Anti-Diabetic Nephropathy Activities of Polysaccharides Obtained from Termitornyces albuminosus via Regulation of NF-κB Signaling in db/db Mice


Abstract
:1. Introduction
2. Results
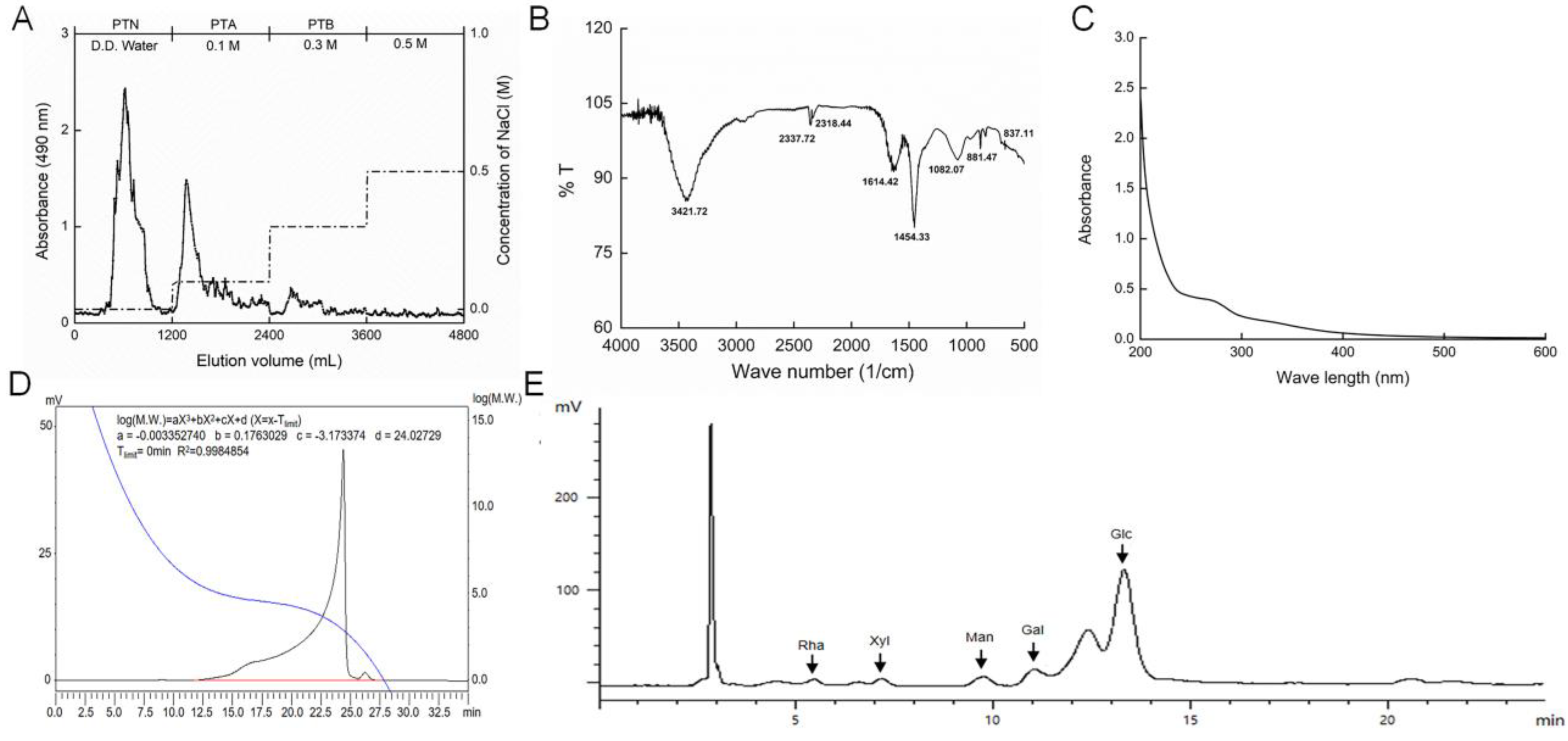
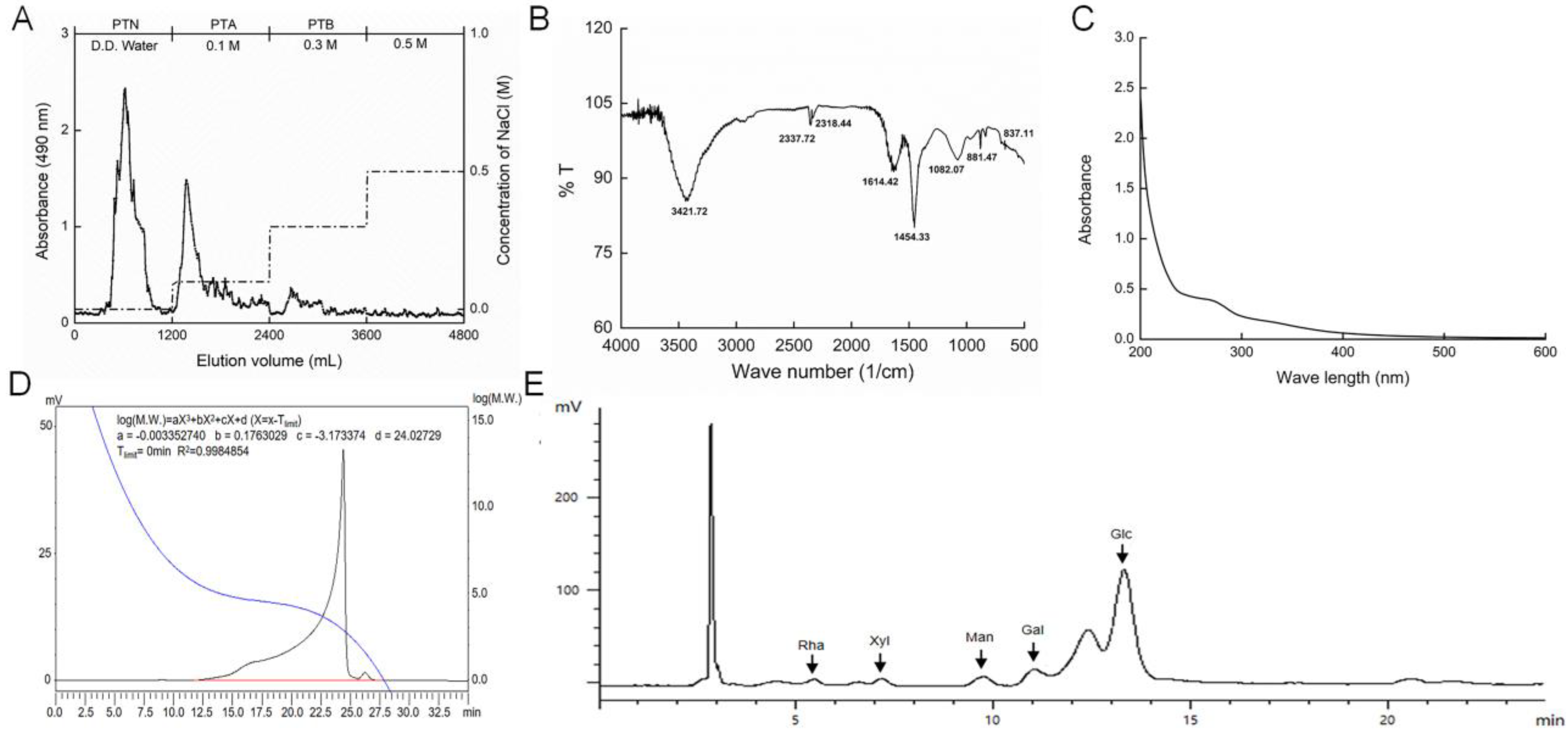
2.1. Characterization of PTA
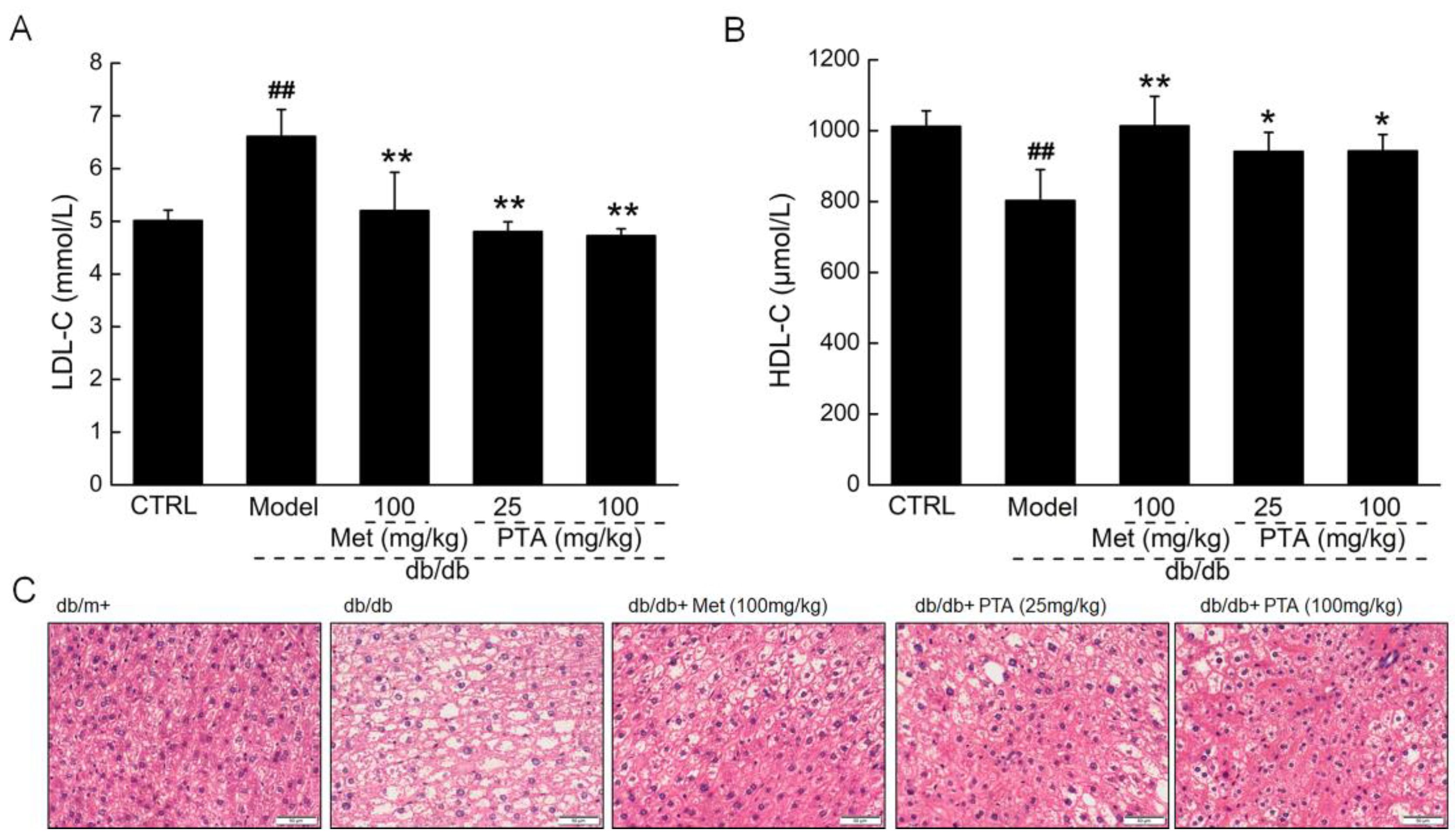
2.2. PTA Regulates Lipid Metabolism in db/db Mice
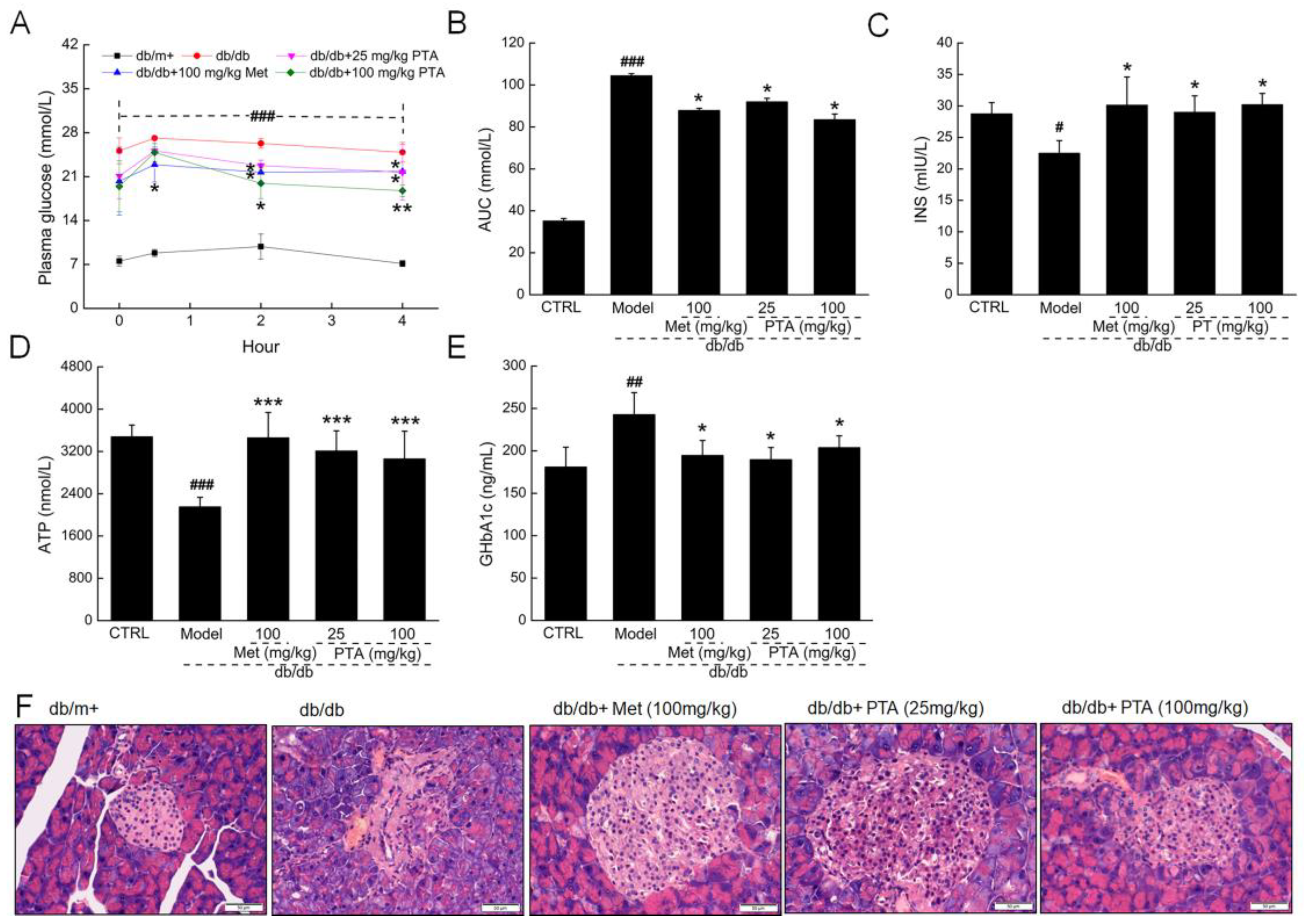
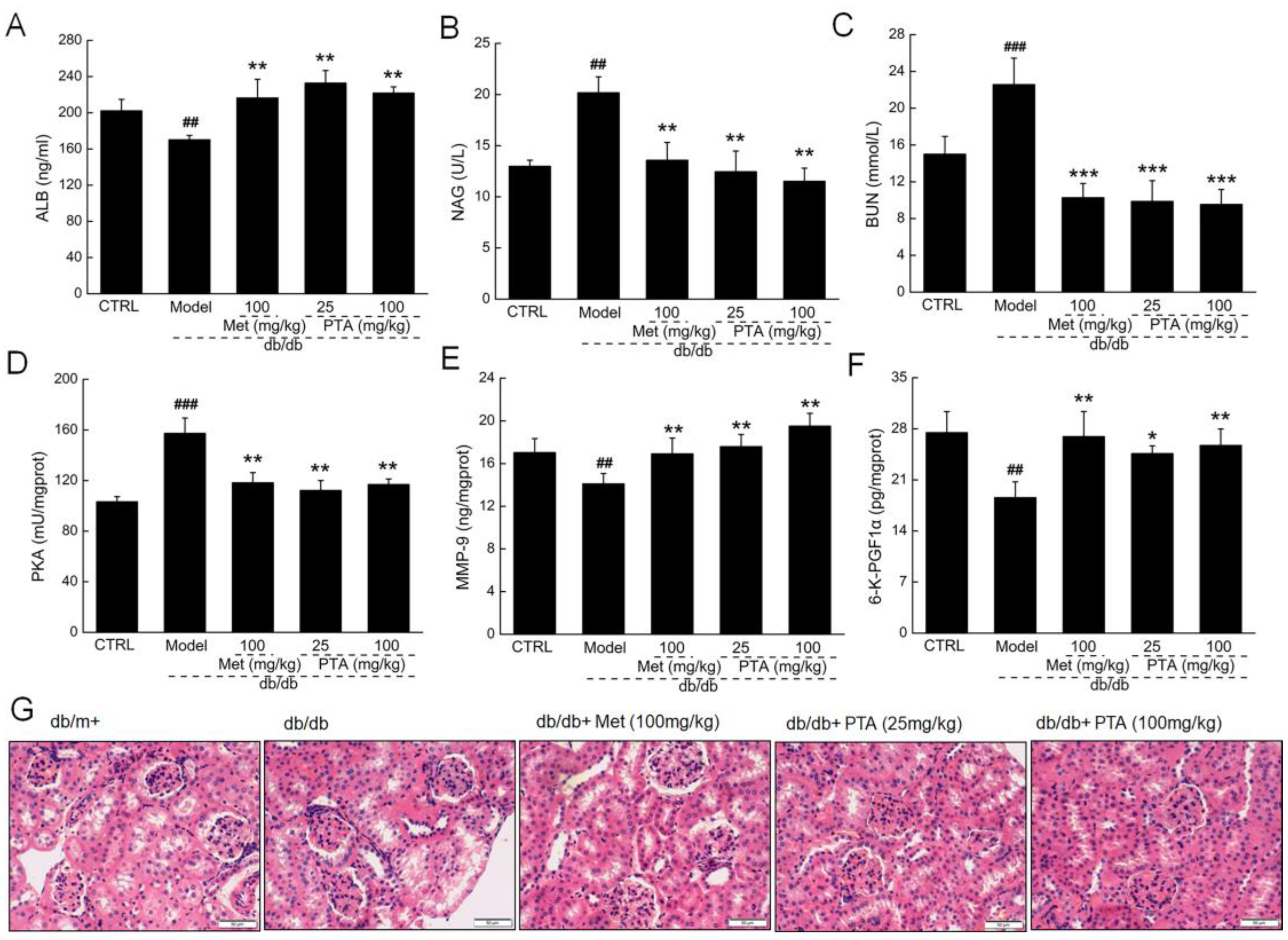
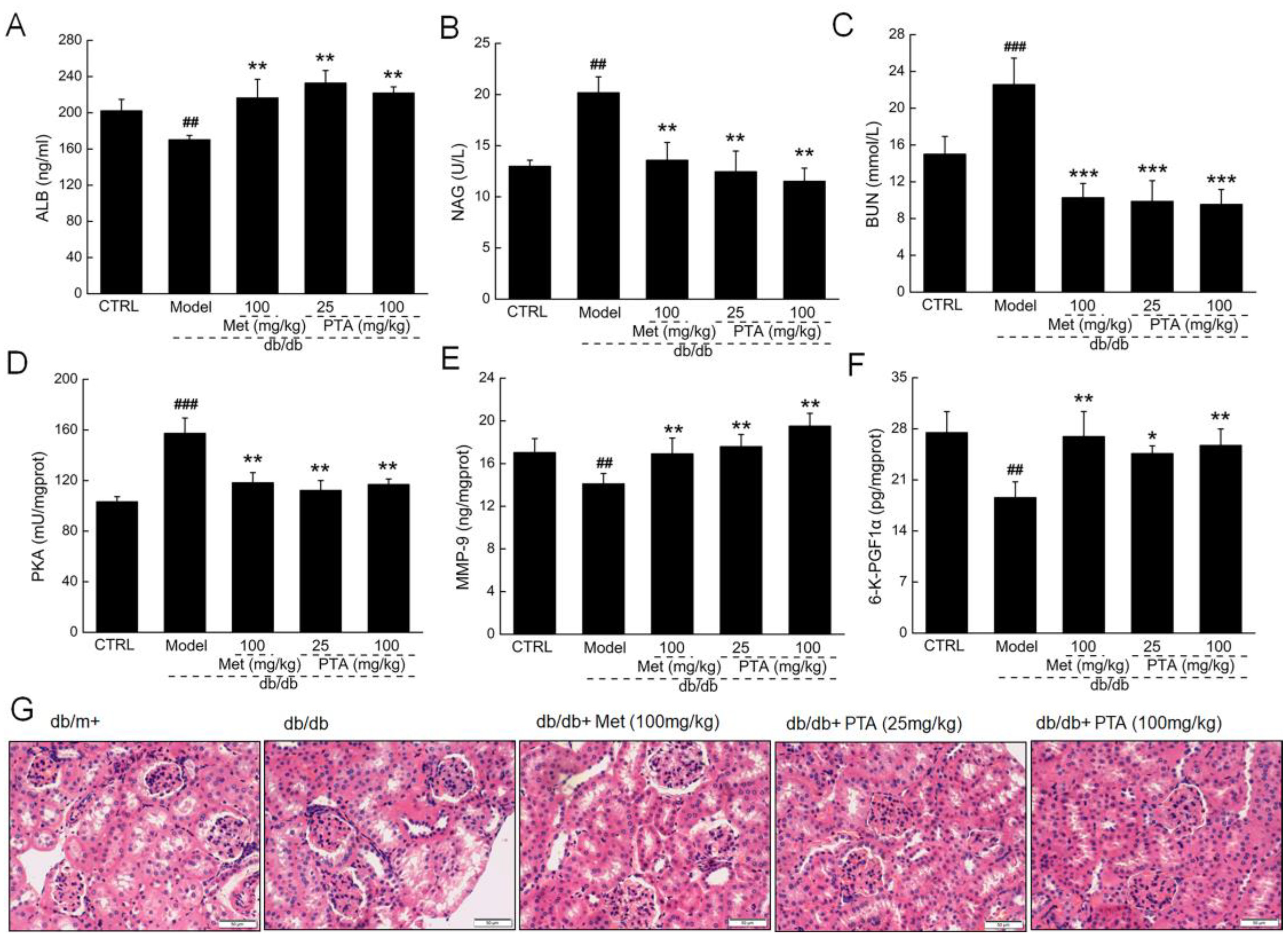
2.3. Renal Protection by PTA in db/db Mice
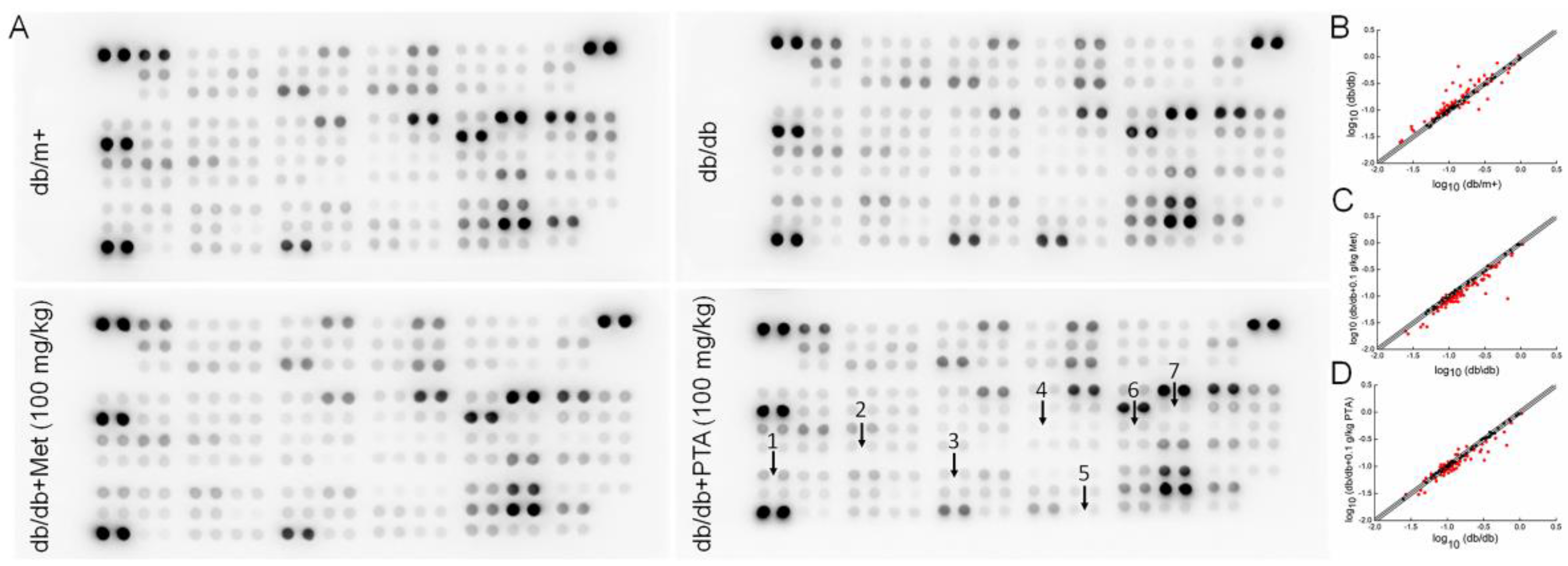
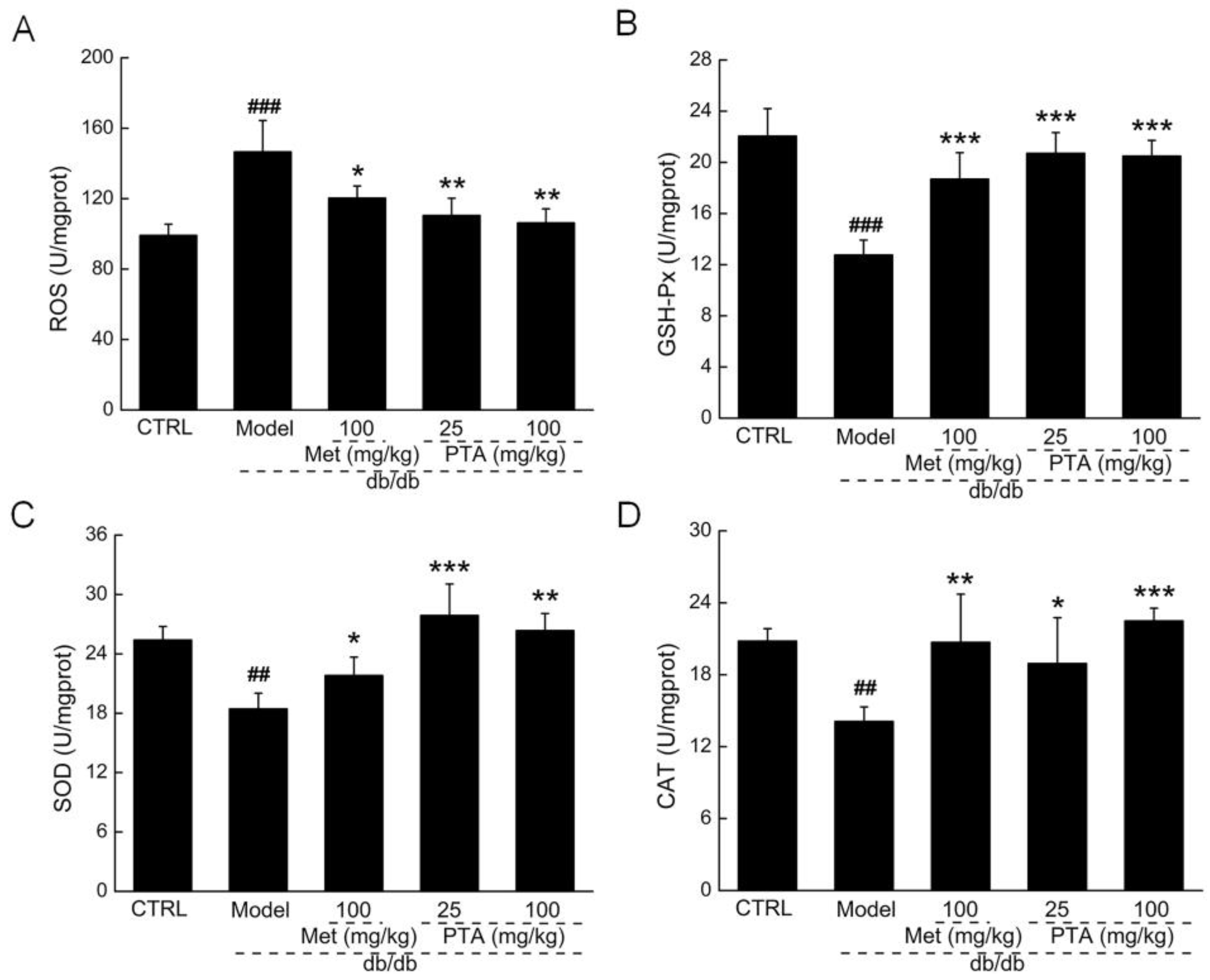
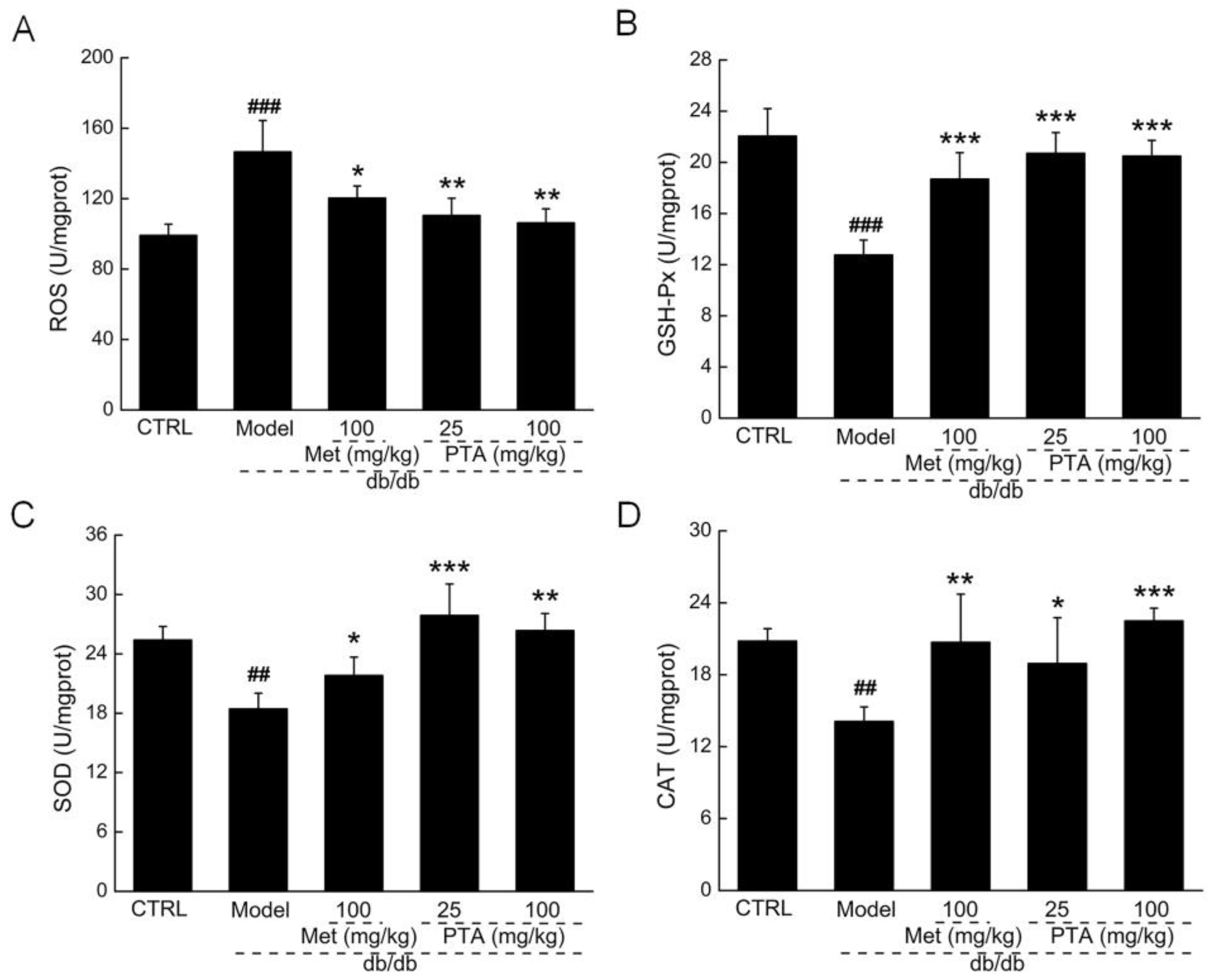
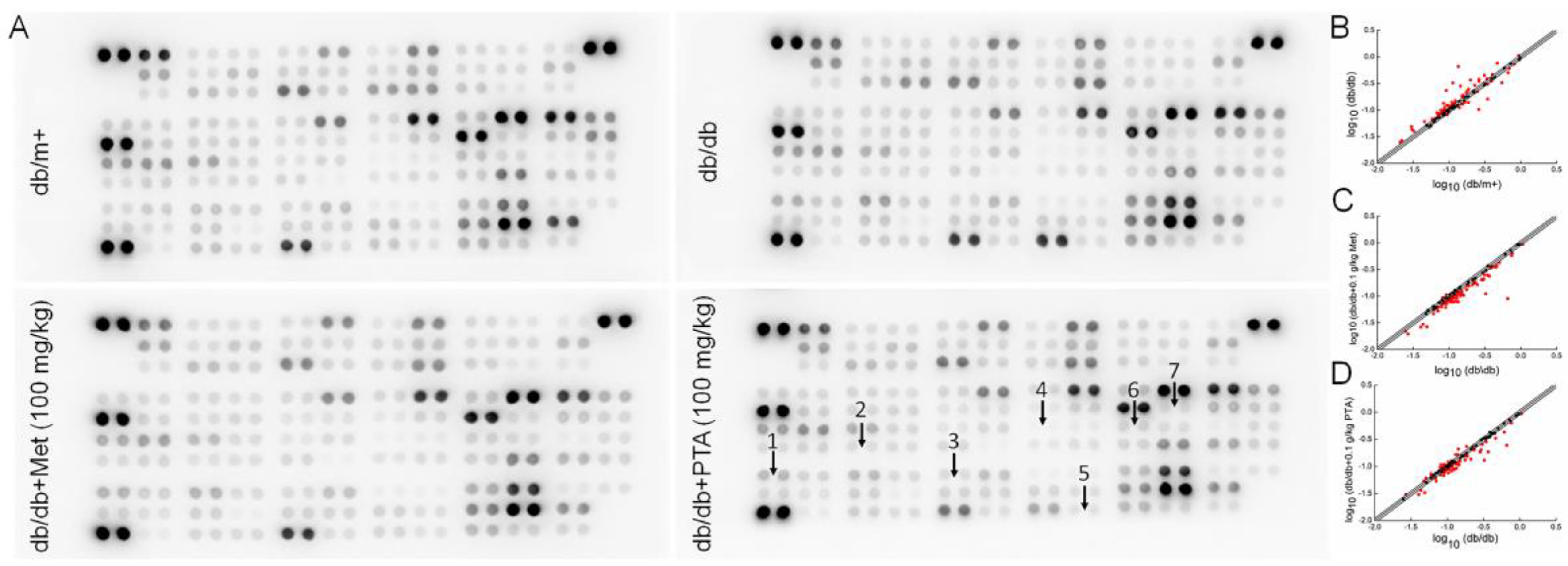
2.4. Anti-Inflammation and Anti-Oxidation Pathways are Involved in PTA-Mediated Renal Protection
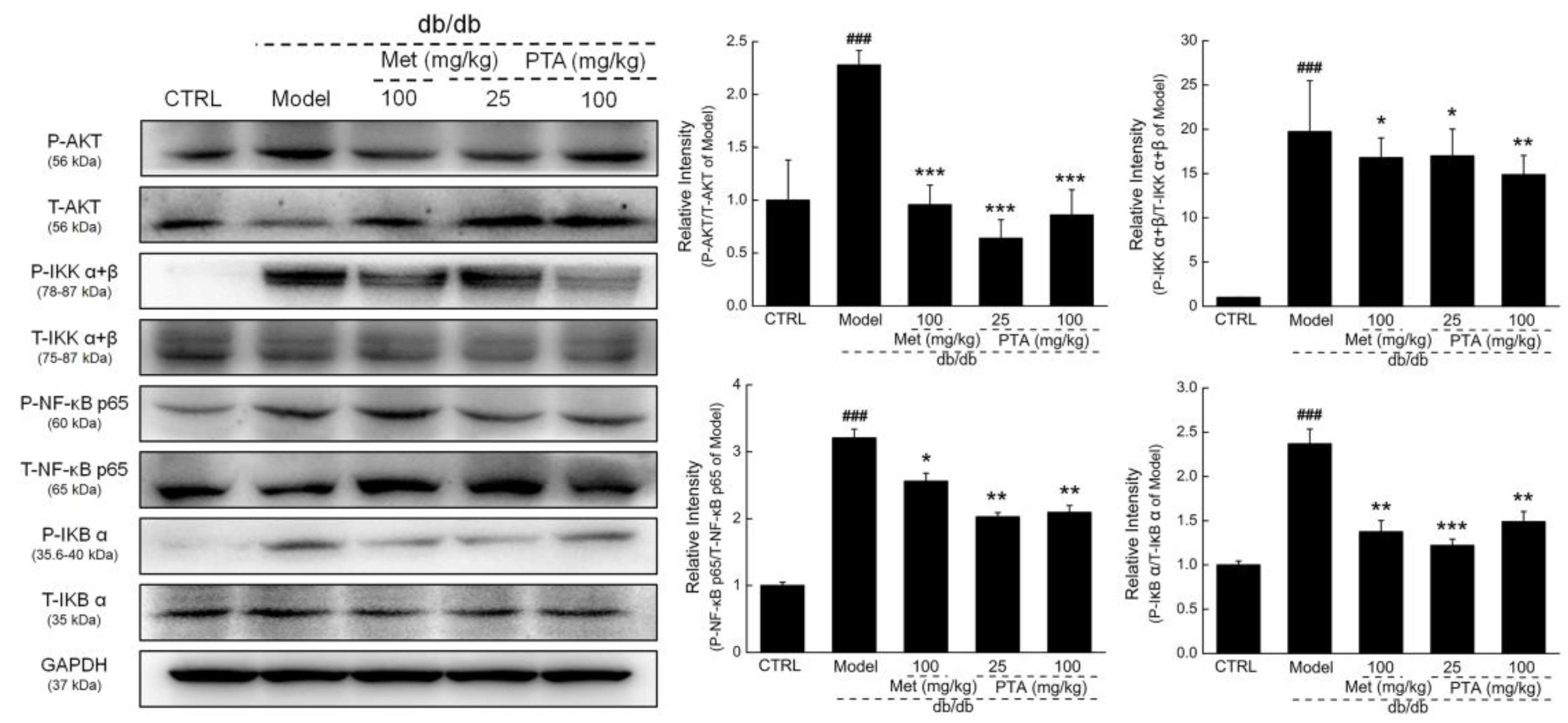
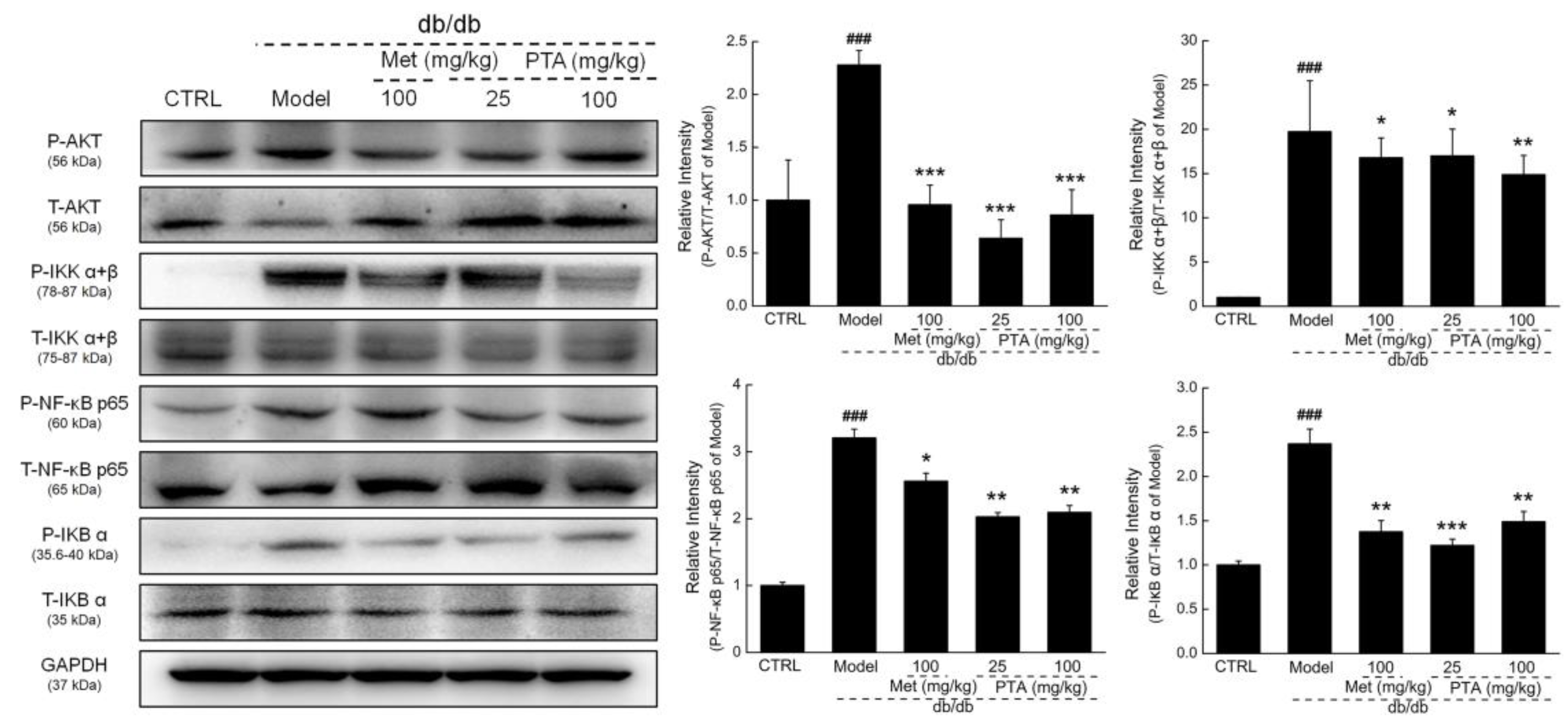
2.5. NF-κB Signaling Might be Involved in PTA-Mediated Renal Protection
3. Discussion
4. Materials and Methods
4.1. Preparation of Polysaccharides from T. albuminosus Fruiting Body
4.2. Characteristic Analysis of PTA
4.2.1. Ultraviolet (UV) Spectra Measurement.
4.2.2. FT-IR Spectra Analysis.
4.2.3. Molecular Weight Analysis.
4.2.4. Monosaccharides Analysis.
4.3. Experimental Protocol Performed in db/db Mice
4.4. Histopathological Examination
4.5. Antibody Array Assay of Kidney
4.6. Biochemical Indexes Measurement
4.7. Western Blotting
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| AKT | Protein Kinase B |
| ALB | Albumin |
| Ara | L-Arabinose |
| ATP | Adenosine Triphosphate |
| AUC | Area Under the Curve |
| BCA | Butyleyanoacrylate |
| BSA | Bovine Serum Albumin |
| BUN | Blood Urea Nitrogen |
| CAT | Catalase |
| CTRL | Control Group |
| DEAE−52 | Diethylethanolamine-52 |
| DM | Diabetes Mellitus |
| DN | Diabetic Nephropathy |
| ECL | Electrochemiluminescence |
| ELISA | Enzyme-Linked Immunosorbent Assay |
| FT-IR | Fourier Transform Infrared |
| Gal | D-Galactose |
| GHbA1c | Glycosylated Hemoglobin A1c |
| Glc | D-Glucose |
| H and E | Hematoxylin Eosin |
| GAPDH | Glyceraldehyde-3-Phosphate Dehydrogenase |
| GSH-Px | Glutathione Peroxide |
| HDL-C | High Density Lipid Cholesterol |
| HPLC | High Performance Liquid Chromatography |
| IFN-γ | Interferon-Γ |
| IκBα | Inhibitor Of ΚB Alpha |
| IKKα+β | Inhibitor Of ΚB Kinase Alpha + Beta |
| ILs | Interleukins |
| INS | Insulin |
| LDL-C | Low Density Lipid Cholesterol |
| LEP | Leptin |
| Man | D-Mannose |
| MCP-1 | Monocyte Chemotactic Protein-1 |
| M-CSF | Macrophage Colony-Stimulating Factor |
| Met | Metformin |
| MMP-9 | Matrix Metalloproteinase-9 |
| NAG | N-Acetyl-Β-D-Glucosidase |
| NF-κB | Nuclear Factor-ΚB |
| OGTT | Oral Glucose Tolerance Test |
| PKA | cAMP-Dependent Protein Kinase |
| PMSF | Phenylmethanesulfonyl Fluoride |
| PVDF | Polyvinylidene Difluoride |
| Rha | L-Rhamnose |
| RIPA | Radio Immunoprecipitation Assay |
| ROS | Reactive Oxygen Species |
| SDS-PAGE | Sodium Dodecyl Sulfate–Polyacrylamide Gel Electrophoresis |
| SOD | Superoxide Dismutase |
| T2DM | Type 2 DM |
| TNF-α | Tumor Necrosis Factor-A |
| UV | Ultraviolet |
| Xyl | D-Xylose |
| 6-K-PGF1α | 6-Keto Prostaglandin F 1A |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Week | db/m+ | db/db | db/db | |||
|---|---|---|---|---|---|---|
| Met (100 mg/kg) | PTA (25 mg/kg) | PTA (100 mg/kg) | ||||
| Body weights (g) | 1 | 20.4 ± 2.2 | 41.6 ± 2.3### | 40.6 ± 2.9 | 40.8 ± 3.0 | 40.3 ± 2.6 |
| 2 | 18.4 ± 2.5 | 42.7 ± 2.6### | 39.3 ± 3.2* | 40.0 ± 3.0* | 40.1 ± 2.4* | |
| 3 | 20.3 ± 2.1 | 45.4 ± 1.9### | 42.9 ± 3.0* | 41.8 ± 2.8** | 42.6 ± 1.8** | |
| 4 | 21.4 ± 1.1 | 46.6 ± 2.2### | 43.8 ± 3.7* | 43.8 ± 2.9* | 44.3 ± 2.1* | |
| 5 | 22.1 ± 0.9 | 48.8 ± 2.9### | 45.4 ± 2.9* | 45.3 ± 2.4** | 46.0 ± 2.1* | |
| 6 | 22.0 ± 0.8 | 51.5 ± 2.6### | 45.9 ± 3.1** | 45.6 ± 3.2*** | 46.9 ± 2.1** | |
| 7 | 23.0 ± 0.8 | 51.1 ± 3.2### | 45.3 ± 3.0** | 45.9 ± 2.9** | 47.8 ± 2.4* | |
| 8 | 22.7 ± 1.4 | 51.1 ± 3.1### | 45.5 ± 1.9*** | 46.1 ± 2.7** | 48.0 ± 2.8* | |
| Plasma glucose (mmol/L) | 1 | 5.0 ± 2.3 | 17.0 ± 6.2### | 16.9 ± 6.4 | 15.6 ± 6.5 | 17.9 ± 3.5 |
| 2 3 | 6.6 ± 1.3 6.8 ± 1.6 | 20.4 ± 3.8### 20.2 ± 3.9### | 17.8 ± 3.3 18.8 ± 3.2 | 16.6 ± 3.6* 17.3 ± 2.1 | 17.9 ± 1.7 17.4 ± 2.3 | |
| 4 5 | 6.7 ± 0.9 6.8 ± 0.8 | 20.3 ± 3.9### 21.0 ± 3.6### | 19.0 ± 1.1 17.3 ± 2.9* | 18.1 ± 3.2 17.6 ± 3.1 | 17.2 ± 2.5* 16.7 ± 2.3** | |
| 6 7 | 6.1 ± 1.0 7.0 ± 1.3 | 21.2 ± 2.2### 21.7 ± 2.2### | 17.2 ± 2.3** 17.8 ± 2.6** | 18.9 ± 2.3* 18.9 ± 2.7* | 16.2 ± 2.7*** 16.1 ± 2.1*** | |
| 8 | 6.7 ± 0.7 | 23.4 ± 2.3### | 17.1 ± 2.0*** | 17.4 ± 3.0*** | 16.4 ± 2.2*** | |
| Coordinate | Target | Fold | |
|---|---|---|---|
| db/db (vs.db/m+) | db/db+100 mg/kg PTA (vs.db/db) | ||
| A 3; A 4 | Adiponectin/Acrp30 | −24.8% | −0.4% |
| A 5; A 6 | Amphiregulin | 18.6% | −6.7% |
| A 7; A 8 | Angiopoietin-1 | 15.0% | −3.9% |
| A 9; A 10 | Angiopoietin-2 | 29.8% | 3.4% |
| A 11; A 12 | Angiopoietin-like 3 | 43.6% | −7.8% |
| A 13; A 14 | BAFF/BLyS/TNFSF13B | −1.9% | 21.2% |
| A 15; A 16 | C1q R1/CD93 | 14.1% | 9.9% |
| A 17; A 18 | CCL2/JE/MCP-1 | −3.6% | 30.9% |
| A 19; A 20 | CCL3/CCL4 MIP-1 alpha/beta | 0.7% | 13.9% |
| A 21; A 22 | CCL5/RANTES | −6.0% | −12.3% |
| B 3; B 4 | CCL6/C10 | 31.9% | −17.4% |
| B 5; B 6 | CCL11/Eotaxin | 23.3% | −5.9% |
| B 7; B 8 | CCL12/MCP-5 | 12.7% | −9.8% |
| B 9; B 10 | CCL17/TARC | 17.1% | −11.0% |
| B 11; B 12 | CCL19/MIP-3 beta | 3.2% | 1.9% |
| B 13; B 14 | CCL20/MIP-3 alpha | −12.6% | 13.9% |
| B 15; B 16 | CCL21/6Ckine | 3.2% | 0.9% |
| B 17; B 18 | CCL22/MDC | −3.4% | 20.2% |
| B 19; B 20 | CD14 | 5.5% | 11.1% |
| B 21; B 22 | CD40/TNFRSF5 | 29.0% | −11.6% |
| C 3; C 4 | CD160 | 8.9% | 5.7% |
| C 5; C 6 | Chemerin | 20.7% | −5.1% |
| C 7; C 8 | Chitinase 3-like 1 | 187.7% | −60.1% |
| C 9; C 10 | Coagulation Factor III/ Tissue Factor | −4.9% | −1.2% |
| C 11; C 12 | Complement Component C5/C5a | −7.6% | 1.7% |
| C 13; C 14 | Complement Factor D | −60.4% | 17.6% |
| C 15; C 16 | C-Reactive Protein/CRP | 44.5% | −8.6% |
| C 17; C 18 | CX3CL1/Fractalkine | −20.9% | 6.8% |
| C 19; C 20 | CXCL1/KC | 2.4% | 7.6% |
| C 21; C 22 | CXCL2/MIP-2 | 11.3% | 6.9% |
| D 1; D 2 | CXCL9/MIG | 4.8% | −25.2% |
| D 3; D 4 | CXCL10/IP-10 | 8.7% | −20.1% |
| D 5; D 6 | CXCL11/I-TAC | −0.3% | −6.8% |
| D 7; D 8 | CXCL13/BLC/BCA-1 | 31.4% | −28.2% |
| D 9; D 10 | CXCL16 | 20.5% | −6.3% |
| D 11; D 12 | Cystatin C | −1.0% | −6.7% |
| D 13; D 14 | DKK-1 | −8.5% | 16.9% |
| D 15; D 16 | DPPIV/CD26 | −13.1% | 3.0% |
| D 17; D 18 | EGF | −4.5% | −3.2% |
| D 19; D 20 | Endoglin/CD105 | −6.2% | −2.3% |
| D 21; D 22 | Endostatin | 3.2% | −9.9% |
| D 23; D 24 | Fetuin A/AHSG | −11.2% | −16.1% |
| E 1; E 2 | FGF acidic | −2.6% | −2.9% |
| E 3; E 4 | FGF-21 | −7.4% | −5.0% |
| E 5; E 6 | Flt-3 Ligand | −6.4% | 1.8% |
| E 7; E 8 | Gas 6 | 9.7% | −7.1% |
| E 9; E 10 | G-CSF | 2.7% | 7.0% |
| E 11; E 12 | GDF-15 | 4.5% | 18.4% |
| E 13; E 14 | GM-CSF | −8.8% | 19.0% |
| E 15; E 16 | HGF | −16.4% | 26.5% |
| E 17; E 18 | ICAM-1/CD54 | −10.9% | 1.1% |
| E 19; E 20 | IFN-gamma | −14.9% | 45.9% |
| E 21; E 22 | IGFBP-1 | 32.6% | −2.9% |
| E 23; E 24 | IGFBP-2 | −53.4% | −2.0% |
| F 1; F 2 | IGFBP-3 | −3.0% | 4.9% |
| F 3; F 4 | IGFBP-5 | −4.3% | 4.5% |
| F 5; F 6 | IGFBP-6 | −11.0% | −5.2% |
| F 7; F 8 | IL-1 alpha/IL-1F1 | 5.0% | −1.8% |
| F 9; F 10 | IL-1 beta/ IL-1F2 | 17.8% | −10.7% |
| F 11; F 12 | IL-1ra/IL-1F3 | 58.9% | −31.1% |
| F 13; F 14 | IL-2 | 19.2% | 0.4% |
| F 15; F 16 | IL-3 | −9.8% | 12.1% |
| F 17; F 18 | IL-4 | −25.4% | 18.2% |
| F 19; F 20 | IL-5 | −0.7% | −10.7% |
| F 21; F 22 | IL-6 | 32.4% | 0.7% |
| F 23; F 24 | IL-7 | 4.4% | −8.8% |
| G 1; G 2 | IL-10 | 6.0% | 3.9% |
| G 3; G 4 | IL-11 | −4.1% | 8.2% |
| G 5; G 6 | IL-12p40 | 3.6% | 6.4% |
| G 7; G 8 | IL-13 | 8.6% | −4.4% |
| G 9; G 10 | IL-15 | 17.8% | −12.5% |
| G 11; G 12 | IL-17A | 22.4% | 12.9% |
| G 13; G 14 | IL-22 | 5.3% | −2.2% |
| G 15; G 16 | IL-23 | 0.3% | 3.1% |
| G 17; G 18 | IL-27p28 | −8.5% | 0.7% |
| G 19; G 20 | IL-28 | −10.8% | −1.9% |
| G 21; G 22 | IL-33 | 20.7% | 26.1% |
| G 23; G 24 | LDL R | 9.6% | −2.5% |
| H 1; H 2 | Leptin | 156.9% | −14.2% |
| H 3; H 4 | LIF | 11.9% | −17.0% |
| H 5; H 6 | Lipocalin-2/NGAL | 97.3% | −34.9% |
| H 7; H 8 | LIX | 31.5% | −29.8% |
| H 9; H 10 | M-CSF | 21.6% | −11.2% |
| H 11; H 12 | MMP-2 | 20.1% | −18.8% |
| H 13; H 14 | MMP-3 | 24.6% | −25.1% |
| H 15; H 16 | MMP-9 | 64.0% | −33.7% |
| H 17; H 18 | Myeloperoxidase | 83.5% | −49.4% |
| H 19; H 20 | Osteopontin (OPN | 62.5% | −17.2% |
| H 21; H 22 | Osteoprotegerin/ TNFRSF11B | 13.2% | −14.3% |
| H 23; H 24 | PD-ECGF/ Thymidine phosphorylase | 13.7% | −2.1% |
| I 1; I 2 | PDGF-BB | 21.4% | −36.0% |
| I 3; I 4 | Pentraxin 2/SAP | 14.3% | −15.2% |
| I 5; I 6 | Pentraxin 3/ TSG-14 | 21.5% | −17.7% |
| I 7; I 8 | Periostin/OSF-2 | 42.9% | −16.2% |
| I 9; I 10 | Pref-1/DLK-1/FA1 | 28.1% | −19.2% |
| I 11; I 12 | Proliferin | 16.8% | −3.7% |
| I 13; I 14 | Proprotein Convertase 9/ PCSK9 | 3.5% | −26.6% |
| I 15; I 16 | RAGE | 16.6% | −16.2% |
| I 17; I 18 | RBP4 | 15.4% | −28.8% |
| I 19; I 20 | Reg3G | 13.9% | −11.2% |
| I 21; I 22 | Resistin | −39.9% | −0.5% |
| J 3; J 4 | E-Selectin/CD62E | 7.8% | −23.9% |
| J 5; J 6 | P-Selectin/CD62P | −14.3% | −7.8% |
| J 7; J 8 | Serpin E1/PAI-1 | 20.4% | −27.4% |
| J 9; J 10 | Serpin F1/PEDF | 4.1% | −29.5% |
| J 11; J 12 | Thrombopoietin | 17.0% | −12.3% |
| J 13; J 14 | TIM-1/KIM-1/ HAVCR | 388.0% | −62.9% |
| J 15; J 16 | TNF-alpha | 32.0% | −24.4% |
| J 17; J 18 | VCAM-1/CD106 | 28.3% | −30.9% |
| J 19; J 20 | VEGF | −3.9% | −11.7% |
| J 21; J 22 | WISP-1/CCN4 | −2.2% | −18.9% |
References
- Lebovitz, H.E. Etiology and pathogenesis of diabetes mellitus. Pediatric Clin. N. Am. 1984, 31, 521–530. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, M.; Xu, D.; Zhang, F.; Yao, B.; Yan, Y.; Zhao, N.; Xu, W.; Qin, G. Body mass index and the risk of mortality among Chinese adults with Type 2 diabetes. Diabet. Med. 2018, 35, 1562–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, S.L. Diabetes and dyslipidaemia. Diabetes Obes. Metab. 2006, 8, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Guo, Z.; Dong, J.; Sheng, S.; Wang, Y.; Yu, L.; Wang, H.; Tang, L. miR-374a regulates inflammatory response in diabetic nephropathy by targeting MCP-1 expression. Front. Pharmacol. 2018, 9, 900. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Long, J.; Jiang, W.; Shi, Y.; He, X.; Zhou, Z.; Li, Y.; Yeung, R.O.; Wang, J.; Matsushita, K.; et al. Trends in Chronic Kidney Disease in China. N. Engl. J. Med. 2016, 375, 905–906. [Google Scholar] [CrossRef]
- Ritz, E.; Zeng, X.X.; Rychlík, I. Clinical manifestation and natural history of diabetic nephropathy. Contrib Nephrol. 2011, 170, 19–27. [Google Scholar]
- Chou, S.-T.; Tseng, S.-T. Oxidative stress markers in type 2 diabetes patients with diabetic nephropathy. Clin. Exp. Nephrol. 2017, 21, 283–292. [Google Scholar] [CrossRef]
- Volpe, C.M.O.; Villar-Delfino, P.H.; Dos Anjos, P.M.F.; Nogueira-Machado, J.A. Cellular death, reactive oxygen species (ROS) and diabetic complications. Cell Death Dis. 2018, 9, 119. [Google Scholar] [CrossRef]
- Wada, J.; Makino, H. Inflammation and the pathogenesis of diabetic nephropathy. Clin. Sci. 2013, 124, 139–152. [Google Scholar] [CrossRef]
- Sancar-Bas, S.; Gezginci-Oktayoglu, S.; Bolkent, S. Exendin-4 attenuates renal tubular injury by decreasing oxidative stress and inflammation in streptozotocin-induced diabetic mice. Growth Factors 2015, 33, 419–429. [Google Scholar] [CrossRef]
- Banba, N.; Nakamura, T.; Matsumura, M.; Kuroda, H.; Hattori, Y.; Kasai, K. Possible relationship of monocyte chemoattractant protein-1 with diabetic nephropathy. Kidney Int. 2000, 58, 684–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Waili, N.; Al-Waili, H.; Al-Waili, T.; Salom, K. Natural antioxidants in the treatment and prevention of diabetic nephropathy; a potential approach that warrants clinical trials. Redox Rep. 2017, 22, 99–118. [Google Scholar] [CrossRef] [PubMed]
- Brenneman, J.; Hill, J.; Pullen, S. Emerging therapeutics for the treatment of diabetic nephropathy. Bioorg. Med. Chem. Lett. 2016, 26, 4394–4402. [Google Scholar] [CrossRef] [PubMed]
- Merrill, J.P. Dialysis versus transplantation in the treatment of end-stage renal disease. Annu. Rev. Med. 1978, 29, 343–358. [Google Scholar] [CrossRef]
- Hariharan, S.; Smith, R.D.; Viero, R.; First, M.R. DIABETIC NEPHROPATHY AFTER RENAL TRANSPLANTATION: Clinical and Pathologic Features1. Transplantation 1996, 62, 632–635. [Google Scholar] [CrossRef]
- Rhee, S.Y.; Kim, H.J.; Ko, S.-H.; Hur, K.-Y.; Kim, N.-H.; Moon, M.K.; Park, S.-O.; Lee, B.-W.; Choi, K.M.; Kim, J.H. Monotherapy in patients with type 2 diabetes mellitus. Diabetes Metab. J. 2017, 41, 349–356. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, Y.; Ke, Y.; Liu, Y.; Luo, X.; Li, C.; Zhang, Z.; Liu, A.; Shen, L.; Chen, H. Antidiabetic activity of polysaccharides from Suillellus luridus in streptozotocin-induced diabetic mice. Int. J. Biol. Macromol. 2018, 119, 134–140. [Google Scholar] [CrossRef]
- Yang, S.; Yan, J.; Yang, L.; Meng, Y.; Wang, N.; He, C.; Fan, Y.; Zhou, Y. Alkali-soluble polysaccharides from mushroom fruiting bodies improve insulin resistance. Int. J. Biol. Macromol. 2019, 126, 466–474. [Google Scholar] [CrossRef]
- Wang, Y.; Lai, L.; Teng, L.; Li, Y.; Cheng, J.; Chen, J.; Deng, C. Mechanism of the anti-inflammatory activity by a polysaccharide from Dictyophora indusiata in lipopolysaccharide-stimulated macrophages. Int. J. Biol. Macromol. 2019, 126, 1158–1166. [Google Scholar] [CrossRef]
- Wang, J.; Hu, W.; Li, L.; Huang, X.; Liu, Y.; Wang, D.; Teng, L. Antidiabetic activities of polysaccharides separated from Inonotus obliquus via the modulation of oxidative stress in mice with streptozotocin-induced diabetes. PLoS ONE 2017, 12, e0180476. [Google Scholar] [CrossRef]
- Jiang, X.; Teng, S.; Wang, X.; Li, S.; Zhang, Y.; Wang, D. The Antidiabetic and Antinephritic Activities of Tuber melanosporum via Modulation of Nrf2-Mediated Oxidative Stress in the db/db Mouse. Oxidative Med. Cell. Longev. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Song, J.; Wang, D.; Zhang, N.; Lu, J.; Meng, Q.; Zhou, Y.; Wang, N.; Liu, Y.; Wang, D.; et al. The anti-membranous glomerulonephritic activity of purified polysaccharides from Irpex lacteus Fr. Int. J. Biol. Macromol. 2016, 84, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Ying, T. Isolation, molecular characterization and antioxidant activity of a water-soluble polysaccharide extracted from the fruiting body of Termitornyces albuminosus (Berk.) Heim. Int. J. Biol. Macromol. 2019, 122, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, S.; Zhang, J.; Che, G.; Zhou, M.; Liu, M.; Zhang, C.; Xu, N.; Lin, L.; Liu, Y.; et al. The antihyperlipidemic activities of enzymatic and acidic intracellular polysaccharides by Termitomyces albuminosus. Carbohydr. Polym. 2016, 151, 1227–1234. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, J.; Zhang, J.; Wang, X.; Liu, M.; Zhang, C.; Jia, L. Hepatoprotective and in vitro antioxidant effects of native depolymerised-exopolysaccharides derived from Termitomyces albuminosus. Sci. Rep. 2017, 7, 3910. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, E.A.; El Enshasy, H.; Wadaan, M.A.; Aziz, R. Mushrooms: A potential natural source of anti-inflammatory compounds for medical applications. Mediat. Inflamm. 2014, 2014, 805841. [Google Scholar] [CrossRef] [PubMed]
- Östenson, C.-G.; Khan, A.; Abdel-Halim, S.; Guenifi, A.; Suzuki, K.; Goto, Y.; Efendic, S. Abnormal insulin secretion and glucose metabolism in pancreatic islets from the spontaneously diabetic GK rat. Diabetologia 1993, 36, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Tung, Y.T.; Chen, H.L.; Wu, H.S.; Ho, M.H.; Chong, K.Y.; Chen, C.M. Kefir Peptides Prevent Hyperlipidemia and Obesity in High-Fat-Diet-Induced Obese Rats via Lipid Metabolism Modulation. Mol. Nutr. Food Res. 2018, 62. [Google Scholar] [CrossRef]
- Song, Q.; Zhang, Y.; Zhang, Y.; Zhang, X.; Gao, Y.; Zhang, J.; Chu, L.; Zhao, S. Potential mechanisms underlying the protective effects of salvianic acid A against atherosclerosis in vivo and vitro. Biomed. Pharmacother. 2019, 109, 945–956. [Google Scholar] [CrossRef]
- Al Hroob, A.M.; Abukhalil, M.H.; Alghonmeen, R.D.; Mahmoud, A.M. Ginger alleviates hyperglycemia-induced oxidative stress, inflammation and apoptosis and protects rats against diabetic nephropathy. Biomed. Pharmacother. 2018, 106, 381–389. [Google Scholar] [CrossRef]
- Dzydzan, O.; Bila, I.; Kucharska, A.Z.; Brodyak, I.; Sybirna, N. Antidiabetic effects of extracts of red and yellow fruits of cornelian cherries (Cornus mas L.) on rats with streptozotocin-induced diabetes mellitus. Food Funct. 2019. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.S.; Khosravi, M.; Grocott, M.P.; Mythen, M.G. Concepts in hypoxia reborn. Crit. Care 2010, 14, 315. [Google Scholar] [CrossRef] [PubMed]
- Gerber, P.A.; Rutter, G.A. The role of oxidative stress and hypoxia in pancreatic beta-cell dysfunction in diabetes mellitus. Antioxid. Redox Signal. 2017, 26, 501–518. [Google Scholar] [CrossRef] [PubMed]
- Ouriel, K.; Smedira, N.G.; Ricotta, J.J. Protection of the kidney after temporary ischemia: Free radical scavengers. J. Vasc. Surg. 1985, 2, 49–53. [Google Scholar] [CrossRef] [Green Version]
- Hasnain, S.Z.; Prins, J.B.; McGuckin, M.A. Oxidative and endoplasmic reticulum stress in β-cell dysfunction in diabetes. J. Mol. Endocrinol. 2016, 56, R33–R54. [Google Scholar] [CrossRef]
- Insel, P.A.; Liljenquist, J.E.; Tobin, J.D.; Sherwin, R.S.; Watkins, P.; Andres, R.; Berman, M. Insulin control of glucose metabolism in man: A new kinetic analysis. J. Clin. Investig. 1975, 55, 1057–1066. [Google Scholar] [CrossRef]
- Ashcroft, F.M.; Rohm, M.; Clark, A.; Brereton, M.F. Is type 2 diabetes a glycogen storage disease of pancreatic β cells? Cell Metab. 2017, 26, 17–23. [Google Scholar] [CrossRef]
- Tangvarasittichai, S. Oxidative stress, insulin resistance, dyslipidemia and type 2 diabetes mellitus. World J. Diabetes 2015, 6, 456–480. [Google Scholar] [CrossRef]
- Lomonaco, R.; Chen, J.; Cusi, K. An endocrine perspective of nonalcoholic fatty liver disease (NAFLD). Ther. Adv. Endocrinol. Metab. 2011, 2, 211–225. [Google Scholar] [CrossRef]
- Palazhy, S.; Viswanathan, V. Lipid Abnormalities in Type 2 Diabetes Mellitus Patients with Overt Nephropathy. Diabetes Metab. J. 2017, 41, 128–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, M.L.; Lu, X.M.; Cong, W.T.; Xing, X.; Tan, Y.; Li, Y.Q.; Li, X.K.; Jin, L.T.; Wang, X.J.; Dong, J.C.; et al. Multiple Low-Dose Radiation Prevents Type 2 Diabetes-Induced Renal Damage through Attenuation of Dyslipidemia and Insulin Resistance and Subsequent Renal Inflammation and Oxidative Stress. PLoS ONE 2014, 9, 17. [Google Scholar] [CrossRef] [PubMed]
- Arango Duque, G.; Descoteaux, A. Macrophage cytokines: Involvement in immunity and infectious diseases. Front. Immunol. 2014, 5, 491. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Yan, X.; Nagata, N.; Aritake, K.; Katsumata, Y.; Matsuhashi, T.; Nakamura, M.; Hirai, H.; Urade, Y.; Asano, K. PGD2-CRTH2 pathway promotes tubulointerstitial fibrosis. J. Am. Soc. Nephrol. 2012, 23, 1797–1809. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, T. The hunt for the source of primary interleukin-4: How we discovered that natural killer t cells and basophils determine T helper type 2 cell differentiation in vivo. Front. Immunol. 2018, 9, 716. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-C.; Sytwu, H.-K.; Lu, K.-C.; Lin, Y.-F. Role of T cells in type 2 diabetic nephropathy. Exp. Diabetes Res. 2011, 2011. [Google Scholar] [CrossRef]
- Chow, F.; Ozols, E.; Nikolic-Paterson, D.J.; Atkins, R.C.; Tesch, G.H. Macrophages in mouse type 2 diabetic nephropathy: Correlation with diabetic state and progressive renal injury. Kidney Int. 2004, 65, 116–128. [Google Scholar] [CrossRef] [Green Version]
- Sangoi, M.B.; de Carvalho, J.A.M.; Tatsch, E.; Hausen, B.S.; Bollick, Y.S.; Londero, S.W.K.; Duarte, T.; Scolari, R.; Duarte, M.; Premaor, M.O.; et al. Urinary inflammatory cytokines as indicators of kidney damage in type 2 diabetic patients. Clin. Chim. Acta 2016, 460, 178–183. [Google Scholar] [CrossRef]
- Wang, H.Y.; Zhao, J.G.; Wei, Z.G.; Zhang, Y.Q. The renal protection of flavonoid-rich ethanolic extract from silkworm green cocoon involves in inhibiting TNF-alpha-p38 MAP kinase signalling pathway in type 2 diabetic mice. Biomed. Pharm. 2019, 118, 7. [Google Scholar] [CrossRef]
- Mitchell, A.M.; Christian, L.M. Repetitive negative thinking, meaning in life, and serum cytokine levels in pregnant women: Varying associations by socioeconomic status. J. Behav. Med. 2019, 42, 960–972. [Google Scholar] [CrossRef]
- Manna, K.; Mishra, S.; Saha, M.; Mahapatra, S.; Saha, C.; Yenge, G.; Gaikwad, N.; Pal, R.; Oulkar, D.; Banerjee, K.; et al. Amelioration of diabetic nephropathy using pomegranate peel extract-stabilized gold nanoparticles: Assessment of NF-kappaB and Nrf2 signaling system. Int. J. Nanomed. 2019, 14, 1753–1777. [Google Scholar] [CrossRef]
- Jha, J.C.; Banal, C.; Chow, B.S.M.; Cooper, M.E.; Jandeleit-Dahm, K. Diabetes and Kidney Disease: Role of Oxidative Stress. Antioxid. Redox. Signal 2016, 25, 657–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, G.P.; Mijares, M.R.; Chavez, K.; Suarez, A.I.; Compagnone, R.S.; Chirinos, P.; De Sanctis, J.B. Caracasine acid, an ent-3,4-seco-kaurene, promotes apoptosis and cell differentiation through NFkB signal pathway inhibition in leukemia cells. Eur. J. Pharm. 2019, 862, 172624. [Google Scholar] [CrossRef] [PubMed]
- El-Mesery, M.; Rosenthal, T.; Rauert-Wunderlich, H.; Schreder, M.; Stuhmer, T.; Leich, E.; Schlosser, A.; Ehrenschwender, M.; Wajant, H.; Siegmund, D. The NEDD8-activating enzyme inhibitor MLN4924 sensitizes a TNFR1(+) subgroup of multiple myeloma cells for TNF-induced cell death. Cell Death Dis. 2019, 10, 611. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Jiang, X.; Teng, S.S.; Zhang, Y.Q.; Liu, Y.; Li, X.; Li, Y. The Antidiabetic and Antinephritic Activities of Auricularia cornea (An Albino Mutant Strain) via Modulation of Oxidative Stress in the db/db Mice. Front. Immunol. 2019, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Sui, Y.; Liu, W.; Tian, W.; Li, X.Q.; Cao, W. A branched arabinoglucan from Angelica sinensis ameliorates diabetic renal damage in rats. Phytother. Res. 2019, 33, 818–831. [Google Scholar] [CrossRef] [PubMed]
- Kou, L.; Du, M.; Liu, P.; Zhang, B.; Zhang, Y.; Yang, P.; Shang, M.; Wang, X. Anti-diabetic and anti-nephritic activities of Grifola frondosa mycelium polysaccharides in diet-streptozotocin-induced diabetic rats via modulation on oxidative stress. Appl. Biochem. Biotechnol. 2019, 187, 310–322. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.W.; Wu, C.Y.; Cheng, H.T. Merit of Astragalus polysaccharide in the improvement of early diabetic nephropathy with an effect on mRNA expressions of NF-kappa B and I kappa B in renal cortex of streptozotoxin-induced diabetic rats. J. Ethnopharmacol. 2007, 114, 387–392. [Google Scholar] [CrossRef]


| Kidney (pg/mgprot) | db/m+ | db/db | db/db | ||
|---|---|---|---|---|---|
| Met (0.1g/kg) | PTA (0.025g/kg) | PTA (0.1g/kg) | |||
| IL-2 | 54.8 ± 2.5 | 72.4 ± 9.3## | 59.7 ± 6.9* | 53.6 ± 3.9** | 59.4 ± 7.0* |
| IL-4 | 2.7 ± 0.1 | 2.4 ± 0.1# | 2.9 ± 0.3** | 2.8 ± 0.3** | 3.0 ± 0.5** |
| IL-12 | 0.80 ± 0.08 | 1.35 ± 0.14### | 0.93 ± 0.11*** | 0.96 ± 0.09*** | 1.05 ± 0.06** |
| TNF-α | 100.2 ± 6.7 | 132.7 ± 11.7## | 116.1 ± 14.0 | 100.7 ± 10.0** | 102.4 ± 4.3** |
| LEP | 0.66 ± 0.05 | 0.37 ± 0.02### | 0.65 ± 0.04*** | 0.65 ± 0.09*** | 0.66 ± 0.09*** |
| M-CSF | 21.2 ± 1.9 | 26.1 ± 0.6## | 20.9 ± 1.6** | 18.9 ± 2.2*** | 20.9 ± 1.9** |
| IFN-γ | 2.5 ± 0.3 | 3.4 ± 0.2## | 2.4 ± 0.2*** | 2.2 ± 0.1*** | 2.7 ± 0.2** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.; Feng, Q.; Liao, H.; Yu, X.; Liu, Y.; Wang, D. Anti-Diabetic Nephropathy Activities of Polysaccharides Obtained from Termitornyces albuminosus via Regulation of NF-κB Signaling in db/db Mice. Int. J. Mol. Sci. 2019, 20, 5205. https://doi.org/10.3390/ijms20205205
Yang C, Feng Q, Liao H, Yu X, Liu Y, Wang D. Anti-Diabetic Nephropathy Activities of Polysaccharides Obtained from Termitornyces albuminosus via Regulation of NF-κB Signaling in db/db Mice. International Journal of Molecular Sciences. 2019; 20(20):5205. https://doi.org/10.3390/ijms20205205
Chicago/Turabian StyleYang, Chang, Qi Feng, Huan Liao, Xinlei Yu, Yang Liu, and Di Wang. 2019. "Anti-Diabetic Nephropathy Activities of Polysaccharides Obtained from Termitornyces albuminosus via Regulation of NF-κB Signaling in db/db Mice" International Journal of Molecular Sciences 20, no. 20: 5205. https://doi.org/10.3390/ijms20205205
APA StyleYang, C., Feng, Q., Liao, H., Yu, X., Liu, Y., & Wang, D. (2019). Anti-Diabetic Nephropathy Activities of Polysaccharides Obtained from Termitornyces albuminosus via Regulation of NF-κB Signaling in db/db Mice. International Journal of Molecular Sciences, 20(20), 5205. https://doi.org/10.3390/ijms20205205