Effects of Gangliosides on Spermatozoa, Oocytes, and Preimplantation Embryos

 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Gangliosides
3. Protective Effect of Gangliosides in Germ Cells and Early Embryos
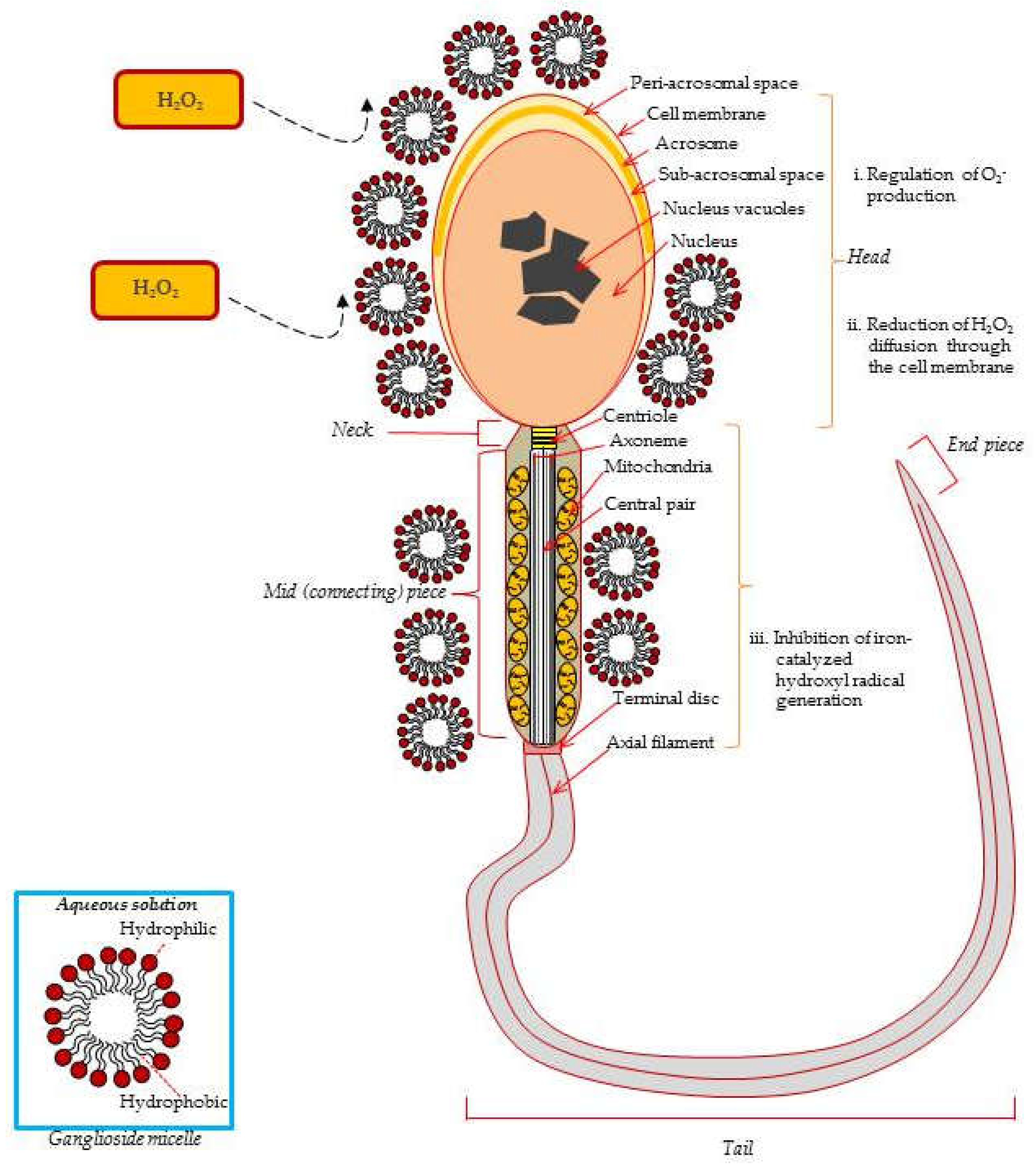
4. Effects of Gangliosides on Spermatozoa.
5. Gangliosides on Oocytes and Preimplantation Embryos
6. Conclusions
Funding
Conflicts of Interest
References
- Edidin, M. The state of lipid rafts: From model membranes to cell. Ann. Rev. Biophys. Struct. 2003, 32, 257–283. [Google Scholar] [CrossRef] [PubMed]
- Brown, D. Lipid rafts, detergent-resistant membranes, and raft targeting signals. Physiology 2006, 21, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Mclntosh, T.J. Overview of membrane rafts. Meth. Mol. Biol. 2007, 398, 1–9. [Google Scholar]
- Simons, K.; Ikonen, E. Functional rafts in cell membranes. Nature 1997, 387, 569–572. [Google Scholar] [CrossRef]
- Silvius, J.R. Partitioning of membrane molecules between raft and non-raft domains: Insights from membrane studies. Biochem. Biophys. Acta. 2005, 1746, 193–202. [Google Scholar] [CrossRef]
- Campbell, T.; Davy, A.; Liu, Y.; Arcellana-Panlilio, M.; Robbins, S. Distinct membrane compartmentalization and signaling of ephrin-A5 and ephrin-B1. Biochem. Biophys. Res. Commun. 2008, 375, 362–366. [Google Scholar] [CrossRef]
- Godoy, V.; Riquelme, G. Distinct lipid rafts in subdomains form human placental apical syncytiotrophoblast membranes. J. Membr. Biol. 2008, 224, 21–31. [Google Scholar] [CrossRef]
- Sonnino, S.; Mauri, L.; Chigorno, V.; Prinetti, A. Gangliosides as components of lipid membrane domains. Glycobiology 2007, 17, 1R–13R. [Google Scholar] [CrossRef]
- Kotani, M.; Kawashima, I.; Ozawa, H.; Terashima, T.; Tai, T. Differential distribution of major gangliosides in rat central nervous system detected by specific monoclonal antibodies. Glycobiology 1993, 3, 137–146. [Google Scholar] [CrossRef]
- Hakomori, S. Structure, organization and function of glycosphingolipids in membrane. Cur. Opin. Hematol. 2003, 10, 16–24. [Google Scholar] [CrossRef]
- Biswas, S.; Biswas, K.; Richmond, A.; Ko, J.; Chosh, S.; Simmons, M.; Rayman, P.; Rimi, B.; Gill, I.; Tannenbaum, C.S.; et al. Elevated levels of select gangliosides in T cells from renal cell carcinoma patients us associated with T cell dysfunction. J. Immunol. 2009, 183, 5050–5058. [Google Scholar] [CrossRef] [PubMed]
- Webb, T.J.; Li, X.; Giuntoli, R.L.; Lopez, P.H.; Heuser, C.; Schnaar, R.L.; Tsuji, M.; Kurts, C.; Oelke, M.; Schneck, J.P. Molecular identification of GD3 as a suppressor of the innate immune response in ovarian cancer. Cancer Res. 2012, 72, 3744–3752. [Google Scholar] [CrossRef] [PubMed]
- Sa, G.T.; Das, T.; Moon, C.; Hilston, C.M.; Rayman, P.A.; Rini, B.I.; Tannenbaum, C.S.; Finke, J.H. GD3, an overexpressed tumor-derived ganglioside, mediates the apoptosis of activated but not resting T cells. Cancer Res. 2009, 69, 3095–3104. [Google Scholar] [CrossRef] [PubMed]
- Dennery, P.A. Effects of oxidative stress on embryonic development. Birth defects Res. C. Embryo Today Rev. 2007, 81, 155–162. [Google Scholar] [CrossRef]
- Tripathi, A.; Khatun, S.; Pandey, A.N.; Mishra, S.K.; Chaube, R.; Shrivastav, T.G.; Chaube, S.K. Intracellular levels of hydrogen peroxide and nitric oxide in oocytes at various stages of meiotic cell cycle and apoptosis. Free Radic. Res. 2009, 43, 287–294. [Google Scholar] [CrossRef]
- Maulik, N.; Das, D.K.; Gogineni, M.; Cordis, G.A.; Avrova, N.; Denosova, N. Reduction of myocardial ischemic reperfusion injury by sialylated glycosphingolipids, gangliosides. J. Cardiovasc. Pharmacol. 1993, 22, 74–81. [Google Scholar] [CrossRef]
- Bondy, S.C.; Mckee, M.; Davoodbhoy, Y.M. Prevention of chemically induced changes in synaptosomal membrane order by ganglioside GM1 and α-tocooherol. Biochim. Biophys. Acta. 1990, 1026, 213–219. [Google Scholar] [CrossRef]
- Tyurin, V.A.; Tyurina, Y.Y.; Avrova, N.F. Ganglioside-dependent factor, inhibiting lipid peroxidation in rat brain synaptosomes. Neurochem. Int. 1992, 20, 401–407. [Google Scholar] [CrossRef]
- Tyurina, Y.Y.; Tyurin, V.A.; Avrova, N.F. Ganglioside GM1 protects cAMP 3`5`: Phosphodiesterase from inactivation caused by lipid peroxidation in brain synaptosomes of rats. Mol. Chem. Neuropathol. 1993, 19, 205–217. [Google Scholar] [CrossRef]
- Avrova, N.F.; Zakharova, I.O.; Tyurin, V.A.; Tyurina, Y.Y.; Gamaley, I.A.; Schepetkin, I.A. Different metabolic effects of ganglioside GM1 in brain synaptosome and phagocytic cells. Neurochem. Res. 2002, 27, 27,751–759. [Google Scholar] [CrossRef]
- Yamamoto, H.A.; Mohanan, P.V. Ganglioside GT1b and melatonin inhibit brain mitochondrial DNA damage and seizures induced by kainic acid and mice. Brain Res. 2003, 21, 100–106. [Google Scholar] [CrossRef]
- Gong, G.; Yin, L.; Yuan, L.; Sui, D.; Sun, Y.; Fu, H.; Chen, L.; Wang, X. Ganglioside GM1 protects against high altitude cerebral edema in rats by suppressing the oxidative stress and inflammatory response via the PI3K/AKT-Nrf2 pathway. Mol. Immunol. 2018, 95, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Gavella, M.; Kveder, M.; Lipovac, V. Modulation of ROS production in human leukocytes by ganglioside micelles. Braz. J. Med. Biol. Res. 2010, 43, 942–949. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kim, S.M.; Jung, J.U.; Ryu, J.S.; Jin, J.W.; Yang, H.J.; Ko, K.; You, H.K.; Jung, K.Y.; Choo, Y.K. Effects of gangliosides on the differentiation of human mesenchymal stem cells into osteoblasts by modulation epidermal growth factor receptors. Biochem. Biophys. Res. Commun. 2008, 371, 866–871. [Google Scholar] [CrossRef]
- Xu, Y.; Tan, L.J.; Grachtchouk, V.; Voorhees, J.J.; Fisher, G.J. Receptor-type protein-tysosine phosphatase-kappa regulated epidermal growth factors receptor function. J. Biol. Chem. 2005, 280, 42694–42700. [Google Scholar] [CrossRef]
- Ben-Yosef, D.; Galiani, D.; Dekel, N.; Shalgi, R. Rat oocytes induced to mature by epidermal growth factor are successfully fertilized. Mol. Cell Endocrionol. 1992, 88, 135–141. [Google Scholar] [CrossRef]
- De La Fuente, R.; O.’Brien, M.J.; Eppig, J.J. Epidermal growth factor enhances preimplantation developmental competence of maturing mouse oocytes. Hum. Reprod. 1999, 14, 3060–3068. [Google Scholar] [CrossRef]
- Illera, M.J.; Lorenzo, P.L.; Illera, J.C.; Petters, R.M. Developmental competence of immature pig oocytes under the influence of EGF, IGF-I, follicular fluid and gonadotropins during IVM-IVF processes. Int. J. Biol. 1998, 42, 1169–1172. [Google Scholar]
- Singh, B.; Meng, L.; Rutledge, J.M.; Armstrongs, D.T. Effects of epidermal growth factor and follicle-stimulating hormone during in vitro maturation on cytoplasmic maturation of porcine oocytes. Mol. Reprod. Dev. 1997, 46, 401–407. [Google Scholar] [CrossRef]
- Goud, P.T.; Goud, A.P.; Quan, C.; Laverge, H.; Van der Elst, J.; De Sutter, P.; Dhont, M. In-vitro maturation of human germinal vesicle stage oocytes role of cumulus cells and epidermal growth factor in the culture medium. Hum. Reprod. 1998, 13, 1638–1644. [Google Scholar] [CrossRef]
- Fujimoto, Y.; Izumoto, S.; Suzuki, T.; Kinoshita, M.; Kagawa, N.; Wada, K.; Hashimoto, N.; Maruno, M.; Nakatsuji, Y.; Yoshimine, T. Ganglioside GM3 inhibits proliferation and invasion of glioma. J. Neuroonco. 2005, 71, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Chung, T.W.; Kim, S.J.; Choi, H.J.; Kim, K.J.; Kim, M.J.; Kim, S.H.; Lee, H.J.; Ko, J.H.; Lee, Y.C.; Suzuki, A.; et al. Ganglioside GM3 inhibits VEGF/VEGFR-2-mediated angiogenesis: Direct interaction of GM3 with VEGFR-2. Glycobiology 2009, 19, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Meuillet, E.J.; Mania-Farnell, B.; George, D.; Inokuchi, J.I.; Bremer, E.G. Modulation of EGF receptor activity by changes in the GM3 content in a human epidermoid carcinoma cell line, A431. Exp. Cell Res. 2000, 256, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, R.; Ohyama, C.; Aoki, H.; Takahashi, T.; Satoh, M.; Saito, S.; Hoshi, S.; Ishii, A.; Saito, M.; Arai, Y. Ganglioside GM3 overexpression induces apoptosis and reduces malignant potential in murine bladder cancer. Cancer Res. 2002, 62, 3850–3854. [Google Scholar]
- Huang, X.; Li, Y.; Zhang, J.; Xu, Y.; Tian, Y.; Ma, K. Ganglioside GM3 inhibits hepatoma cell motility via down-regulation activity of EGFR and PI3K/AKT signaling pathway. J. cell. Biocehm. 2013, 114, 1616–1624. [Google Scholar]
- Park, H.J.; Chae, S.K.; Kim, J.W.; Yang, S.G.; Jung, J.M.; Kim, M.J.; Wee, G.; Lee, D.S.; Kim, S.U.; Koo, D.B. Ganglioside GM3 induces cumulus apoptosis through inhibition of epidermal growth factors receptor-mediated PI3K/AKT signaling pathways during in vitro maturation of pig oocytes. Mol. Reprod. Dev. 2017, 84, 702–711. [Google Scholar] [CrossRef]
- Liu, Y.; Wond, A.; Bob, D.; Weiss, M.; Sliwinski, R.; Vilar, J.; Notario, V.; Sutherland, M.; Colberg-poley, A.M.; Ladisch, S. Ganglioside synthase knockout in oncogene-transformed fibroblasts depletes gangliosides and impairs tumor growth. Oncogen 2010, 29, 3297–3306. [Google Scholar] [CrossRef]
- Saito, M.; Ito, M.; Sugiyama, K. A specific loss of C-series gangliosides in pancreas of streptozotocin-induced diabetic rats. Life Sci. 1999, 64, 1803–1810. [Google Scholar] [CrossRef]
- Saito, M.; Sugiyama, K. A distinct ganglioside composition of rat pancreatic islets. Arch. Biochem. Biophys. 2000, 376, 371–376. [Google Scholar] [CrossRef]
- Van Blerkom, J.; Caltrider, K. Sperm attachment and penetration competence in the human oocyte: A possible aetiology of fertilization failure involving the organization of oolemmal lipid raft microdomains influenced by the ΔΨm of subplasmalemmal mitochondria. Reprod. Biomed. Online 2013, 27, 27,690–701. [Google Scholar] [CrossRef]
- Kolter, T.; Sandhoff, K. Recent advances in the biochemistry of sphingolipidoses. Brain Pathol. 1998, 79–100. [Google Scholar] [CrossRef] [PubMed]
- Jeckel, D.; Karrenbauer, A.; Burger, K.N.; van Meer, G.; Wieland, F. Glucosylceramide is synthesized at the cytosolic surface of various Golgi subfractions. J. cell. Biol. 1992, 117, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Futerman, A.H.; Pagano, R.E. Determination of the intracellular sites and topology of glucosylceramide synthesis in rat liver. Biochem. J. 1991, 280, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Marks, D.L.; Wu, K.; Paul, P.; Kamisaka, Y.; Watanabe, R.; Pagano, R.E. Oligomerization and topology of the Golgi membrane protein glucosylceramide synthase. J. Biol. Chem. 1999, 274, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, T.; Wada, R.A.; Sasaki, T.; Deng, C.; Bierfreund, U.; Sandhoff, K.; Proia, R.L. A vital role for glycosphingolipid synthesis during development and differentiation. Proc. Natl. Acad. Sci. USA 1999, 96, 9142–9147. [Google Scholar] [CrossRef] [PubMed]
- Kolesnick, R.N.; Kronke, M. Regulation of ceramide production and apoptosis. Annu. Rev. Physiol. 1998, 60, 643–665. [Google Scholar] [CrossRef]
- Hannun, Y.A.; Obeid, L.M. Ceramide and the eukaryotic stress response. Biochem. Soc. Trans. 1997, 25, 1171–1175. [Google Scholar] [CrossRef]
- Schauer, R. Sialic acids as link to Japanese scientists. Proc. Jpn. Acad. Ser. B. Phys. Biol. Sci. 2016, 92, 109–120. [Google Scholar] [CrossRef]
- Tanner, M.E. The enzymes of sialic acid biosynthesis. Bioorg. Chem. 2005, 33, 216–228. [Google Scholar] [CrossRef]
- YU, R.K.; Tsai, Y.; Ariga, T.; Yanagisawa, M. Structures, biosynthesis, and functions of gangliosides- an overview. J.Oleo.Sci. 2011, 60, 537–544. [Google Scholar] [CrossRef]
- Kuhn, R.; Wiegandt, H. Die Konstitution der Ganglio-N-Tetraose und des gangliosids G1. Chember. 1963, 96, 866. [Google Scholar]
- Mahadik, S.P.; Karpiak, S.K. Gangliosides in treatment of neural injury and disease. Drug Dev. Res. 1998, 15, 337–360. [Google Scholar] [CrossRef]
- Inokuchi, J. Membrane microdomains and insulin resistance. FEBS. Lett. 2010, 584, 1864–1871. [Google Scholar] [CrossRef] [PubMed]
- Regina Todeschini, A.; Hakomori, S.I. Functional role of glycosphingolipids and gangliosides in control of cell adhesion, motility, and growth, through glycosynaptic microdomains. Biochim. Biophys. Acta. 2008, 1780, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Ohmi, Y.; Ohkawa, Y.; Yamauchi, Y.; Tajima, O.; Furukawa, K.; Furukawa, K. Essential roles of gangliosides in the formation and maintenance of membrane microdomains in brain tissues. Neurochem. Res. 2012, 37, 1185–1191. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, K.; Hamamura, K.; Ohkawa, Y.; Ohmi, Y.; Furukawa, K. Disialyl gangliosides enhance tumor phenotypes with differential modalities. Glycoconj. J. 2012, 29, 579–584. [Google Scholar] [CrossRef]
- Hakomori, S.I.; Handa, K. GM3 and cancer. Glycoconj. J. 2015, 32, 1–8. [Google Scholar] [CrossRef]
- Ledeen, R.W.; Wu, G.; Andre, S.; Bleich, D.; Huet, D.; Kaltner, H.; Koptiz, J.; Gablus, H.J. Beyond glycoproteins as galectin counterreceptors:tumor-effector T cell growth control via ganglioside GM1 [corrected]. Ann. N. Y. Acad. Sci. 2012, 1253, 206–221. [Google Scholar] [CrossRef]
- Lipina, C.; Hundal, H.S. Ganglioside GM3 as a gatekeeper of obesity-associated insulin resistance: Evidence and mechanisms. Febs. Lett. 2015, 589, 3221–3227. [Google Scholar] [CrossRef]
- Allende, M.L.; Proia, R.L. Simplifying complexity: Genetically resculpting glycosphingolipid synthesis pathways in mice to reveal function. Glycoconj. J. 2014, 31, 613–622. [Google Scholar] [CrossRef]
- Avrova, N.F.; Victorov, I.V.; Tyurin, V.A.; Zakharova, I.O.; Sokolova, T.V.; Andreeva, N.A.; Stelmaschuk, E.V.; Tyurina, Y.Y.; Gonchar, V.S. Inhibition of glutamate-induced intensification of free radical reactions by ganglioside: Possible role in their protective effect in rat cerebella granule cells and phagocytic cell. Neurochem. Res. 1998, 23, 945–952. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.U.; Jeon, Y.; Youn, J.D.; Cai, L.; Kim, E.; Yoo, H.; Kim, K.J.; Park, K.M.; Jin, M.; Kim, H.; et al. Effect of ganglioside GT1b on the in vitro maturation of porcine oocytes and embryonic development. J. Reprod. Dec. 2015, 61, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Tamburino, L.; Marchiani, S.; Montoya, M.; Elia Marino, F.; Natail, I.; Cambi, M.; Forti, D.; Baldi, E.; Muratori, M. Mechanisms and clinical correlates of sperm DNA damage. Asianj. Androl. 2012, 14, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Thomson, L.K.; Fleming, S.D.; Aitken, R.J.; De, G.N.; Zieschang, J.A.; Clark, A.M. Cryopreservation-induced human sperm DNA damage is predominantly mediated by oxidative stress rather than apoptosis. Hum. Reprod. 2009, 24, 2061–2070. [Google Scholar] [CrossRef]
- Zribi, N.; Chakroun, N.K.; El Euch, H.; Gargouri, J.; Bahloul, A.; Ammar Keskes, L. Effects of cryopreservation on human sperm deoxyribonucleic acid integrity. Fertil. Steril. 2010, 93, 159–166. [Google Scholar] [CrossRef]
- Kim, B.H.; Jung, J.U.; Ko, K.; Kim, W.S.; Ryu, J.S.; Jun, J.W.; Yang, H.J.; Kim, J.S.; Kwon, H.C.; Nam, S.Y.; et al. Expression of ganglioside GT1b in mouse embryos at different development stages after cryopreservation. Arch. Pham. Res. 2008, 31, 88–95. [Google Scholar] [CrossRef]
- James, P.S.; Wolfe, C.A.; Mackie, A.; Ladha, S.; Prentice, A.; Jones, R. Lipid dynamics in the plasma membrane of fresh and cryopreserved human spermatozoa. Hum. Reprod. 1999, 14, 1827–1832. [Google Scholar] [CrossRef]
- Chakrabarty, J.; Banerjee, D.; Pal, J.; De, J.; Ghosh, A.; Majumder, G.C. Shedding off specific lipid constituents from sperm cell membrane during cryopreservation. Cryobiology 2007, 54, 27–35. [Google Scholar] [CrossRef]
- Sautter, J.; Hoglinger, G.U.; Oertel, W.H.; Earl, C.D. Systemic treatment with GM1 ganglioside improves survival and function of cryopreserved embryonic midbrain grafted to the 6-hydroxydopamind-lesioned rat striatum. Exp. Neurol. 2000, 164, 121–129. [Google Scholar] [CrossRef]
- Gavella, M.; Lipova, V.; Rakos, R.; Colak, B. Reduction of oxidative changes in human spermatozoa by exogenous gangliosides. Andrologia 2005, 37, 17–24. [Google Scholar] [CrossRef]
- Jung, K.Y.; Kim, B.H.; Hwang, M.R.; Cho, J.R.; Kim, H.M.; Lee, Y.C.; Kim, C.H.; Kim, J.K.; Kim, B.J.; Choo, Y.K. Differential distribution of gnaglioside GM3 in seminiferous tubule and epididymis of adult rats. Arch. Pham. Res. 2001, 4, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Takamiya, K.; Yamamoto, A.; Furukawa, K.; Yamashiro, S.; Shin, M.; Okada, M.; Fukumoto, S.; Haraguchi, M.; Takeda, N.; Fujimura, K.; et al. Mice with disrupted GM2/GD2 synthase gene lack complex gangliosides but exhibit only subtle defects in their nervous system. Proc. Natl. Acad. Sci. USA 1996, 93, 10662–10667. [Google Scholar] [CrossRef] [PubMed]
- Takamiya, K.; Yamamoto, A.; Furukawa, K.; Zhao, J.; Fukumoto, S.; Yamashiro, S.; Okada, M.; Haraguchi, M.; Shin, M.; Kishikawa, M.; et al. Complex gangliosides are essential in spermatogenesis of mice: Possible roles in the transport of testosterone. Proc. Natl. Acad. Sci. USA 1998, 1998. 95, 12147–12152. [Google Scholar] [CrossRef]
- Aitken, R.J.; Gordon, E.; Harkiss, D.; Twigg, J.P.; Milne, P.; Jennings, Z.; Irvine, D.S. Relative impact of oxidative stress on the functional competence and genomic integrity of human spermatozoa. Biol. Reprod. 1998, 59, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Baldi, E.; Luconi, M.; Bonaccorsi, L.; Muratori, M.; Forti, G. Intracellular events and signaling pathways involved in sperm acquisition of fertilizing capacity and acrosome reaction. Front. Biosci. 2000, 5, E110–E123. [Google Scholar] [CrossRef]
- Visconti, P.E.; Bailey, J.L.; Moore, G.D.; Pan, D.; Olds-Clarke, P.; Kopf, G.S. Capacitation of mouse spermatozoa. I. Correction between the capacitation state and protein tyrosine Phosphorylation. Development 1995, 121, 1129–1137. [Google Scholar] [PubMed]
- Shadan, S.; James, P.S.; Howes, E.A.; Jones, R. Cholesterol efflux alters lipid raft stability and distribution during capacitation of boar spermatozoa. Biol. Reprod. 2004, 71, 253–265. [Google Scholar] [CrossRef]
- Roberts, K.P.; Wamstad, J.A.; Ensrud, K.M.; Hamilton, D.W. Inhibition of capacitation-associated tyrosine phosphorylation signaling in rat sperm by epididymal protein Crisp-1. Biol. Reprod. 2003, 69, 572–581. [Google Scholar] [CrossRef]
- Dunning, K.R.; Watson, L.N.; Zhang, V.J.; Brown, H.M.; Kaczmarek, A.K.; Robker, R.L.; Russell, D.L. Activation of Mouse Cumulus-Oocyte Complex Maturation In Vitro Through EGF-Like Activity of Versican. Biol. Reprod. 2015, 92, 116. [Google Scholar] [CrossRef]
- Selvaraj, V.; Buttke, D.E.; Asano, A.; McElwee, J.L.; Wolff, C.A.; Nelson, J.L.; Klaus, A.V.; Hunnicutt, G.R.; Travis, A.J. GM1 dynamics as a marker for membrane changes associated with the process of capacitation in murine and bovine spermatozoa. J. Androl. 2007, 28, 588–599. [Google Scholar] [CrossRef]
- Moody, M.A.; Cardona, C.; Simpson, A.J.; Smith, T.T.; Travis, A.J.; Ostermeier, G.C. Validation of a laboratory-developed test of human sperm capacitation. Mol. Reprod. Dev. 2017, 84, 408–422. [Google Scholar] [CrossRef] [PubMed]
- Choo, Y.K.; Chiba, T.C.; Tai, T.; Ogiso, M.; Hoshi, M. Differential distribution of gangliosides in adult rat ovary during the oestrous cycle. Glycobiology 1995, 5, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Choo, Y.K. Distribution of ganglioside GM3 in the rat ovary after gonadotropin stimulation. Mol. Cells 1999, 9, 365–375. [Google Scholar] [PubMed]
- Rebbaa, A.; Hurh, J.; Yamamoto, H.; Kersey, D.S.; Bremer, E.G. Ganglioside GM3 inhibition of EGF receptor mediated signal transduction. Glycobiology 1996, 6, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Chung, T.W.; Choi, H.J.; Kim, S.J.; Kwak, C.H.; Song, K.H.; Jun, U.H.; Chang, Y.C.; Chang, H.W.; Lee, Y.C.; Ha, K.T.; et al. The ganglioside GM3 in associated with cisplatin-induced apoptosis in human colon cancer cells. PLoS. ONE. 2014, 9, e92786. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, T.; Hashiramoto, A.; Haluzik, M.; Mizukami, H.; Beck, S.; Norton, A.; Kono, M.; Tsuji, S.; Daniotti, J.L.; Werth, N.; et al. Enhanced insulin sensitivity in mice lacking ganglioside GM3.Proc. Natl. Acad. Sci. USA 2003, 100, 3445–3459. [Google Scholar] [CrossRef] [PubMed]
- Ju, E.J.; Kwak, D.H.; Lee, D.H.; Kim, S.M.; Kim, J.S.; Choi, H.G.; Jung, K.Y.; Lee, S.U.; Do, S.I.; Park, Y.I.; et al. Pathophysiological implication of ganglioside GM3 in early mouse embryonic development through apoptosis. Arch. Pham. Res. 2005, 28, 1057–1064. [Google Scholar] [CrossRef]
- Kwak, D.H.; Jung, K.Y.; Lee, Y.C.; Choo, Y.K. Expressional changes of ganglioside GM3 during ovarian maturation and early embryonic development in db/db mice. Dev. Growth. Differ. 2003, 45, 95–102. [Google Scholar] [CrossRef]
- Zhuo, D.; Guan, F. Ganglioside GM1 promotes contact inhibition of growth by regulating the localization of epidermal growth factor receptor from glycospingolipid-enriched microdomain to caveolae. Cell Prolif. 2019, 52, e12639. [Google Scholar] [CrossRef]
- Mirkin, B.L.; Clark, S.H.; Zhang, C. Inhibition of human neuroblastoma cell proliferation and EGF receptor phosphorylation by gangliosides GM1, GM3, GD1A and GT1B. Cell Prolif. 2002, 35, 105–115. [Google Scholar] [CrossRef]
- Mitsuda, T.; Furukawa, K.; Fukumoto, S.; Miyazaki, H.; Urano, T.; Furukawa, K. Overexpression of ganglioside GM1 results in the dispersion of platelet-derived growth factor receptor from glycolipid-enriched microdomains and in the suppression of cell growth signals. J. Biol. Chem. 2002, 277, 11239–11246. [Google Scholar] [CrossRef] [PubMed]
- Nishio, M.; Fukumoto, S.; Furukawa, K.; Ichimura, A.; Miyazaki, H.; Kusunokis, S.; Urano, T.; Furukawa, K. Overexpressed GM1 suppresses nerve growth factor (NGF) signals by modulating the intracellular localization of NGF receptors and membrane fluidity in PC12 cells. J. Biol. Chem. 2004, 279, 33368–33378. [Google Scholar] [CrossRef] [PubMed]
- Comiskey, M.; Warner, C.M. Spatio-temporal localization of membrane lipid rafts in mouse oocytes and cleaving preimplantation embryos. Dev Biol. 2007, 303, 727–739. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sato, B.; Katagiri, Y.U.; Miyado, K.; Okino, N.; Ito, M.; Akutsu, H.; Okita, H.; Umezawa, A.; Fujimoto, J.; Toshimori, K.; et al. Lipid rafts enriched in monosialylGb5Cer carrying the stage-specific embryonic antigen-4 epitope are involved in development of mouse preimplantation embryos at cleavage stage. BMC. Dev. Biol. 2011, 11, 22. [Google Scholar] [CrossRef] [PubMed]
- Bouvier, J.D.; Seyfired, T.N. Ganglioside composition of normal mutant mouse embryos. J. Neurochem. 1989, 52, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Ngamukote, S.; Yanagisawa, M.; Ariga, T.; Ando, S.; Yu, R.K. Developmental changes of glycoshingolipids and expression of glycogens in mouse brains. J. Neurochem. 2007, 103, 2327–2341. [Google Scholar] [CrossRef]
- Kahl, C.R.; Means, A.R. Regulation of cell cycle progression by calcium/calmodulin dependent pathways. Endroc. Rev. 2003, 24, 719–736. [Google Scholar] [CrossRef]
- Hudmon, A.; Schulman, H. Neuronal CA2+/calmodulin-dependent protein kinase II: The role of structure and autoregulation in cellular function. Biochem. J. 2002, 364, 593–611. [Google Scholar] [CrossRef]
- Fan, H.Y.; Sun, Q.Y. Involvement of mitogen-activated protein kinase cascade during oocyte maturation and fertilization in mammals. Biol. Reprod. 2003, 69, 1552–1564. [Google Scholar] [CrossRef]
- Su, Y.Q.; Eppig, J.J. Evidence that multifunctional calcium/calmodulin-dependent protein kinase II (CaM KII) participates in the meiotic maturation of mouse oocytes. Mol. Reprod. Dev. 2002, 61, 560–569. [Google Scholar] [CrossRef]
- Chen, N.; Furuya, S.; Dol, H.; Hashimoto, Y.; Kudo, Y.; Higashi, H. Ganglioside/calmodulin kinaseII signal inducing cdc-42 mediated neuronal actin reorganization. Neuroscience 2003, 120, 163–176. [Google Scholar] [CrossRef]
- Kim, J.W.; Park, H.J.; Chae, S.k.; Ahn, J.H.; Do, G.Y.; Choo, Y.K.; Park, J.J.; Jung, B.D.; Kim, S.U.; Chang, K.T.; et al. Ganglioside GD1a promotes oocyte maturation, furthers preimplantation development, and increases blastocyst quality in pigs. J. Reprod. Dev. 2016, 62, 249–255. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Liu, Y.; Li, R.; Ladisch, S. Exogenous ganglioside GD1a enhances epidermal growth factor receptor binding and dimerization. J. Biol. Chem. 2004, 279, 36481–36489. [Google Scholar] [CrossRef] [PubMed]
- Miljan, E.A.; Meuillet, E.J.; Mania-Farnell, B.; George, D.; Yamamoto, H.; Simon, H.; Bremer, E.G. Interaction of the extracellular domain of the epidermal growth factor receptor with ganglioside. J. Biol. Chem. 2002, 277, 10108–10113. [Google Scholar] [CrossRef] [PubMed]
- Bremer, E.G.; Schlessinger, J.; Hakomori, S. Ganglioside-mediated modulation of cell growth. Specific effects of GM3 on tyrosine phosphorylation of the epidermal growth factor receptor. J.Biol.Chem. 1986, 261, 2434–2440. [Google Scholar]
- Vivanco, I.; Sawyers, C.L. The phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat. Rev. Cancer 2002, 2, 489–501. [Google Scholar] [CrossRef]
- Byeon, H.G.; Ku, M.; Yang, J. Beyond EGFR inhibition: Multilateral combat strategies to stop the progression of head and neck cancer. Exp. Mol. Med. 2019, 51, 8. [Google Scholar] [CrossRef]
- Tsai, S.C.; Lu, C.C.; Lee, C.Y.; Lin, Y.C.; Chung, J.G.; Kuo, S.C.; Amagaya, S.; Chen, F.N.; Chen, M.Y.; Chan, S.F. AKT serine/threonine protein kinase modulates bulfalin-triggered intrinsic pathway of apoptosis in CAL 27 oral cancer cells. Int. J. Oncol. 2012, 41, 1683–1692. [Google Scholar] [CrossRef][Green Version]
- Xiong, S.; Mu, T.; Wang, G.; Jiang, X. Mitochondria-mediated apoptosis in mammals. Protein Cell 2014, 5, 737–749. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, B.H.; Ju, W.S.; Kim, J.-S.; Kim, S.-U.; Park, S.J.; Ward, S.M.; Lyu, J.H.; Choo, Y.-K. Effects of Gangliosides on Spermatozoa, Oocytes, and Preimplantation Embryos. Int. J. Mol. Sci. 2020, 21, 106. https://doi.org/10.3390/ijms21010106
Kim BH, Ju WS, Kim J-S, Kim S-U, Park SJ, Ward SM, Lyu JH, Choo Y-K. Effects of Gangliosides on Spermatozoa, Oocytes, and Preimplantation Embryos. International Journal of Molecular Sciences. 2020; 21(1):106. https://doi.org/10.3390/ijms21010106
Chicago/Turabian StyleKim, Bo Hyun, Won Seok Ju, Ji-Su Kim, Sun-Uk Kim, Soon Ju Park, Sean M. Ward, Ju Hyeong Lyu, and Young-Kug Choo. 2020. "Effects of Gangliosides on Spermatozoa, Oocytes, and Preimplantation Embryos" International Journal of Molecular Sciences 21, no. 1: 106. https://doi.org/10.3390/ijms21010106
APA StyleKim, B. H., Ju, W. S., Kim, J.-S., Kim, S.-U., Park, S. J., Ward, S. M., Lyu, J. H., & Choo, Y.-K. (2020). Effects of Gangliosides on Spermatozoa, Oocytes, and Preimplantation Embryos. International Journal of Molecular Sciences, 21(1), 106. https://doi.org/10.3390/ijms21010106