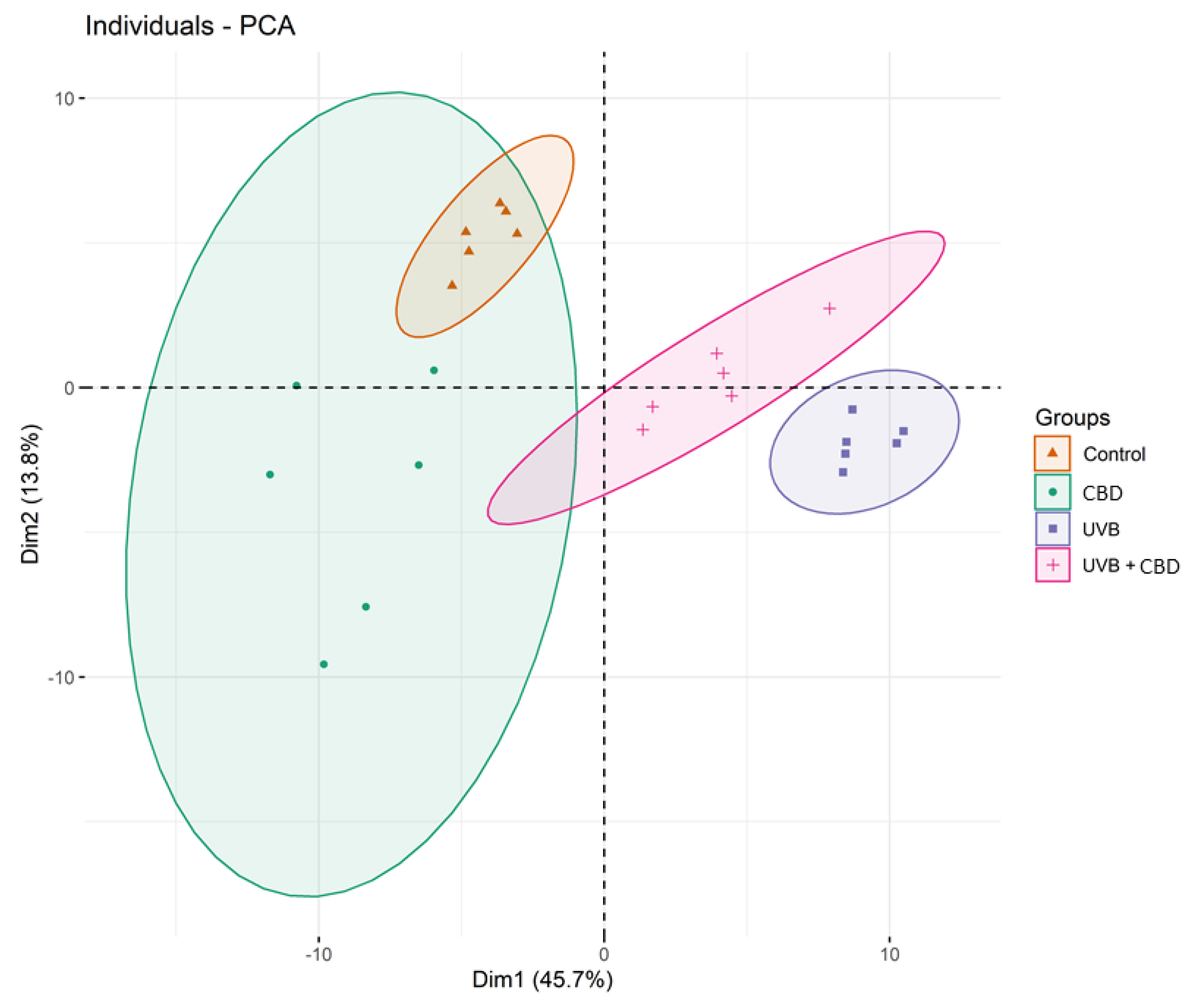
2.2. Comparison of the Phospholipid Profile of Healthy Keratinocytes Not Treated and Treated with CBD or/and UVB (Control vs. CBD vs. UVB vs. UVB + CBD)
2.3. Comparison of the Phospholipid Profile of Psoriatic Keratinocytes Untreated and Treated with CBD (4 μM) or/and UVB (60 mJ/cm2) (Ps vs. Ps + CBD vs. Ps + UVB vs. Ps + UVB + CBD)
3. Discussion
4. Materials and Methods
4.1. Reagents/Chemicals
4.2. Collection of Skin Samples
4.3. Cell Culture and Treatment
- Group 1 [Control]: Keratinocytes from the skin of healthy subjects cultured in standard medium.
- Group 2 [CBD]: Keratinocytes of the skin of healthy subjects cultured for 24 h in a medium containing 4 µM of CBD.
- Group 3 [UVB]: Keratinocytes of the skin of healthy subjects exposed to UVB radiation.
- Group 4 [UVB + CBD]: Keratinocytes of the skin of healthy subjects exposed to UVB radiation and then cultured for 24 h in a medium containing 4 µM CBD.
- Group 5 [Ps]: Keratinocytes of the skin of psoriatic patients cultured in standard medium.
- Group 6 [Ps + CBD]: Keratinocytes of the skin of psoriatic patients cultured for 24 h in a medium containing 4 µM CBD.
- Group 7 [Ps + UVB]: Keratinocytes of the skin of psoriatic patients exposed to UVB radiation.
- Group 8 [Ps + UVB + CBD]: Keratinocytes of the skin of psoriatic patients exposed to UVB irradiation and cultured for 24 h in a medium containing 4 µM CBD.
4.4. Lipidomic Analysis
4.4.1. Lipid Extraction
4.4.2. Quantification of Phospholipids by Measurement of Phosphorus
4.4.3. UPLC-ESI-MS and MS/MS Analysis
4.4.4. Identification of Phospholipids
4.4.5. Data Treatment
4.4.6. Statistical Analysis
4.5. Physicochemical Properties of Keratinocyte Cell Membranes
4.5.1. Analysis of the Zeta Potential
4.5.2. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 4-HNE | 4-hydroxy-2-onenal |
| ANOVA | Analysis of variance |
| CBD | Cannabidiol |
| ESI | Electrospray ionization |
| HILIC | Hydrophilic interaction liquid chromatography |
| HPLC | High performance liquid chromatography |
| LPC | Lysophosphatidylcholine |
| LPE | Lysophosphoethanolamine |
| MDA | Malondialdehyde |
| PASI | Psoriasis area and severity index |
| PCA | Principal component analysis |
| PC | Phosphatidylcholine |
| PE | Phosphatidylethanolamine |
| PEo | Ether-linked phosphoethanolamine |
| PI | Phosphatidylinositol |
| PS | Phosphatidylserines |
| PLA2 | Phospholipase A2 |
| PLS-DA | Partial least squares-discriminate analysis |
| PUFAs | Polyunsaturated fatty acids |
| QTOF | Quadrupole time of flight mass spectrometer |
| ROS | Reactive oxygen species |
| SM | Sphingomyelin |
References
- Fernandis, A.Z.; Wenk, M.R. Membrane lipids as signaling molecules. Curr. Opin. Lipidol. 2007, 18, 121–128. [Google Scholar] [CrossRef]
- Hubler, M.J.; Kennedy, A.J. Role of lipids in the metabolism and activation of immune cells. J. Nutr. Biochem. 2016, 34, 1–7. [Google Scholar] [CrossRef]
- Berdyshev, E.; Goleva, E.; Bronova, I.A.; Dyjack, N.; Rios, C.; Jung, J.; Taylor, P.; Jeong, M.; Hall, C.F.; Richers, B.N.; et al. Lipid abnormalities in atopic skin are driven by type 2 cytokines. JCI Insight 2018, 3, 3. [Google Scholar] [CrossRef]
- Li, S.; Villarreal, M.; Stewart, S.; Choi, J.; Ganguli-Indra, G.; Babineau, D.; Philpot, C.; David, G.; Yoshida, T.; Boguniewicz, M.; et al. Altered composition of epidermal lipids correlates with Staphylococcus aureus colonization status in atopic dermatitis. Br. J. Dermatol. 2017, 177, e125–e127. [Google Scholar] [CrossRef] [PubMed]
- Pietrzak, A.; Michalak-Stoma, A.; Chodorowska, G.; Szepietowski, J.C. Lipid Disturbances in Psoriasis: An Update. Mediat. Inflamm. 2010, 2010, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wójcik, P.; Biernacki, M.; Wroński, A.; Łuczaj, W.; Waeg, G.; Zarkovic, N.; Skrzydlewska, E. Altered Lipid Metabolism in Blood Mononuclear Cells of Psoriatic Patients Indicates Differential Changes in Psoriasis Vulgaris and Psoriatic Arthritis. Int. J. Mol. Sci. 2019, 20, 4249. [Google Scholar] [CrossRef] [PubMed]
- Rendon, A.; Schäkel, K. Psoriasis Pathogenesis and Treatment. Int. J. Mol. Sci. 2019, 20, 1475. [Google Scholar] [CrossRef]
- Akhyani, M.; Ehsani, A.; Robati, R.M.; Robati, A. The lipid profile in psoriasis: A controlled study. J. Eur. Acad. Dermatol. Venereol. 2007, 21, 1330–1332. [Google Scholar] [CrossRef]
- Bajaj, D.R.; Mahesar, S.M.; Devrajani, B.R.; Iqbal, M.P. Lipid profile in patients with psoriasis presenting at Liaquat University Hospital Hyderabad. J. Pak. Med. Assoc. 2009, 59, 512–515. [Google Scholar]
- Farshchian, M.; Zamanian, A.; Monsef, A.-R.; Mahjub, H. Serum lipid level in Iranian patients with psoriasis. J. Eur. Acad. Dermatol. Venereol. 2007, 21, 802–805. [Google Scholar] [CrossRef]
- Zeng, C.; Wen, B.; Hou, G.; Lei, L.; Mei, Z.; Jia, X.; Chen, X.; Zhu, W.; Li, J.; Kuang, Y.; et al. Lipidomics profiling reveals the role of glycerophospholipid metabolism in psoriasis. GigaScience 2017, 6, 1–11. [Google Scholar] [CrossRef]
- Koyano, S.; Hatamochi, A.; Yamazaki, S.; Ishikawa, J.; Kitahara, T.; Narita, H.; Kondo, N.; Masukawa, Y. Psoriasis Patients Have Abnormal Ceramide Profile in Stratum Corneum. Nishi Nihon Hifuka 2010, 72, 494–499. [Google Scholar] [CrossRef]
- Łuczaj, W.; Wroński, A.; Domingues, P.; Domingues, M.R.; Skrzydlewska, E. Domingues Lipidomic Analysis Reveals Specific Differences between Fibroblast and Keratinocyte Ceramide Profile of Patients with Psoriasis Vulgaris. Molecules 2020, 25, 630. [Google Scholar] [CrossRef]
- Tawada, C.; Kanoh, H.; Nakamura, M.; Mizutani, Y.; Fujisawa, T.; Banno, Y.; Seishima, M. Interferon-γ Decreases Ceramides with Long-Chain Fatty Acids: Possible Involvement in Atopic Dermatitis and Psoriasis. J. Investig. Dermatol. 2014, 134, 712–718. [Google Scholar] [CrossRef] [PubMed]
- Tawada, C.; Kanoh, H.; Banno, Y.; Nakamura, Y.; Seishima, M. Analysis of ceramide profiles in stratum corneum of atopic dermatitis and psoriasis by MALDI-TOF-MS. J. Dermatol. 2012, 39, 67. [Google Scholar]
- Jarocka-Karpowicz, I.; Biernacki, M.; Wroński, A.; Gęgotek, A.; Skrzydlewska, E. Cannabidiol Effects on Phospholipid Metabolism in Keratinocytes from Patients with Psoriasis Vulgaris. Biomolecules 2020, 10, 367. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.B.; Jerome, D.; Yeung, J. Diagnosis and management of psoriasis. Can. Fam. Physician Med. Fam. Can. 2017, 63, 278–285. [Google Scholar]
- Lee, C.-H.; Wu, S.-B.; Hong, C.-H.; Yu, H.-S.; Wei, Y.-H. Molecular Mechanisms of UV-Induced Apoptosis and Its Effects on Skin Residential Cells: The Implication in UV-Based Phototherapy. Int. J. Mol. Sci. 2013, 14, 6414–6435. [Google Scholar] [CrossRef] [PubMed]
- Morales, P.; Reggio, P.H.; Jagerovic, N. An Overview on Medicinal Chemistry of Synthetic and Natural Derivatives of Cannabidiol. Front. Pharmacol. 2017, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Burstein, S.H. Cannabidiol (CBD) and its analogs: A review of their effects on inflammation. Bioorganic Med. Chem. 2015, 23, 1377–1385. [Google Scholar] [CrossRef] [PubMed]
- Pellati, F.; Borgonetti, V.; Brighenti, V.; Biagi, M.; Benvenuti, S.; Corsi, L. Cannabis sativa L. and Nonpsychoactive Cannabinoids: Their Chemistry and Role against Oxidative Stress, Inflammation, and Cancer. BioMed Res. Int. 2018, 2018, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Atalay, S.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Antioxidative and Anti-Inflammatory Properties of Cannabidiol. Antioxidants 2019, 9, 21. [Google Scholar] [CrossRef] [PubMed]
- Jastrząb, A.; Gęgotek, A.; Skrzydlewska, E. Cannabidiol Regulates the Expression of Keratinocyte Proteins Involved in the Inflammation Process through Transcriptional Regulation. Cells 2019, 8, 827. [Google Scholar] [CrossRef]
- Sheriff, T.; Lin, M.J.; Dubin, D.; Khorasani, H. The potential role of cannabinoids in dermatology. J. Dermatol. Treat. 2019, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, J.D.; Williamson, E.M. Cannabinoids inhibit human keratinocyte proliferation through a non-CB1/CB2 mechanism and have a potential therapeutic value in the treatment of psoriasis. J. Dermatol. Sci. 2007, 45, 87–92. [Google Scholar] [CrossRef]
- Baxter, A.; Hulett, M.D.; Poon, I.K.H. The phospholipid code: A key component of dying cell recognition, tumor progression and host–microbe interactions. Cell Death Differ. 2015, 22, 1893–1905. [Google Scholar] [CrossRef]
- Watson, H. Biological membranes. Essays Biochem. 2015, 59, 43–69. [Google Scholar] [CrossRef]
- Vance, J.E. Historical perspective: Phosphatidylserine and phosphatidylethanolamine from the 1800s to the present. J. Lipid Res. 2018, 59, 923–944. [Google Scholar] [CrossRef]
- Choudhary, V.; Olala, L.O.; Qin, H.; Helwa, I.; Pan, Z.-Q.; Tsai, Y.-Y.; Frohman, M.A.; Kaddour-Djebbar, I.; Bollag, W.B. Aquaporin-3 Re-Expression Induces Differentiation in a Phospholipase D2-Dependent Manner in Aquaporin-3-Knockout Mouse Keratinocytes. J. Investig. Dermatol. 2015, 135, 499–507. [Google Scholar] [CrossRef]
- Cai, T.; Yang, F. Phospholipid and Phospholipidomics in Health and Diseases. In Translational Bioinformatics; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2018; pp. 177–202. [Google Scholar]
- Franco, J.; Ferreira, C.; Sobreira, T.J.P.; Sundberg, J.P.; HogenEsch, H. Profiling of epidermal lipids in a mouse model of dermatitis: Identification of potential biomarkers. PLoS ONE 2018, 13, e196595. [Google Scholar] [CrossRef]
- Santinha, D.; Dória, M.L.; Neves, B.M.; Maciel, E.; Martins, J.; Helguero, L.; Domingues, P.; Cruz, M.T.; Domingues, M.R. Prospective phospholipid markers for skin sensitization prediction in keratinocytes: A phospholipidomic approach. Arch. Biochem. Biophys. 2013, 533, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Jonas, A. Lecithin cholesterol acyltransferase. Biochim. Biophys. Acta (BBA) Mol. Cell Boil. Lipids 2000, 1529, 245–256. [Google Scholar] [CrossRef]
- Cigliano, L.; Maresca, B.; Salvatore, A.; Nino, M.; Monfrecola, G.; Ayala, F.; Carlucci, A.; Pugliese, R.; Pedone, C.; Abrescia, P. Haptoglobin from psoriatic patients exhibits decreased activity in binding haemoglobin and inhibiting lecithin-cholesterol acyltransferase activity. J. Eur. Acad. Dermatol. Venereol. 2008, 22, 417–425. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Holzer, M.; Wolf, P.; Inzinger, M.; Trieb, M.; Curcic, S.; Pasterk, L.; Weger, W.; Heinemann, A.; Marsche, G. Anti-psoriatic therapy recovers high-density lipoprotein composition and function. J. Investig. Dermatol. 2013, 134, 635–642. [Google Scholar] [CrossRef]
- Wanner, R.; Peiser, M.; Wittig, B. Keratinocytes Rapidly Readjust Ceramide–Sphingomyelin Homeostasis and Contain a Phosphatidylcholine–Sphingomyelin Transacylase. J. Investig. Dermatol. 2004, 122, 773–782. [Google Scholar] [CrossRef]
- Uchida, Y.; Hara, M.; Nishio, H.; Sidransky, E.; Inoue, S.; Otsuka, F.; Suzuki, A.; Elias, P.M.; Holleran, W.M.; Hamanaka, S. Epidermal sphingomyelins are precursors for selected stratum corneum ceramides. J. Lipid Res. 2000, 41, 2071–2082. [Google Scholar]
- Alessandrini, F.; Pfister, S.; Kremmer, E.; Gerber, J.-K.; Ring, J.; Behrendt, H. Alterations of Glucosylceramide-β-Glucosidase Levels in the Skin of Patients with Psoriasis Vulgaris. J. Investig. Dermatol. 2004, 123, 1030–1036. [Google Scholar] [CrossRef]
- Cha, H.J.; He, C.; Zhao, H.; Dong, Y.; An, I.-S.; Kim, Y.J. Intercellular and intracellular functions of ceramides and their metabolites in skin (Review). Int. J. Mol. Med. 2016, 38, 16–22. [Google Scholar] [CrossRef]
- Bondar, O.V.; Saifullina, D.V.; Shakhmaeva, I.I.; Mavlyutova, I.I.; Abdullin, T.I. Kazan (Volga Region) Federal University Monitoring of the Zeta Potential of Human Cells upon Reduction in Their Viability and Interaction with Polymers. Acta Nat. 2012, 4, 78–81. [Google Scholar] [CrossRef]
- Haupt, A.; Minc, N. Gradients of phosphatidylserine contribute to plasma membrane charge localization and cell polarity in fission yeast. Mol. Boil. Cell 2017, 28, 210–220. [Google Scholar] [CrossRef]
- Yeung, T.; Gilbert, G.E.; Shi, J.; Silvius, J.; Kapus, A.; Grinstein, S. Membrane Phosphatidylserine Regulates Surface Charge and Protein Localization. Science 2008, 319, 210–213. [Google Scholar] [CrossRef] [PubMed]
- Chaurio, R.A.; Janko, C.; Munoz, L.; Frey, B.; Herrmann, M.; Gaipl, U. Phospholipids: Key Players in Apoptosis and Immune Regulation. Molecules 2009, 14, 4892–4914. [Google Scholar] [CrossRef] [PubMed]
- Kay, J.G.; Grinstein, S. Sensing Phosphatidylserine in Cellular Membranes. Sensors 2011, 11, 1744–1755. [Google Scholar] [CrossRef] [PubMed]
- Wójcik, P.; Zarkovic, N.; Gęgotek, A.; Skrzydlewska, E. Involvement of Metabolic Lipid Mediators in the Regulation of Apoptosis. Biomolecules 2020, 10, 402. [Google Scholar] [CrossRef]
- Soliman, E.; Henderson, K.L.; Danell, A.S.; Van Dross, R. Arachidonoyl-ethanolamide activates endoplasmic reticulum stress-apoptosis in tumorigenic keratinocytes: Role of cyclooxygenase-2 and novel J-series prostamides. Mol. Carcinog. 2015, 55, 117–130. [Google Scholar] [CrossRef]
- Kagan, V.E.; Bayır, H.; Tyurina, Y.Y.; Bolevich, S.B.; Maguire, J.J.; Fadeel, B.; Balasubramanian, K. Elimination of the unnecessary: Intra- and extracellular signaling by anionic phospholipids. Biochem. Biophys. Res. Commun. 2017, 482, 482–490. [Google Scholar] [CrossRef]
- Huang, T.; Lin, X.; Meng, X.; Lin, M. Phosphoinositide-3 Kinase/Protein Kinase-B/Mammalian Target of Rapamycin Pathway in Psoriasis Pathogenesis. A Potential Therapeutic Target? Acta Derm. Venereol. 2014, 94, 371–379. [Google Scholar] [CrossRef]
- Pike, M.C.; Lee, C.S.; Elder, J.T.; Voorhees, J.J.; Fisher, G.J. Increased Phosphatidylinositol Kinase Activity in Psoriatic Epidermis. J. Investig. Dermatol. 1989, 92, 791–797. [Google Scholar] [CrossRef]
- Roller, A.; Perino, A.; Dapavo, P.; Soro, E.; Okkenhaug, K.; Hirsch, E.; Ji, H. Blockade of Phosphatidylinositol 3-Kinase (PI3K)δ or PI3Kγ Reduces IL-17 and Ameliorates Imiquimod-Induced Psoriasis-like Dermatitis. J. Immunol. 2012, 189, 4612–4620. [Google Scholar] [CrossRef]
- DeLeo, V.; Scheide, S.; Meshulam, J.; Hanson, D.; Cardullo, A. Ultraviolet Radiation Alters Choline Phospholipid Metabolism in Human Keratinocytes. J. Investig. Dermatol. 1988, 91, 303–308. [Google Scholar] [CrossRef]
- Gruber, F. The Skin Lipidome Under Environmental Stress—Technological Platforms, Molecular Pathways and Translational Opportunities. In Skin Stress Response Pathways; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2016; pp. 1–27. [Google Scholar]
- Dalmau, N.; Andrieu-Abadie, N.; Tauler, R.; Bedia, C. Phenotypic and lipidomic characterization of primary human epidermal keratinocytes exposed to simulated solar UV radiation. J. Dermatol. Sci. 2018, 92, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Olivier, E.; Dutot, M.; Regazzetti, A.; Dargere, D.; Auzeil, N.; Laprevote, O.; Rat, P. Lipid deregulation in UV irradiated skin cells: Role of 25-hydroxycholesterol in keratinocyte differentiation during photoaging. J. Steroid Biochem. Mol. Boil. 2017, 169, 189–197. [Google Scholar] [CrossRef]
- Chakravarthy, M.V.; Lodhi, I.J.; Yin, L.; Malapaka, R.R.V.; Xu, H.E.; Turk, J.; Semenkovich, C.F. Identification of a Physiologically Relevant Endogenous Ligand for PPARα in Liver. Cell 2009, 138, 476–488. [Google Scholar] [CrossRef] [PubMed]
- Straus, D.S.; Glass, C.K. Anti-inflammatory actions of PPAR ligands: New insights on cellular and molecular mechanisms. Trends Immunol. 2007, 28, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Rockenfeller, P.; Carmona-Gutierrez, D.; Pietrocola, F.; Kroemer, G.; Madeo, F. Ethanolamine: A novel anti-aging agent. Mol. Cell. Oncol. 2015, 3, e1019023. [Google Scholar] [CrossRef] [PubMed]
- Scheau, C.; Badarau, I.A.; Mihai, G.L.; Scheau, A.-E.; Costache, D.O.; Constantin, C.; Calina, D.; Caruntu, C.; Costache, R.S.; Caruntu, A. Cannabinoids in the Pathophysiology of Skin Inflammation. Molecules 2020, 25, 652. [Google Scholar] [CrossRef]
- Dean, J.M.; Lodhi, I.J. Structural and functional roles of ether lipids. Protein Cell 2017, 9, 196–206. [Google Scholar] [CrossRef]
- McIntyre, T.M. Bioactive oxidatively truncated phospholipids in inflammation and apoptosis: Formation, targets, and inactivation. Biochim. Biophys. Acta (BBA) Bioenerg. 2012, 1818, 2456–2464. [Google Scholar] [CrossRef]
- Broniec, A.; Klosinski, R.; Pawlak, A.M.; Wrona-Krol, M.; Thompson, D.; Sarna, T.J. Interactions of plasmalogens and their diacyl analogs with singlet oxygen in selected model systems. Free. Radic. Boil. Med. 2011, 50, 892–898. [Google Scholar] [CrossRef]
- Wallner, S.; Schmitz, G. Plasmalogens the neglected regulatory and scavenging lipid species. Chem. Phys. Lipids 2011, 164, 573–589. [Google Scholar] [CrossRef]
- Dobrzyńska, I.; Kotynska, J.; Szachowicz-Petelska, B.; Figaszewski, Z.A. Determination of association constants of monovalent ions to sphingomyelin and phosphatidylinositol liposomal membranes by microelectrophoresis. Soft Mater. 2016, 15, 113–120. [Google Scholar] [CrossRef]
- Kotynska, J.; Dobrzyńska, I.; Figaszewski, Z.A. Association of alkali metal cations with phosphatidylcholine liposomal membrane surface. Eur. Biophys. J. 2016, 46, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, B.; Laurino, C.; Vadalà, M. A therapeutic effect of cbd-enriched ointment in inflammatory skin diseases and cutaneous scars. La Clin. Ter. 2019, 170, e93–e99. [Google Scholar]
- Fotakis, G.; Timbrell, J.A. In vitro cytotoxicity assays: Comparison of LDH, neutral red, MTT and protein assay in hepatoma cell lines following exposure to cadmium chloride. Toxicol. Lett. 2006, 160, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Casares, L.; García, V.; Garrido-Rodríguez, M.; Millán, E.; Collado, J.A.; García-Martín, A.; Peñarando, J.; Calzado, M.A.; De La Vega, L.; Muñoz, E. Cannabidiol induces antioxidant pathways in keratinocytes by targeting BACH1. Redox Boil. 2020, 28, 101321. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Bartlett, E.; Lewis, D. Spectrophotometric determination of phosphate esters in the presence and absence of orthophosphate. Anal. Biochem. 1970, 36, 159–167. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [PubMed]
- Lé, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Dinno, A. Dunn. Test: Dunn’s Test of Multiple Comparisons Using Rank Sums. R Package Version 1.3.5. 2017. Available online: https://cran.r-project.org/web/packages/dunn.test/index.html (accessed on 4 January 2019).
- Kolde, R. Pheatmap: Pretty Heatmaps. R Package Version 1.0.12. 2019. Available online: https://CRAN.R-project.org/package=pheatmap (accessed on 4 January 2019).
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.gbif.org/tool/81287/r-a-language-and-environment-for-statistical-computing (accessed on 4 January 2019).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Łuczaj, W.; Dobrzyńska, I.; Wroński, A.; Domingues, M.R.; Domingues, P.; Skrzydlewska, E. Cannabidiol-Mediated Changes to the Phospholipid Profile of UVB-Irradiated Keratinocytes from Psoriatic Patients. Int. J. Mol. Sci. 2020, 21, 6592. https://doi.org/10.3390/ijms21186592
Łuczaj W, Dobrzyńska I, Wroński A, Domingues MR, Domingues P, Skrzydlewska E. Cannabidiol-Mediated Changes to the Phospholipid Profile of UVB-Irradiated Keratinocytes from Psoriatic Patients. International Journal of Molecular Sciences. 2020; 21(18):6592. https://doi.org/10.3390/ijms21186592
Chicago/Turabian StyleŁuczaj, Wojciech, Izabela Dobrzyńska, Adam Wroński, M Rosário Domingues, Pedro Domingues, and Elżbieta Skrzydlewska. 2020. "Cannabidiol-Mediated Changes to the Phospholipid Profile of UVB-Irradiated Keratinocytes from Psoriatic Patients" International Journal of Molecular Sciences 21, no. 18: 6592. https://doi.org/10.3390/ijms21186592
APA StyleŁuczaj, W., Dobrzyńska, I., Wroński, A., Domingues, M. R., Domingues, P., & Skrzydlewska, E. (2020). Cannabidiol-Mediated Changes to the Phospholipid Profile of UVB-Irradiated Keratinocytes from Psoriatic Patients. International Journal of Molecular Sciences, 21(18), 6592. https://doi.org/10.3390/ijms21186592