The Roles of Indian Hedgehog Signaling in TMJ Formation

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Recent Experimental Findings
2.1. Abnormal TMJ Development in Ihh-Null Mice at Embryonic and Early Postnatal Life
2.2. Role of Ihh in TMJ Growth and Maintenance during Postnatal Stages
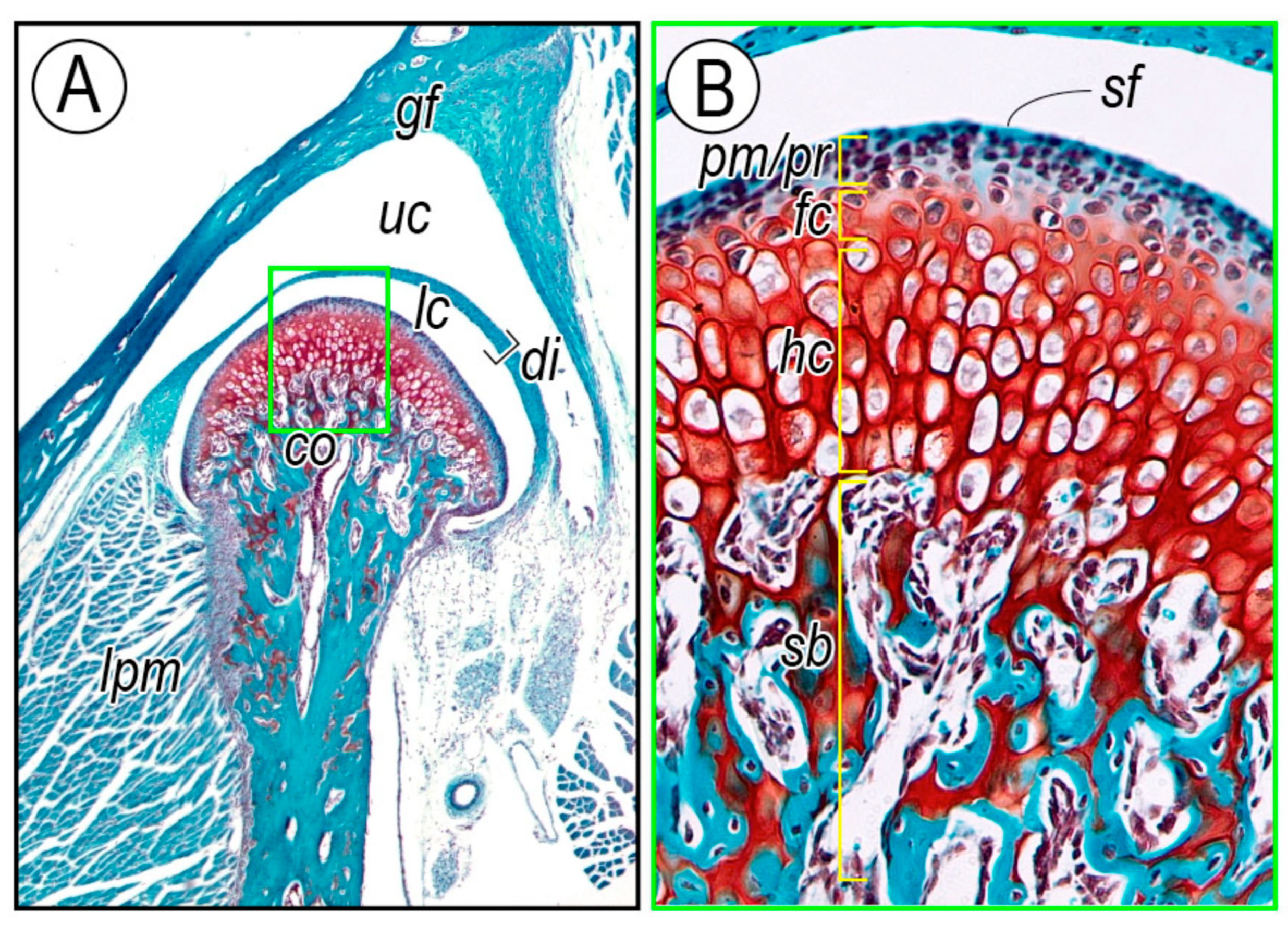
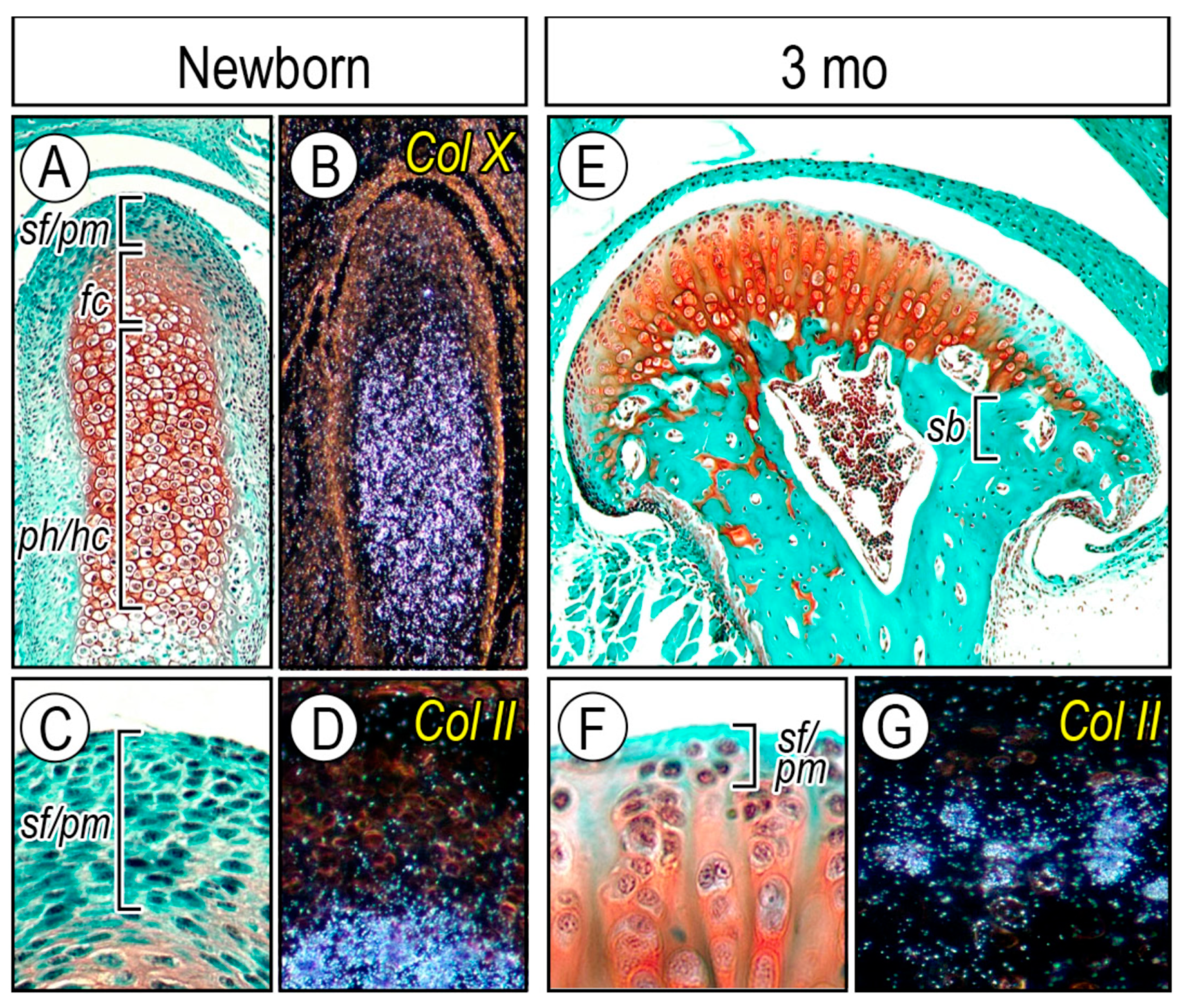
2.2.1. Cellular Organization of Condylar Cartilage in Postnatal Stages
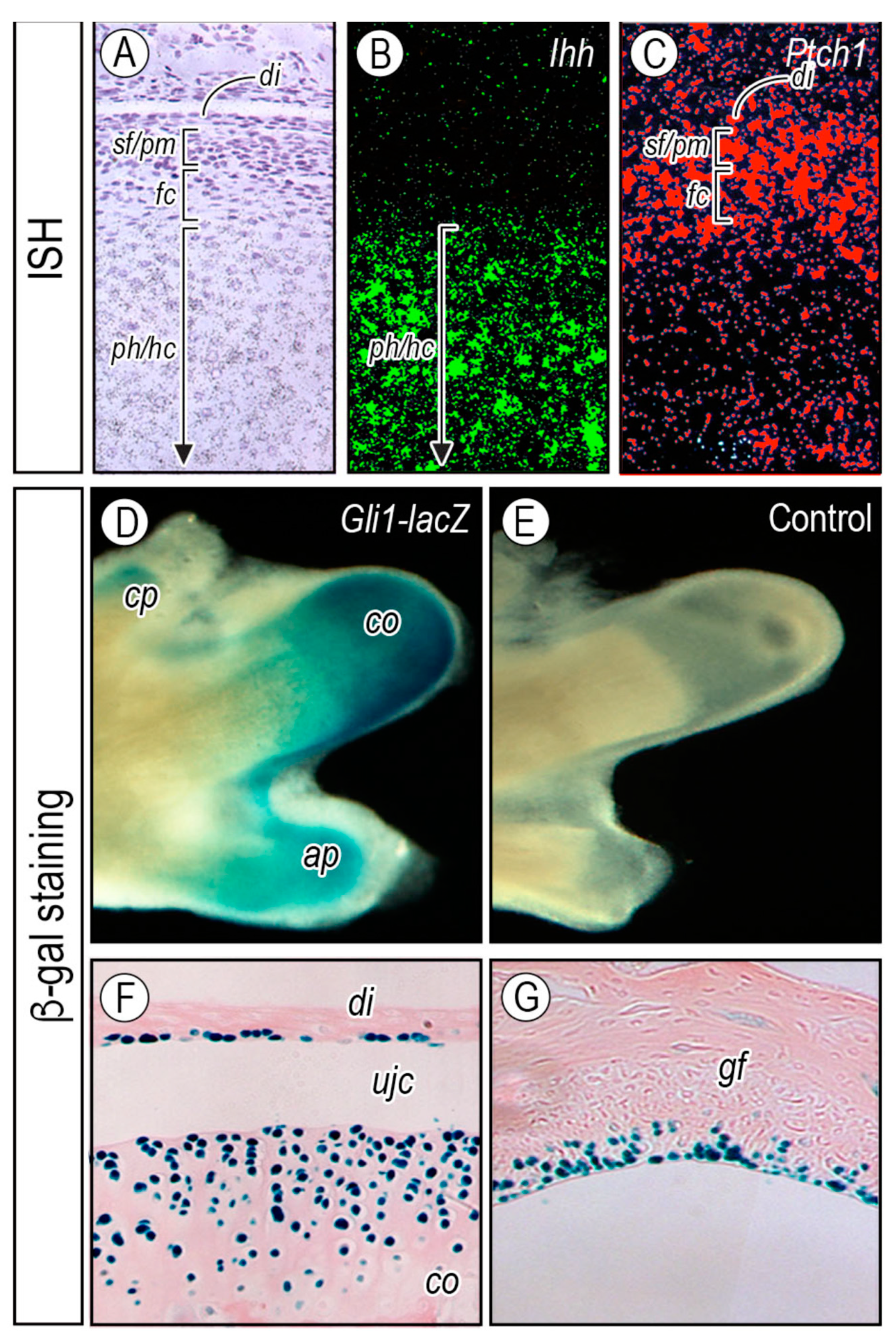
2.2.2. Topography of Hedgehog Signaling
2.2.3. Effect of Conditional Ihh Signaling Ablation in Postnatal Stages
2.2.4. Hh Signaling in Degenerative TMJs
3. Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rux, D.; Decker, R.S.; Koyama, E.; Pacifici, M. Joints in the appendicular skeleton: Developmental mechanisms and evolutionary influences. Curr. Top. Dev. Biol. 2019, 133, 119–151. [Google Scholar] [CrossRef] [PubMed]
- Pacifici, M.; Decker, R.S.; Koyama, E. Limb Synovial Joint Development From the Hips Down: Implications for Articular Cartilage Repair and Regeneration. In Developmental Biology and Musculoskeletal Tissue Engineering; Academic Press: Cambridge, MA, USA, 2018; pp. 67–101. Available online: https://www.sciencedirect.com/science/article/pii/B9780128114674000048 (accessed on 12 December 2019).
- Koyama, E.; Yasuda, T.; Minugh-Purvis, N.; Kinumatsu, T.; Yallowitz, A.R.; Wellik, D.M.; Pacifici, M. Hox11 genes establish synovial joint organization and phylogenetic characteristics in developing mouse zeugopod skeletal elements. Development 2010, 137, 3795–3800. [Google Scholar] [CrossRef] [PubMed]
- Hinton, R.J. Genes that regulate morphogenesis and growth of the temporomandibular joint: A review. Dev. Dyn. 2014, 243, 864–874. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Iwata, J. Mouse genetic models for temporomandibular joint development and disorders. Oral Dis. 2016, 22, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Symons, N.B. A histochemical study of the secondary cartilage of the mandibular condyle in the rat. Arch. Oral Biol. 1965, 10, 579–584. [Google Scholar] [CrossRef]
- Koyama, E.; Shibukawa, Y.; Nagayama, M.; Sugito, H.; Young, B.; Yuasa, T.; Okabe, T.; Ochiai, T.; Kamiya, N.; Rountree, R.B.; et al. A distinct cohort of progenitor cells participates in synovial joint and articular cartilage formation during mouse limb skeletogenesis. Dev. Biol. 2008, 316, 62–73. [Google Scholar] [CrossRef]
- Shibata, S.; Fukada, K.; Suzuki, S.; Ogawa, T.; Yamashita, Y. In situ hybridization and immunohistochemistry of bone sialoprotein and secreted phosphoprotein 1 (osteopontin) in the developing mouse mandibular condylar cartilage compared with limb bud cartilage. J. Anat. 2002, 200, 309–320. [Google Scholar] [CrossRef]
- Vinkka, H. Secondary cartilages in the facial skeleton of the rat. Proc. Finn. Dent. Soc. 1982, 78 (Suppl. 7), 1–137. [Google Scholar]
- Radlanski, R.J.; Renz, H.; Klarkowski, M.C. Prenatal development of the human mandible. 3D reconstructions, morphometry and bone remodelling pattern, sizes 12-117 mm CRL. Anat. Embryol. 2003, 207, 221–232. [Google Scholar] [CrossRef]
- Shibukawa, Y.; Young, B.; Wu, C.; Yamada, S.; Long, F.; Pacifici, M.; Koyama, E. Temporomandibular joint formation and condyle growth require Indian hedgehog signaling. Dev. Dyn. 2007, 236, 426–434. [Google Scholar] [CrossRef]
- Wadhwa, S.; Kapila, S. TMJ disorders: Future innovations in diagnostics and therapeutics. J. Dent. Educ. 2008, 72, 930–947. [Google Scholar] [PubMed]
- Luder, H.U.; Leblond, C.P.; von der Mark, K. Cellular stages in cartilage formation as revealed by morphometry, radioautography and type II collagen immunostaining of the mandibular condyle from weanling rats. Am. J. Anat 1988, 182, 197–214. [Google Scholar] [CrossRef] [PubMed]
- Silbermann, M.; Frommer, J. The nature of endochondral ossification in the mandibular condyle of the mouse. Anat. Rec. 1972, 172, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Sarnat, B.G. Developmental facial abnormalities and the temporomandibular joint. Dent. Clin. N. Am. 1966, 79, 587–600. [Google Scholar] [CrossRef]
- Petrovic, A.G. Mechanisms and regulation of mandibular condylar growth. Acta Morphol. Neerl. Scand. 1972, 10, 25–34. [Google Scholar] [PubMed]
- Kantomaa, T.; Tuominen, M.; Pirttiniemi, P. Effect of mechanical forces on chondrocyte maturation and differentiation in the mandibular condyle of the rat. J. Dent. Res. 1994, 73, 1150–1156. [Google Scholar] [CrossRef]
- Jing, Y.; Zhou, X.; Han, X.; Jing, J.; von der Mark, K.; Wang, J.; de Crombrugghe, B.; Hinton, R.J.; Feng, J.Q. Chondrocytes Directly Transform into Bone Cells in Mandibular Condyle Growth. J. Dent. Res. 2015, 94, 1668–1675. [Google Scholar] [CrossRef]
- Frommer, J. Prenatal Development of the Mandibular Joint in Mice. Anat. Rec. 1964, 150, 449–461. [Google Scholar] [CrossRef]
- Bhaskar, S.N. Growth pattern of the rat mandible from 13 days insemination age to 30 days after birth. Am. J. Anat 1953, 92, 1–53. [Google Scholar] [CrossRef]
- Mori-Akiyama, Y.; Akiyama, H.; Rowitch, D.H.; de Crombrugghe, B. Sox9 is required for determination of the chondrogenic cell lineage in the cranial neural crest. Proc. Natl. Acad. Sci. USA 2003, 100, 9360–9365. [Google Scholar] [CrossRef]
- Gu, S.; Wei, N.; Yu, L.; Fei, J.; Chen, Y. Shox2-deficiency leads to dysplasia and ankylosis of the temporomandibular joint in mice. Mech. Dev. 2008, 125, 729–742. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, T.; Nah, H.D.; Laurita, J.; Kinumatsu, T.; Shibukawa, Y.; Shibutani, T.; Minugh-Purvis, N.; Pacifici, M.; Koyama, E. Muenke syndrome mutation, FgfR3P(2)(4)(4)R, causes TMJ defects. J. Dent. Res. 2012, 91, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Koyama, E.; Saunders, C.; Salhab, I.; Decker, R.S.; Chen, I.; Um, H.; Pacifici, M.; Nah, H.D. Lubricin is Required for the Structural Integrity and Post-natal Maintenance of TMJ. J. Dent. Res. 2014, 93, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Bechtold, T.E.; Saunders, C.; Decker, R.S.; Um, H.B.; Cottingham, N.; Salhab, I.; Kurio, N.; Billings, P.C.; Pacifici, M.; Nah, H.D.; et al. Osteophyte formation and matrix mineralization in a TMJ osteoarthritis mouse model are associated with ectopic hedgehog signaling. Matrix Biol. 2016, 52-54, 339–354. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, C.; Rohr, J.; Liu, H.; He, F.; Yu, J.; Sun, C.; Li, L.; Gu, S.; Chen, Y. Tissue interaction is required for glenoid fossa development during temporomandibular joint formation. Dev. Dyn. 2011, 240, 2466–2473. [Google Scholar] [CrossRef]
- Lee, R.T.; Zhao, Z.; Ingham, P.W. Hedgehog signalling. Development 2016, 143, 367–372. [Google Scholar] [CrossRef]
- Alman, B.A. The role of hedgehog signalling in skeletal health and disease. Nat. Rev. Rheumatol. 2015, 11, 552–560. [Google Scholar] [CrossRef]
- Yao, E.; Chuang, P.T. Hedgehog signaling: From basic research to clinical applications. J. Formos. Med. Assoc. 2015, 114, 569–576. [Google Scholar] [CrossRef]
- Abramyan, J. Hedgehog Signaling and Embryonic Craniofacial Disorders. J. Dev. Biol. 2019, 7. [Google Scholar] [CrossRef]
- Bitgood, M.J.; McMahon, A.P. Hedgehog and Bmp genes are coexpressed at many diverse sites of cell-cell interaction in the mouse embryo. Dev. Biol. 1995, 172, 126–138. [Google Scholar] [CrossRef]
- Koyama, E.; Leatherman, J.L.; Noji, S.; Pacifici, M. Early chick limb cartilaginous elements possess polarizing activity and express hedgehog-related morphogenetic factors. Dev. Dyn. 1996, 207, 344–354. [Google Scholar] [CrossRef]
- Nakamura, T.; Aikawa, T.; Iwamoto-Enomoto, M.; Iwamoto, M.; Higuchi, Y.; Pacifici, M.; Kinto, N.; Yamaguchi, A.; Noji, S.; Kurisu, K.; et al. Induction of osteogenic differentiation by hedgehog proteins. Biochem. Biophys. Res. Commun. 1997, 237, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Long, F.; Chung, U.I.; Ohba, S.; McMahon, J.; Kronenberg, H.M.; McMahon, A.P. Ihh signaling is directly required for the osteoblast lineage in the endochondral skeleton. Development 2004, 131, 1309–1318. [Google Scholar] [CrossRef] [PubMed]
- Vortkamp, A.; Lee, K.; Lanske, B.; Segre, G.V.; Kronenberg, H.M.; Tabin, C.J. Regulation of rate of cartilage differentiation by Indian hedgehog and PTH-related protein. Science 1996, 273, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Lanske, B.; Karaplis, A.C.; Lee, K.; Luz, A.; Vortkamp, A.; Pirro, A.; Karperien, M.; Defize, L.H.; Ho, C.; Mulligan, R.C.; et al. PTH/PTHrP receptor in early development and Indian hedgehog-regulated bone growth. Science 1996, 273, 663–666. [Google Scholar] [CrossRef] [PubMed]
- St-Jacques, B.; Hammerschmidt, M.; McMahon, A.P. Indian hedgehog signaling regulates proliferation and differentiation of chondrocytes and is essential for bone formation. Genes Dev. 1999, 13, 2072–2086. [Google Scholar] [CrossRef]
- Chung, U.I.; Schipani, E.; McMahon, A.P.; Kronenberg, H.M. Indian hedgehog couples chondrogenesis to osteogenesis in endochondral bone development. J. Clin. Investig. 2001, 107, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Alcedo, J.; Ayzenzon, M.; Von Ohlen, T.; Noll, M.; Hooper, J.E. The Drosophila smoothened gene encodes a seven-pass membrane protein, a putative receptor for the hedgehog signal. Cell 1996, 86, 221–232. [Google Scholar] [CrossRef]
- van den Heuvel, M.; Ingham, P.W. smoothened encodes a receptor-like serpentine protein required for hedgehog signalling. Nature 1996, 382, 547–551. [Google Scholar] [CrossRef]
- Chen, W.; Burgess, S.; Hopkins, N. Analysis of the zebrafish smoothened mutant reveals conserved and divergent functions of hedgehog activity. Development 2001, 128, 2385–2396. [Google Scholar]
- Goodrich, L.V.; Milenkovic, L.; Higgins, K.M.; Scott, M.P. Altered neural cell fates and medulloblastoma in mouse patched mutants. Science 1997, 277, 1109–1113. [Google Scholar] [CrossRef] [PubMed]
- Haycraft, C.J.; Serra, R. Cilia involvement in patterning and maintenance of the skeleton. Curr. Top. Dev. Biol. 2008, 85, 303–332. [Google Scholar] [CrossRef] [PubMed]
- Sreekumar, V.; Norris, D.P. Cilia and development. Curr. Opin. Genet. Dev. 2019, 56, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Nachury, M.V.; Mick, D.U. Establishing and regulating the composition of cilia for signal transduction. Nat. Rev. Mol. Cell Biol. 2019, 20, 389–405. [Google Scholar] [CrossRef]
- Ruiz i Altaba, A. Gli proteins encode context-dependent positive and negative functions: Implications for development and disease. Development 1999, 126, 3205–3216. [Google Scholar]
- Sasaki, H.; Nishizaki, Y.; Hui, C.; Nakafuku, M.; Kondoh, H. Regulation of Gli2 and Gli3 activities by an amino-terminal repression domain: Implication of Gli2 and Gli3 as primary mediators of Shh signaling. Development 1999, 126, 3915–3924. [Google Scholar]
- Wong, S.Y.; Reiter, J.F. The primary cilium at the crossroads of mammalian hedgehog signaling. Curr. Top. Dev. Biol. 2008, 85, 225–260. [Google Scholar] [CrossRef]
- Hynes, M.; Stone, D.M.; Dowd, M.; Pitts-Meek, S.; Goddard, A.; Gurney, A.; Rosenthal, A. Control of cell pattern in the neural tube by the zinc finger transcription factor and oncogene Gli-1. Neuron 1997, 19, 15–26. [Google Scholar] [CrossRef]
- Karlstrom, R.O.; Tyurina, O.V.; Kawakami, A.; Nishioka, N.; Talbot, W.S.; Sasaki, H.; Schier, A.F. Genetic analysis of zebrafish gli1 and gli2 reveals divergent requirements for gli genes in vertebrate development. Development 2003, 130, 1549–1564. [Google Scholar] [CrossRef]
- Lee, J.; Platt, K.A.; Censullo, P.; Ruiz i Altaba, A. Gli1 is a target of Sonic hedgehog that induces ventral neural tube development. Development 1997, 124, 2537–2552. [Google Scholar]
- Koyama, E.; Ochiai, T.; Rountree, R.B.; Kingsley, D.M.; Enomoto-Iwamoto, M.; Iwamoto, M.; Pacifici, M. Synovial joint formation during mouse limb skeletogenesis: Roles of Indian hedgehog signaling. Ann. N. Y. Acad. Sci. 2007, 1116, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Decker, R.S.; Koyama, E.; Pacifici, M. Genesis and morphogenesis of limb synovial joints and articular cartilage. Matrix Biol. 2014, 39, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Kubiak, M.; Ditzel, M. A Joint Less Ordinary: Intriguing Roles for Hedgehog Signalling in the Development of the Temporomandibular Synovial Joint. J. Dev. Biol. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Maeda, Y.; Nakamura, E.; Nguyen, M.T.; Suva, L.J.; Swain, F.L.; Razzaque, M.S.; Mackem, S.; Lanske, B. Indian Hedgehog produced by postnatal chondrocytes is essential for maintaining a growth plate and trabecular bone. Proc. Natl. Acad. Sci. USA 2007, 104, 6382–6387. [Google Scholar] [CrossRef] [PubMed]
- Hilton, M.J.; Tu, X.; Cook, J.; Hu, H.; Long, F. Ihh controls cartilage development by antagonizing Gli3, but requires additional effectors to regulate osteoblast and vascular development. Development 2005, 132, 4339–4351. [Google Scholar] [CrossRef]
- Purcell, P.; Joo, B.W.; Hu, J.K.; Tran, P.V.; Calicchio, M.L.; O’Connell, D.J.; Maas, R.L.; Tabin, C.J. Temporomandibular joint formation requires two distinct hedgehog-dependent steps. Proc. Natl. Acad. Sci. USA 2009, 106, 18297–18302. [Google Scholar] [CrossRef] [PubMed]
- Mo, R.; Freer, A.M.; Zinyk, D.L.; Crackower, M.A.; Michaud, J.; Heng, H.H.; Chik, K.W.; Shi, X.M.; Tsui, L.C.; Cheng, S.H.; et al. Specific and redundant functions of Gli2 and Gli3 zinc finger genes in skeletal patterning and development. Development 1997, 124, 113–123. [Google Scholar]
- Michikami, I.; Fukushi, T.; Honma, S.; Yoshioka, S.; Itoh, S.; Muragaki, Y.; Kurisu, K.; Ooshima, T.; Wakisaka, S.; Abe, M. Trps1 is necessary for normal temporomandibular joint development. Cell Tissue Res. 2012, 348, 131–140. [Google Scholar] [CrossRef]
- Yang, L.; Gu, S.; Ye, W.; Song, Y.; Chen, Y. Augmented Indian hedgehog signaling in cranial neural crest cells leads to craniofacial abnormalities and dysplastic temporomandibular joint in mice. Cell Tissue Res. 2016, 364, 105–115. [Google Scholar] [CrossRef]
- Bertolacini, C.D.; Ribeiro-Bicudo, L.A.; Petrin, A.; Richieri-Costa, A.; Murray, J.C. Clinical findings in patients with GLI2 mutations—Phenotypic variability. Clin. Genet. 2012, 81, 70–75. [Google Scholar] [CrossRef]
- Mak, K.K.; Kronenberg, H.M.; Chuang, P.T.; Mackem, S.; Yang, Y. Indian hedgehog signals independently of PTHrP to promote chondrocyte hypertrophy. Development 2008, 135, 1947–1956. [Google Scholar] [CrossRef] [PubMed]
- Karp, S.J.; Schipani, E.; St-Jacques, B.; Hunzelman, J.; Kronenberg, H.; McMahon, A.P. Indian hedgehog coordinates endochondral bone growth and morphogenesis via parathyroid hormone related-protein-dependent and -independent pathways. Development 2000, 127, 543–548. [Google Scholar] [PubMed]
- Embree, M.C.; Chen, M.; Pylawka, S.; Kong, D.; Iwaoka, G.M.; Kalajzic, I.; Yao, H.; Shi, C.; Sun, D.; Sheu, T.J.; et al. Exploiting endogenous fibrocartilage stem cells to regenerate cartilage and repair joint injury. Nat. Commun. 2016, 7, 13073. [Google Scholar] [CrossRef] [PubMed]
- Kaul, R.; O’Brien, M.H.; Dutra, E.; Lima, A.; Utreja, A.; Yadav, S. The Effect of Altered Loading on Mandibular Condylar Cartilage. PLoS ONE 2016, 11, e0160121. [Google Scholar] [CrossRef] [PubMed]
- Ishizuka, Y.; Shibukawa, Y.; Nagayama, M.; Decker, R.; Kinumatsu, T.; Saito, A.; Pacifici, M.; Koyama, E. TMJ degeneration in SAMP8 mice is accompanied by deranged Ihh signaling. J. Dent. Res. 2014, 93, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Kurio, N.; Saunders, C.; Bechtold, T.E.; Salhab, I.; Nah, H.D.; Sinha, S.; Billings, P.C.; Pacifici, M.; Koyama, E. Roles of Ihh signaling in chondroprogenitor function in postnatal condylar cartilage. Matrix Biol. 2018, 67, 15–31. [Google Scholar] [CrossRef]
- Bai, C.B.; Auerbach, W.; Lee, J.S.; Stephen, D.; Joyner, A.L. Gli2, but not Gli1, is required for initial Shh signaling and ectopic activation of the Shh pathway. Development 2002, 129, 4753–4761. [Google Scholar]
- Ochiai, T.; Shibukawa, Y.; Nagayama, M.; Mundy, C.; Yasuda, T.; Okabe, T.; Shimono, K.; Kanyama, M.; Hasegawa, H.; Maeda, Y.; et al. Indian hedgehog roles in post-natal TMJ development and organization. J. Dent. Res. 2010, 89, 349–354. [Google Scholar] [CrossRef]
- Henry, S.P.; Jang, C.W.; Deng, J.M.; Zhang, Z.; Behringer, R.R.; de Crombrugghe, B. Generation of aggrecan-CreERT2 knockin mice for inducible Cre activity in adult cartilage. Genesis 2009, 47, 805–814. [Google Scholar] [CrossRef]
- Chen, W.H.; Hosokawa, M.; Tsuboyama, T.; Ono, T.; Iizuka, T.; Takeda, T. Age-related changes in the temporomandibular joint of the senescence accelerated mouse. SAM-P/3 as a new murine model of degenerative joint disease. Am. J. Pathol. 1989, 135, 379–385. [Google Scholar]
- Hosokawa, M.; Kasai, R.; Higuchi, K.; Takeshita, S.; Shimizu, K.; Hamamoto, H.; Honma, A.; Irino, M.; Toda, K.; Matsumura, A.; et al. Grading score system: A method for evaluation of the degree of senescence in senescence accelerated mouse (SAM). Mech. Ageing Dev. 1984, 26, 91–102. [Google Scholar] [CrossRef]
- Scrivani, S.J.; Keith, D.A.; Kaban, L.B. Temporomandibular disorders. N. Engl. J. Med. 2008, 359, 2693–2705. [Google Scholar] [CrossRef] [PubMed]
- Dimitroulis, G. Management of temporomandibular joint disorders: A surgeon’s perspective. Aust. Dent. J. 2018, 63 (Suppl. 1), S79–S90. [Google Scholar] [CrossRef]
- Mercuri, L.G. Osteoarthritis, osteoarthrosis, and idiopathic condylar resorption. Oral Maxillofac. Surg. Clin. N. Am. 2008, 20, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Schiffman, E.; Ohrbach, R.; Truelove, E.; Look, J.; Anderson, G.; Goulet, J.P.; List, T.; Svensson, P.; Gonzalez, Y.; Lobbezoo, F.; et al. Diagnostic Criteria for Temporomandibular Disorders (DC/TMD) for Clinical and Research Applications: Recommendations of the International RDC/TMD Consortium Network* and Orofacial Pain Special Interest Groupdagger. J. Oral Fac. Pain Headache 2014, 28, 6–27. [Google Scholar] [CrossRef] [PubMed]
- Elsaid, K.A.; Machan, J.T.; Waller, K.; Fleming, B.C.; Jay, G.D. The impact of anterior cruciate ligament injury on lubricin metabolism and the effect of inhibiting tumor necrosis factor alpha on chondroprotection in an animal model. Arthritis Rheum. 2009, 60, 2997–3006. [Google Scholar] [CrossRef]
- Elsaid, K.A.; Fleming, B.C.; Oksendahl, H.L.; Machan, J.T.; Fadale, P.D.; Hulstyn, M.J.; Shalvoy, R.; Jay, G.D. Decreased lubricin concentrations and markers of joint inflammation in the synovial fluid of patients with anterior cruciate ligament injury. Arthritis Rheum. 2008, 58, 1707–1715. [Google Scholar] [CrossRef]
- Teeple, E.; Elsaid, K.A.; Fleming, B.C.; Jay, G.D.; Aslani, K.; Crisco, J.J.; Mechrefe, A.P. Coefficients of friction, lubricin, and cartilage damage in the anterior cruciate ligament-deficient guinea pig knee. J. Orthop. Res. 2008, 26, 231–237. [Google Scholar] [CrossRef]
- Kosinska, M.K.; Ludwig, T.E.; Liebisch, G.; Zhang, R.; Siebert, H.C.; Wilhelm, J.; Kaesser, U.; Dettmeyer, R.B.; Klein, H.; Ishaque, B.; et al. Articular Joint Lubricants during Osteoarthritis and Rheumatoid Arthritis Display Altered Levels and Molecular Species. PLoS ONE 2015, 10, e0125192. [Google Scholar] [CrossRef]
- Jay, G.D.; Waller, K.A. The biology of lubricin: Near frictionless joint motion. Matrix Biol. 2014, 39, 17–24. [Google Scholar] [CrossRef]
- Das, N.; Schmidt, T.A.; Krawetz, R.J.; Dufour, A. Proteoglycan 4: From Mere Lubricant to Regulator of Tissue Homeostasis and Inflammation: Does proteoglycan 4 have the ability to buffer the inflammatory response? Bioessays 2019, 41, e1800166. [Google Scholar] [CrossRef] [PubMed]
- Bahabri, S.A.; Suwairi, W.M.; Laxer, R.M.; Polinkovsky, A.; Dalaan, A.A.; Warman, M.L. The camptodactyly-arthropathy-coxa vara-pericarditis syndrome: Clinical features and genetic mapping to human chromosome 1. Arthritis Rheum. 1998, 41, 730–735. [Google Scholar] [CrossRef]
- Marcelino, J.; Carpten, J.D.; Suwairi, W.M.; Gutierrez, O.M.; Schwartz, S.; Robbins, C.; Sood, R.; Makalowska, I.; Baxevanis, A.; Johnstone, B.; et al. CACP, encoding a secreted proteoglycan, is mutated in camptodactyly-arthropathy-coxa vara-pericarditis syndrome. Nat. Genet. 1999, 23, 319–322. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.; Waller, K.A.; Cui, Y.; Allen, J.M.; Smits, P.; Zhang, L.X.; Ayturk, U.M.; Hann, S.; Lessard, S.G.; Zurakowski, D.; et al. Lubricin restoration in a mouse model of congenital deficiency. Arthritis Rheumatol. 2015, 67, 3070–3081. [Google Scholar] [CrossRef] [PubMed]
- Rhee, D.K.; Marcelino, J.; Baker, M.; Gong, Y.; Smits, P.; Lefebvre, V.; Jay, G.D.; Stewart, M.; Wang, H.; Warman, M.L.; et al. The secreted glycoprotein lubricin protects cartilage surfaces and inhibits synovial cell overgrowth. J. Clin. Investig. 2005, 115, 622–631. [Google Scholar] [CrossRef]
- Hill, A.; Duran, J.; Purcell, P. Lubricin protects the temporomandibular joint surfaces from degeneration. PLoS ONE 2014, 9, e106497. [Google Scholar] [CrossRef]
- Chen, J.; Gupta, T.; Barasz, J.A.; Kalajzic, Z.; Yeh, W.C.; Drissi, H.; Hand, A.R.; Wadhwa, S. Analysis of microarchitectural changes in a mouse temporomandibular joint osteoarthritis model. Arch. Oral Biol. 2009, 54, 1091–1098. [Google Scholar] [CrossRef][Green Version]
- Wadhwa, S.; Embree, M.; Ameye, L.; Young, M.F. Mice deficient in biglycan and fibromodulin as a model for temporomandibular joint osteoarthritis. Cells Tissues Organs 2005, 181, 136–143. [Google Scholar] [CrossRef]
- Embree, M.C.; Kilts, T.M.; Ono, M.; Inkson, C.A.; Syed-Picard, F.; Karsdal, M.A.; Oldberg, A.; Bi, Y.; Young, M.F. Biglycan and fibromodulin have essential roles in regulating chondrogenesis and extracellular matrix turnover in temporomandibular joint osteoarthritis. Am. J. Pathol. 2010, 176, 812–826. [Google Scholar] [CrossRef]
- Larheim, T.A.; Abrahamsson, A.K.; Kristensen, M.; Arvidsson, L.Z. Temporomandibular joint diagnostics using CBCT. Dentomaxillofac. Radiol. 2015, 44, 20140235. [Google Scholar] [CrossRef]
- Rehan, O.M.; Saleh, H.A.K.; Raffat, H.A.; Abu-Taleb, N.S. Osseous changes in the temporomandibular joint in rheumatoid arthritis: A cone-beam computed tomography study. Imaging Sci. Dent. 2018, 48, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Murata, K.; Kokubun, T.; Onitsuka, K.; Oka, Y.; Kano, T.; Morishita, Y.; Ozone, K.; Kuwabara, N.; Nishimoto, J.; Isho, T.; et al. Controlling joint instability after anterior cruciate ligament transection inhibits transforming growth factor-beta-mediated osteophyte formation. Osteoarthr. Cartil. 2019, 27, 1185–1196. [Google Scholar] [CrossRef] [PubMed]
- van der Kraan, P.M.; van den Berg, W.B. Osteophytes: Relevance and biology. Osteoarthr. Cartil. 2007, 15, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Blaney Davidson, E.N.; Vitters, E.L.; van der Kraan, P.M.; van den Berg, W.B. Expression of transforming growth factor-beta (TGFbeta) and the TGFbeta signalling molecule SMAD-2P in spontaneous and instability-induced osteoarthritis: Role in cartilage degradation, chondrogenesis and osteophyte formation. Ann. Rheum. Dis. 2006, 65, 1414–1421. [Google Scholar] [CrossRef]
- Jingushi, S.; Shida, J.; Iwamoto, Y.; Kinoshita, T.; Hiyama, Y.; Tamura, M.; Izumi, T. Transient exposure of fibroblast growth factor-2 induced proliferative but not destructive changes in mouse knee joints. Connect. Tissue Res. 2006, 47, 242–248. [Google Scholar] [CrossRef]
- Okazaki, K.; Jingushi, S.; Ikenoue, T.; Urabe, K.; Sakai, H.; Ohtsuru, A.; Akino, K.; Yamashita, S.; Nomura, S.; Iwamoto, Y. Expression of insulin-like growth factor I messenger ribonucleic acid in developing osteophytes in murine experimental osteoarthritis and in rats inoculated with growth hormone-secreting tumor. Endocrinology 1999, 140, 4821–4830. [Google Scholar] [CrossRef]
- Bechtold, T.E.; Saunders, C.; Mundy, C.; Um, H.; Decker, R.S.; Salhab, I.; Kurio, N.; Billings, P.C.; Pacifici, M.; Nah, H.D.; et al. Excess BMP Signaling in Heterotopic Cartilage Forming in Prg4-null TMJ Discs. J. Dent. Res. 2016, 95, 292–301. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, M.; Liu, Q.; Zhang, H.; Zhang, J.; Lu, L.; Xie, M.; Chen, D.; Wang, M. Inhibition of Ihh Reverses Temporomandibular Joint Osteoarthritis via a PTH1R Signaling Dependent Mechanism. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef]
- Cohen, M.M., Jr. The hedgehog signaling network. Am. J. Med. Genet. A 2003, 123A, 5–28. [Google Scholar] [CrossRef]
- Pepinsky, R.B.; Zeng, C.; Wen, D.; Rayhorn, P.; Baker, D.P.; Williams, K.P.; Bixler, S.A.; Ambrose, C.M.; Garber, E.A.; Miatkowski, K.; et al. Identification of a palmitic acid-modified form of human Sonic hedgehog. J. Biol. Chem. 1998, 273, 14037–14045. [Google Scholar] [CrossRef]
- Porter, J.A.; Ekker, S.C.; Park, W.J.; von Kessler, D.P.; Young, K.E.; Chen, C.H.; Ma, Y.; Woods, A.S.; Cotter, R.J.; Koonin, E.V.; et al. Hedgehog patterning activity: Role of a lipophilic modification mediated by the carboxy-terminal autoprocessing domain. Cell 1996, 86, 21–34. [Google Scholar] [CrossRef]
- Chen, M.H.; Li, Y.J.; Kawakami, T.; Xu, S.M.; Chuang, P.T. Palmitoylation is required for the production of a soluble multimeric Hedgehog protein complex and long-range signaling in vertebrates. Genes Dev. 2004, 18, 641–659. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Goetz, J.A.; Suber, L.M.; Scott, W.J., Jr.; Schreiner, C.M.; Robbins, D.J. A freely diffusible form of Sonic hedgehog mediates long-range signalling. Nature 2001, 411, 716–720. [Google Scholar] [CrossRef] [PubMed]
- Gallet, A.; Ruel, L.; Staccini-Lavenant, L.; Therond, P.P. Cholesterol modification is necessary for controlled planar long-range activity of Hedgehog in Drosophila epithelia. Development 2006, 133, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Goetz, J.A.; Singh, S.; Suber, L.M.; Kull, F.J.; Robbins, D.J. A highly conserved amino-terminal region of sonic hedgehog is required for the formation of its freely diffusible multimeric form. J. Biol. Chem. 2006, 281, 4087–4093. [Google Scholar] [CrossRef] [PubMed]
- Billings, P.C.; Pacifici, M. Interactions of signaling proteins, growth factors and other proteins with heparan sulfate: Mechanisms and mysteries. Connect. Tissue Res. 2015, 56, 272–280. [Google Scholar] [CrossRef]
- Radhakrishnan, A.; Sun, L.P.; Kwon, H.J.; Brown, M.S.; Goldstein, J.L. Direct binding of cholesterol to the purified membrane region of SCAP: Mechanism for a sterol-sensing domain. Mol. Cell 2004, 15, 259–268. [Google Scholar] [CrossRef]
- Tukachinsky, H.; Kuzmickas, R.P.; Jao, C.Y.; Liu, J.; Salic, A. Dispatched and scube mediate the efficient secretion of the cholesterol-modified hedgehog ligand. Cell Rep. 2012, 2, 308–320. [Google Scholar] [CrossRef]
- Kawakami, T.; Kawcak, T.; Li, Y.J.; Zhang, W.; Hu, Y.; Chuang, P.T. Mouse dispatched mutants fail to distribute hedgehog proteins and are defective in hedgehog signaling. Development 2002, 129, 5753–5765. [Google Scholar] [CrossRef]
- Jakobs, P.; Exner, S.; Schurmann, S.; Pickhinke, U.; Bandari, S.; Ortmann, C.; Kupich, S.; Schulz, P.; Hansen, U.; Seidler, D.G.; et al. Scube2 enhances proteolytic Shh processing from the surface of Shh-producing cells. J. Cell Sci. 2014, 127, 1726–1737. [Google Scholar] [CrossRef]
- Yasuda, T.; Mundy, C.; Kinumatsu, T.; Shibukawa, Y.; Shibutani, T.; Grobe, K.; Minugh-Purvis, N.; Pacifici, M.; Koyama, E. Sulfotransferase Ndst1 is needed for mandibular and TMJ development. J. Dent. Res. 2010, 89, 1111–1116. [Google Scholar] [CrossRef] [PubMed]
- Crawford, B.E.; Garner, O.B.; Bishop, J.R.; Zhang, D.Y.; Bush, K.T.; Nigam, S.K.; Esko, J.D. Loss of the heparan sulfate sulfotransferase, Ndst1, in mammary epithelial cells selectively blocks lobuloalveolar development in mice. PLoS ONE 2010, 5, e10691. [Google Scholar] [CrossRef] [PubMed]
- Grobe, K.; Inatani, M.; Pallerla, S.R.; Castagnola, J.; Yamaguchi, Y.; Esko, J.D. Cerebral hypoplasia and craniofacial defects in mice lacking heparan sulfate Ndst1 gene function. Development 2005, 132, 3777–3786. [Google Scholar] [CrossRef] [PubMed]
- Long, H.Q.; Tian, P.F.; Guan, Y.X.; Liu, L.X.; Wu, X.P.; Li, B. Expression of Ihh signaling pathway in condylar cartilage after bite-raising in adult rats. J. Mol. Histol. 2019, 50, 459–470. [Google Scholar] [CrossRef]
- Kinumatsu, T.; Shibukawa, Y.; Yasuda, T.; Nagayama, M.; Yamada, S.; Serra, R.; Pacifici, M.; Koyama, E. TMJ development and growth require primary cilia function. J. Dent. Res. 2011, 90, 988–994. [Google Scholar] [CrossRef]
- Buckland, J. Osteoarthritis: Blocking hedgehog signaling might have therapeutic potential in OA. Nat. Rev. Rheumatol. 2010, 6, 61. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bechtold, T.E.; Kurio, N.; Nah, H.-D.; Saunders, C.; Billings, P.C.; Koyama, E. The Roles of Indian Hedgehog Signaling in TMJ Formation. Int. J. Mol. Sci. 2019, 20, 6300. https://doi.org/10.3390/ijms20246300
Bechtold TE, Kurio N, Nah H-D, Saunders C, Billings PC, Koyama E. The Roles of Indian Hedgehog Signaling in TMJ Formation. International Journal of Molecular Sciences. 2019; 20(24):6300. https://doi.org/10.3390/ijms20246300
Chicago/Turabian StyleBechtold, Till E., Naito Kurio, Hyun-Duck Nah, Cheri Saunders, Paul C. Billings, and Eiki Koyama. 2019. "The Roles of Indian Hedgehog Signaling in TMJ Formation" International Journal of Molecular Sciences 20, no. 24: 6300. https://doi.org/10.3390/ijms20246300
APA StyleBechtold, T. E., Kurio, N., Nah, H.-D., Saunders, C., Billings, P. C., & Koyama, E. (2019). The Roles of Indian Hedgehog Signaling in TMJ Formation. International Journal of Molecular Sciences, 20(24), 6300. https://doi.org/10.3390/ijms20246300