Downregulation of the Netrin-1 Receptor UNC5b Underlies Increased Placental Angiogenesis in Human Gestational Diabetes Mellitus


Abstract
1. Introduction
2. Results
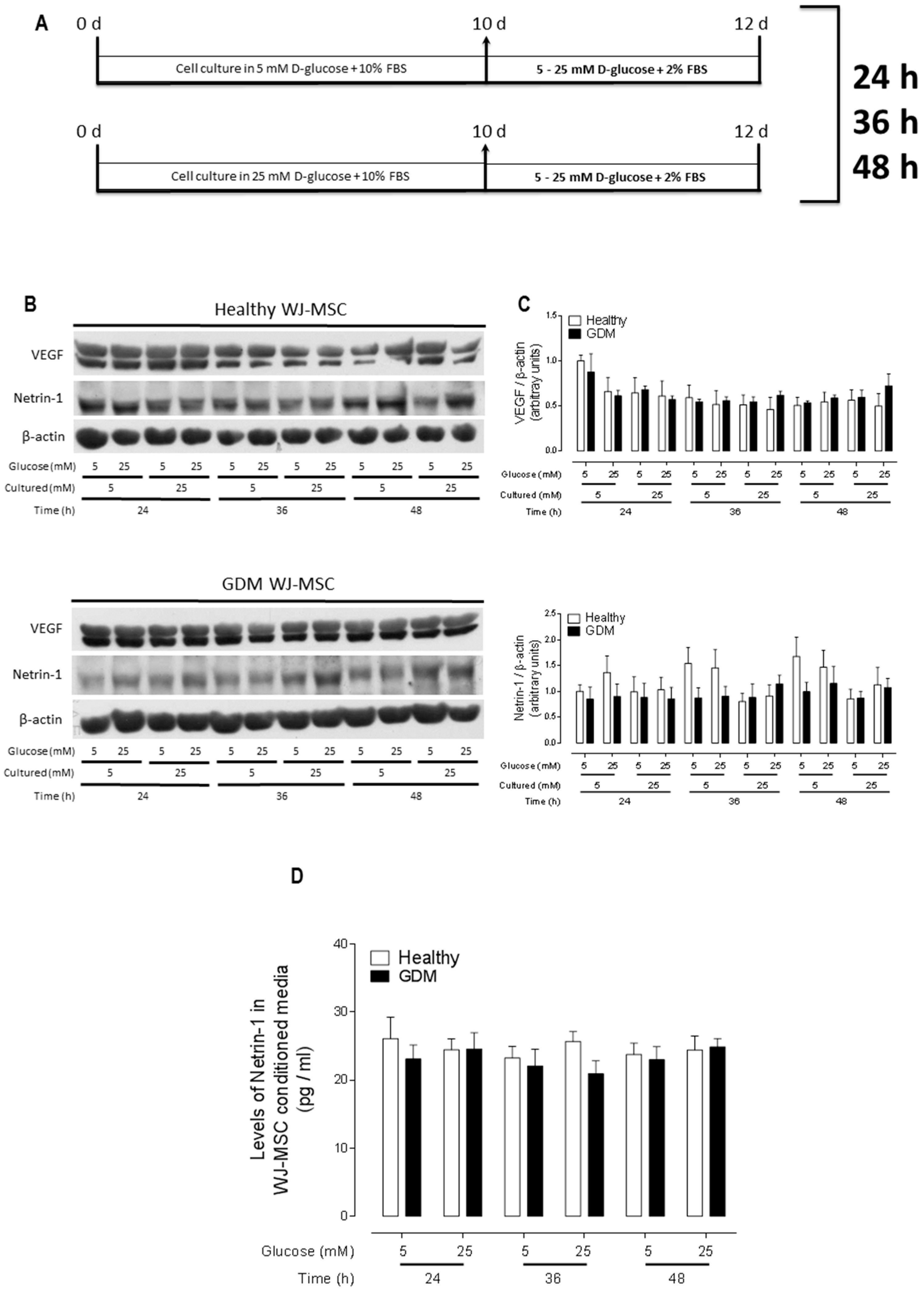
2.1. Healthy and GDM WJ-MSC Express and Secrete Similar Amounts of Netrin-1
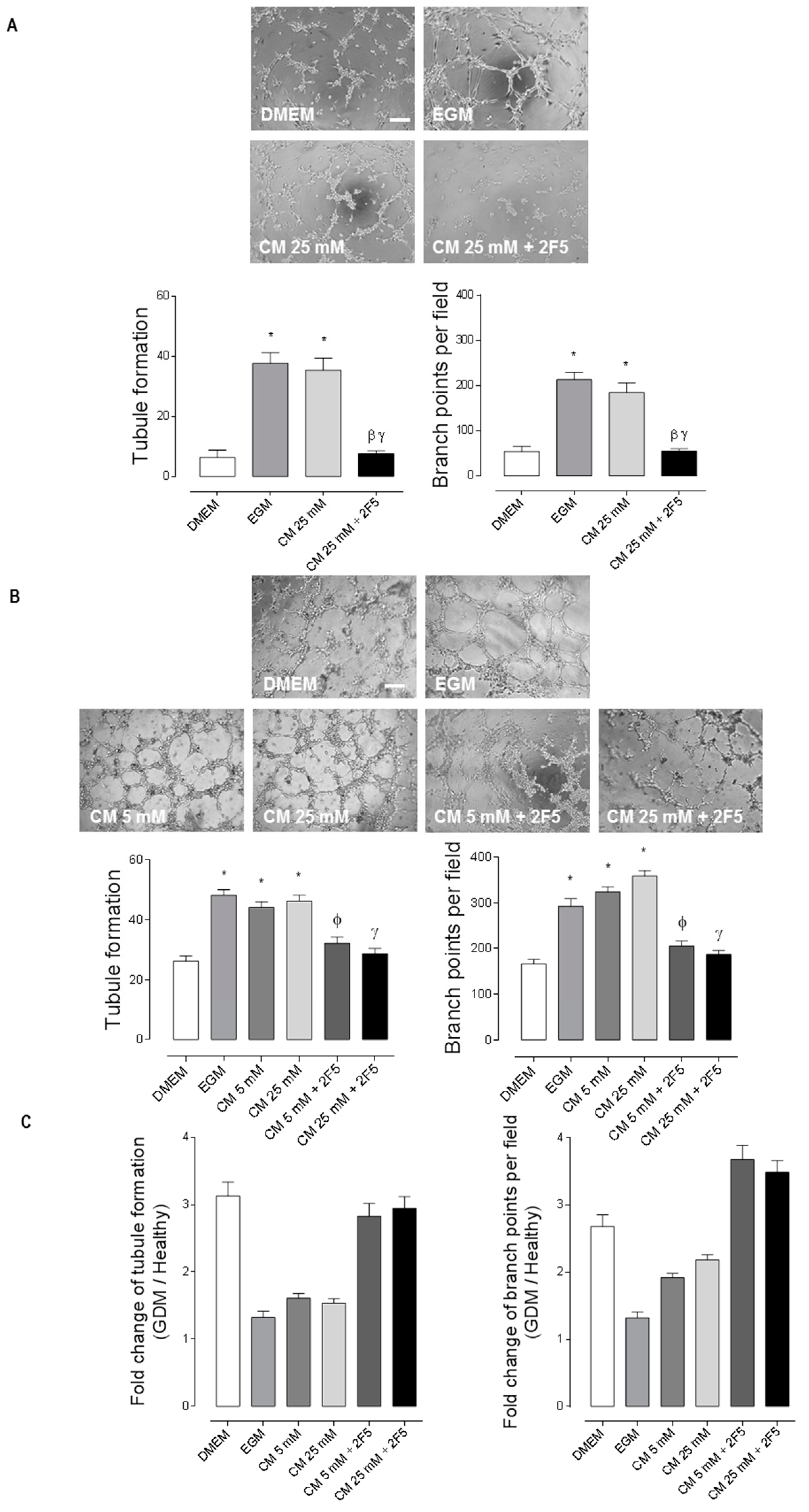
2.2. WJ-MSC’s Netrin-1 Has an Angiogenic Effect in GDM-HUVEC
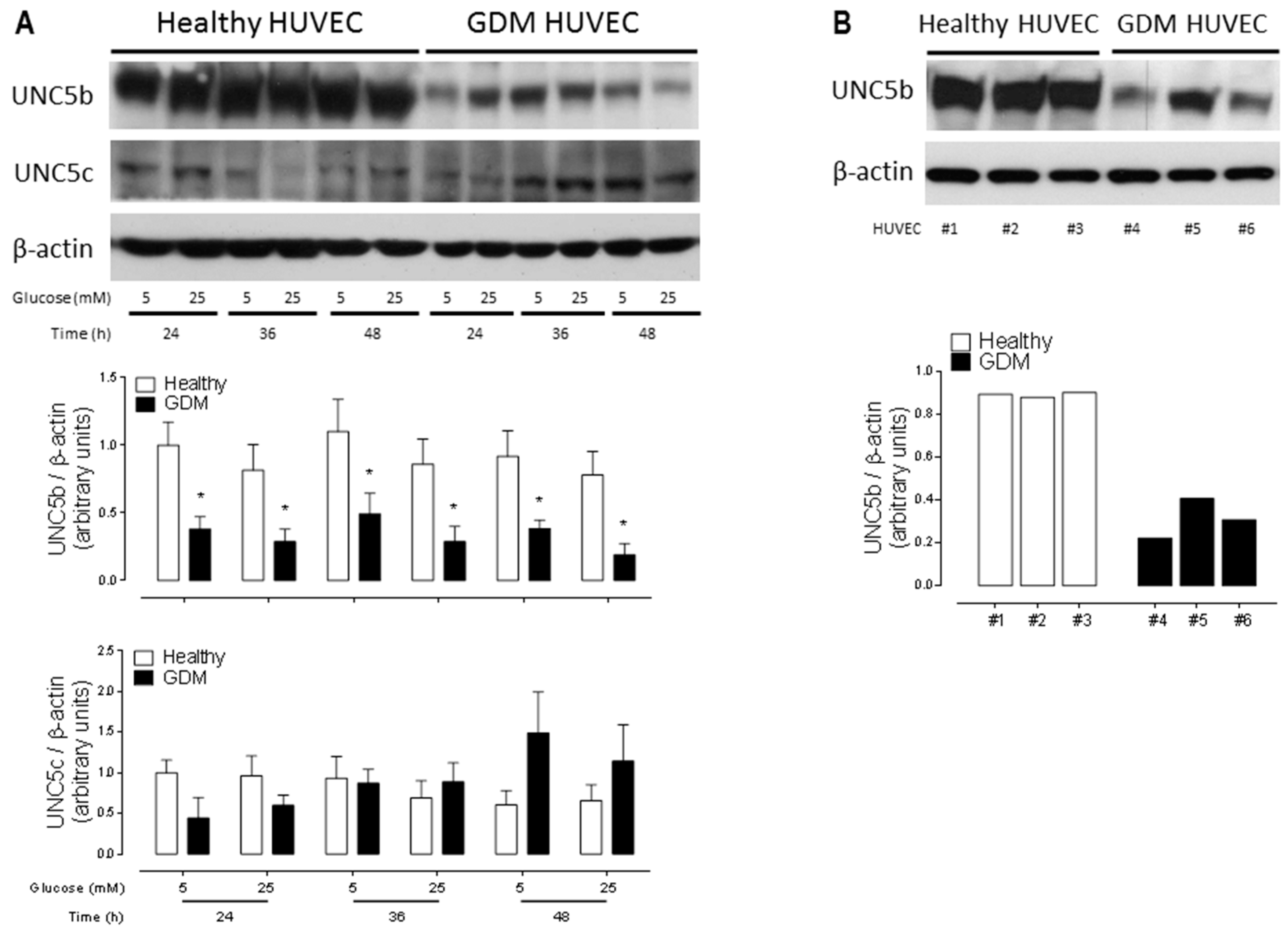
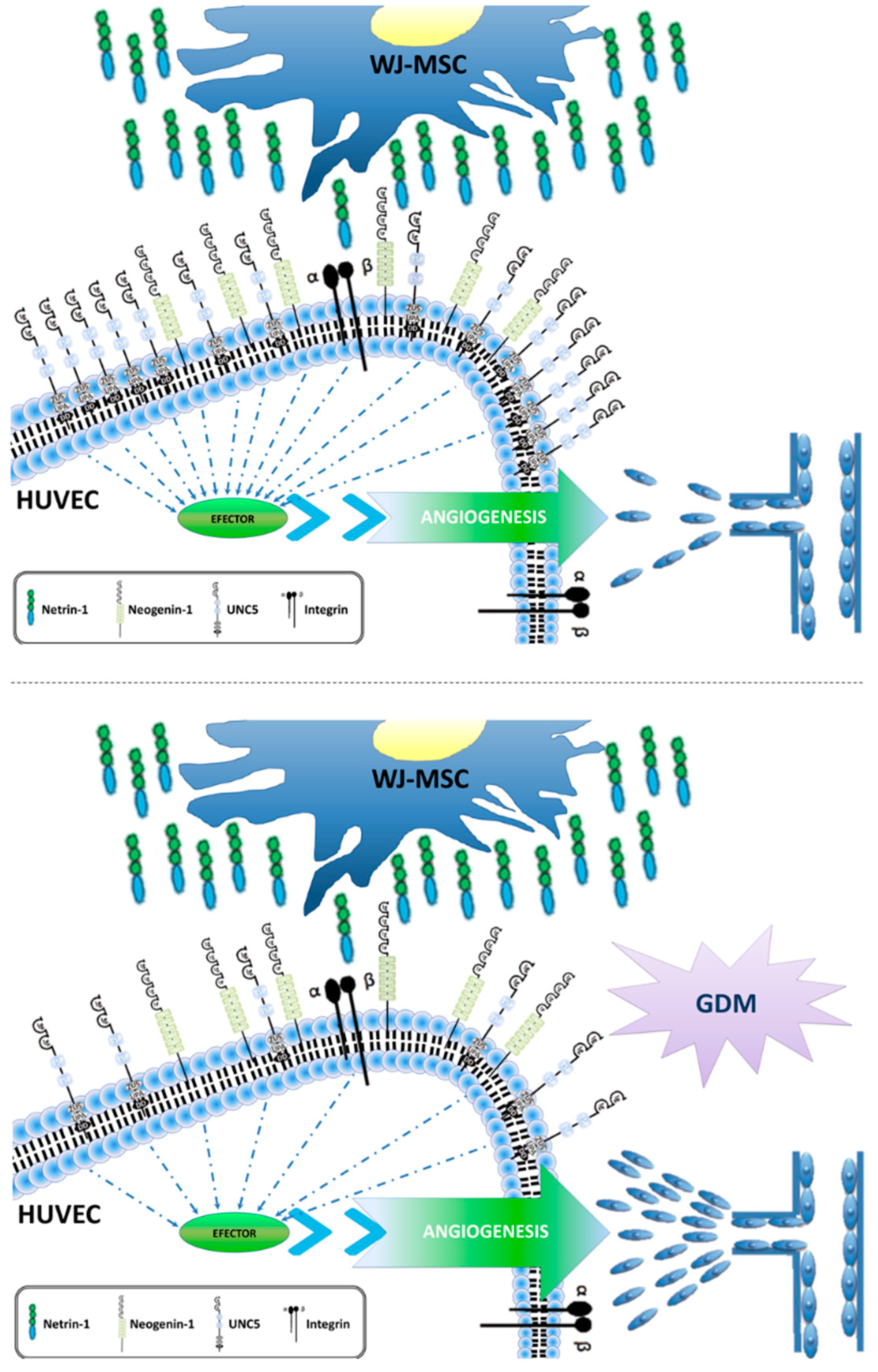
2.3. Downregulation of the Anti-Angiogenic UNC5b Receptor Could Explain the Increased Angiogenesis Observed in GDM HUVEC
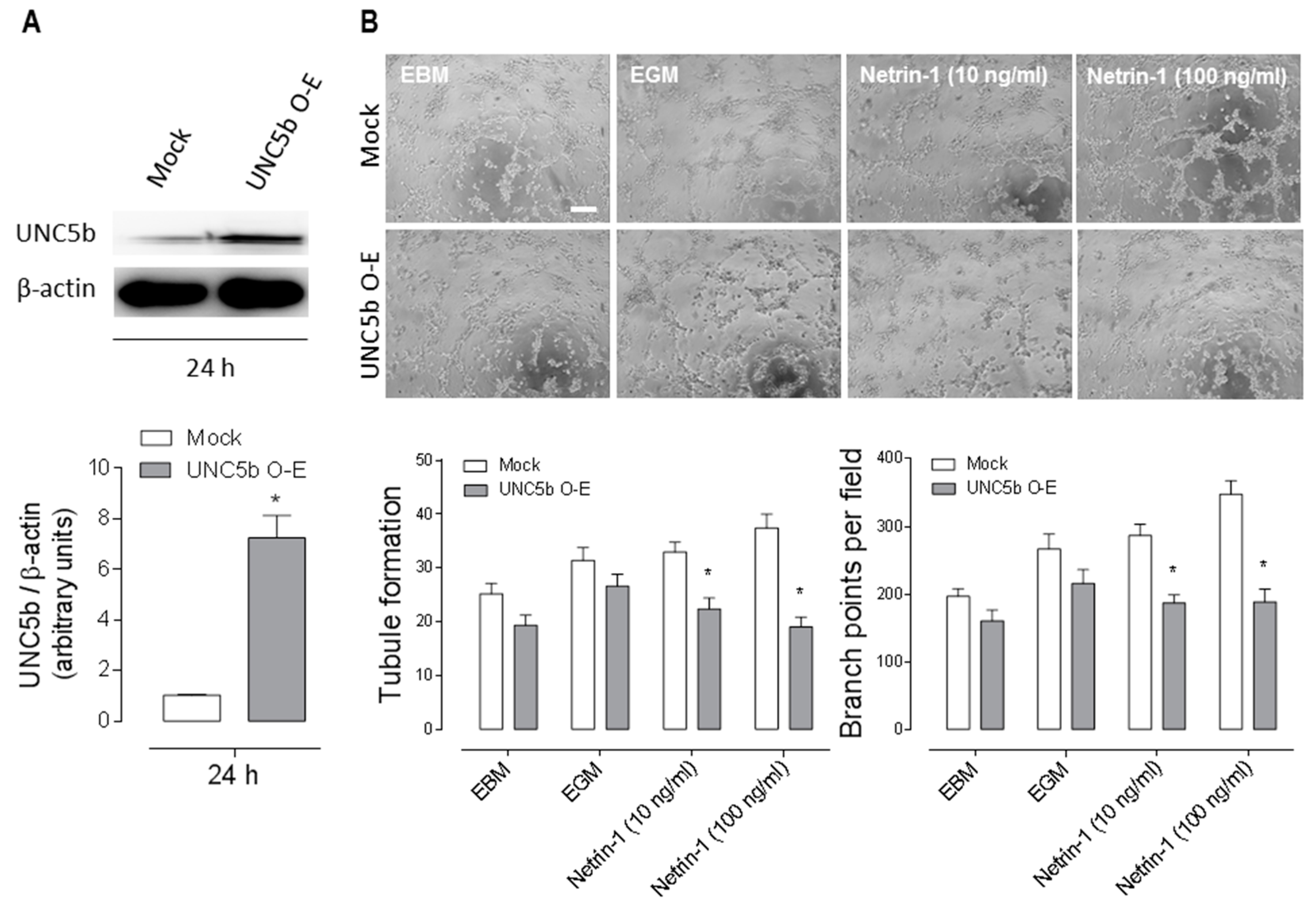
2.4. Overexpression of the Anti-Angiogenic UNC5b Receptor Reduces Angiogenesis In Vitro
3. Discussion
4. Materials and Methods
4.1. Isolation and Cell Culture of WJ-MSC and HUVEC
4.2. Tube Formation Assay
4.3. Western Blot
4.4. Netrin-1 Determination in Conditioned Media (CM)
4.5. Overexpression of UNC5b and Tube Formation Assay
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Disclaimers
References
- Desoye, G.; Hauguel-de Mouzon, S. The Human Placenta in Gestational Diabetes Mellitus. Diabetes Care 2007, 30, S120–S126. [Google Scholar] [CrossRef] [PubMed]
- Reece, E.A. The fetal and maternal consequences of gestational diabetes mellitus. J. Matern Neonatal Med. Taylor Francis 2010, 23, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Jirkovská, M.; Kubínová, L.; Janáček, J.; Moravcová, M.; Krejčí, V.; Karen, P. Topological Properties and Spatial Organization of Villous Capillaries in Normal and Diabetic Placentas. J. Vasc. Res. 2002, 39, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.J.E.-J. The Morphology of Villous Capillary Bed in Normal and Diabetic Placenta. Rijeka IntechOpen 2012. [Google Scholar] [CrossRef]
- Huynh, J.; Dawson, D.; Roberts, D.; Bentley-Lewis, R. A systematic review of placental pathology in maternal diabetes mellitus. Placenta 2015, 36, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Sobrevia, L.; Abarzúa, F.; Nien, J.K.; Salomón, C.; Westermeier, F.; Puebla, C.; Cifuentes, F.; Guzmán-Gutiérrez, E.; Leiva, A.; Casanello, P. Review: Differential placental macrovascular and microvascular endothelial dysfunction in gestational diabetes. Placenta 2011, 32, S159–S164. [Google Scholar] [CrossRef] [PubMed]
- Serafini, T.; Kennedy, T.E.; Gaiko, M.J.; Mirzayan, C.; Jessell, T.M.; Tessier-Lavigne, M. The netrins define a family of axon outgrowth-promoting proteins homologous to C. elegans UNC-6. Cell 1994, 78, 409–424. [Google Scholar] [CrossRef]
- Dakouane-Giudicelli, M.; Duboucher, C.; Fortemps, J.; Missey-Kolb, H.; Brulé, D.; Giudicelli, Y.; de Mazancourt, P. Characterization and Expression of Netrin-1 and Its Receptors UNC5B and DCC in Human Placenta. J. Histochem. Cytochem. 2009, 58, 73–82. [Google Scholar] [CrossRef]
- Park, K.W.; Crouse, D.; Lee, M.; Karnik, S.K.; Sorensen, L.K.; Murphy, K.J.; Kuo, C.J.; Li, D.Y. The axonal attractant Netrin-1 is an angiogenic factor. Proc. Natl. Acad. Sci. USA 2004, 101, 16210–16215. [Google Scholar] [CrossRef]
- Larrieu-Lahargue, F.; Thomas, K.R.; Li, D.Y. Netrin ligands and receptors: Lessons from neurons to the endothelium. Trends Cardiovasc. Med. 2012, 22, 44–47. [Google Scholar] [CrossRef]
- Navankasattusas, S.; Whitehead, K.J.; Suli, A.; Sorensen, L.K.; Lim, A.H.; Zhao, J.; Park, K.W.; Wythe, J.D.; Thomas, K.R.; Chien, C.-B.; et al. The netrin receptor UNC5B promotes angiogenesis in specific vascular beds. Development 2008, 135, 659–667. [Google Scholar] [CrossRef]
- Finci, L.I.; Krüger, N.; Sun, X.; Zhang, J.; Chegkazi, M.; Wu, Y.; Schenk, G.; Mertens, H.D.T.; Svergun, D.I.; Zhang, Y.; et al. The crystal structure of netrin-1 in complex with DCC reveals the bifunctionality of netrin-1 as a guidance cue. Neuron 2014, 83, 839–849. [Google Scholar] [CrossRef]
- Prieto, C.P.; Ortiz, M.C.; Villanueva, A.; Villarroel, C.; Edwards, S.S.; Elliott, M.; Lattus, J.; Aedo, S.; Meza, D.; Lois, P.; et al. Netrin-1 acts as a non-canonical angiogenic factor produced by human Wharton’s jelly mesenchymal stem cells (WJ-MSC). Stem Cell Res. Ther. 2017, 8, 43. [Google Scholar] [CrossRef]
- Nanaev, A.K.; Kohnen, G.; Milovanov, A.P.; Domogatsky, S.P.; Kaufmann, P. Stromal differentiation and architecture of the human umbilical cord. Placenta 1997, 18, 53–64. [Google Scholar] [CrossRef]
- Edwards, S.S.; Zavala, G.; Prieto, C.P.; Elliott, M.; Martínez, S.; Egaña, J.T.; Bono, M.R.; Palma, V. Functional analysis reveals angiogenic potential of human mesenchymal stem cells from Wharton’s jelly in dermal regeneration. Angiogenesis 2014, 17, 851–866. [Google Scholar] [CrossRef]
- Westermeier, F.; Salomón, C.; Farías, M.; Arroyo, P.; Fuenzalida, B.; Sáez, T.; Salsoso, R.; Sanhueza, C.; Guzmán-Gutiérrez, E.; Pardo, F.; et al. Insulin requires normal expression and signaling of insulin receptor A to reverse gestational diabetes-reduced adenosine transport in human umbilical vein endothelium. FASEB J. Fed. Am. Soc. Exp. Biol. 2014, 29, 37–49. [Google Scholar] [CrossRef]
- Boutsikou, T.; Giotaki, M.; Gourgiotis, D.; Boutsikou, M.; Briana, D.D.; Marmarinos, A.; Baka, S.; Hassiakos, D.; Malamitsi-Puchner, A. Cord blood netrin-1 and -4 concentrations in term pregnancies with normal, restricted and increased fetal growth. J. Matern Neonatal Med. Taylor Francis 2014, 27, 1849–1853. [Google Scholar] [CrossRef]
- Hunt, M.A.; Currie, M.J.; Robinson, B.A.; Dachs, G.U. Optimizing transfection of primary human umbilical vein endothelial cells using commercially available chemical transfection reagents. J. Biomol. Tech. 2010, 21, 66–72. [Google Scholar]
- Larrivée, B.; Freitas, C.; Trombe, M.; Lv, X.; Delafarge, B.; Yuan, L.; Bouvrée, K.; Bréant, C.; Del Toro, R.; Bréchot, N.; et al. Activation of the UNC5B receptor by Netrin-1 inhibits sprouting angiogenesis. Genes Dev. 2007, 21, 2433–2447. [Google Scholar] [CrossRef]
- Mission, J.F.; Catov, J.; Deihl, T.E.; Feghali, M.; Scifres, C. Early Pregnancy Diabetes Screening and Diagnosis: Prevalence, Rates of Abnormal Test Results, and Associated Factors. Obstet. Gynecol. 2017, 130, 1136–1142. [Google Scholar] [CrossRef]
- Castellucci, M.; Kosanke, G.; Verdenelli, F.; Huppertz, B.; Kaufmann, P. Villous sprouting: Fundamental mechanisms of human placental development. Hum. Reprod. Update 2000, 6, 485–494. [Google Scholar] [CrossRef]
- Tai-Nagara, I.; Yoshikawa, Y.; Numata, N.; Ando, T.; Okabe, K.; Sugiura, Y.; Ieda, M.; Takakura, N.; Nakagawa, O.; Zhou, B.; et al. Placental labyrinth formation in mice requires endothelial FLRT2/UNC5B signaling. Development 2017, 144, 2392–2401. [Google Scholar] [CrossRef]
- Shimizu, A.; Nakayama, H.; Wang, P.; König, C.; Akino, T.; Sandlund, J.; Coma, S.; Italiano, J.E., Jr.; Mammoto, A.; Bielenberg, D.R.; et al. Netrin-1 promotes glioblastoma cell invasiveness and angiogenesis by multiple pathways including activation of RhoA, cathepsin B, and cAMP-response element-binding protein. J. Biol. Chem. 2013, 288, 2210–2222. [Google Scholar] [CrossRef]
- Rajasekharan, S.; Baker, K.A.; Horn, K.E.; Jarjour, A.A.; Antel, J.P.; Kennedy, T.E. Netrin 1 and Dcc regulate oligodendrocyte process branching and membrane extension via Fyn and RhoA. Development 2009, 136, 415–426. [Google Scholar] [CrossRef]
- Antoine-Bertrand, J.; Ghogha, A.; Luangrath, V.; Bedford, F.K.; Lamarche-Vane, N. The activation of ezrin-radixin-moesin proteins is regulated by netrin-1 through Src kinase and RhoA/Rho kinase activities and mediates netrin-1-induced axon outgrowth. Mol. Biol. Cell 2011, 22, 3734–3746. [Google Scholar] [CrossRef]
- Mediero, A.; Ramkhelawon, B.; Perez-Aso, M.; Moore, K.J.; Cronstein, B.N. Netrin-1 is a critical autocrine/paracrine factor for osteoclast differentiation. J. Bone Miner. Res. 2015, 30, 837–854. [Google Scholar] [CrossRef]
- Liu, C.; Ke, X.; Wang, Y.; Feng, X.; Li, Q.; Zhang, Y.; Zhu, J.; Li, Q. The level of netrin-1 is decreased in newly diagnosed type 2 diabetes mellitus patients. BMC Endocr. Disord. 2016, 16, 33. [Google Scholar] [CrossRef]
- Toque, H.A.; Fernandez-Flores, A.; Mohamed, R.; Caldwell, R.B.; Ramesh, G.; Caldwell, R.W. Netrin-1 is a novel regulator of vascular endothelial function in diabetes. PLoS ONE 2017, 12, e0186734. [Google Scholar] [CrossRef]
- Tu, T.; Zhang, C.; Yan, H.; Luo, Y.; Kong, R.; Wen, P.; Ye, Z.; Chen, J.; Feng, J.; Liu, F.; et al. CD146 acts as a novel receptor for netrin-1 in promoting angiogenesis and vascular development. Cell Res. 2015, 25, 275–287. [Google Scholar] [CrossRef]
- Ramkhelawon, B.; Hennessy, E.J.; Ménager, M.; Ray, T.D.; Sheedy, F.J.; Hutchison, S.; Wanschel, A.; Oldebeken, S.; Geoffrion, M.; Spiro, W.; et al. Netrin-1 promotes adipose tissue macrophage retention and insulin resistance in obesity. Nat. Med. 2014, 20, 377–384. [Google Scholar] [CrossRef]
- Miloudi, K.; Binet, F.; Wilson, A.; Cerani, A.; Oubaha, M.; Menard, C.; Henriques, S.; Mawambo, G.; Dejda, A.; Nguyen, P.T.; et al. Truncated netrin-1 contributes to pathological vascular permeability in diabetic retinopathy. J. Clin. Investig. 2016, 126, 3006–3022. [Google Scholar] [CrossRef]
- Castets, M.; Coissieux, M.-M.; Delloye-Bourgeois, C.; Bernard, L.; Delcros, J.-G.; Bernet, A.; Laudet, V.; Mehlen, P. Inhibition of Endothelial Cell Apoptosis by Netrin-1 during Angiogenesis. Dev. Cell. 2009, 16, 614–620. [Google Scholar] [CrossRef]
- Kefeli, U.; Ucuncu Kefeli, A.; Cabuk, D.; Isik, U.; Sonkaya, A.; Acikgoz, O.; Ozden, E.; Uygun, K. Netrin-1 in cancer: Potential biomarker and therapeutic target? Tumor Biol. 2017, 39. [Google Scholar] [CrossRef]
- Lee, J.H.; Parveen, A.; Do, M.H.; Lim, Y.; Shim, S.H.; Kim, S.Y. Lespedeza cuneata protects the endothelial dysfunction via eNOS phosphorylation of PI3K/Akt signaling pathway in HUVECs. Phytomedicine 2018, 48, 1–9. [Google Scholar] [CrossRef]
- Karar, J.; Maity, A. PI3K/AKT/mTOR Pathway in Angiogenesis. Front. Mol. Neurosci. 2011, 4, 51. [Google Scholar] [CrossRef]
- Troncoso, F.; Acurio, J.; Herlitz, K.; Aguayo, C.; Bertoglia, P.; Guzman-Gutierrez, E.; Loyola, M.; Gonzalez, M.; Rezgaoui, M.; Desoye, G.; et al. Gestational diabetes mellitus is associated with increased pro-migratory activation of vascular endothelial growth factor receptor 2 and reduced expression of vascular endothelial growth factor receptor 1. PLoS ONE 2017, 12, e0182509. [Google Scholar] [CrossRef]
- Yang, Y.; Zou, L.; Xu, K.S. Expression of netrin-1 in placenta from patients with pre-eclampsia and the relation to placental angiogenesis. Zhonghua Fu Chan Ke Za Zhi 2006, 41, 597–600. [Google Scholar]
- Qian-hua, W.; Shao-ping, Z.; Jian-wen, Z.; Yun, Y.; Li, Z. Reduced expression of netrin-1 is associated with fetal growth restriction. Mol. Cell Biochem. 2011, 350, 81–87. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Healthy (n = 40) | GDM (n = 25) |
|---|---|---|
| Age (years) | 30 ± 7 | 31 ± 7 |
| Height (m) | 1.54 ± 0.04 | 1.55 ± 0.05 |
| Weight (kg) | 65.6 ± 6 | 63.7 ± 3 |
| Body mass index (kg/m2) | 27.8 ± 0.1 | 26.4 ± 0.2 |
| Fasting blood glucose | 78 ± 6.7 | 78.3 ± 3.7 |
| Postprandial blood Glucose (2 h) | 99 ± 16.7 | 151 ± 9.6 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prieto, C.P.; Casas, B.S.; Falcón, P.; Villanueva, A.; Lois, P.; Lattus, J.; Palma, V. Downregulation of the Netrin-1 Receptor UNC5b Underlies Increased Placental Angiogenesis in Human Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2019, 20, 1408. https://doi.org/10.3390/ijms20061408
Prieto CP, Casas BS, Falcón P, Villanueva A, Lois P, Lattus J, Palma V. Downregulation of the Netrin-1 Receptor UNC5b Underlies Increased Placental Angiogenesis in Human Gestational Diabetes Mellitus. International Journal of Molecular Sciences. 2019; 20(6):1408. https://doi.org/10.3390/ijms20061408
Chicago/Turabian StylePrieto, Catalina P., Bárbara S. Casas, Paulina Falcón, Andrea Villanueva, Pablo Lois, José Lattus, and Verónica Palma. 2019. "Downregulation of the Netrin-1 Receptor UNC5b Underlies Increased Placental Angiogenesis in Human Gestational Diabetes Mellitus" International Journal of Molecular Sciences 20, no. 6: 1408. https://doi.org/10.3390/ijms20061408
APA StylePrieto, C. P., Casas, B. S., Falcón, P., Villanueva, A., Lois, P., Lattus, J., & Palma, V. (2019). Downregulation of the Netrin-1 Receptor UNC5b Underlies Increased Placental Angiogenesis in Human Gestational Diabetes Mellitus. International Journal of Molecular Sciences, 20(6), 1408. https://doi.org/10.3390/ijms20061408