Hyperacetylation of Cardiac Mitochondrial Proteins Is Associated with Metabolic Impairment and Sirtuin Downregulation after Chronic Total Body Irradiation of ApoE -/- Mice




Abstract
:1. Introduction
2. Results
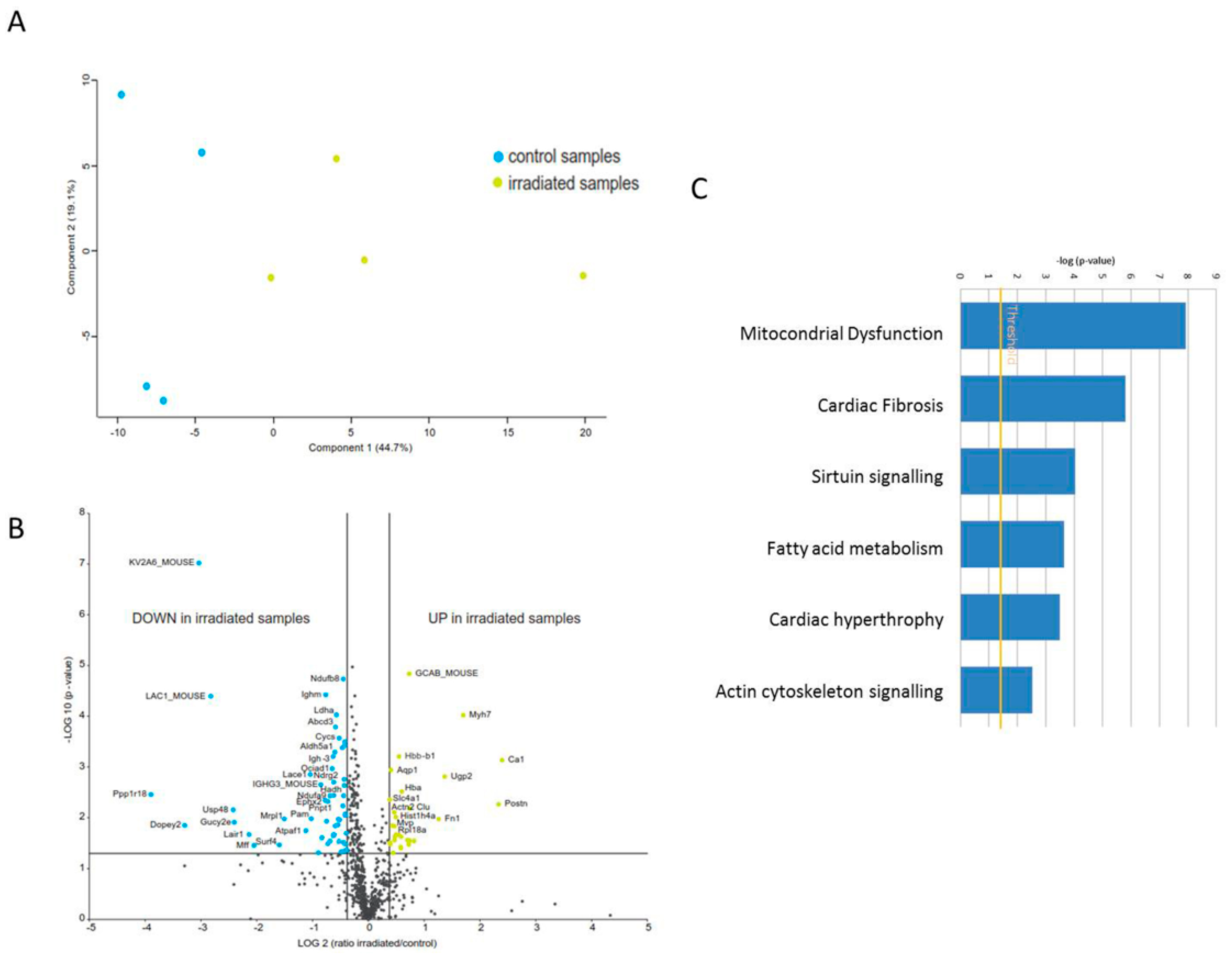
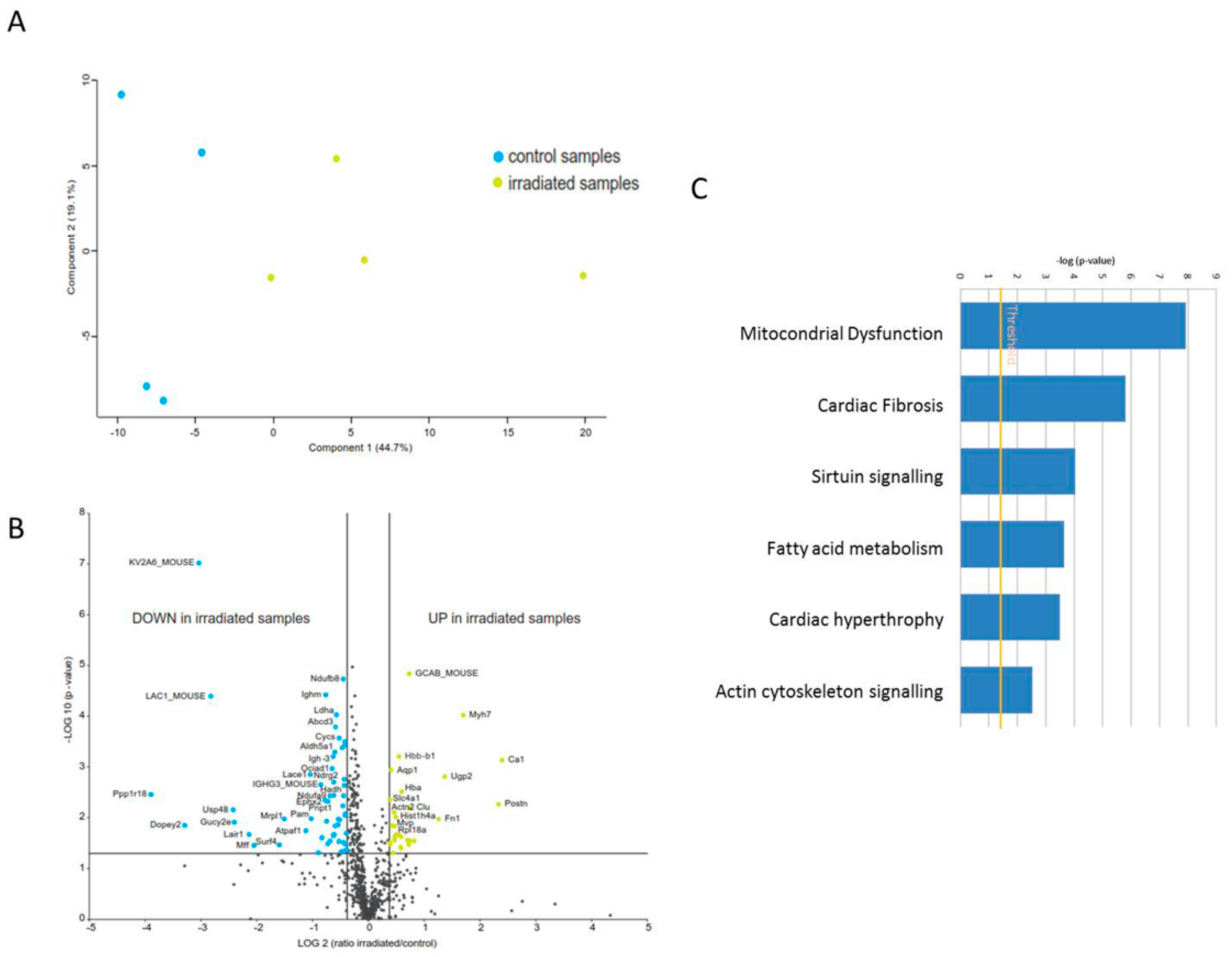
2.1. The Cardiac Mitochondrial Proteome Is Altered after Chronic Irradiation
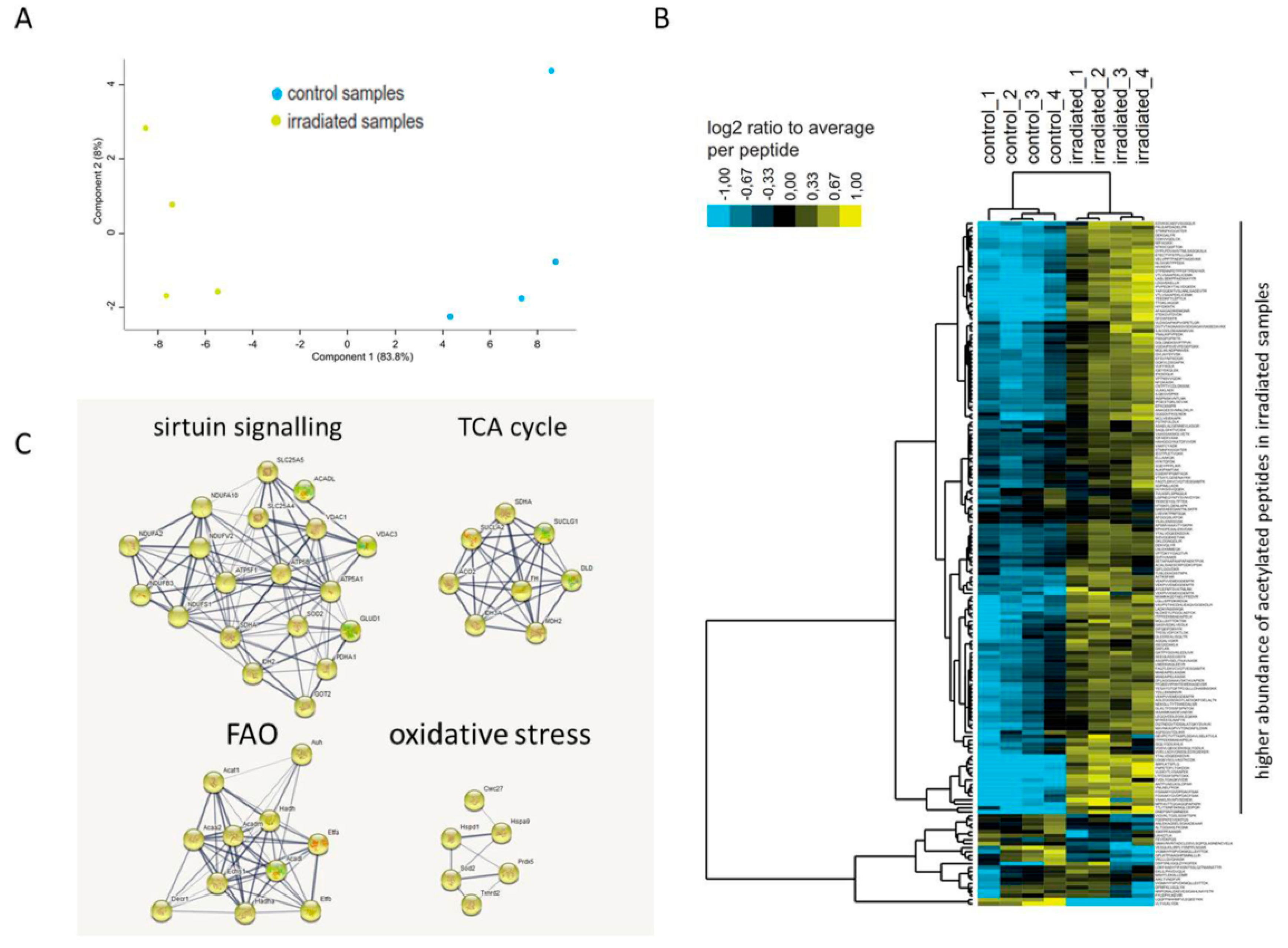
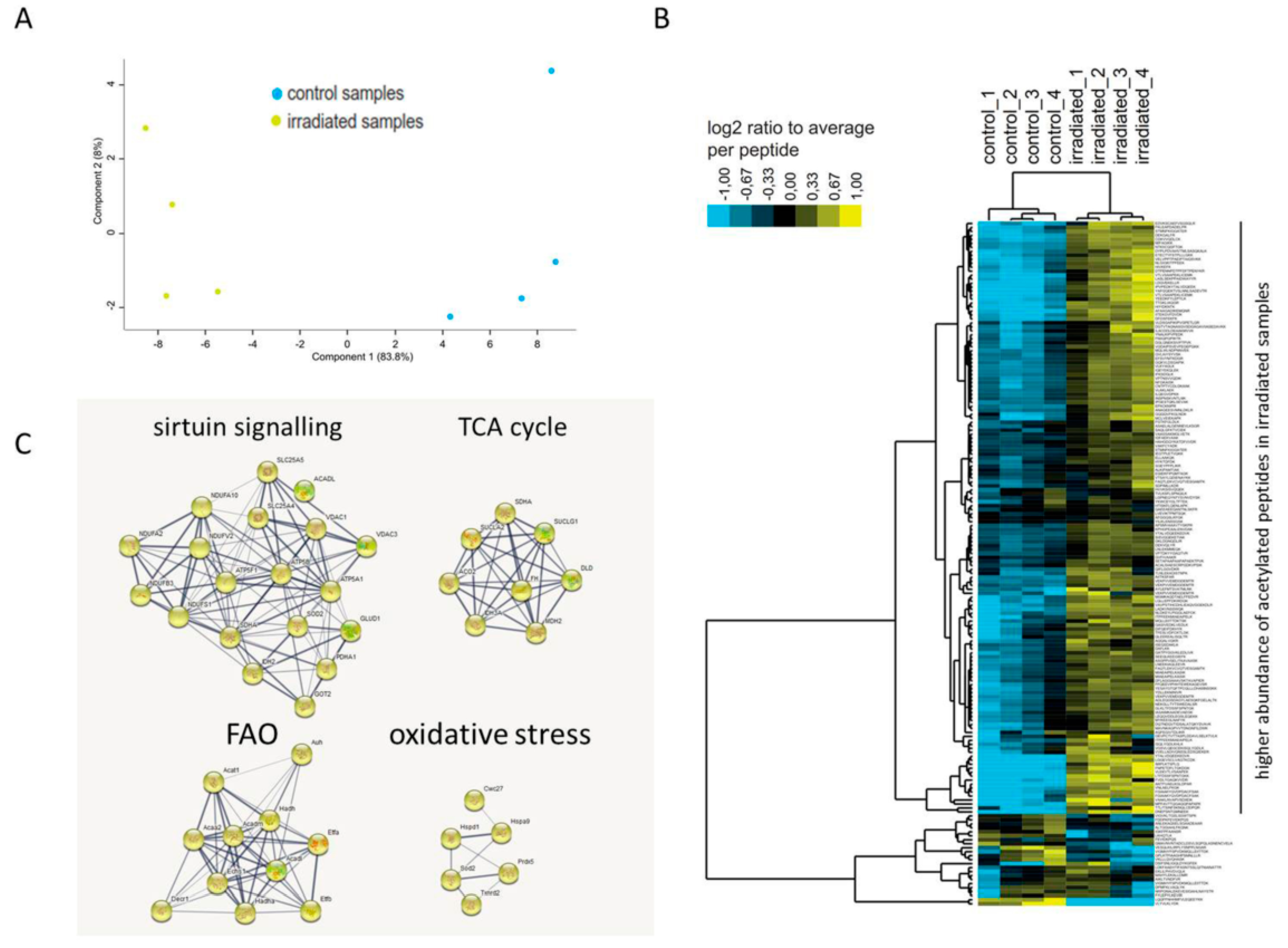
2.2. Long-Term Chronic Irradiation Causes Mitochondrial Protein Hyperacetylation
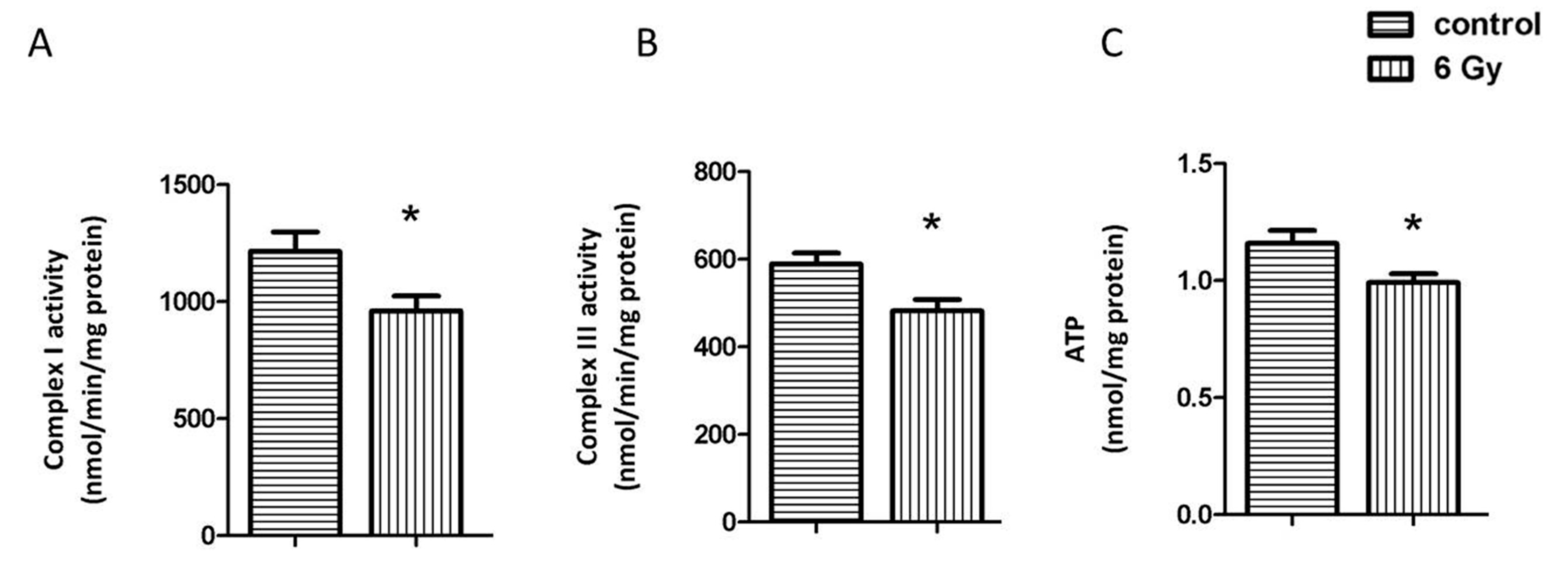
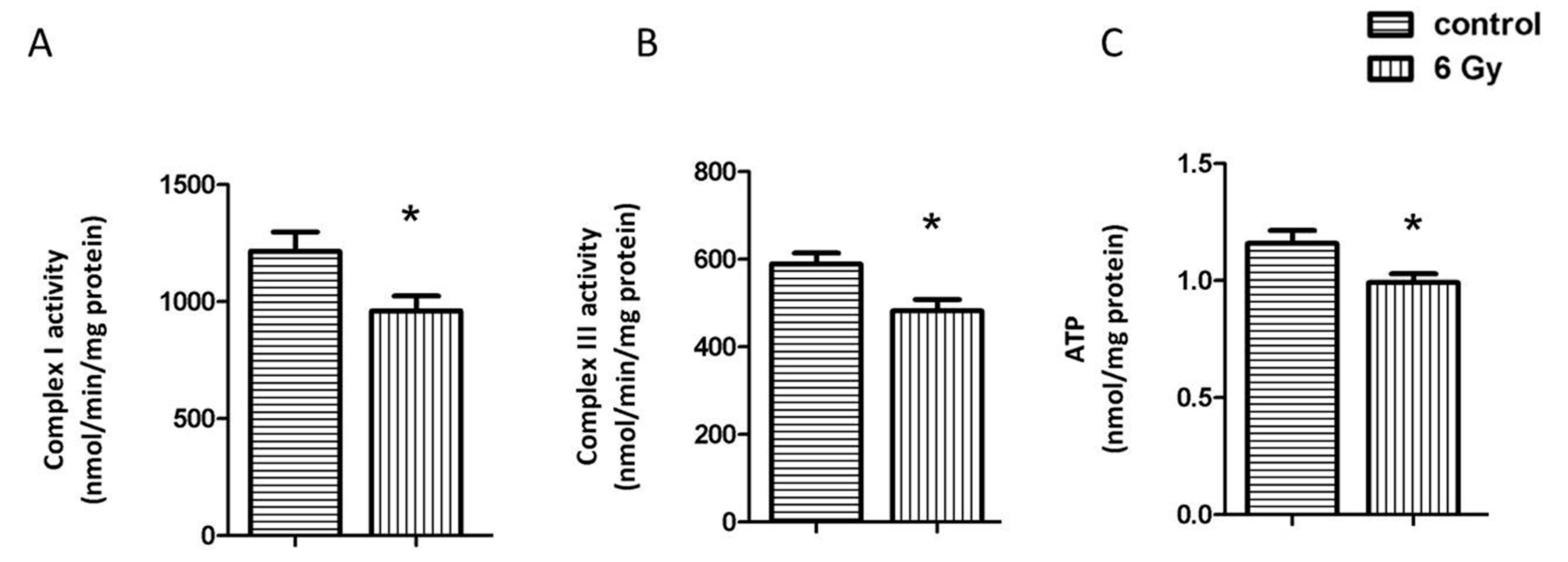
2.3. Chronic Irradiation Decreases the Activity of Respiratory Complexes I and III and Cardiac ATP Level
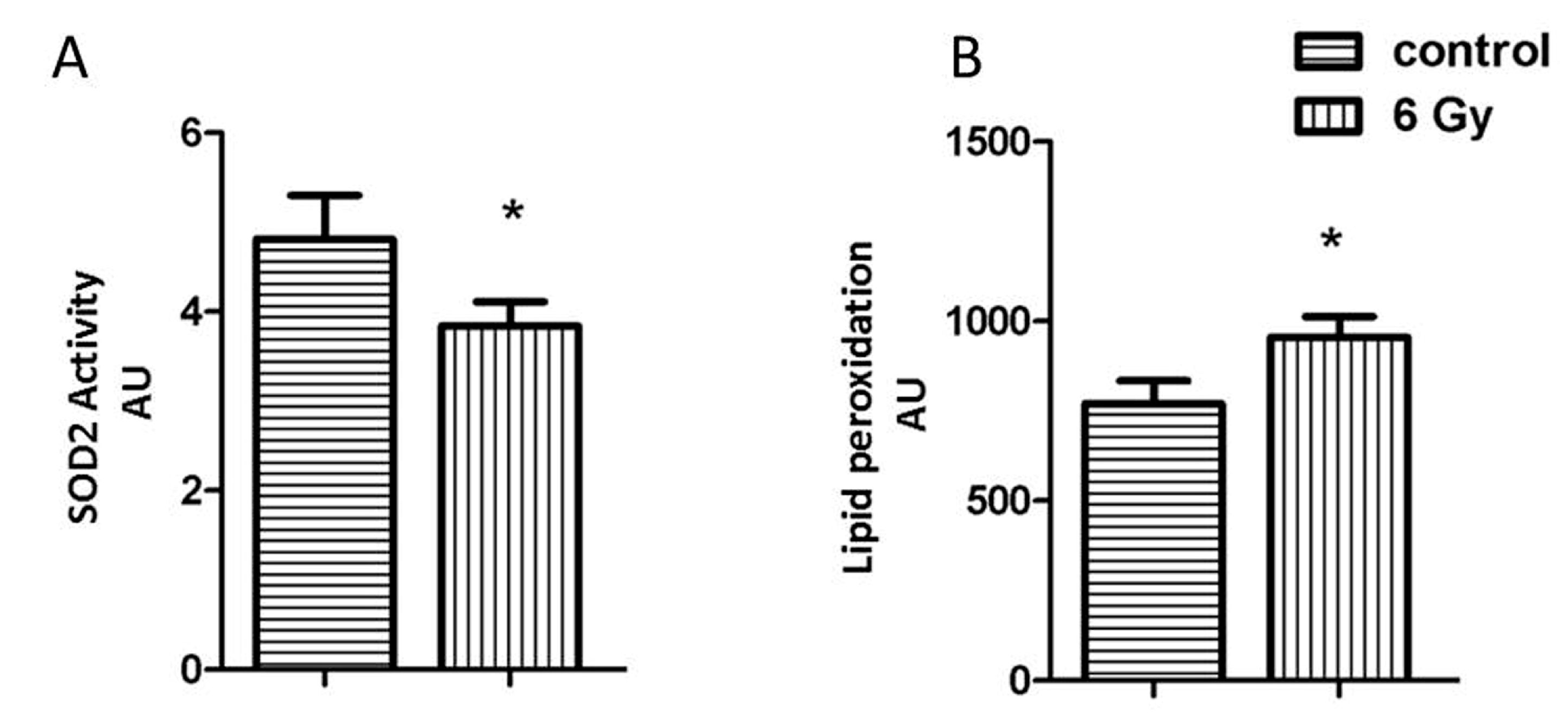
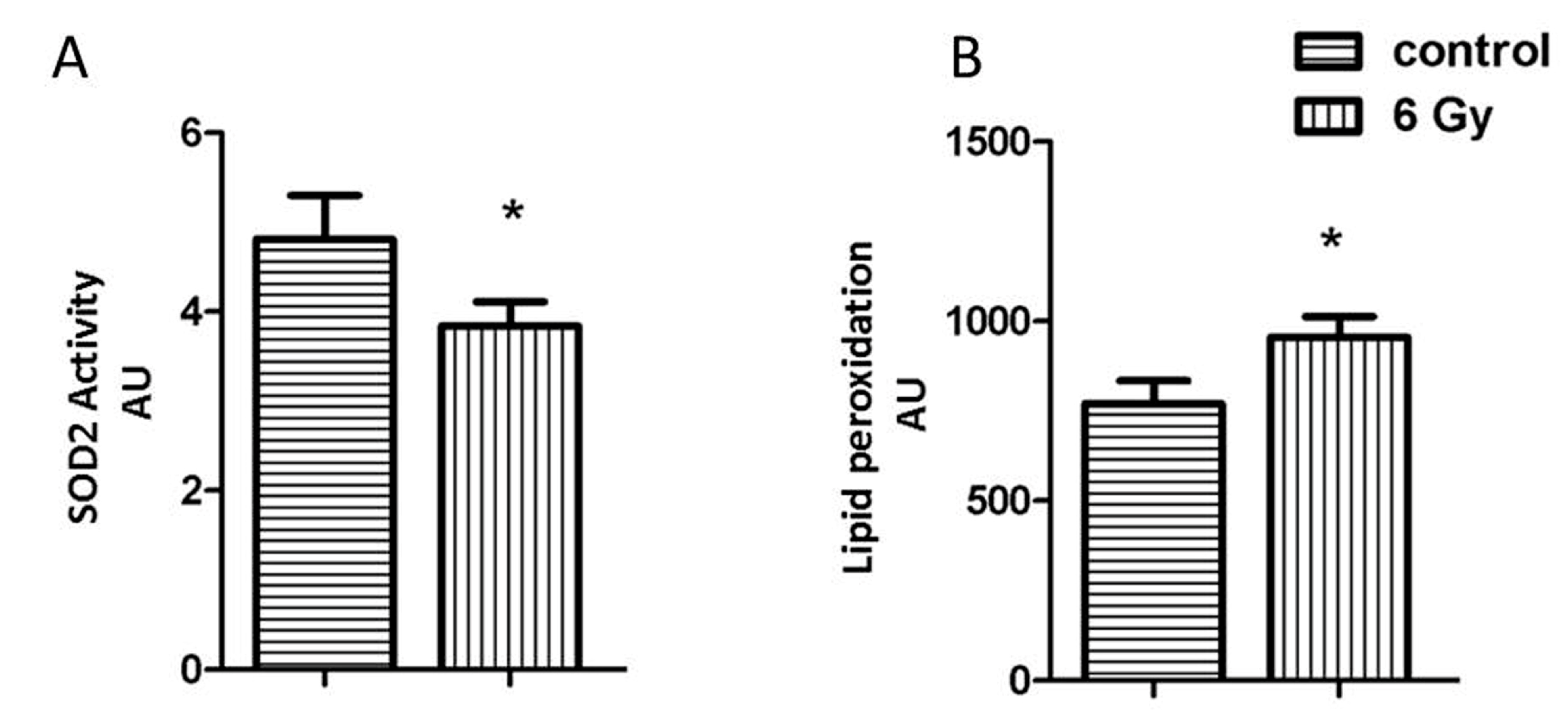
2.4. Acetylation Impairs the Oxidative Stress Response in Irradiated Mitochondria
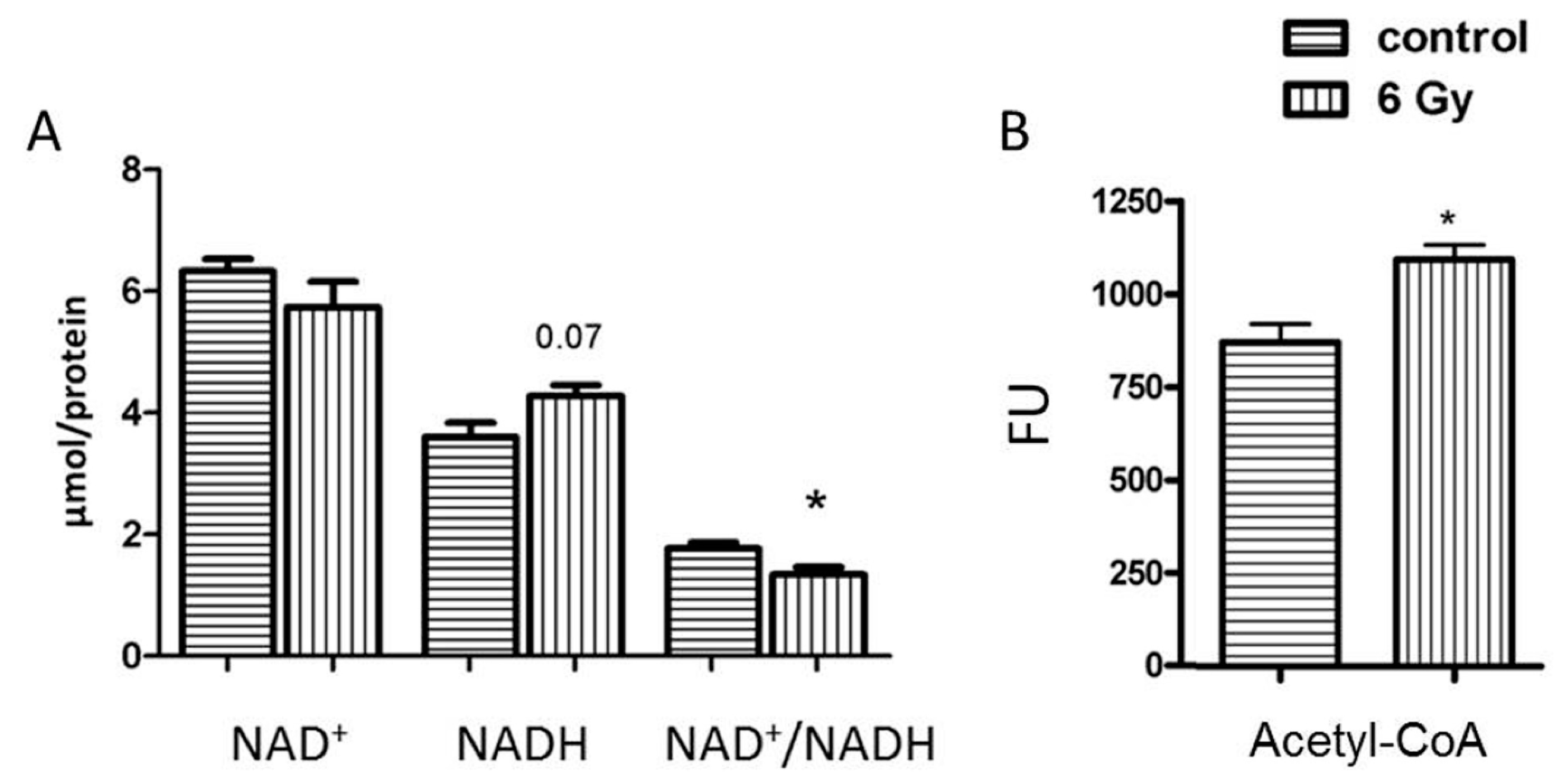
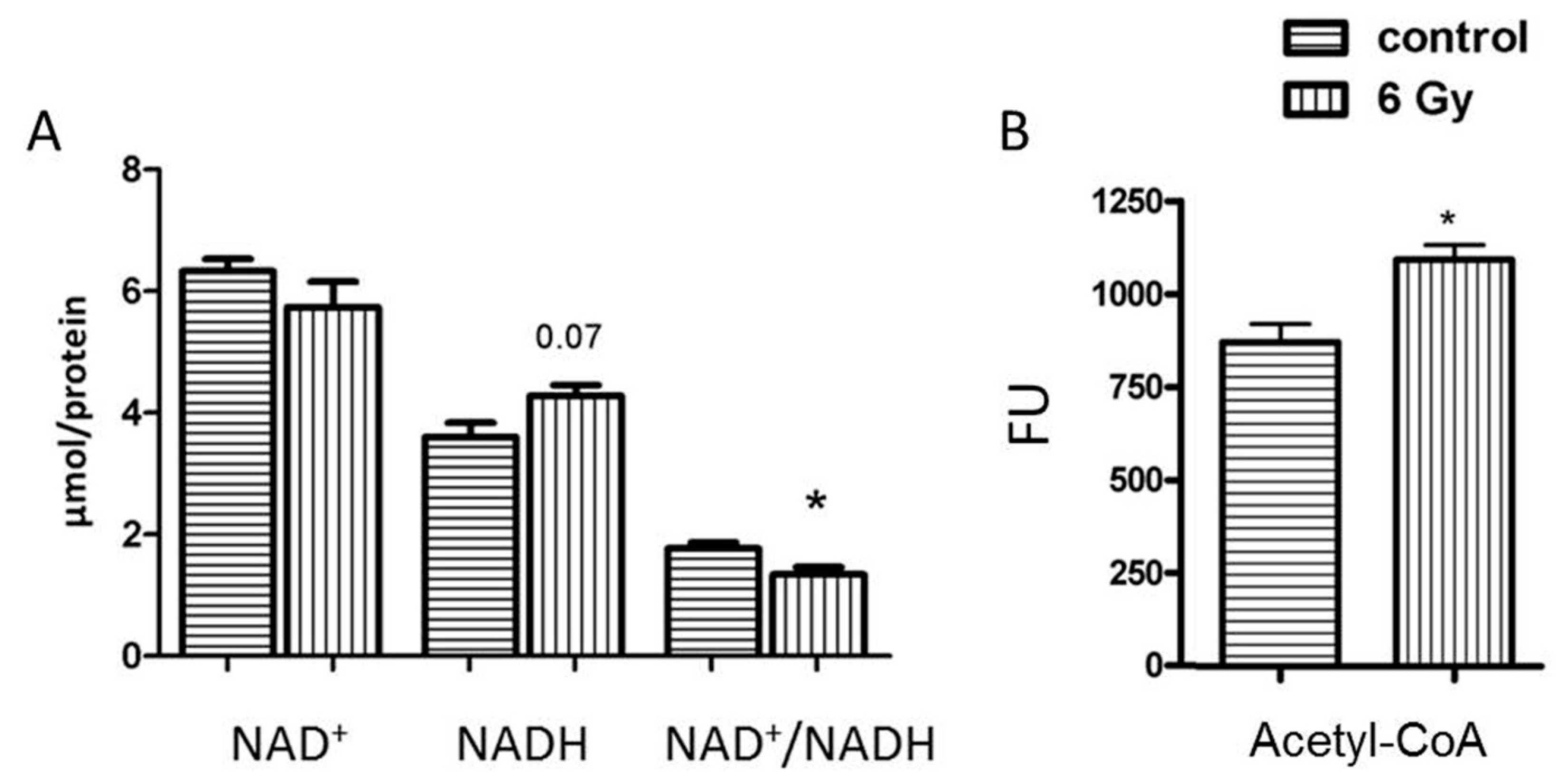
2.5. Mitochondrial NAD+/NADH Level Is Reduced after Chronic Irradiation
2.6. The Level of Acetyl-CoA Is Enhanced in Irradiated Mitochondria
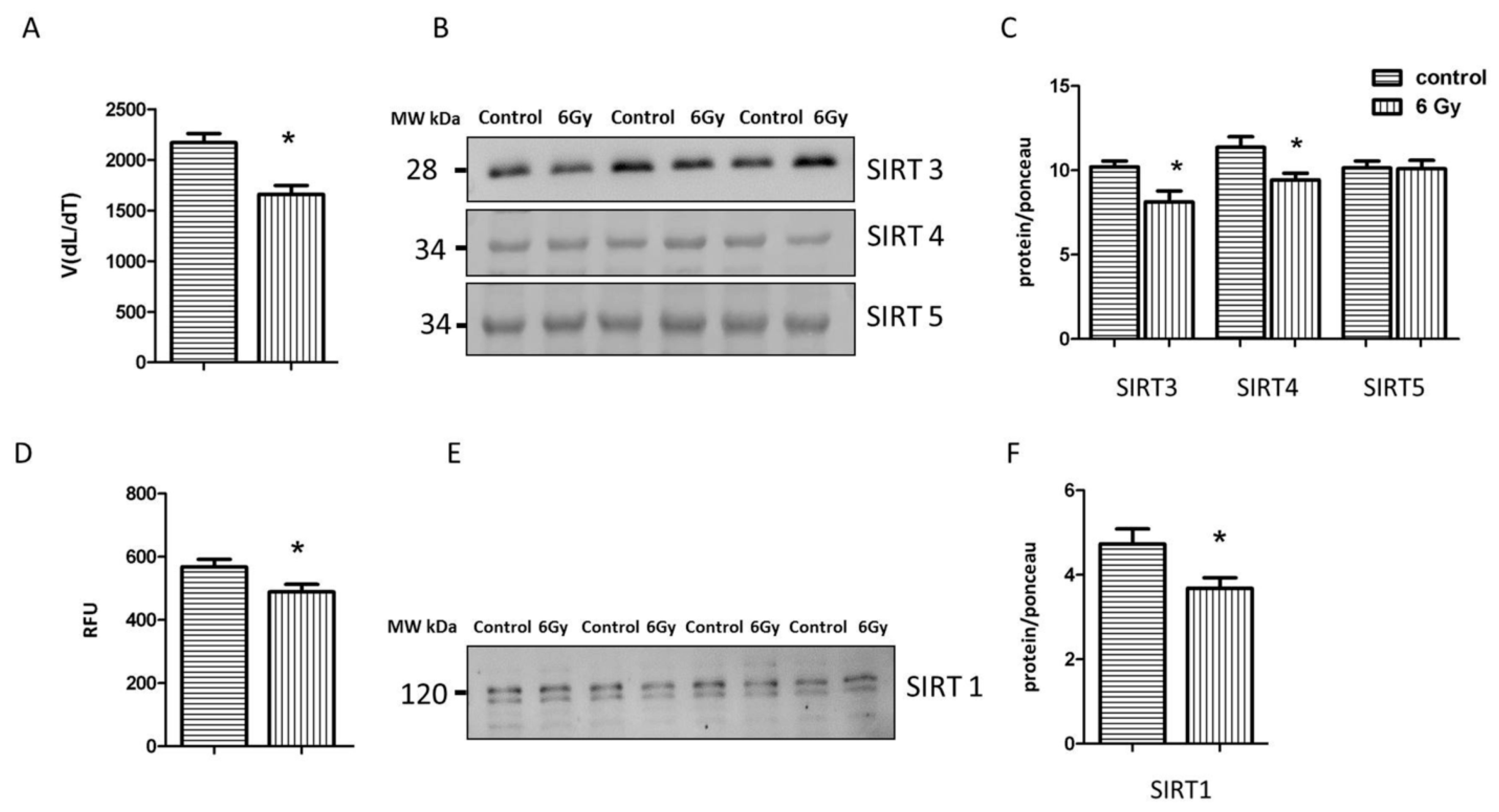
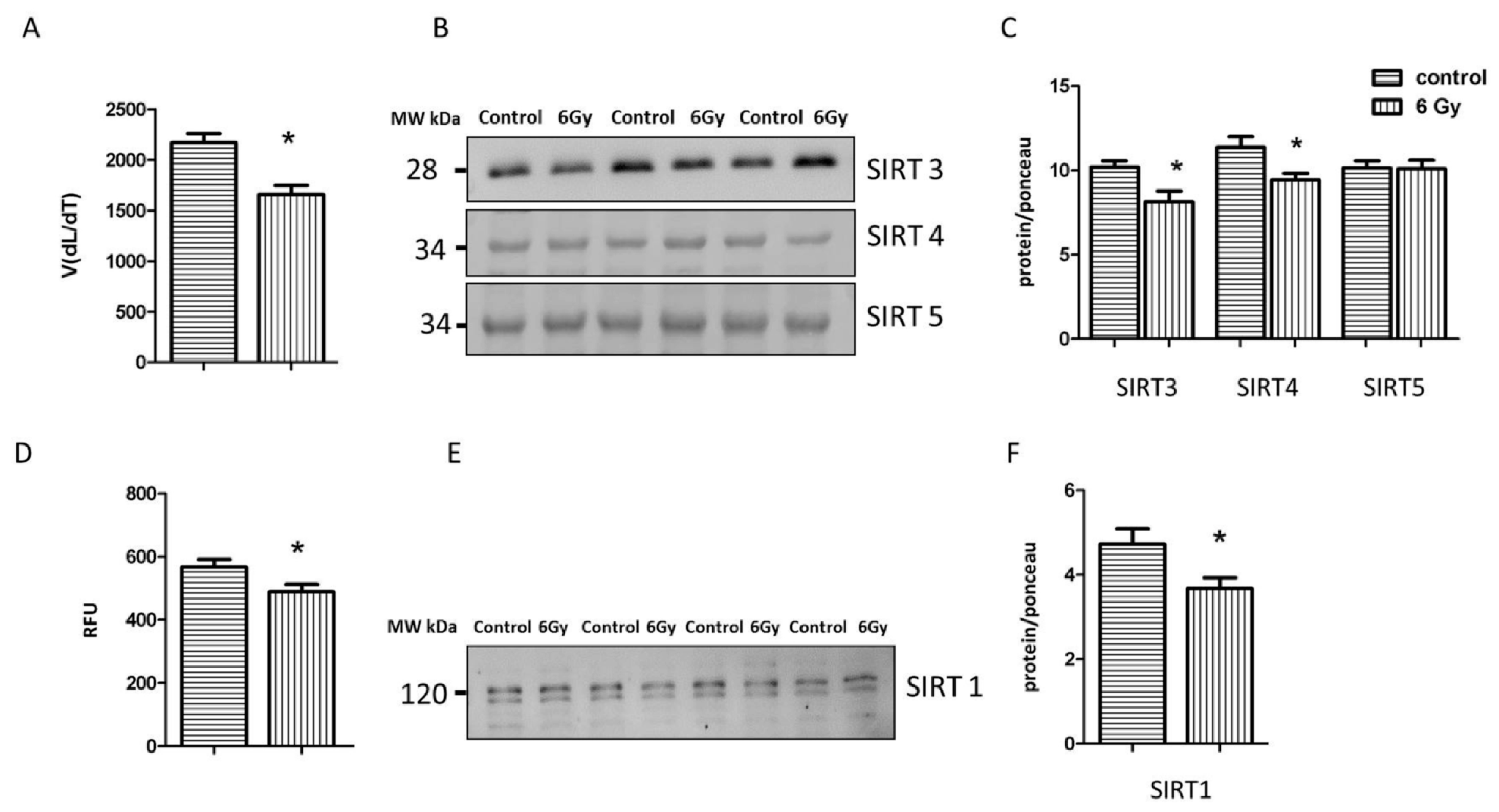
2.7. Irradiation Negatively Influences the Activity and Expression of Sirtuins
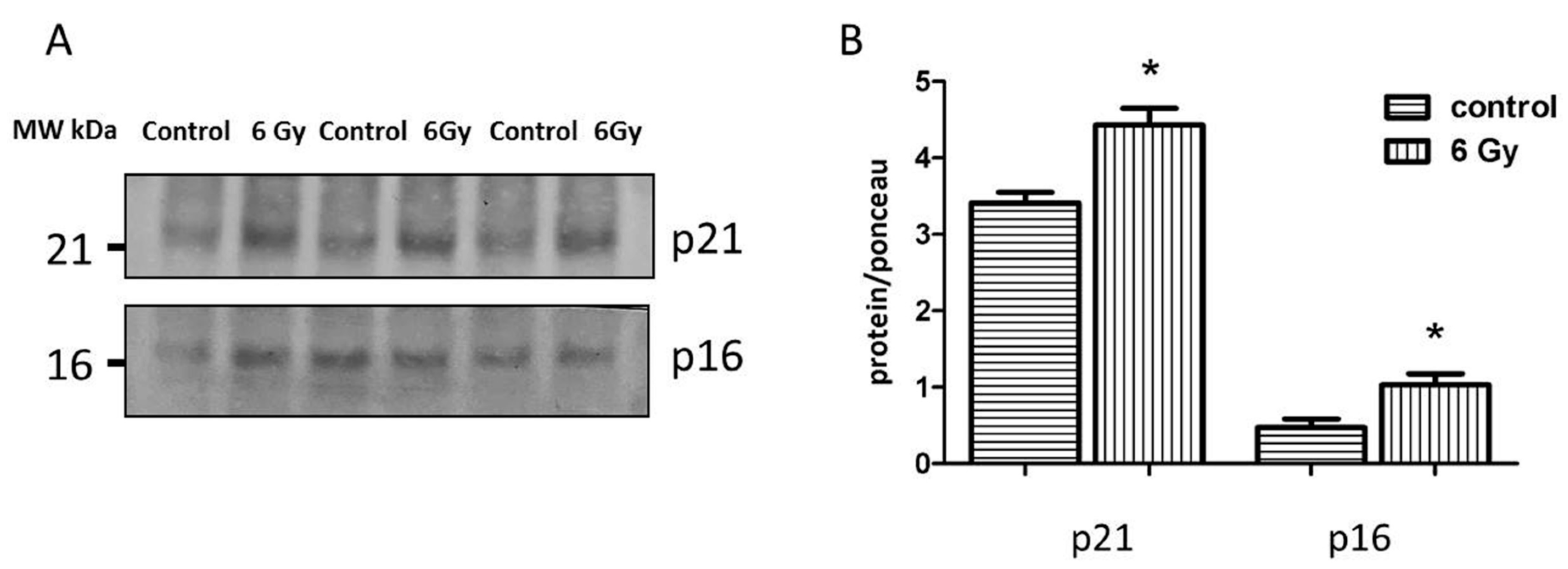
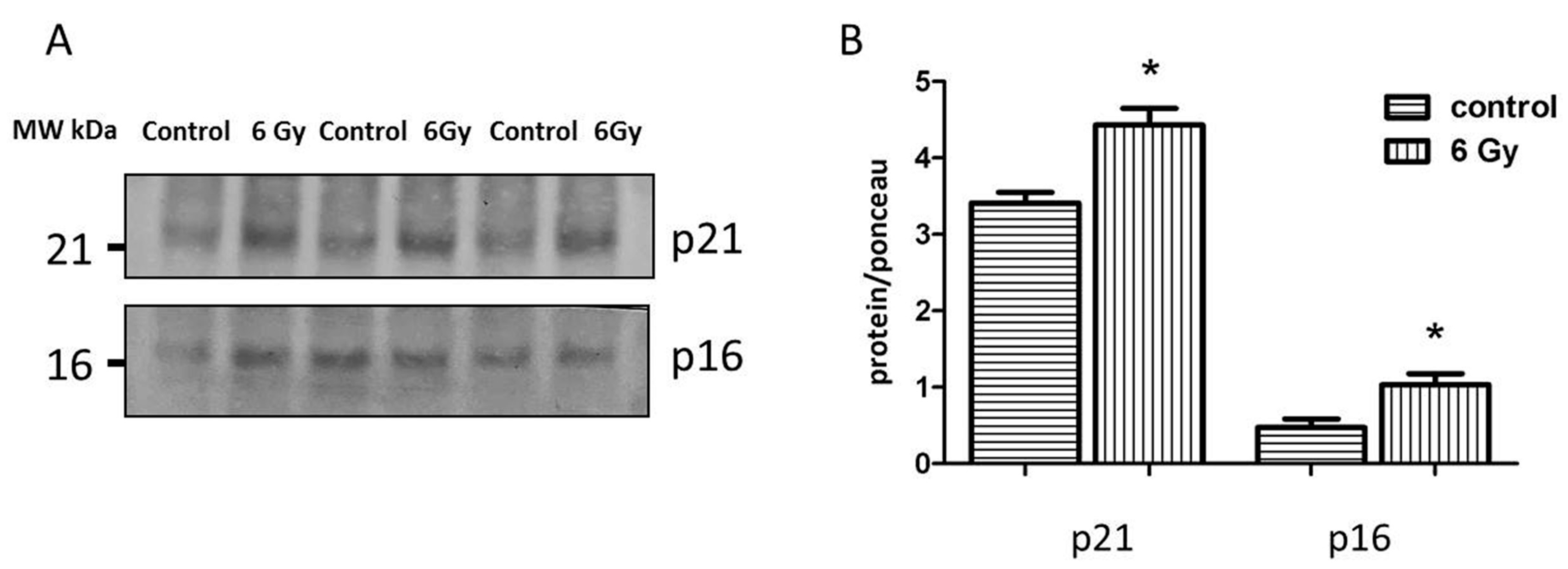
2.8. Reduced Sirtuin Activities Contribute to Cardiac Senescence
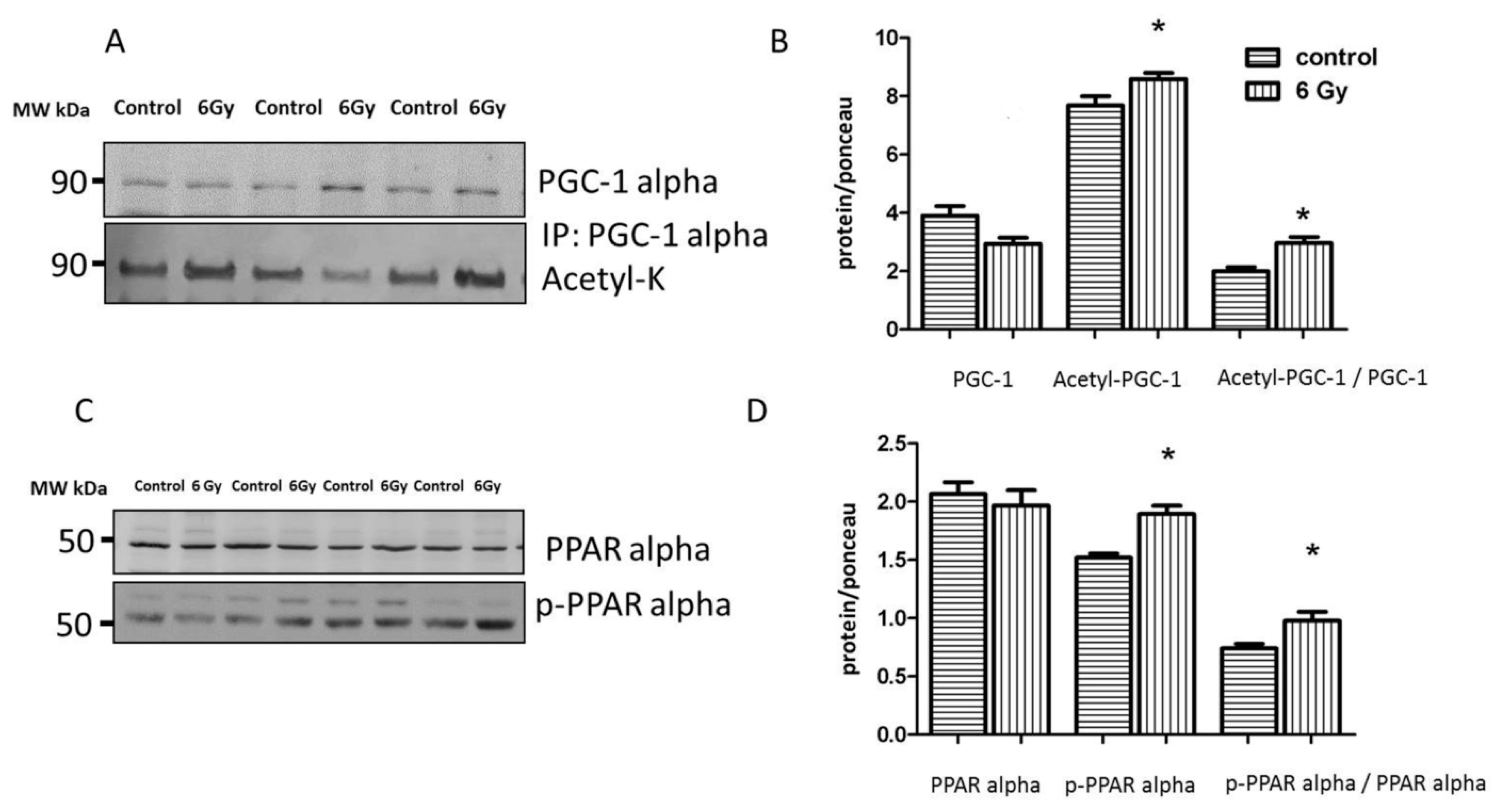
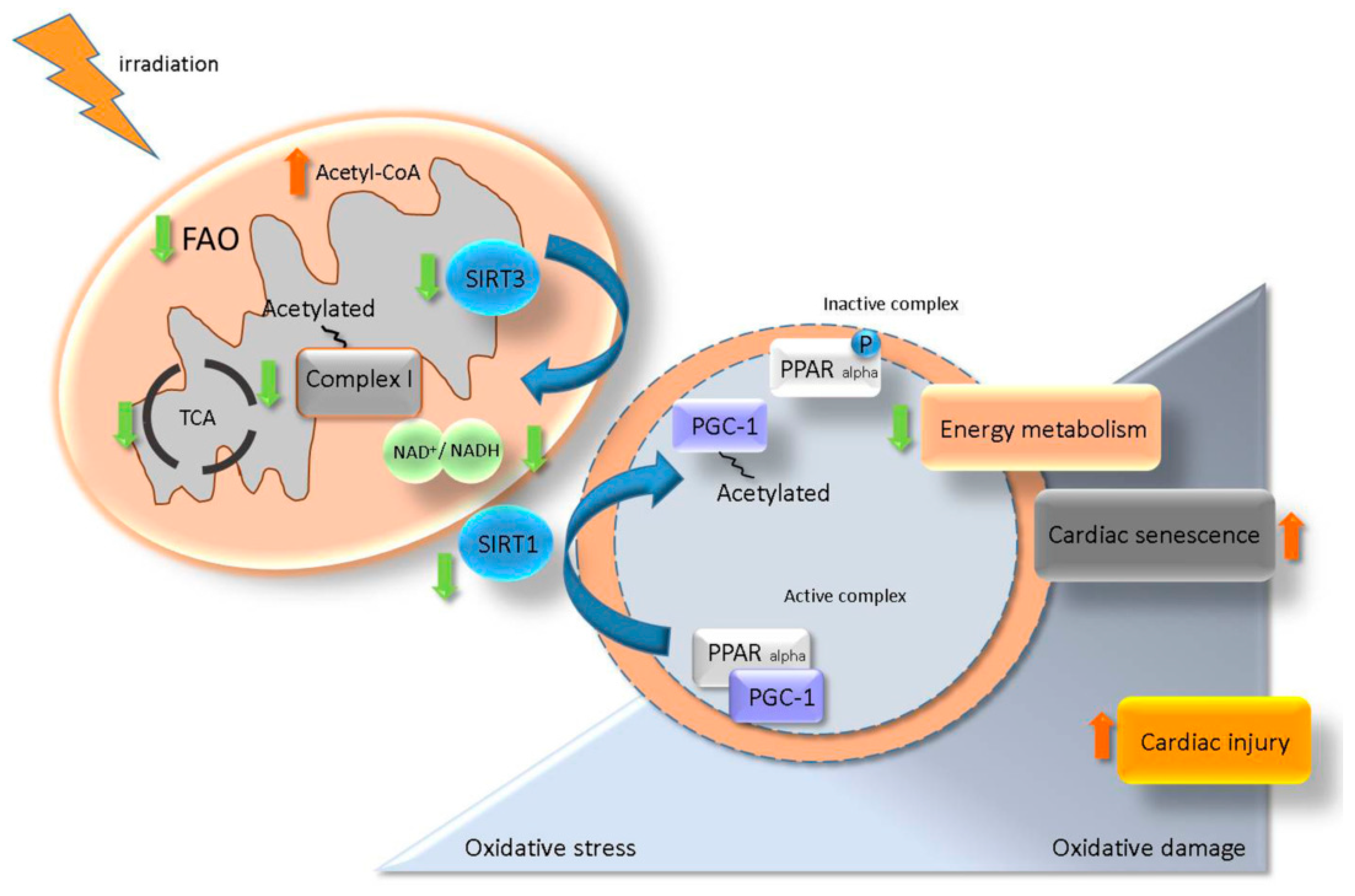
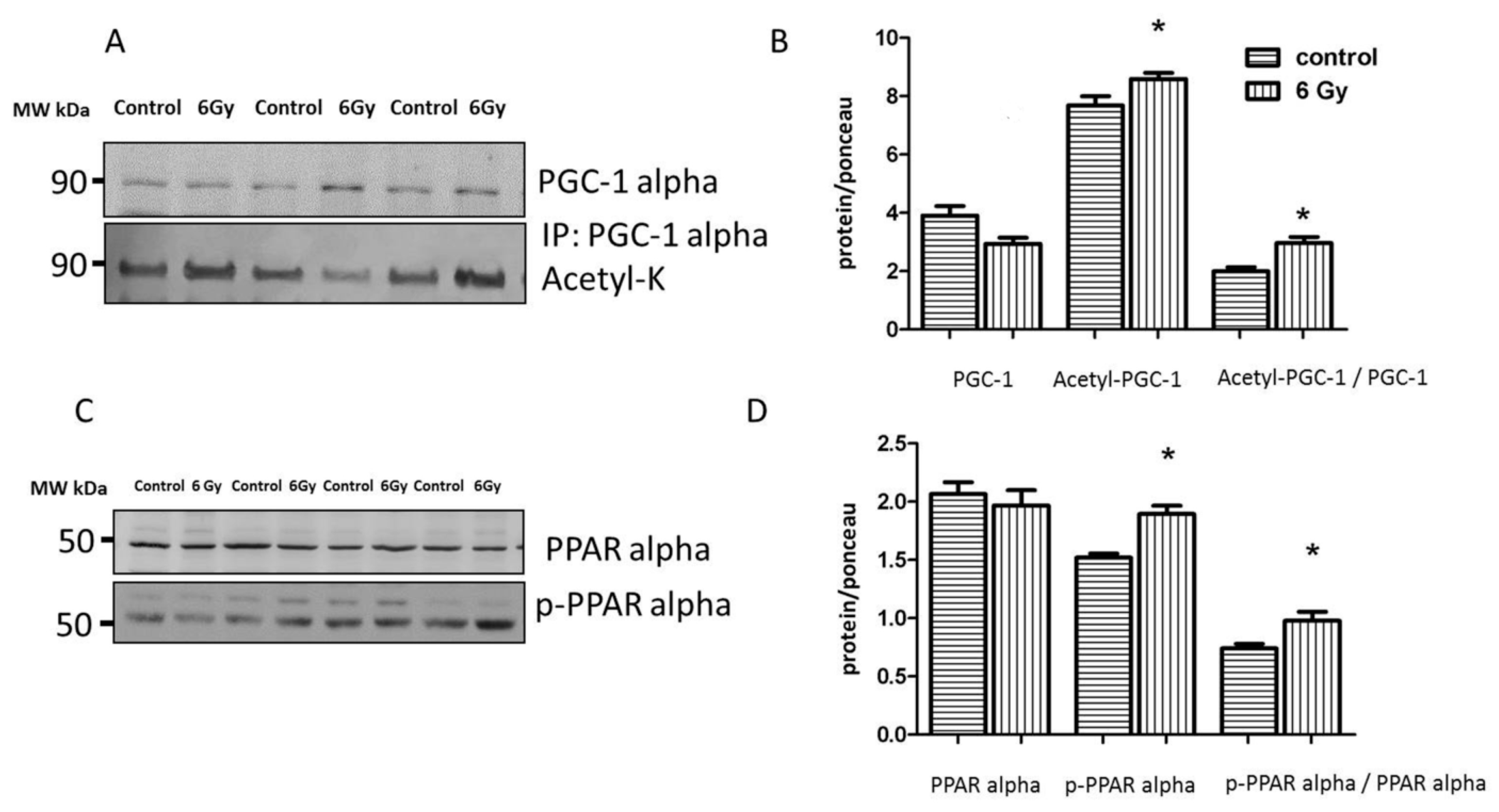
2.9. Mitochondrial Acetylation Is Involved in Radiation-Induced Cardiac Metabolism Alteration
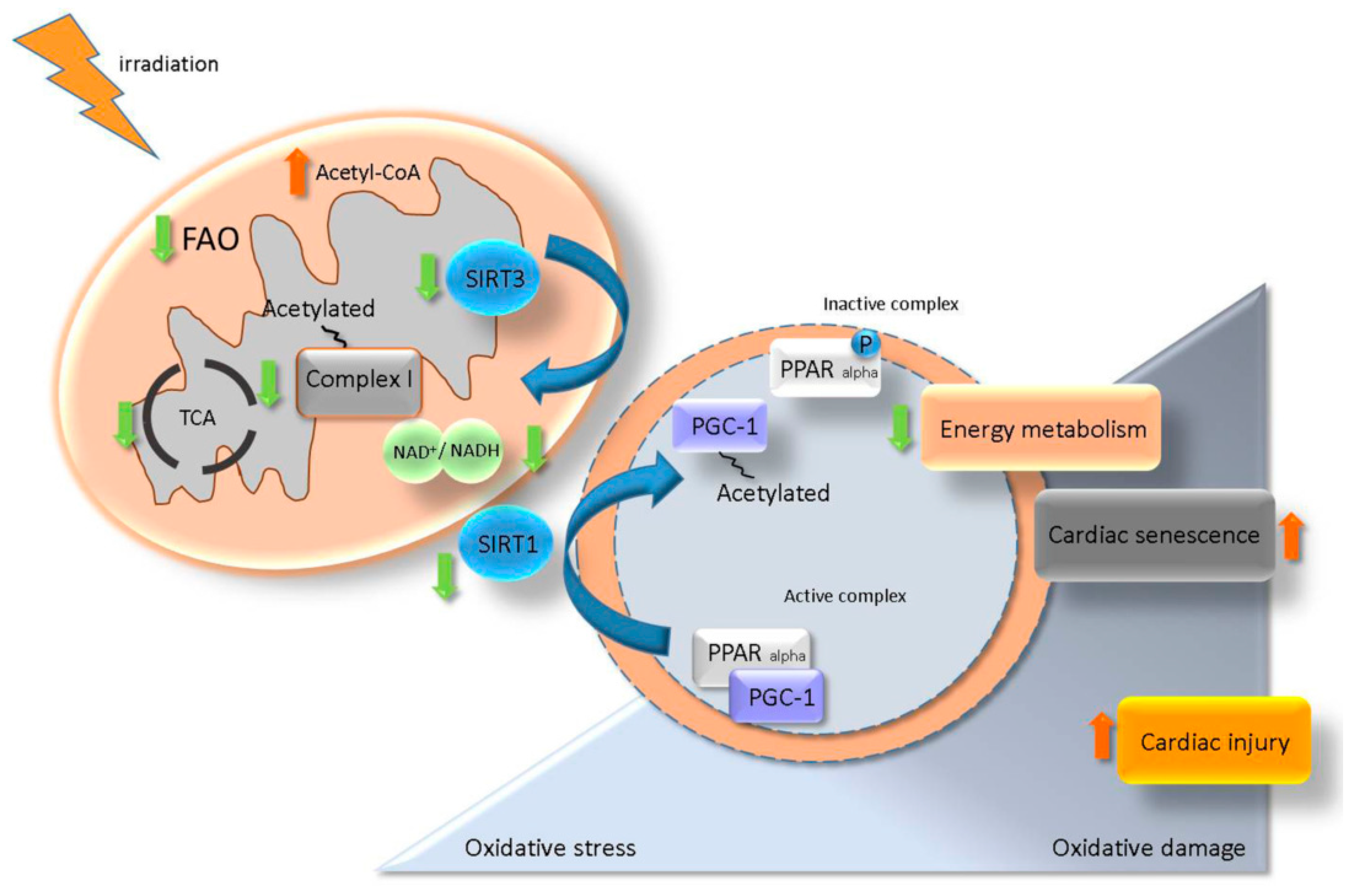
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Irradiation
4.3. Cardiac Mitochondria Isolation
4.4. Proteome Analysis
4.5. Acetylome Analysis
4.6. Principal Components Analysis
4.7. Heatmap Generation
4.8. Protein-Protein Interaction and Signalling Network
4.9. Sirtuin Activity Assay
4.10. Complex I and Complex III Activity
4.11. ATP Assay
4.12. Acetyl-CoA Assay
4.13. NAD+/ NADH Assay
4.14. SOD2 Activity Assay
4.15. Lipid Peroxidation Assay
4.16. Immunoblot Analysis
4.17. Statistical Analysis
4.18. Data Availability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Little, M.P. Radiation and circulatory disease. Mutat. Res. 2016, 770, 299–318. [Google Scholar] [CrossRef] [Green Version]
- Little, M.P.; Azizova, T.V.; Bazyka, D.; Bouffler, S.D.; Cardis, E.; Chekin, S.; Chumak, V.V.; Cucinotta, F.A.; de Vathaire, F.; Hall, P.; et al. Systematic review and meta-analysis of circulatory disease from exposure to low-level ionizing radiation and estimates of potential population mortality risks. Environ. Health Perspect. 2012, 120, 1503–1511. [Google Scholar] [CrossRef]
- Tapio, S. Pathology and biology of radiation-induced cardiac disease. J. Rad. Res. 2016, 57, 439–448. [Google Scholar] [CrossRef]
- Swerdlow, A.J.; Higgins, C.D.; Smith, P.; Cunningham, D.; Hancock, B.W.; Horwich, A.; Hoskin, P.J.; Lister, A.; Radford, J.A.; Rohatiner, A.Z.; et al. Myocardial infarction mortality risk after treatment for Hodgkin disease: A collaborative British cohort study. J. Natl. Cancer Inst. 2007, 99, 206–214. [Google Scholar] [CrossRef]
- Darby, S.C.; Ewertz, M.; McGale, P.; Bennet, A.M.; Blom-Goldman, U.; Bronnum, D.; Correa, C.; Cutter, D.; Gagliardi, G.; Gigante, B.; et al. Risk of ischemic heart disease in women after radiotherapy for breast cancer. N. Engl. J. Med. 2013, 368, 987–998. [Google Scholar] [CrossRef]
- Carr, Z.A.; Land, C.E.; Kleinerman, R.A.; Weinstock, R.W.; Stovall, M.; Griem, M.L.; Mabuchi, K. Coronary heart disease after radiotherapy for peptic ulcer disease. Int. J. Radiat. Oncol. Biol. Phys. 2005, 61, 842–850. [Google Scholar] [CrossRef]
- Tukenova, M.; Guibout, C.; Oberlin, O.; Doyon, F.; Mousannif, A.; Haddy, N.; Guerin, S.; Pacquement, H.; Aouba, A.; Hawkins, M.; et al. Role of cancer treatment in long-term overall and cardiovascular mortality after childhood cancer. J. Clin. Oncol. 2010, 28, 1308–1315. [Google Scholar] [CrossRef]
- Azizova, T.V.; Muirhead, C.R.; Moseeva, M.B.; Grigoryeva, E.S.; Vlasenko, E.V.; Hunter, N.; Haylock, R.G.; O’Hagan, J.A. Ischemic heart disease in nuclear workers first employed at the Mayak PA in 1948-1972. Health Phys. 2012, 103, 3–14. [Google Scholar] [CrossRef]
- Azizova, T.V.; Grigoryeva, E.S.; Haylock, R.G.; Pikulina, M.V.; Moseeva, M.B. Ischaemic heart disease incidence and mortality in an extended cohort of Mayak workers first employed in 1948-1982. Br. J. Radiol. 2015, 88, 20150169. [Google Scholar] [CrossRef]
- Azizova, T.V.; Muirhead, C.R.; Druzhinina, M.B.; Grigoryeva, E.S.; Vlasenko, E.V.; Sumina, M.V.; O’Hagan, J.A.; Zhang, W.; Haylock, R.G.; Hunter, N. Cardiovascular diseases in the cohort of workers first employed at Mayak PA in 1948-1958. Radiat. Res. 2010, 174, 155–168. [Google Scholar] [CrossRef]
- Gillies, M.; Richardson, D.B.; Cardis, E.; Daniels, R.D.; O’Hagan, J.A.; Haylock, R.; Laurier, D.; Leuraud, K.; Moissonnier, M.; Schubauer-Berigan, M.K.; et al. Mortality from Circulatory Diseases and other Non-Cancer Outcomes among Nuclear Workers in France, the United Kingdom and the United States (INWORKS). Radiat. Res. 2017, 188, 276–290. [Google Scholar] [CrossRef]
- Zhang, W.; Haylock, R.G.E.; Gillies, M.; Hunter, N. Mortality from heart diseases following occupational radiation exposure: Analysis of the National Registry for Radiation Workers (NRRW) in the United Kingdom. J. Radiol. Prot. 2019, 39, 327–353. [Google Scholar] [CrossRef]
- Ruhm, W.; Azizova, T.; Bouffler, S.; Cullings, H.M.; Grosche, B.; Little, M.P.; Shore, R.S.; Walsh, L.; Woloschak, G.E. Typical doses and dose rates in studies pertinent to radiation risk inference at low doses and low dose rates. J. Radiat. Res. 2018, 59, ii1–ii10. [Google Scholar] [CrossRef]
- Hunter, N.; Kuznetsova, I.S.; Labutina, E.V.; Harrison, J.D. Solid cancer incidence other than lung, liver and bone in Mayak workers: 1948-2004. Br. J. Cancer 2013, 109, 1989–1996. [Google Scholar] [CrossRef]
- Azimzadeh, O.; Sievert, W.; Sarioglu, H.; Yentrapalli, R.; Barjaktarovic, Z.; Sriharshan, A.; Ueffing, M.; Janik, D.; Aichler, M.; Atkinson, M.J.; et al. PPAR Alpha: A Novel Radiation Target in Locally Exposed Mus musculus Heart Revealed by Quantitative Proteomics. J. Proteome Res. 2013, 12, 2700–2714. [Google Scholar] [CrossRef]
- Barjaktarovic, Z.; Shyla, A.; Azimzadeh, O.; Schulz, S.; Haagen, J.; Dorr, W.; Sarioglu, H.; Atkinson, M.J.; Zischka, H.; Tapio, S. Ionising radiation induces persistent alterations in the cardiac mitochondrial function of C57BL/6 mice 40 weeks after local heart exposure. Radiother. Oncol. 2013, 106, 404–410. [Google Scholar] [CrossRef]
- Barjaktarovic, Z.; Schmaltz, D.; Shyla, A.; Azimzadeh, O.; Schulz, S.; Haagen, J.; Dorr, W.; Sarioglu, H.; Schafer, A.; Atkinson, M.J.; et al. Radiation-induced signaling results in mitochondrial impairment in mouse heart at 4 weeks after exposure to X-rays. PLoS ONE 2011, 6, e27811. [Google Scholar] [CrossRef]
- Azimzadeh, O.; Azizova, T.; Merl-Pham, J.; Subramanian, V.; Bakshi, M.V.; Moseeva, M.; Zubkova, O.; Hauck, S.M.; Anastasov, N.; Atkinson, M.J.; et al. A dose-dependent perturbation in cardiac energy metabolism is linked to radiation-induced ischemic heart disease in Mayak nuclear workers. Oncotarget 2017, 8, 9067–9078. [Google Scholar] [CrossRef]
- Hebert, A.S.; Dittenhafer-Reed, K.E.; Yu, W.; Bailey, D.J.; Selen, E.S.; Boersma, M.D.; Carson, J.J.; Tonelli, M.; Balloon, A.J.; Higbee, A.J.; et al. Calorie restriction and SIRT3 trigger global reprogramming of the mitochondrial protein acetylome. Mol. Cell 2013, 49, 186–199. [Google Scholar] [CrossRef]
- Hirschey, M.D.; Shimazu, T.; Goetzman, E.; Jing, E.; Schwer, B.; Lombard, D.B.; Grueter, C.A.; Harris, C.; Biddinger, S.; Ilkayeva, O.R.; et al. SIRT3 regulates mitochondrial fatty-acid oxidation by reversible enzyme deacetylation. Nature 2010, 464, 121–125. [Google Scholar] [CrossRef] [Green Version]
- Hirschey, M.D.; Shimazu, T.; Jing, E.; Grueter, C.A.; Collins, A.M.; Aouizerat, B.; Stancakova, A.; Goetzman, E.; Lam, M.M.; Schwer, B.; et al. SIRT3 deficiency and mitochondrial protein hyperacetylation accelerate the development of the metabolic syndrome. Mol. Cell 2011, 44, 177–190. [Google Scholar] [CrossRef]
- Parodi-Rullan, R.M.; Chapa-Dubocq, X.R.; Javadov, S. Acetylation of Mitochondrial Proteins in the Heart: The Role of SIRT3. Front. Physiol. 2018, 9, 1094. [Google Scholar] [CrossRef] [Green Version]
- Fukushima, A.; Lopaschuk, G.D. Acetylation control of cardiac fatty acid beta-oxidation and energy metabolism in obesity, diabetes, and heart failure. Biochim. Biophys. Acta 2016, 1862, 2211–2220. [Google Scholar] [CrossRef]
- Haigis, M.C.; Mostoslavsky, R.; Haigis, K.M.; Fahie, K.; Christodoulou, D.C.; Murphy, A.J.; Valenzuela, D.M.; Yancopoulos, G.D.; Karow, M.; Blander, G.; et al. SIRT4 inhibits glutamate dehydrogenase and opposes the effects of calorie restriction in pancreatic beta cells. Cell 2006, 126, 941–954. [Google Scholar] [CrossRef]
- Anderson, K.A.; Huynh, F.K.; Fisher-Wellman, K.; Stuart, J.D.; Peterson, B.S.; Douros, J.D.; Wagner, G.R.; Thompson, J.W.; Madsen, A.S.; Green, M.F.; et al. SIRT4 Is a Lysine Deacylase that Controls Leucine Metabolism and Insulin Secretion. Cell Metab. 2017, 25, 838–855. [Google Scholar] [CrossRef]
- Frye, R.A. Phylogenetic classification of prokaryotic and eukaryotic Sir2-like proteins. Biochem. Biophys. Res. Commun. 2000, 273, 793–798. [Google Scholar] [CrossRef]
- Saunders, L.R.; Verdin, E. Sirtuins: Critical regulators at the crossroads between cancer and aging. Oncogene 2007, 26, 5489–5504. [Google Scholar] [CrossRef]
- Nemoto, S.; Fergusson, M.M.; Finkel, T. SIRT1 functionally interacts with the metabolic regulator and transcriptional coactivator PGC-1{alpha}. J. Biol. Chem. 2005, 280, 16456–16460. [Google Scholar] [CrossRef]
- Vega, R.B.; Huss, J.M.; Kelly, D.P. The coactivator PGC-1 cooperates with peroxisome proliferator-activated receptor alpha in transcriptional control of nuclear genes encoding mitochondrial fatty acid oxidation enzymes. Mol. Cell. Biol. 2000, 20, 1868–1876. [Google Scholar] [CrossRef]
- Canto, C.; Gerhart-Hines, Z.; Feige, J.N.; Lagouge, M.; Noriega, L.; Milne, J.C.; Elliott, P.J.; Puigserver, P.; Auwerx, J. AMPK regulates energy expenditure by modulating NAD+ metabolism and SIRT1 activity. Nature 2009, 458, 1056–1060. [Google Scholar] [CrossRef]
- Kong, X.; Wang, R.; Xue, Y.; Liu, X.; Zhang, H.; Chen, Y.; Fang, F.; Chang, Y. Sirtuin 3, a new target of PGC-1alpha, plays an important role in the suppression of ROS and mitochondrial biogenesis. PLoS ONE 2010, 5, e11707. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ren, X.; Zhang, Q.; Li, Z.; Ma, S.; Bao, J.; Li, Z.; Bai, X.; Zheng, L.; Zhang, Z.; et al. PGC-1alpha/ERRalpha-Sirt3 Pathway Regulates DAergic Neuronal Death by Directly Deacetylating SOD2 and ATP Synthase beta. Antioxid. Redox Signal. 2016, 24, 312–328. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Tu, B.P. Acetyl-CoA and the regulation of metabolism: Mechanisms and consequences. Curr. Opin. Cell Biol. 2015, 33, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Lo Sasso, G.; Schlage, W.K.; Boue, S.; Veljkovic, E.; Peitsch, M.C.; Hoeng, J. The Apoe(-/-) mouse model: A suitable model to study cardiovascular and respiratory diseases in the context of cigarette smoke exposure and harm reduction. J. Transl. Med. 2016, 14, 146. [Google Scholar] [CrossRef]
- Mancuso, M.; Pasquali, E.; Braga-Tanaka, I., 3rd; Tanaka, S.; Pannicelli, A.; Giardullo, P.; Pazzaglia, S.; Tapio, S.; Atkinson, M.J.; Saran, A. Acceleration of atherogenesis in ApoE -/- mice exposed to acute or low-dose-rate ionizing radiation. Oncotarget 2015, 6, 31263–31271. [Google Scholar] [CrossRef]
- Kumarathasan, P.; Vincent, R.; Blais, E.; Saravanamuthu, A.; Gupta, P.; Wyatt, H.; Mitchel, R.; Hannan, M.; Trivedi, A.; Whitman, S. Cardiovascular changes in atherosclerotic ApoE-deficient mice exposed to Co60 (gamma) radiation. PLoS ONE 2013, 8, e65486. [Google Scholar] [CrossRef]
- Calvo, S.E.; Clauser, K.R.; Mootha, V.K. MitoCarta2.0: An updated inventory of mammalian mitochondrial proteins. Nucleic Acids Res. 2016, 44, D1251–D1257. [Google Scholar] [CrossRef]
- Chan, S.H.; Wu, K.L.; Chang, A.Y.; Tai, M.H.; Chan, J.Y. Oxidative impairment of mitochondrial electron transport chain complexes in rostral ventrolateral medulla contributes to neurogenic hypertension. Hypertension 2009, 53, 217–227. [Google Scholar] [CrossRef]
- Karamanlidis, G.; Lee, C.F.; Garcia-Menendez, L.; Kolwicz, S.C., Jr.; Suthammarak, W.; Gong, G.; Sedensky, M.M.; Morgan, P.G.; Wang, W.; Tian, R. Mitochondrial complex I deficiency increases protein acetylation and accelerates heart failure. Cell Metab. 2013, 18, 239–250. [Google Scholar] [CrossRef]
- Jeninga, E.H.; Schoonjans, K.; Auwerx, J. Reversible acetylation of PGC-1: Connecting energy sensors and effectors to guarantee metabolic flexibility. Oncogene 2010, 29, 4617–4624. [Google Scholar] [CrossRef]
- Barger, P.M.; Brandt, J.M.; Leone, T.C.; Weinheimer, C.J.; Kelly, D.P. Deactivation of peroxisome proliferator-activated receptor-alpha during cardiac hypertrophic growth. J. Clin. Investig. 2000, 105, 1723–1730. [Google Scholar] [CrossRef] [PubMed]
- Papiez, A.; Azimzadeh, O.; Azizova, T.; Moseeva, M.; Anastasov, N.; Smida, J.; Tapio, S.; Polanska, J. Integrative multiomics study for validation of mechanisms in radiation-induced ischemic heart disease in Mayak workers. PLoS ONE 2018, 13, e0209626. [Google Scholar] [CrossRef] [PubMed]
- Polevoda, B.; Sherman, F. The diversity of acetylated proteins. Genome Biol. 2002, 3, reviews0006. [Google Scholar] [CrossRef] [PubMed]
- Matsushima, S.; Sadoshima, J. The role of sirtuins in cardiac disease. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1375–H1389. [Google Scholar] [CrossRef]
- Ali, I.; Conrad, R.J.; Verdin, E.; Ott, M. Lysine Acetylation Goes Global: From Epigenetics to Metabolism and Therapeutics. Chem. Rev. 2018, 118, 1216–1252. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Xu, W.; Jiang, W.; Yu, W.; Lin, Y.; Zhang, T.; Yao, J.; Zhou, L.; Zeng, Y.; Li, H.; et al. Regulation of cellular metabolism by protein lysine acetylation. Science 2010, 327, 1000–1004. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, Y.; Yang, C.; Xiong, H.; Lin, Y.; Yao, J.; Li, H.; Xie, L.; Zhao, W.; Yao, Y.; et al. Acetylation of metabolic enzymes coordinates carbon source utilization and metabolic flux. Science 2010, 327, 1004–1007. [Google Scholar] [CrossRef]
- Chen, T.; Liu, J.; Li, N.; Wang, S.; Liu, H.; Li, J.; Zhang, Y.; Bu, P. Mouse SIRT3 attenuates hypertrophy-related lipid accumulation in the heart through the deacetylation of LCAD. PLoS ONE 2015, 10, e0118909. [Google Scholar] [CrossRef]
- Grillon, J.M.; Johnson, K.R.; Kotlo, K.; Danziger, R.S. Non-histone lysine acetylated proteins in heart failure. Biochim. Biophys. Acta 2012, 1822, 607–614. [Google Scholar] [CrossRef] [Green Version]
- Lewin, M.B.; Timiras, P.S. Lipid changes with aging in cardiac mitochondrial membranes. Mech. Ageing Dev. 1984, 24, 343–351. [Google Scholar] [CrossRef]
- Hafner, A.V.; Dai, J.; Gomes, A.P.; Xiao, C.Y.; Palmeira, C.M.; Rosenzweig, A.; Sinclair, D.A. Regulation of the mPTP by SIRT3-mediated deacetylation of CypD at lysine 166 suppresses age-related cardiac hypertrophy. Aging 2010, 2, 914–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundaresan, N.R.; Gupta, M.; Kim, G.; Rajamohan, S.B.; Isbatan, A.; Gupta, M.P. Sirt3 blocks the cardiac hypertrophic response by augmenting Foxo3a-dependent antioxidant defense mechanisms in mice. J. Clin. Investig. 2009, 119, 2758–2771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cencioni, C.; Spallotta, F.; Mai, A.; Martelli, F.; Farsetti, A.; Zeiher, A.M.; Gaetano, C. Sirtuin function in aging heart and vessels. J. Mol. Cell. Cardiol. 2015, 83, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Donato, A.J.; Magerko, K.A.; Lawson, B.R.; Durrant, J.R.; Lesniewski, L.A.; Seals, D.R. SIRT-1 and vascular endothelial dysfunction with ageing in mice and humans. J. Physiol. 2011, 589, 4545–4554. [Google Scholar] [CrossRef]
- Ghosh, S.; Zhou, Z. SIRTain regulators of premature senescence and accelerated aging. Protein Cell 2015, 6, 322–333. [Google Scholar] [CrossRef] [Green Version]
- Grabowska, W.; Sikora, E.; Bielak-Zmijewska, A. Sirtuins, a promising target in slowing down the ageing process. Biogerontology 2017, 18, 447–476. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, I.; Minamino, T. Cellular senescence in cardiac diseases. J. Cardiol. 2019, 74, 313–319. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Ma, Y.; Song, L.; Yu, L.; Zhang, L.; Zhang, Y.; Xing, Y.; Yin, Y.; Ma, H. SIRT3 deficiency exacerbates p53/Parkinmediated mitophagy inhibition and promotes mitochondrial dysfunction: Implication for aged hearts. Int. J. Mol. Med. 2018, 41, 3517–3526. [Google Scholar]
- Lin, S.; Wang, Y.; Zhang, X.; Kong, Q.; Li, C.; Li, Y.; Ding, Z.; Liu, L. HSP27 Alleviates Cardiac Aging in Mice via a Mechanism Involving Antioxidation and Mitophagy Activation. Oxid. Med. Cell. Longev. 2016, 2016, 2586706. [Google Scholar] [CrossRef]
- Lu, J.; Zhang, H.; Chen, X.; Zou, Y.; Li, J.; Wang, L.; Wu, M.; Zang, J.; Yu, Y.; Zhuang, W.; et al. A small molecule activator of SIRT3 promotes deacetylation and activation of manganese superoxide dismutase. Free Radic. Biol. Med. 2017, 112, 287–297. [Google Scholar] [CrossRef]
- Tao, R.; Coleman, M.C.; Pennington, J.D.; Ozden, O.; Park, S.H.; Jiang, H.; Kim, H.S.; Flynn, C.R.; Hill, S.; Hayes McDonald, W.; et al. Sirt3-mediated deacetylation of evolutionarily conserved lysine 122 regulates MnSOD activity in response to stress. Mol. Cell 2010, 40, 893–904. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.J.; Katunga, L.A.; Willis, M.S. Mitochondria as a source and target of lipid peroxidation products in healthy and diseased heart. Clin. Exp. Pharmacol. Physiol. 2012, 39, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Dai, S.H.; Li, X.; Luo, P.; Zhu, J.; Wang, Y.H.; Fei, Z.; Jiang, X.F. Sirt1-Sirt3 axis regulates human blood-brain barrier permeability in response to ischemia. Redox. Biol. 2018, 14, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.; Seok, S.; Yau, P.; Li, X.; Kemper, B.; Kemper, J.K. Obesity and aging diminish sirtuin 1 (SIRT1)-mediated deacetylation of SIRT3, leading to hyperacetylation and decreased activity and stability of SIRT3. J. Biol. Chem. 2017, 292, 17312–17323. [Google Scholar] [CrossRef] [Green Version]
- Carnevale, I.; Pellegrini, L.; D’Aquila, P.; Saladini, S.; Lococo, E.; Polletta, L.; Vernucci, E.; Foglio, E.; Coppola, S.; Sansone, L.; et al. SIRT1-SIRT3 Axis Regulates Cellular Response to Oxidative Stress and Etoposide. J. Cell. Physiol. 2017, 232, 1835–1844. [Google Scholar] [CrossRef]
- Verdin, E.; Hirschey, M.D.; Finley, L.W.; Haigis, M.C. Sirtuin regulation of mitochondria: Energy production, apoptosis, and signaling. Trends Biochem. Sci. 2010, 35, 669–675. [Google Scholar] [CrossRef]
- Khan, S.A.; Sathyanarayan, A.; Mashek, M.T.; Ong, K.T.; Wollaston-Hayden, E.E.; Mashek, D.G. ATGL-catalyzed lipolysis regulates SIRT1 to control PGC-1alpha/PPAR-alpha signaling. Diabetes 2015, 64, 418–426. [Google Scholar] [CrossRef]
- Barjaktarovic, Z.; Kempf, S.J.; Sriharshan, A.; Merl-Pham, J.; Atkinson, M.J.; Tapio, S. Ionizing radiation induces immediate protein acetylation changes in human cardiac microvascular endothelial cells. J. Radiat. Res. 2015, 56, 623–632. [Google Scholar] [CrossRef] [Green Version]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef]
- Spellman, P.T.; Sherlock, G.; Zhang, M.Q.; Iyer, V.R.; Anders, K.; Eisen, M.B.; Brown, P.O.; Botstein, D.; Futcher, B. Comprehensive identification of cell cycle-regulated genes of the yeast Saccharomyces cerevisiae by microarray hybridization. Mol. Biol. Cell 1998, 9, 3273–3297. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids. Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed]
- Kramer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | ID | Description | No. Of Hyperacetylated Peptides | No. Of Hypoacetylated Peptides | GO Description | GO Accession |
|---|---|---|---|---|---|---|
| 1 | ACAA2 | 3-ketoacyl-CoA thiolase | 2 | 0 | FAO | GO:0006635 |
| 2 | ACADL | Long-chain specific acyl-CoA dehydrogenase | 2 | 0 | FAO | GO:0006635 |
| 3 | ACADM | Medium-chain specific acyl-CoA dehydrogenase | 1 | 0 | FAO | GO:0006635 |
| 4 | ACAT1 | Acetyl-CoA acetyltransferase | 1 | 0 | FAO | GO:0006635 |
| 5 | ACO2 | Aconitate hydratase | 6 | 1 | TCA metabolic process | GO:0072350 |
| 6 | ACOT13 | Acyl-coenzyme A thioesterase 13 | 4 | 0 | acyl-CoA hydrolase activity | GO:0047617 |
| 7 | ACOT5 | Acyl-coenzyme A thioesterase 5 | 1 | 0 | acyl-CoA hydrolase activity | GO:0047617 |
| 8 | ALDH4A1 | Delta-1-pyrroline-5-carboxylate dehydrogenase | 1 | 0 | TCA metabolic process | GO:0072350 |
| 9 | ATP5A1 | ATP synthase subunit alpha | 1 | 0 | ATP metabolic process | GO:0046034 |
| 10 | ATP5B | ATP synthase subunit beta | 2 | 0 | ATP metabolic process | GO:0046034 |
| 11 | ATP5F1 | ATP synthase F(0) complex subunit B1 | 3 | 0 | ATP metabolic process | GO:0046034 |
| 12 | ATP5H | ATP synthase subunit d | 9 | 0 | ATP metabolic process | GO:0046034 |
| 13 | ATP5L | ATP synthase subunit g | 2 | 0 | ATP metabolic process | GO:0046034 |
| 14 | ATP5O | ATP synthase subunit O | 1 | 0 | ATP metabolic process | GO:0046034 |
| 15 | AUH | Methylglutaconyl-CoA hydratase | 1 | 0 | FAO | GO:0006635 |
| 16 | CBLB | E3 ubiquitin-protein ligase | 1 | 0 | ubiquitin protein ligase binding | GO:0031625 |
| 17 | COX4I1 | Cytochrome c oxidase subunit 4 isoform 1 | 2 | 0 | ATP metabolic process | GO:0046034 |
| 18 | CUX1 | Homeobox protein cut-like 1 | 1 | 0 | DNA binding | GO:0003677 |
| 19 | CWC27 | Peptidyl-prolyl cis-trans isomerase | 1 | 0 | protein folding | GO:0006457 |
| 20 | CYCS | Cytochrome c | 1 | 0 | ATP metabolic process | GO:0046034 |
| 21 | DECR1 | 2-4-dienoyl-CoA reductase | 1 | 0 | FAO | GO:0006635 |
| 22 | DLD | Dihydrolipoyl dehydrogenase | 3 | 1 | ATP metabolic process | GO:0046034 |
| 23 | DLST | Dihydrolipoyllysine-residue succinyltransferase component of 2-oxoglutarate dehydrogenase complex | 1 | 0 | TCA metabolic process | GO:0072350 |
| 24 | ECHS1 | Enoyl-CoA hydratase | 1 | 0 | FAO | GO:0006635 |
| 25 | ETFA | Electron transfer flavoprotein subunit alpha | 3 | 1 | FAO | GO:0006635 |
| 26 | ETFB | Electron transfer flavoprotein subunit beta | 1 | 0 | FAO | GO:0006635 |
| 27 | FAM187A | Ig-like V-type domain-containing protein | 1 | 0 | enzyme binding | GO:0019899 |
| 28 | FGD6 | RhoGEF and PH domain-containing protein 6 | 1 | 0 | regulation of GTPase activity | GO:0043087 |
| 29 | FH | Fumarate hydratase | 5 | 0 | TCA metabolic process | GO:0072350 |
| 30 | GBAS | Protein NipSnap homolog 2 | 1 | 0 | ATP metabolic process | GO:0046034 |
| 31 | GLUD1 | Glutamate dehydrogenase 1 | 1 | 0 | TCA metabolic process | GO:0072350 |
| 32 | GOT2 | Aspartate aminotransferase | 3 | 1 | amino acid binding | GO:0016597 |
| 33 | HADH | Hydroxyacyl-coenzyme A dehydrogenase | 1 | 0 | FAO | GO:0006635 |
| 34 | HADHA | Trifunctional enzyme subunit alpha | 7 | 0 | FAO | GO:0006635 |
| 35 | HIST1H1C | Histone H1 | 1 | 0 | DNA binding | GO:0003677 |
| 36 | HIST1H1E | Histone H1 | 1 | 0 | ATP binding | GO:0005524 |
| 37 | HMGCL | Hydroxymethylglutaryl-CoA lyase | 1 | 0 | fatty-acyl-CoA binding | GO:0000062 |
| 38 | HSPA9 | Stress-70 protein | 1 | 0 | unfolded protein binding | GO:0051082 |
| 39 | HSPD1 | 60 kDa heat shock protein | 1 | 0 | unfolded protein binding | GO:0051082 |
| 40 | IDH2 | Isocitrate dehydrogenase [NADP] | 12 | 0 | TCA metabolic process | GO:0072350 |
| 41 | IDH3A | Isocitrate dehydrogenase [NADP] | 0 | 1 | TCA metabolic process | GO:0072350 |
| 42 | LRRC40 | Leucine-rich repeat-containing protein 40 | 1 | 0 | cellular process | GO:0009987 |
| 43 | MDH1B | Putative malate dehydrogenase 1B | 1 | 0 | TCA metabolic process | GO:0072350 |
| 44 | MDH2 | Malate dehydrogenase | 6 | 0 | TCA metabolic process | GO:0072350 |
| 45 | MMEL1 | Membrane metallo-endopeptidase-like 1 | 1 | 0 | endopeptidase activity | GO:0004175 |
| 46 | MRGPRA1 | Mas-related G-protein coupled receptor member A1 | 0 | 1 | G protein-coupled receptor activity | GO:0004930 |
| 47 | MYH6 | Myosin-6 | 2 | 1 | actin-dependent ATPase activity | GO:0030898 |
| 48 | NDUFA2 | NADH dehydrogenase [ubiquinone] 1 alpha subcomplex subunit 2 | 1 | 0 | NADH dehydrogenase (ubiquinone) activity | GO:0008137 |
| 49 | NDUFAB1 | Acyl carrier protein | 0 | 1 | acyl binding | GO:0000035 |
| 50 | NDUFB3 | NADH dehydrogenase [ubiquinone] 1 beta subcomplex subunit 3 | 1 | 0 | NADH dehydrogenase (ubiquinone) activity | GO:0008137 |
| 51 | NDUFS1 | NADH-ubiquinone oxidoreductase 75 kDa subunit | 2 | 0 | NADH dehydrogenase (ubiquinone) activity | GO:0008137 |
| 52 | NDUFV2 | NADH dehydrogenase [ubiquinone] flavoprotein 2 | 1 | 0 | NADH dehydrogenase (ubiquinone) activity | GO:0008137 |
| 53 | NIT2 | Omega-amidase NIT2 | 1 | 0 | omega-amidase activity | GO:0050152 |
| 54 | OXCT1 | Succinyl-CoA:3-ketoacid coenzyme A transferase 1 | 1 | 0 | 3-oxoacid CoA-transferase activity | GO:0008260 |
| 55 | PAPOLB | Poly(A) polymerase beta | 1 | 0 | RNA polymerase binding | GO:0070063 |
| 56 | PDHA1 | Pyruvate dehydrogenase E1 component subunit alpha | 1 | 0 | TCA metabolic process | GO:0072350 |
| 57 | PRDX5 | Peroxiredoxin-5 | 2 | 0 | response to oxygen radical | GO:0000305 |
| 58 | PROSC | Proline synthase co-transcribed bacterial homolog protein | 1 | 0 | pyridoxal phosphate binding | GO:0030170 |
| 59 | RPIA | Ribose-5-phosphate isomerase | 1 | 0 | carbohydrate binding | GO:0030246 |
| 60 | SDHA | Succinate dehydrogenase [ubiquinone] flavoprotein subunit | 4 | 0 | TCA metabolic process | GO:0072350 |
| 61 | SLC25A3 | Phosphate carrier protein | 1 | 0 | phosphate transmembrane transporter | GO:0005315 |
| 62 | SLC25A4 | ADP/ATP translocase 1 | 2 | 1 | ATP:ADP antiporter activity | GO:0005471 |
| 63 | SLC25A5 | ADP/ATP translocase 2 | 1 | 0 | ATP:ADP antiporter activity | GO:0005471 |
| 64 | SOD2 | Superoxide dismutase [Mn] | 1 | 0 | response to oxygen radical | GO:0000305 |
| 65 | SUCLA2 | Succinate--CoA ligase [ADP-forming] subunit beta | 1 | 0 | TCA metabolic process | GO:0072350 |
| 66 | SUCLG1 | Succinate--CoA ligase [ADP/GDP-forming] subunit alpha | 1 | 0 | TCA metabolic process | GO:0072350 |
| 67 | TXNRD2 | Thioredoxin reductase 2 | 1 | 0 | response to oxygen radical | GO:0000305 |
| 68 | UQCRB | Cytochrome b-c1 complex subunit 7 | 1 | 0 | ATP metabolic process | GO:0046034 |
| 69 | UQCRC1 | Cytochrome b-c1 complex subunit 1 | 1 | 0 | ATP metabolic process | GO:0046034 |
| 70 | VDAC1 | Voltage-dependent anion-selective channel protein 1 | 5 | 0 | anion channel activity | GO:0005253 |
| 71 | VDAC3 | Voltage-dependent anion-selective channel protein 3 | 1 | 0 | anion channel activity | GO:0005253 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barjaktarovic, Z.; Merl-Pham, J.; Braga-Tanaka, I.; Tanaka, S.; Hauck, S.M.; Saran, A.; Mancuso, M.; Atkinson, M.J.; Tapio, S.; Azimzadeh, O. Hyperacetylation of Cardiac Mitochondrial Proteins Is Associated with Metabolic Impairment and Sirtuin Downregulation after Chronic Total Body Irradiation of ApoE -/- Mice. Int. J. Mol. Sci. 2019, 20, 5239. https://doi.org/10.3390/ijms20205239
Barjaktarovic Z, Merl-Pham J, Braga-Tanaka I, Tanaka S, Hauck SM, Saran A, Mancuso M, Atkinson MJ, Tapio S, Azimzadeh O. Hyperacetylation of Cardiac Mitochondrial Proteins Is Associated with Metabolic Impairment and Sirtuin Downregulation after Chronic Total Body Irradiation of ApoE -/- Mice. International Journal of Molecular Sciences. 2019; 20(20):5239. https://doi.org/10.3390/ijms20205239
Chicago/Turabian StyleBarjaktarovic, Zarko, Juliane Merl-Pham, Ignacia Braga-Tanaka, Satoshi Tanaka, Stefanie M. Hauck, Anna Saran, Mariateresa Mancuso, Michael J. Atkinson, Soile Tapio, and Omid Azimzadeh. 2019. "Hyperacetylation of Cardiac Mitochondrial Proteins Is Associated with Metabolic Impairment and Sirtuin Downregulation after Chronic Total Body Irradiation of ApoE -/- Mice" International Journal of Molecular Sciences 20, no. 20: 5239. https://doi.org/10.3390/ijms20205239