Abstract
One of the main mechanisms carried out by the cells to counteract several forms of stress is the activation of the nuclear factor erythroid 2-related factor (Nrf2) signaling. Nrf2 signaling controls the expression of many genes through the binding of a specific cis-acting element known as the antioxidant response element (ARE). Activation of Nrf2/ARE signaling can mitigate several pathologic mechanisms associated with an autoimmune response, digestive and metabolic disorders, as well as respiratory, cardiovascular, and neurodegenerative diseases. Indeed, several studies have demonstrated that Nrf2 pathway plays a key role in inflammation and in cancer development in many organs, including the intestine. Nrf2 appears to be involved in inflammatory bowel disease (IBD), an immune-mediated chronic and disabling disease, with a high risk of developing intestinal fibrotic strictures and cancer. Currently, drugs able to increase cytoprotective Nrf2 function are in clinical trials or already being used in clinical practice to reduce the progression of some degenerative conditions. The role of Nrf2 in cancer development and progression is controversial, and drugs able to inhibit abnormal levels of Nrf2 are also under investigation. The goal of this review is to analyze and discuss Nrf2-dependent signals in the initiation and progression of intestinal fibrosis and cancers occurring in IBD.
Keywords:
inflammatory bowel disease; IBD; fibrosis; intestinal fibrosis; cancer; colorectal cancer; Nrf2 1. Introduction
Inflammatory bowel disease (IBD) is a spectrum of diseases, including Crohn’s disease (CD) and ulcerative colitis (UC), that lead to numerous complications, among which the most common is represented by intestinal fibrosis and cancer. Intestinal fibrosis is a process characterized by excessive deposition of extracellular matrix (ECM) proteins by activated myofibroblasts [1,2,3].
Intestinal fibrogenesis is driven by complex mechanisms since in the gut there are many cell types (as fibroblasts, subepithelial myofibroblasts, smooth muscle cells, epithelial and endothelial cells) that may become activated ECM-producing myofibroblasts. In fibrosis, the abnormal accumulation of ECM can be determined either by its excessive production by activated myofibroblasts or by its reduced degradation. Physiologically, the degradation of ECM components is regulated by the delicate equilibrium between proteolytic enzymes, like matrix metalloproteinases (MMPs) and tissue inhibitors of MMP (TIMPs). Perturbation of the delicate MMP/TIMP balance results in the excessive deposition of ECM proteins, which consequently leads to fibrosis.
A pivotal role in intestinal fibrosis, other than in many types of cancer, is certainly played by transforming growth factor β (TGF-β), not only through its canonical interaction with small mothers against decapentaplegic proteins (Smads) but also by a complex network with other profibrotic and antifibrotic molecules [1,4,5,6]. Recently, several studies have focused their attention on Nrf2, an important transcription factor able to interact with TGF-β and that appears to be involved in the fibrogenesis process in many organs, including the bowel [7,8,9,10,11].
Additionally, although the exact mechanism of action seems still controversial, it was established that Nrf2 participates in the development, progression, and metastasis of several tumors, including colorectal cancer [12,13,14,15,16,17,18].
Understanding the exact role of Nrf2 signaling in the pathophysiology of these diseases could be crucial in developing new effective drugs [19,20,21].
Among the multiple aspects of Nrf2, in this review, we discuss its role in intestinal fibrosis and in colorectal cancer occurring during IBD and the possibility that it may be a new and valid therapeutic target.
2. What Is Nrf2?
Nowadays, the human body is repeatedly exposed to many external stimuli, such as environmental pollens, food additives, drugs, ultraviolet light, and ionizing radiation. In addition to these exogenous stresses, cells are also subject to biological insults due to free radicals, reactive oxygen species (ROS), and reactive nitrogen species (RNS) [22,23,24].
Under normal conditions, these molecules are synthesized in mitochondria, peroxisomes, and endoplasmic reticulum at physiological concentrations, while during pathological processes, they are released at high levels into the vascular system, becoming toxic [24] and leading to specific dysfunctions such as inflammation, autoimmune reaction, and cancer, as well as cardiovascular, airway, digestive, metabolic, and neurodegenerative diseases [23]. In order to neutralize these harmful effects, cells have developed a wide range of defensive mechanisms, including the activation of Nrf2 [24,25,26].
In 1994, Moi et al. identified Nrf2 as a transcription factor of beta-globin gene and, thereafter, several studies have revealed its key role in the regulation of many detoxifying enzymes, appearing to be a main player of the cytoprotective response against oxidative stress [26,27,28]. Nrf2 is a member of the cap’n’collar subfamily of basic region leucine zipper transcription factors that together with Nrf1, Nrf3, and NF-E2 p45 subunit, is able to regulate gene expression [22,24]. Under homeostatic conditions, Nrf2 transcriptional activity is mediated by the binding with its inhibitor Kelch-like ECH-associated protein 1 (Keap1) which mediates the interaction between Nrf2 and Cul3-based E3-ubiquitin ligase complex, leading to continuous ubiquitination and proteasome degradation of Nrf2. This mechanism ensures constant levels of Nrf2 in the cells (Figure 1) [22,25]. Instead, in response to repetitive endogens and exogenous stimuli, Keap1 inhibition fails and Nrf2 translocate from the cytoplasm to the nucleus where dimerizes with members of the musculoaponeurotic fibrosarcoma proteins (MAF). The heterodimer recognizes a specific sequence known as the antioxidant responsive element (ARE), situated in the promoter region of Nrf-2 target genes, and activates their transcription (Figure 1).
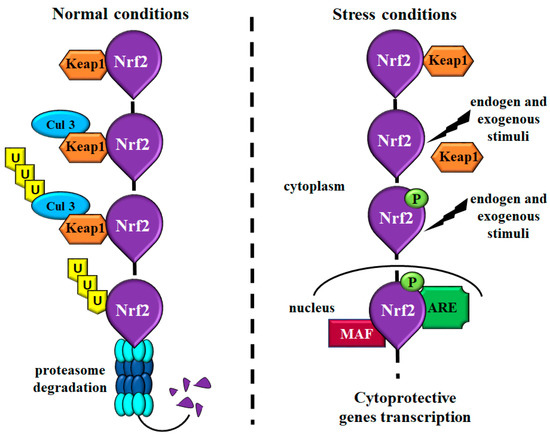
Figure 1.
Schematic representation showing the main Nrf2 molecular mechanisms. Under normal condition, Nrf2 binds its inhibitor Keap1 that mediates the interaction with Cul3 leading to Nrf2 ubiquitination and proteasome degradation. These events provide a regulated modulation of Nrf2 levels in the cells. Under stress conditions and in response to different insults, Keap1 is inactivated, and Nrf2 is phosphorylated and translocated into the nucleus. In the nucleus, Nrf2 binds MAF proteins and, consequently, ARE sequences inducing the transcription of cytoprotective genes. Abbreviations: Cul3 = Cul3-based E3-ubiquitin ligase; Keap1 = Kelch-like ECH-associated protein 1; Nrf2 = nuclear factor-erythroid 2-related factor; MAF = musculoaponeurotic fibrosarcoma proteins; ARE = antioxidant responsive element; P = phosphorylated.
Initially, ARE was described as a regulatory element for a limited number of genes such as NAD(P)H dehydrogenase-(quinone 1) and glutathione transferase, but new functions of Nrf2 are constantly arising [29,30,31,32,33,34,35].
Currently, it is well known that there are several proteins encoded by ARE genes, including those involved in drug and xenobiotic detoxification and in oxidative stress response. In cellular redox homeostasis, Nrf2 can regulate critical functions such as biosynthesis of glutathione (GSH), thioredoxin (TXN), in addition to the regeneration of nicotinamide adenine dinucleotide phosphate oxidase (NADPH) and the production of ROS. Nrf2 could have a protective role in many autoimmune diseases (i.e., vitiligo, asthma, multiple sclerosis, and systemic lupus erythematosus) since an increased production of reactive immunogenic macromolecules intermediates in these disorders seems closely linked to the loss of several phase II detoxification enzymes regulated by the activation of Nrf2 genes [23,36].
Additionally, Nrf2/ARE signaling participates in heme metabolism, cellular redox homeostasis, the control of inflammatory responses, tissue remodeling, fibrosis, carcinogenesis, and metastasis [37,38].
3. Role of Nrf2 in Inflammation and Fibrosis in Various Organs
Several studies have demonstrated that Keap1/Nrf2 signaling plays a pivotal role in the attenuation of both acute and chronic inflammation in several diseases (i.e., rheumatoid arthritis, asthma, emphysema, gastritis, and atherosclerosis) through its ability to normalize mitochondrial function, restore redox homeostasis, and suppress the production of inflammatory mediators [39,40,41,42,43,44].
Particularly, in an early phase of the inflammatory process, the Nrf2/ARE pathway seems to exert an inhibitory effect in the production of pro-inflammatory molecules such as cytokines, chemokines, cell adhesion proteins, MMPs, cyclooxygenase-2 (COX-2), and prostaglandins [45,46,47,48,49,50].
Due to the intimate link between inflammation and fibrosis, several studies have analyzed the potential role of Nrf2 to mitigate the release of several inflammatory molecules involved in abnormal collagen syntheses and deposition [51,52,53,54,55,56,57,58] (Table 1).

Table 1.
Studies assessing Nrf2 involvement in inflammation and fibrosis in various organs.
4. Role of Nrf2 Signaling in Intestinal Inflammation and Fibrosis
After an injury, the organism activates a sequence of events to maintain the integrity of tissue morphology and functionality: (1) recruitment of inflammatory cells, (2) release of fibrogenic cytokines and, lastly, (3) activation of ECM-depositing cells [59].
Inflammation is the first biological response to adverse stimuli, such as pathogens (i.e., bacteria and viruses) and external damage (i.e., tissue scrapes and effects of chemicals and radiations) to limit and sometimes to eliminate the causes of cellular injuries. According to the type of cells involved and the injury duration, the inflammatory process can be classified as acute (rapidly and self-limiting response) or chronic (response prolonged for weeks, months or, in a specific scenario, for a lifetime). Protracted insults can lead to abnormal deposition of ECM proteins and consequently to a progressive pathological condition such as fibrosis. However, although several studies have tried to clarify the relationship between inflammation and fibrosis in IBD, the precise pathophysiological mechanism is still unknown and no effective drugs, as alternative to surgery, are available when stenosis has developed. In an early phase, inflammatory cells are recruited into the injury site to restore organ integrity and function. However, the inflammatory response protraction over time becomes chronic, leading to myofibroblast activation and uncontrollable ECM deposition and fibrosis [1,4,6]. It is well known that the key regulator of intestinal fibrosis is TGF-β, which exerts its function through its canonical (Smads) and non-canonical (i.e., mitogen-activated protein kinase (MAPK), sphingosine 1-phosphate, and mammalian target of rapamycin) pathways [60,61].
Moreover, recent findings have highlighted other crucial molecules able to interact with TGF-β and involved in the development of intestinal fibrosis, such as Nrf2 and its downstream genes (Table 2).
Table 2.
Studies assessing Nrf2 involvement in intestinal inflammation and fibrosis.
4.1. Nrf2 in Experimental Models of Colitis Induced in Transgenic Mice
In 2006, for the first time, Khor et al. showed in an experimental dextran sulfate sodium (DSS)-induced colitis model that Nrf2 knockout (KO) mice (Nrf2−/−) were more susceptible to acquiring histological features of colitis (such as a short length of the colon and rectal bleeding) compared to Nrf2 wild-type (WT) mice (Nrf2+/+). In the same study, overexpression of many antioxidant and phase II detoxifying enzymes (heme oxygenase-1, UDP-glucuronosyltransferase 1A1, and glutathione S-transferase Mu-1) was associated with an increase in Nrf2 levels in DSS WT mice compared to KO mice. Depletion of Nrf2 reduced the mRNA and protein expression of these enzymes and contributed to an increased level of pro-inflammatory mediators, supporting the hypothesis that Nrf2 plays a significant role in colonic damage [62].
Remarkably, the role of the Nrf2/inflammation axis in the progression of DSS-induced colitis has been demonstrated by genetic mouse experiments. In fact, Nrf2−/− KO mice challenged by DSS presented more severe histopathological signs of colitis, showing a high expression of COX-2, HO-1, and γ-glutamylcysteine synthetase (γ-GCS) compared to WT controls. On the other side, DSS-induced colitis in COX-2−/− KO mice was less severe compared to WT control mice, showing a decrease of heme oxygenase-1 (HO-1) and increase of NADPH quinone oxidoreductase-1 (NQO-1), demonstrating the tight association between inflammation and the antioxidant system [63].
Interestingly, in the same year, Li et al. found that high expression of Nrf2 and HO-1 were correlated with the level of mitogen-activated protein kinase phosphatase-1 (Mkp-1), not only in the colon of mice with DSS-induced colitis but even in colorectal biopsies of patients affected by UC and CD. Moreover, Mkp-1−/− mice were more sensitive to DSS-induced colitis, showing severe signs of inflammation. The authors concluded that Mkp-1 and Nrf2/HO-1 create a protective function against inflammation [64].
It is known that during an inflammatory process, a range of damage-sensing receptors—including the inflammasome multiprotein complexes NOD-like receptor (NLR) family—are activated. Among these receptors, the most studied is NLPR3 and its dysregulation seems to contribute to IBD development [65]. In DSS-induced colitis using NLPR3−/− KO mice, Wang et al. analyzed the anti-inflammatory effect of a novel Nrf2/ARE inducer, called compound 1. This molecule was able to reduce colorectal inflammation in DSS-induced colitis in NLPR3+/+ WT mice while it did not exert anti-inflammatory effects in DSS-induced colitis in NLPR3−/− KO animals. This evidence highlighted that the protective effect of compound 1 was due to its inhibitory action on NLRP3 inflammasome and, concomitantly, Nrf2 activation [65].
4.2. Nrf2 Inhibition in Experimental Models of Colitis
Several studies revealed that heme oxygenase, an important enzyme regulated by Nrf2, ameliorated inflammation in many experimental models [66,67,68,69] although its specific role in IBD remains unclear.
To clarify this mechanism, Wang et al. analyzed the HO-1 involvement on trinitrobenzene sulfonic acid (TNBS)-induced colitis in mice. HO activity and HO-1 gene expression were significantly higher in mice receiving TNBS compared to controls. Administration of mesoporphyrin (SnMP), an HO inhibitor, potentiated the colonic damage along with a reduction in HO-1 activity. Furthermore, the reduction of HO-1 expression by SnMP also enhanced ROS and inducible nitric oxide synthase (iNOS) expression, both of which were dramatically increased after the TNBS enema. These results indicate a protective role of HO-1 and Nrf2 in TNBS-induced colitis by decreasing free radical production and inhibiting iNOS expression in colonic mucosa [70].
More recent investigations conducted both in vivo (by DSS-induced colitis as an experimental model of IBD) and in vitro (by colonic cells NCM460) revealed that CPUY192018, an inhibitor of the Keap1–Nrf2 interaction, reduced the expression of inflammatory cytokines and exerted a cytoprotective effect. These data suggested that molecules able to regulate Keap1–Nrf2–ARE signaling alleviated features of experimental colitis and could represent a new therapeutic strategy for IBD [71].
4.3. Nrf2 Activation in Experimental Models of Colitis
In an in vivo study of acetic acid-induced colitis in rats, Yalniz et al. demonstrated the ability of nadroparin sodium to prevent and attenuate the expression of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-kB). The authors demonstrated that anti-inflammatory and antioxidative effects of nadroparin were mediated by Nrf2/HO-1/NF-kB pathways, showing that Nrf2/ARE signaling was involved in intestinal inflammatory processes [72].
In DSS-induced colitis, Liu et al. analyzed the effects of dimethyl fumarate (DMF), an immunomodulatory and anti-inflammatory drug currently used for relapsing forms of multiple sclerosis. The authors showed that DMF administration attenuated signs of DSS damage (i.e., body weight loss, colon length reduction), activated Nrf2 and its downstream genes, and inhibited pro-inflammatory cytokines and the NLRP3 inflammasome. Due to the ability of DMF to interfere with Nrf2 signaling, these results demonstrated a potential use of this drug also in the treatment of colitis [73].
Recently, in a mouse model of DSS-induced colitis, Yang et al. evaluated the impact of hyperoside (Hyp), a flavonoid with anti-inflammatory, antiapoptotic, and antioxidant proprieties. The results showed that Hyp reduced the expression of pro-inflammatory cytokines such as TNF-α, interleukin-6, COX-2, and NF-kB and increased the levels of anti-inflammatory cytokines such as interleukin-10 [74]. Additionally, the levels of pro-apoptotic proteins such as caspase-3 and Bax were reduced while antiapoptotic proteins, such as Bcl2, were overexpressed. The study demonstrated that Hyp ameliorates colitis by inducing expression of Nrf2 and its target genes. This effect was probably due to the ability of this substance to attenuate the inflammatory process and apoptosis through the activation of Nrf2 signaling [74].
Park et al. demonstrated the anti-inflammatory and antioxidative ability of a natural herb known as Perilla frutescens (PF). In DSS-induced colitis in mice, PF administration improved the features of colitis and reduced the expression of several pro-inflammatory molecules. PF was also able to inhibit the activation of NF-kB and STAT3 and, at the same time, to increase Nrf2 and HO-1 in the colon, displaying the protective effect of Nrf2 [75].
Another molecular partner of Nrf2 has been revealed by Jing et al. The study demonstrated that the features of colitis in DSS-treated rats were improved by administration of berberine, an alkaloid extracted from Berberis species. Berberine upregulated Nrf2 expression and increased P-glycoprotein (P-gp) both in vivo and in vitro in Caco-2 cells. Nrf2 silencing in Caco-2 cells abolished the upregulation of P-gp and, concomitantly, the beneficial effect of berberine [76]. Similar protective effects, through activation of Nrf2 signaling in DSS-induced colitis in mice, were described for luteolin, oligonol, and sinomenine, which were able to activate also the Nrf2-downstream partner NQO-1 [77,78,79]. Particularly, Li et al. revealed that luteolin was able to mitigate signs of fibrosis as well as the expression of inflammatory mediators (iNOS, TNF-α, IL-6) by activating Nrf2 and its downstream genes (HO-1 and NQO-1) [77]. Comparable results were found with the oligonol. This compound, by increasing Nrf2 action and the activation of HO-1 and NQO-1, leads to a decrease of IL-1, IL-6, TNF-α, NF-kB, c-Fos, and c-Jun [78]. In another study, Zhuo et al. highlighted the ability of sinomenine to improve DSS-induced colitis, interfering with the Nrf2/NQO-1 signaling and reducing the levels of pro-inflammatory and pro-fibrotic molecules (TNF-α, IL-6, and iNOS) [79]. Similarly, Gao et al. showed the protective role of the LL202, a synthetic flavonoid derivate, which, especially in macrophages, stimulates the Nrf2/HO-1 pathway in both TNBS and DSS-induced colitis [80]. This study also revealed that this natural compound reduced the expression of pro-inflammatory molecules through the activation of Nrf2/HO-1 signaling, amplifying the Nrf2 antioxidant effect [80].
Saber and colleagues explored the effect of olmesartan, an angiotensin II receptor type 1 blocker, in acetic acid-induced colitis in rats. The drug acted as an Nrf2 activator, NF-kB inhibitor, and apoptosis inhibitor [81]. Tussilagone, a molecule able to inhibit NF-κB activation and to induce Nrf2, reduced the main markers of murine colitis induced by DSS (i.e., TNF-α and IL-6) suggesting its potential role in the treatment of colitis [82].
Also, plants typical of Mediterranean scrubs, such as rosemary (Rosmarinus officinalis) and common sage (Salvia officinalis) and synthesized compounds such as carnosol [83] and carnosic acid [84] showed antioxidant and protective effects in both in vitro (colon cells line) and in vivo (DSS-induced colitis in mice) studies. Both carnosol and carnosic acid increased Nrf2 expression.
Sangaraju et al. demonstrated that the phytochemical galangin (GAL) was able to ameliorate DSS-induced colitis in BALB/c mice. GAL administration in DSS colitis in mice induced an increased level of anti-inflammatory cytokines (IL-10) and decreased levels of pro-inflammatory mediators (TNF, IL-6, and iNOS). The study also indicated a protective effect of GAL against intestinal fibrosis by the activation of HO-1 and, consequently, of Nrf2 [85].
A recent paper supported the role of Nrf2 in intestinal fibrosis through the evaluation of apocynin, an NADPH oxidase inhibitor, in DSS-induced colitis in mice. In fact, the recovery of colon length and weight, as well as a decrease in the number of inflammatory foci, was observed in DSS mice upon treatment with apocynin. Moreover, the expression of iNOS, COX-2, TNF-α, and monocyte chemoattractant protein-1 (MCP-1) were decreased in DSS mice treated with apocynin. In the same group, Nrf2 and HO-1 were significantly activated, exerting a protective role [86].
The interplay between colitis and oxidative stress has been reported, with a tight correlation between ROS production and the upregulation of Nrf2. A diet low in methionine (an inducer of ROS) reduced fibrotic scores and the inflammatory profile in DSS-induced colitis in mice compared to DSS mice fed with a regular diet [87]. The beneficial effect of natural compounds in ROS inhibition during the progression of colitis was also demonstrated with the use of catechins, of which legumes and tea are rich. These phytochemicals increased the expression of antioxidant molecules interacting with several pathways, including NF-κB, MAPKs, STAT1/3, and of Nrf2, and also modulated the intestinal flora [88]. Comparable effects have been highlighted for licochalcone A, a chalcone isolated from licorice root widely recognized in traditional Chinese medicine. Licochalcone A was able to reduce oxidative stress and inflammation downregulating NF-kB and upregulating Nrf2 [89]. In fact, in DSS-induced colitis in mice, licochalcone administration reduced the histological scores of colitis and the levels of pro-inflammatory cytokines [89].
The Nrf2 involvement in experimental DSS colitis was also investigated by Li et al. and demonstrated the potential therapeutic role of ZnO nanoparticle (ZnONP) in the treatment of colitis in mice. The data highlighted that ZnONP suppressed ROS and pro-inflammatory cytokine production and activated Nrf2 signaling which was then able to carry out its antioxidant and anti-inflammatory functions. Furthermore, unlike 5-aminosalicylic acid (5-ASA), ZnONP was able to restore the changes in colonic bacteria occurring during colonic injuries and to maintain gut homeostasis. The authors suggested that ZnONP combined with 5-ASA, currently used in IBD treatment, could be a novel therapeutic agent in the treatment of colitis [90].
In TNBS-induced colitis model, other investigators demonstrated that the main markers of fibrosis (α-smooth muscle actin, collagen I, TIMP-1, and TGF-β1/Smad signaling) and ROS levels were suppressed by tert-butylhydroquinone (tBHQ), an agonist of Nrf2 [91]. The authors revealed that TNBS-induced intestinal fibrosis was reversed by tBHQ administration through inhibition of the ROS-dependent TGF-β1/Smad pathway [21].
All these findings revealed that, as occurs in other organs, Nrf2/ARE signaling acts both directly and indirectly (manly cooperating with TGF-β/Smad pathway) to reduce inflammatory and fibrotic processes, and could be a novel effective therapeutic target in IBD [21,73,75,90] (Figure 2).
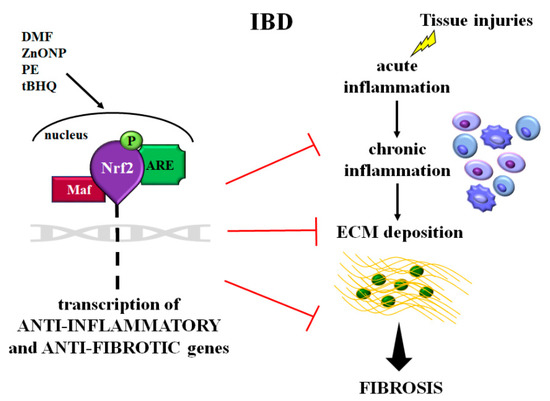
Figure 2.
Schematic representation of the role of Nrf2 in intestinal inflammation and fibrosis occurring in inflammatory bowel disease (IBD). After tissue injuries, the cell’s response is characterized by a cascade of events: acute inflammation, chronic inflammation, and ECM proteins deposition leading to fibrosis. In these pathological events, Nrf2 is activated and then induces the transcription of the anti-inflammatory and antifibrotic genes. Molecules such as DMF, ZnONP, PE, and tBHQ, improving Nrf2 signaling, could be a new therapeutic strategy for intestinal inflammation and fibrosis. Abbreviations: ECM = extracellular matrix; Nrf2 = muclear factor-erythroid 2-related factor; Maf = musculoaponeurotic fibrosarcoma proteins; ARE = antioxidant responsive element; P = phosphorylated; DMF = dimethyl fumarate; ZnONP = ZnO nanoparticle; PE = Perilla frutescens; tBHQ = tert-butylhydroquinone.
5. Role of Nrf2 in the Cancer of Various Organs
The Nrf2 pathway certainly represents one of the main relevant pathways in cell defense and survival signaling, and its involvement in cancer has been extensively investigated in many organs. Data from the literature highlights that this multifunctional molecule exerts a double and controversial role in cancer development [12,13,14,17,18].
On the one hand, in an early stage of the disease, Nrf2 exerts a protective role against chemical-induced carcinogenesis through its ability to reduce ROS accumulation and, consequently, DNA injuries in cells [12,91,92,93,94,95] (Table 3). On the other hand, in the late stage of tumor progression, there is what is called “the dark side” of Nrf2, since it seems to be involved in cancer proliferation, and in chemo- and radioresistance [13,14,17,96,97,98,99,100,101,102] (Table 3).
Table 3.
Studies illustrating positive (advantages) and negative (disadvantages) roles of Nrf2 in cancers of various organs.
6. Role of Nrf2 in IBD-Associated Colorectal Cancer
Colorectal cancer (CRC) is the third most common cancer in men and the second in women worldwide and is the second cause of cancer death in a number of Western countries [103,104,105,106]. CRC is a highly common malignancy in European countries and throughout the world; it has been estimated that 1.13 million new CRC cases are diagnosed every year and that CRC causes about 694,000 deaths per year [106]. CRC is sporadic in 90% of patients; in <10%, it is inherited or is a complication of IBD [107,108].
CRC is linked to a wide range of risk factors such as diet, genetic and epigenetic alteration, immune response, oxidative stress, intestinal microbiota modification, as well as intestinal chronic inflammation [109,110,111,112,113]. Several clinical studies showed that 1 in 6 people affected by ulcerative colitis die of CRC and that chronic inflammation occurring during IBD may increase the risk of CRC development [114,115,116,117,118,119]. Inflammation activates a complex network of cytokines, chemokines, and inflammatory cells (i.e., neutrophils, macrophage, and lymphocytes) generating an environment enriched in ROS and NOS that represents the main factor which contributes to neoplastic transformation in IBD [112,118,120,121,122].
As previously mentioned, several studies showed that Nrf2 plays a “double game” in tumors since, in the first stage of the disease, it exerts a protective role, while in the late stage it may have negative effects in the carcinogenetic process of many organs, including the gut [12,123] (Figure 3) (Table 4).
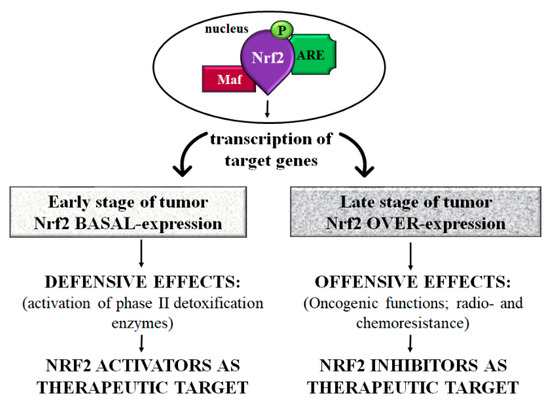
Figure 3.
Scheme of Nrf2 controversial role in colorectal cancer. In the early stage of the tumor, Nrf2 exerts protective effects through the activation of phase II detoxification enzymes. In this step, Nrf2 activators maintaining constant levels of Nrf2 could act as a preventive agent in the development and progression of CRC. On the contrary, at the late stage of the tumor, there is an overexpression of Nrf2 that exerts negative effects characterized by tumor progression and metastasis in addition to radio- and chemoresistance. In this context, using Nrf2 inhibitors to regulate Nrf2 levels might represent a potential adjuvant treatment in CRC. Abbreviations: Nrf2 = nuclear factor-erythroid 2-related factor; Maf = musculoaponeurotic fibrosarcoma proteins; ARE = antioxidant responsive element; P = Phosphorylated.
Table 4.
Studies showing “protective” and “offensive” effects of Nrf2 in colorectal cancer.
6.1. Nrf2 Plays a “Protective Effect” in the Early Stage of Colorectal Cancer
In 2008, Khor et al. used an experimental model of colitis associated with azoxymethane (AOM)/DSS-induced cancer to show that CRC risk is higher in Nrf2 knockout mice. The study highlighted that signs of colitis, as well as the number of tumors per mouse, were increased in Nrf2−/− KO mice compared to wild-type animals. These data demonstrated that Nrf2 was involved not only in protection against inflammation but also against inflammation-associated CRC [12].
Since endogenous estrogen in females has been shown to protect against the development of colon cancer, Song et al. demonstrated that exogenous estrogen replacement in ovariectomized mice showed a protective effect against AOM/DSS-induced colitis and carcinogenesis. The authors revealed that the action of estrogens can be ascribed to a network of estrogen receptors and NF-κB and Nrf2 pathways [124].
Many studies have shown that increased risk of injuries in different organs is correlated to a single-nucleotide polymorphism (SNP) in the promoter region of Nrf2 [43,125,126,127,128]. To clarify the link between Nrf2 and tumor risk, Yokoo and colleagues exposed mice to potassium bromate (KBrO3) to induce neoplastic proliferation comparable to human hereditary colorectal cancer, demonstrating that patients with Nrf2 SNP polymorphisms were more susceptible to colorectal cancer risk. The study showed that the incidence of preneoplastic and neoplastic damage in addition to the overexpression of Nrf2-regulated genes (COX2, proliferating cell nuclear antigen) was significantly higher in Nrf2-deficient mice compared to wild-type animals after a high dose of KBrO3 injection [129].
Jang et al. evaluated the protective effect of simvastatin, a synthetic derivative from the fermentation of Aspergillus terreus, on the expression of Nrf2 in two lines of human colon cancer cells. The study showed that simvastatin induced overexpression of antioxidant enzymes (HO-1, NQO-1, and glutamate–cysteine ligase catalytic subunit) and of Nrf2 in addition to its nuclear translocation. Additionally, using PI3K/Akt and ERK inhibitors, the authors demonstrated the ability of simvastatin in Nrf2 induction in colon cancer cells was mediated by these two pathways [130].
Considering the protective role of Nrf2, the identification of molecules able to sustain constant levels of this factor in an early stage of CRC could be a potential way to prevent cancer development and progression (Figure 3). For this reason, nowadays, many Nrf2 activators of both natural (i.e., sulforaphane, curcumin, resveratrol, kahweol, lycopene, carnosol) [131,132,133,134,135,136] and synthetic (i.e., oltipraz, dimethyl fumarate) origin are constantly tested in order to discover new effective treatments for CRC [137,138,139,140,141]. Indeed, these compounds proved to be able to protect cells from carcinogenic insults by the activation of the phase II detoxification enzymes and, consequently, to increase the Nrf2 cytoprotective response.
The most well-investigated natural molecule was sulforaphane, that if consumed in high doses, reduced the risk of cancer development in many organs, including colon [93,94,134,135,136]. The phenolic compounds epigallocatechin-3-gallate (EGCG), the main active catechin present in green tea, seems to exert antioxidant, anti-inflammatory, and chemopreventive functions in cancer. In in vitro experimental studies using colon cancer cells (Caco-2), EGCG was able to induce Nrf2 through ERK1/2 activation and Akt phosphorylation. The data suggest that due to its anti-inflammatory properties and its ability to interfere with Nrf2 signaling, EGCG could be an effective drug to prevent colorectal cancer development [131]. Trivedi et al. demonstrated that melatonin (MEL) exerted beneficial effects in colitis-associated colon carcinogenesis (CACC). The study showed that MEL was able to decrease autophagy, increase p62 levels, and activate Nrf2, leading to an enhancement of expression of various antioxidant enzymes. The results highlighted that MEL, through its ability to modulate Nrf2 signaling, attenuated the progression of CACC in mice [132]. In a study conducted by Zuo, luteolin (LUT), a dietary flavone, suppressed colorectal carcinogenesis, acting through Nrf2 epigenetic modifications [133]. The same molecule was tested by Kang et al., which revealed that LUT was able to increase the mRNA expression of Nrf2 by DNA demethylase and by the interaction between Nrf2 and p53, exerting a pro-apoptotic effect [142]. The effect of Shaoyao decoction (SYD), another natural compound, was tested by Wang et al. in AOM/DSS-induced CRC in mice. The authors demonstrated that SYD induced a reduction of oxidative stress and inflammation, activating Keap1–Nrf2–ARE signaling. These findings suggest that SYD plays an antioxidant effect and consequently prevents colitis-associated CRC [143]. In a recent paper, Sageretia thea has been reported as a potential drug of colorectal cancer. In fact, Kim et al. demonstrated that S. thea decreased the viability of cancer cells and increased HO-1 expression, activating Nrf2 [144].
Among the synthetic molecules, oltipraz is one of the most studied. In many organs, it was shown to be an effective inhibitor of chemically induced carcinogenesis processes [96,137,138,139,140,145,146,147,148,149]. However, in the bowel, its role is controversial, since chronic doses of this drug seems to increase colorectal cancer risk [142].
6.2. Nrf2 Plays an “Offensive Effect” in Late-Stage Colorectal Cancer
C–X–C chemokine receptor type 4 (CXCR4) and Nrf2 signaling (both normally activated during the cellular response in cancer) appears to be associated with clinical features in patients affected by CRC. High levels of CXCR4 and Nrf2 were correlated with increased tumor recurrence and lymph node and distance metastasis, suggesting that anomalous levels of these two molecules are involved in CRC progression [150].
Multidrug resistance (MDR) represents one of the main problems in CRC treatment and the mechanisms underlying this process are still unclear. In an in vitro study, Zhao et al. analyzed epigenetic modification of the Nrf2 gene to clarify the drug resistance mechanisms observed in colon cancer cells treated with 5-fluorouracil (5-FU). High levels of Nrf2 and its nuclear translocation accompanied the increased HO-1 and ROS levels found in resistant CRC cells (SNU-C5R) compared to controls (SNU-C5). These results suggest that overexpression of Nrf2 and an increase of HO-1 activity are involved in drug resistance occurring in colon cancer [151]. Zhang et al. demonstrated that curcumin was involved in MDR in colorectal cancer cell lines through a direct link with Nrf2. In fact, this compound, in combination with 5-FU, was able to induce apoptosis and reduce Nrf2 and Bcl-2/Bax expression, leading to a reversal effect on MDR in colon cancer [152]. Recently, Zhang et al. revealed, for the first time, the involvement of DUB3—an alternative Nrf2 deubiquitinating enzyme—in chemoresistance occurring in colorectal cancer. The authors demonstrated that DUB3 could be a new therapeutic target for the development of new drugs able to stabilize levels of Nrf2 and counteract its chemotherapy resistance effect [153].
Other studies have focused on the possible correlation between Nrf2 subcellular localization and the development and progression of colorectal cancer. In in vivo experiments, Lin et al. evaluated the expression of antioxidant enzymes (i.e., NQO-1 and HO-1) on colorectal tumor fragments showing that Nrf2 was exclusively expressed in the nucleus (nNrf2) in NQO-1- and HO-1-positive tumors, while it was retained in the cytoplasm (cNrf2) in NQO-1- and HO-1-negative tissues. Additionally, patients with cNrf2 tumors had a poorer outcome compared to those with nNrf2. In in vitro experiments, they demonstrated that cNrf2 increased tumor invasiveness through the upregulation of proteasome non-ATPase regulatory subunit 4 (PSMD4), a crucial enzyme in proteasome complex assembly. These data revealed Nrf2/PSMD4 involvement in colorectal cancer progression [154]. Further confirmation that cNrf2 acts as pro-oncogenic factor came from a study made by Cheng et al. They showed that cNrf2 conferred chemoresistance to 5-FU and oxaliplatin both in vitro (HCT116 cells) and in vivo (animal model of CRC). The resistance mechanism involved activation of PSMD4 which, in turn, enhanced Nrf2 export from the nucleus and, finally, the activation of NF-κB/AKT/β-catenin/ZEB1 cascades. These experimental observations were confirmed in colorectal cancer patients, in whom a higher prevalence of unfavorable chemotherapeutic response was noted in subjects with cNrf2- and PSMD4-positive tumors [155].
There is evidence that the antitumor effects of some compounds are related to their inhibition of Nrf2, while stimulation of tumor growth induced by certain substances is mediated by the activation of Nrf2 [156,157,158]. In a recent study, Thajmohammadi and colleagues demonstrated that stattic (a STAT3 inhibitor) represented an effective molecule to increase the cytotoxic effect of 5-FU in colon cancer cells. The study showed that the simultaneous administration of stattic and 5-FU increased the antitumor effect, decreasing Nrf2 and Bcl-2 levels and, at the same time, increasing the level of Bax in tumor cells. These findings revealed that stattic could be an effective treatment in colon cancer due to its pro-apoptotic function by modulating the Nrf2 expression [156]. Cernigliaro et al., in an in vitro study, reported the effect of ethanol (EtOH) administration on the survival of colon cancer cells. The authors revealed that EtOH increased the expression of antioxidant enzymes and induced Nrf2 nuclear translocation. These data supported the hypothesis that EtOH promotes progression and aggressiveness of colon cancer cells through the activation of Nrf2/HO-1 signaling [157]. In another study, it was reported that glyceollins (soybean-derived phytoalexins) are involved in the development of colon cancer. Using a xenograft BALB/c nude mouse model, the researchers showed that glyceollin administration induced tumor growth by activating Nrf2 signaling and inducing overexpression of HO-1 [158]. Taira et al. demonstrated the ability of marine peroxy sesquiterpenoids, an extract of Sinularia sp. coral, to exert an antitumor effect in human colon cancer cells, suppressing Nrf2-ARE signaling and inducing apoptosis in cancer cells [159]. This natural extract was able to inhibit antiapoptotic molecules (B-cell lymphoma-extra large (Bcl-xL) and phospho-Akt (p-Akt)) and to reduce HO-1, Nrf2, and its phosphorylated form in a colon cancer cell line (HCT116) [159].
In 2018, Sadeghi and colleagues tested the relationship between clinicopathological features of CRC and the expression of Nrf2, Keap1, and ATP binding cassette subfamily B member 1 (ABCB1) [160]. The study highlighted that Nrf2 and ABCB1 genes were overexpressed in CRC fragments compared to controls, showing Nrf2 involvement in CRC pathogenesis. On the contrary, the Nrf2 inhibitor Keap1 was significantly higher in controls with respect to the CRC group. These findings suggest that Nrf2 and P-gp (the main product of the ABCB1 gene) are implicated in CRC development and progression in addition to chemoresistance occurring in the tumor. Inhibitors of the Nrf2-ABCB1/P-gp axis could be useful in increasing the effectiveness of CRC chemotherapeutic drugs.
As Nrf2 may play a central role in tumor progression and in chemo- and radioresistance, Nrf2 inhibitors have emerged as a promising adjuvant therapy to improve chemotherapeutic drug effectiveness (Figure 3). Several Nrf2 inhibitors, including plant extracts (as flavonoids and alkaloids) or synthetic molecules (as ARE expression modulator 1 and metformin), have also been tested in CRC [18,97,161,162]. Particularly, Yao and colleagues revealed, both in vivo and vitro experiments, that wogonin, a flavonoid-like chemical compound, suppress inflammation-associated colon carcinogenesis and cancer development. Wogonin inhibited cell proliferation and production of pro-inflammatory mediators and regulates expression of NF-jB and Nrf2 [163]. Recently, Evans et al. reported that primary CRC and metastatic samples showed high levels of Nrf2 compared to control group. Moreover, in the same study, brusatol (Nrf2 natural inhibitor) injection counteracted tumor growth, suggesting that Nrf2 could be a potential target in CRC treatment [164].
Among synthetic molecules, it was demonstrated that metformin reduced mRNA levels of Nrf2 and patients affected by CRC treated with metformin showed a better prognosis [18,165,166]. Henderson et al. reported that metformin administration improved the prognosis of patients with CRC by reducing tumor recurrences, metastasis, and deaths [165]. Sena et al. treated a colon cancer cell line (HT29) with different concentration of metformin, showing an increase in apoptosis and autophagy processes in treated compared to untreated cells. The study highlighted that the anticancer effects of metformin are correlated with Nrf2 inhibition [166]. All these data suggest that metformin could be an efficacious candidate for the treatment of CRC.
7. Conclusions
Inflammatory bowel diseases, including Crohn’s disease and ulcerative colitis, represent a challenging clinical condition since they can lead to severe complications such as intestinal fibrosis and colorectal cancer which are currently manageable only through surgery.
Limiting IBD progression and the development of chronic complications like fibrosis and cancer represents an important milestone. Identifying the main molecules responsible for the development of IBD-associated intestinal fibrosis and cancer is crucial for the prevention of such severe complications. Nrf2, a transcriptional factor regulating many antioxidant genes, seems to be a good candidate, showing a defensive role against inflammation, fibrosis, and cancer in various organs, including the intestine. Several studies have shown that Nrf2 is directly and strongly involved in the intestinal fibrosis and colorectal cancer occurring in IBD. It emerges that the use of Nrf2 activators to maintain constant levels of this molecule in the first stage of many inflammatory diseases can limit their progression and complications such as fibrosis and cancer. On the other hand, as Nrf2 is overexpressed in advanced cancer, Nrf2 inhibitors may represent an effective therapeutic adjuvant leading to a significant reduction of radio- and chemoresistance. Nevertheless, future studies are needed to clarify the complex role of Nrf2 and the possibility to develop new drugs able to modulate the Nrf2 pathway and to potentiate its protective effect. This scenario could be a valid chance to limit the two main IBD chronic complications—intestinal fibrosis and cancer.
Author Contributions
S.P. performed the literature review, wrote the manuscript and prepared the illustrations; R.S. and A.V. (Antonella Vetuschi) wrote and reviewed the manuscript and suggested important research points; E.G., A.V. (Angelo Viscido), and G.F. reviewed the manuscript and provided critical comments; G.L. suggested the topic of review and supervised, wrote and critically reviewed the manuscript. All authors read and approved the final manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wynn, T.A. Cellular and molecular mechanisms of fibrosis. J. Pathol. 2008, 214, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Speca, S.; Giusti, I.; Rieder, F.; Latella, G. Cellular and molecular mechanisms of intestinal fibrosis. World J. Gastroenterol. 2012, 18, 3635–3661. [Google Scholar] [CrossRef] [PubMed]
- Vetuschi, A.; Latella, G.; Pompili, S.; Gaudio, E.; Sferra, R. Features of intestinal lesions in the clinical course of inflammatory bowel diseases. Ital. J. Anat. Embryol. 2014, 119, 286–303. [Google Scholar] [PubMed]
- Latella, G.; Di Gregorio, J.; Flati, V.; Rieder, F.; Lawrance, I.C. Mechanisms of initiation and progression of intestinal fibrosis in IBD. Scand. J. Gastroenterol. 2015, 50, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Sferra, R.; Pompili, S.; Festuccia, C.; Marampon, F.; Gravina, G.L.; Ventura, L.; Di Cesare, E.; Cicchinelli, S.; Gaudio, E.; Vetuschi, A. The possible prognostic role of histone deacetylase and transforming growth factor β/Smad signaling in high grade gliomas treated by radio-chemotherapy: A preliminary immunohistochemical study. Eur. J. Histochem. 2017, 61, 2732. [Google Scholar] [CrossRef] [PubMed]
- Vetuschi, A.; Pompili, S.; Gaudio, E.; Latella, G.; Sferra, R. PPAR-γ with its anti-inflammatory and anti-fibrotic action could be an effective therapeutic target in IBD. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 8839–8848. [Google Scholar]
- Samarakoon, R.; Overstreet, J.M.; Higgins, P.J. TGF-βsignaling in tissue fibrosis: Redox controls, target genes and therapeutic opportunities. Cell Signal. 2013, 25, 264–268. [Google Scholar] [CrossRef]
- Liu, R.M.; Desai, L.P. Reciprocal regulation of TGF-βand reactive oxygen species: A perverse cycle for fibrosis. Redox Biol. 2015, 6, 565–577. [Google Scholar] [CrossRef]
- Richter, K.; Kietzmann, T. Reactive oxygen species and fibrosis: Further evidence of a significant liaison. Cell Tissue Res. 2016, 365, 591–605. [Google Scholar] [CrossRef]
- Swamy, S.M.; Rajasekaran, N.S.; Thannickal, V.J. Nuclear factor erythroid-2 related factor 2 in aging and lung fibrosis. Am. J. Pathol. 2016, 186, 1712–1723. [Google Scholar] [CrossRef]
- Ahamed, J.; Laurence, J. Role of platelet-derived transforming growth factor-β1 and reactive oxygen species in radiation-induced organ fibrosis. Antioxid. Redox Signal. 2017, 27, 977–988. [Google Scholar] [CrossRef] [PubMed]
- Khor, T.O.; Huang, M.T.; Prawan, A.; Liu, Y.; Hao, X.; Yu, S.; Cheung, W.K.; Chan, J.Y.; Reddy, B.S.; Yang, C.S.; et al. Increased susceptibility of Nrf2 knockout mice to colitis-associated colorectal cancer. Cancer Prev. Res. 2008, 1, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Misra, V.; Thimmulappa, R.K.; Lee, H.; Ames, S.; Hoque, M.O.; Herman, J.G.; Baylin, S.B.; Sidransky, D.; Gabrielson, E.; et al. Dysfunctional KEAP1-NRF2 interaction in non-small-cell lung cancer. PLoS Med. 2006, 3, e420. [Google Scholar] [CrossRef]
- Shibata, T.; Kokubu, A.; Gotoh, M.; Ojima, H.; Ohta, T.; Yamamoto, M.; Hirohashi, S. Genetic alteration of Keap1 confers constitutive Nrf2 activation and resistance to chemotherapy in gallbladder cancer. Gastroenterology 2008, 135, 1358–1368. [Google Scholar] [CrossRef]
- Wang, R.; An, J.; Ji, F.; Jiao, H.; Sun, H.; Zhou, D. Hypermethylation of the Keap1 gene in human lung cancer cell lines and lung cancer tissues. Biochem. Biophys. Res. Commun. 2008, 373, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Solis, L.M.; Behrens, C.; Dong, W.; Suraokar, M.; Ozburn, N.C.; Moran, C.A.; Corvalan, A.H.; Biswal, S.; Swisher, S.G.; Bekele, B.N.; et al. Nrf2 and Keap1 abnormalities in non-small cell lung carcinoma and association with clinicopathologic features. Clin. Cancer Res. 2010, 16, 3743–3753. [Google Scholar] [CrossRef]
- Zhang, P.; Singh, A.; Yegnasubramanian, S.; Esopi, D.; Kombairaju, P.; Bodas, M.; Wu, H.; Bova, S.G.; Biswal, S. Loss of Kelch-like ECH-associated protein 1 function in prostate cancer cells causes chemoresistance and radioresistance and promotes tumor growth. Mol. Cancer Ther. 2010, 9, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Jung, B.J.; Yoo, H.S.; Shin, S.; Park, Y.J.; Jeon, S.M. Dysregulation of NRF2 in Cancer: From Molecular Mechanisms to Therapeutic Opportunities. Biomol. Ther. 2018, 26, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Latella, G. Redox Imbalance in Intestinal Fibrosis: Beware of the TGF-β1, ROS, and Nrf2 Connection. Dig. Dis. Sci. 2018, 63, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Latella, G. Concise Commentary: Is Nrf2 a Master Regulator of Intestinal Fibrosis? Dig. Dis. Sci. 2018, 63, 381–382. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Tan, Y.; Liu, W.; Wang, D.; Pan, D.; Sun, Y.; Zheng, C. NF-E2-Related Factor 2 Suppresses Intestinal Fibrosis by Inhibiting Reactive Oxygen Species-Dependent TGF-β1/SMADs Pathway. Dig. Dis. Sci. 2018, 63, 366–380. [Google Scholar] [CrossRef] [PubMed]
- Osburn, W.O.; Kensler, T.W. Nrf2 signaling: An adaptive response pathway for protection against environmental toxic insults. Rev. Mutat. Res. 2008, 659, 31–39. [Google Scholar] [CrossRef]
- Cuadrado, A.; Manda, G.; Hassan, A.; Alcaraz, M.J.; Barbas, C.; Daiber, A.; Ghezzi, P.; Leόn, R.; Lόpez, M.G.; Oliva, B.; et al. Transcription Factor NRF2 as a Therapeutic Target for Chronic Diseases: A Systems Medicine Approach. Pharmacol. Rev. 2018, 70, 348–383. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, C.; Chio, C.I.I.; Tuveson, D.A. Transcriptional Regulation by Nrf2. Antioxid. Redox Signal. 2018, 29, 1727–1745. [Google Scholar] [CrossRef]
- Cullinan, S.; Gordan, J.; Jin, J.; Harper, J.; Diehl, J. The keap1-BTB protein is an adaptor that bridges Nrf2 to a Cul3-based E3 ligase: Oxidative stress sensing by Cul3-Keap1 ligase. Mol. Cell. Biol. 2004, 24, 8477–8486. [Google Scholar] [CrossRef]
- Ahmed, S.M.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Bas. Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Moi, P.; Chan, K.; Asunis, I.; Cao, A.; Kan, Y.W. Isolation of NF-E2-related factor 2 (Nrf2), a NF-E2-like basic leucine zipper transcriptional activator that binds to the tandem NF-E2/AP1 repeat of the beta-globin locus control region. Proc. Natl. Acad. Sci. USA 1994, 91, 9926–9930. [Google Scholar] [CrossRef]
- Itho, K.; Chiba, T.; Takahashi, S.; Ishii, T.; Igarashi, K.; Katoh, Y.; Oyake, T.; Hayashi, N.; Satoh, K.; Hatayama, I.; et al. An Nrf2/small Maf heterodimer mediates the induction of phase II detoxifying enzyme genes through antioxidant response elements. Biochem. Biophys. Res. Commun. 1997, 236, 313–322. [Google Scholar] [CrossRef]
- Telakowski-Hopkins, C.A.; King, R.G.; Pickett, C.B. Glutathione S-transferase Ya subunit gene: Identification of regulatory elements required for basal level and inducible expression. Proc. Natl. Acad. Sci. USA 1988, 85, 1000–1004. [Google Scholar] [CrossRef] [PubMed]
- Rushmore, T.H.; Morton, M.R.; Pickett, C.B. The antioxidant responsive element. Activation by oxidative stress and identification of the DNA consensus sequence required for functional activity. J. Biol. Chem. 1991, 266, 11632–11639. [Google Scholar]
- Friling, R.S.; Bergelson, S.; Daniel, V. Two adjacent AP1-like binding sites from the electrophile-responsive element of the murine glutathione S-transferase Ya subunit gene. Proc. Natl. Acad. Sci. USA 1992, 89, 668–672. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; Dinkova-Kostova, A.T. The Nrf2 regulatory network provides an interface between redox and intermediary metabolism. Trends Biochem. Sci. 2014, 39, 199–218. [Google Scholar] [CrossRef]
- Hirayama, A.; Yoh, K.; Nagase, S.; Ueda, A.; Itoh, K.; Morito, N.; Hirayama, K.; Takahashi, S.; Yamamoto, M.; Koyama, A. EPR imaging of reducing activity in Nrf2 transcriptional factor-deficient mice. Free Radic. Biol. Med. 2003, 34, 1236–1242. [Google Scholar] [CrossRef]
- Morito, N.; Yoh, K.; Itoh, K.; Hirayama, A.; Koyama, A.; Yamamoto, M.; Takahashi, S. Nrf2 regulates the sensitivity of death receptor signals by affecting intracellular glutathione levels. Oncogene 2003, 22, 9275–9281. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.Y.; Jedlicka, A.E.; Reddy, S.P.; Kensler, T.W.; Yamamoto, M.; Zhang, L.Y.; Kleeberger, S.R. Role of NRF2 in protection against hyperoxic lung injury in mice. Am. J. Respir. Cell Mol. Biol. 2002, 26, 175–182. [Google Scholar] [CrossRef]
- Ma, Q. Role of nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [PubMed]
- Hybertson, B.M.; Gao, B.; Bose, S.K.; McCord, J.M. Oxidative stress in health and disease: The therapeutic potential of Nrf2 activation. Mol. Asp. Med. 2011, 32, 234–246. [Google Scholar] [CrossRef] [PubMed]
- Gorrini, C.; Harris, I.S.; Mark, T.W. Modulation of oxidative stress as an anticancer strategy. Nat. Rev. Drug Discov. 2013, 12, 931–947. [Google Scholar] [CrossRef]
- Tomashefski, J.F. Pulmonary pathology of acute respiratory distress syndrome. Clin. Chest Med. 2000, 21, 435–466. [Google Scholar] [CrossRef]
- Ware, L.B.; Matthay, M.A. The acute respiratory distress syndrome. N. Engl. J. Med. 2000, 342, 1334–1349. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, K.; Handa, H.; Nishizawa, M. Induction of cellular anti-oxidative stress genes through heterodimeric transcription factor Nrf2/ small Maf by antirheumatic gold(I) compounds. J. Biol. Chem. 2001, 276, 34074–34081. [Google Scholar] [CrossRef] [PubMed]
- Abraham, E. Neutrophils and acute lung injury. Crit. Care Med. 2003, 31, S195–S199. [Google Scholar] [CrossRef] [PubMed]
- Arisawa, T.; Tahara, T.; Shibata, T.; Nagasaka, M.; Nakamura, M.; Kamiya, Y.; Fujita, H.; Hasegawa, S.; Takagi, T.; Wang, F.Y.; et al. The relationship between Helicobacter pylori infection and promoter polymorphism of the Nrf2 gene in chronic gastritis. Int. J. Mol. Med. 2007, 19, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Levonen, A.L.; Inkala, M.; Heikura, S.; Jauhiainen, S.; Jykkanen, H.K.; Kansanen, E.; Maatta, K.; Romppanen, E.; Turenen, P.; Rutanen, J.; et al. Nrf2 gene transfer induces antioxidant enzymes and suppresses smooth muscle cell growth in vitro and reduces oxidative stress in rabbit aorta in vivo. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 741–747. [Google Scholar] [CrossRef] [PubMed]
- Nathan, C. Points of control in inflammation. Nature 2002, 420, 846–852. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Mochizuki, M.; Ishii, Y.; Ishii, T.; Shibata, T.; Kawamoto, Y.; Kelly, W.; Sekizawa, K.; Uchida, K.; Yamamoto, M. Transcription Factor Nrf2 Regulates Inflammation by Mediating the Effect of 15-Deoxy-∆12,14-Prostaglandin J2. Mol. Cell. Biol. 2004, 24, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Chan, K.; Kan, Y.W.; Johnson, J.A. Targeted disruption of Nrf2 causes regenerative immune-mediated hemolytic anemia. Proc. Natl. Acad. Sci. USA 2004, 101, 9751–9756. [Google Scholar] [CrossRef] [PubMed]
- Banning, A.; Brigelius-Flohe, R. NF-kappaB, Nrf2 and HO-1 interplay in redox regulated VCAM -1 expression. Antioxid. Redox Signal. 2005, 7, 889–899. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Sohn, D.H.; Jin, X.Y.; Kim, S.W.; Choi, S.C.; Seo, G.S. 2′,4′,6′Tris(methoxymethoxy) chalcone protects against trinitrobenzene sulfonic acid-induced colitis and blocks tumor necrosis factor-alpha-induced intestinal epithelial inflammation via heme oxygenase 1-dependent and independent pathways. Biochem. Pharmacol. 2007, 74, 870–880. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Young-Nam, C.; Young-Joon, S. A protective role of nuclear factor-erythroid 2-related factor-2 (Nrf2) in inflammatory disorders. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2009, 690, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Hellerbrand, C.; Köhler, A.U.; Bugnon, P.; Kan, Y.W.; Werner, S.; Beyer, T.A. The Nrf2 transcription factor protects from toxin-induced liver injury and fibrosis. Lab. Investig. 2008, 88, 1068–1078. [Google Scholar] [CrossRef] [PubMed]
- Sriram, N.; Kalayarasan, S.; Sudhandiran, G. Enhancement of antioxidant defense system by epigallocatechin-3-gallateduring bleomycin-induced experimental pulmonary fibrosis. Biol. Pharm. Bull. 2008, 31, 1306–1311. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, N.; Ishii, Y.; Morishima, Y.; Yageta, Y.; Haraguchi, N.; Itoh, K.; Yamamoto, M.; Hizawa, N. Nrf2 protects against pulmonary fibrosis by regulating the lung oxidant level and Th1/Th2 balance. Respir. Res. 2010, 11, 31. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zou, L.; Li, L.; Wu, T. The Protective Effect of Glycyrrhetinic Acid on Carbon Tetrachloride-Induced Chronic Liver Fibrosis in Mice via Upregulation of Nrf2. PLoS ONE 2013, 8, e53662. [Google Scholar] [CrossRef] [PubMed]
- Prestigiacomo, V.; Suter-Dick, L. Nrf2 protects stellate cells from Smad-dependent cell activation. PLoS ONE 2018, 13, e0201044. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Kang, Y.; Zhou, C.; Cui, R.; Jia, M.; Hu, S.; Ji, X.; Yuan, J.; Cui, H.; Shi, G. Amelioratory Effects of Testosterone Propionate on Age-related Renal Fibrosis via Suppression of TGF-β1/Smad Signaling and Activation of Nrf2ARE Signaling. Sci. Rep. 2018, 8, 10726. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Rong, Y.; Sun, D.; Li, W.; Chen, H.; Cao, B.; Wang, T. Costunolide inhibits pulmonary fibrosis via regulating NF-kB and TGF-β1/Smad2/Nrf2-NOX4 signaling pathways. Biochem. Biophys. Res. Commun. 2019, 510, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.R.; Fan, X.H.; Chen, G.; Zeng, G.W.; Xue, Y.G.; Liu, X.T.; Wang, C.Y. Irisin attenuates angiotensin II-induced cardiac fibrosis via Nrf2 mediated inhibition of ROS/ TGFβ1/Smad2/3 signaling axis. Chem. Biol. Interact. 2019, 302, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.S.; Kalluri, R. Mechanistic connection between inflammation and fibrosis. Kidney Int. 2010, 119, S22–S26. [Google Scholar] [CrossRef] [PubMed]
- Latella, G.; Vetuschi, A.; Sferra, R.; Speca, S.; Gaudio, E. Localization of ανβ6 integrin-TGF-β1/Smad3, mTOR and PPARγ in experimental colorectal fibrosis. Eur. J. Histochem. 2013, 57, e40. [Google Scholar] [CrossRef] [PubMed]
- Sferra, R.; Pompili, S.; Ventura, L.; Dubuquoy, C.; Speca, S.; Gaudio, E.; Latella, G.; Vetuschi, A. Interaction between sphingosine kinase/sphingosine 1 phosphate and transforming growth factor-β/Smads pathways in experimental intestinal fibrosis. An in vivo immunohistochemical study. Eur. J. Histochem. 2018, 31, 62. [Google Scholar] [CrossRef] [PubMed]
- Khor, T.O.O.; Huang, M.T.; Kwon, H.K.; Chan, J.Y.; Reddy, B.S.; Kong, A.N. Nrf2-Deficient Mice Have an Increased Susceptibility to Dextran Sulfate Sodium-Induced Colitis. Cancer Res. 2006, 66, 11580–11584. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; An, J.M.; Kang, E.A.; Han, Y.M.; Kim, Y.S.; Lee, H.J.; Kim, K.J.; Surh, Y.J.; Hahm, K.B. Host nuclear factor erythroid 2-related factor-2 defense system determines the outcome of dextran sulfate sodium-induced colitis in mice. J. Physiol Pharmacol. 2018, 69, 755–767. [Google Scholar] [CrossRef]
- Li, J.; Wang, H.; Zheng, Z.; Luo, L.; Wang, P.; Liu, K.; Namani, A.; Jiang, Z.; Wang, X.J.; Tang, X. Mkp-1 cross-talks with Nrf2/Ho-1 pathway protecting against intestinal inflammation. Free Radic. Biol. Med. 2018, 124, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, H.; Qian, C.; Tang, J.; Zhou, W.; Liu, X.; You, Q.; Hu, R. 3-(2-Oxo-2-phenylethylidene)-2,3,6,7-tetrahydro-1H-pyrazino [2,1-a] isoquinolin-4 (11bH)-one (compound 1), a novel potent Nrf2/ARE inducer, protects against DSS-induced colitis via inhibiting NLRP3 inflammasome. Biochem. Pharmacol. 2016, 101, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Poss, K.D.; Tonegawa, S. Reduced stress defense in heme oxygenase 1-deficient cells. Proc. Natl. Acad. Sci. USA 1996, 20, 10925–10930. [Google Scholar] [CrossRef] [PubMed]
- Willis, D.; Moore, A.R.; Frederick, R.; Willoughby, D.A. Heme oxygenase: A novel target for the modulation of the inflammatory response. Nat. Med. 1996, 87, 87–90. [Google Scholar] [CrossRef]
- Laniado-Schwartzman, M.; Abraham, N.G.; Conners, M.; Dunn, M.W.; Levere, R.D. Heme oxygenase induction with attenuation of experimentally induced corneal inflammation. Biochem. Pharmacol. 1997, 53, 1069–1075. [Google Scholar] [CrossRef]
- Otterbein, L.E.; Bach, F.H.; Alam, J.; Soares, M.; Lu, H.T.; Wysk, M.; Davis, R.J.; Flavell, R.A.; Choi, A.M.K. Carbon monoxide has anti-inflammatory effects involving the mitogen-activated protein kinase pathway. Nat. Med. 2000, 6, 422–428. [Google Scholar] [CrossRef]
- Wang, W.P.; Guo, X.; Koo, M.V.L.; Wong, B.C.Y.; Lam, S.K.; Ye, Y.N.; Cho, C.H. Protective role of heme oxygenase-1 on trinitrobenzene sulfonic acid-induced colitis in rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 281, G586–G594. [Google Scholar] [CrossRef]
- Lu, M.C.; Ji, J.A.; Jiang, Y.L.; Chen, Y.Z.; Yuan, Z.W.; You, Q.D.; Jiang, Z.Y. An inhibitor of the Keap1-Nrf2 protein-protein interaction protects NCM460 colonic cells and alleviates experimental colitis. Sci. Rep. 2016, 6, 26585. [Google Scholar] [CrossRef] [PubMed]
- Yalniz, M.; Demirel, U.; Orhan, C.; Bahcecioglu, I.H.; Ozercan, I.H.; Aygun, C.; Tuzcu, M.; Sahin, K. Nadroparin Sodium Activates Nrf2/HO-1 Pathway in Acetic Acid-Induced Colitis in Rats. Inflammation 2012, 35, 1213–1221. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhou, W.; Zhang, X.; Lu, P.; Du, Q.; Tao, L.; Ding, Y.; Wang, Y.; Hu, R. Dimethyl fumarate ameliorates dextran sulphate sodium-induced murine experimental colitis by activating Nrf2 and suppressing NLRP3 inflammasome activation. Biochem. Pharmacol. 2016, 112, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Shen, L.; Li, Y.; Li, Y.; Yu, S.; Wang, S. Hyperoside attenuates dextran sulfate sodium-induced colitis in mice possibly via activation of the Nrf2 signalling pathway. J. Inflamm. 2017, 14, 25. [Google Scholar] [CrossRef] [PubMed]
- Park, D.D.; Yum, H.W.; Zhong, Z.; Kim, S.H.; Kim, S.H.; Kim, D.H.; Kim, S.J.; Na, H.K.; Sato, A.; Miura, T.; et al. Perilla frutescens Extracts Protects against Dextran Sulphate Sodium-Induced Murine Colitis: NF-kB, STAT3, and Nrf2 as Putative Targets. Front. Pharmacol. 2017, 8, 1–14. [Google Scholar]
- Jing, W.; Safarpour, Y.; Zhang, T.; Guo, P.; Chen, G.; Wu, X.; Fu, Q.; Wang, Y. Berberine Upregulates P-Glycoprotein in Human Caco-2 Cells and in an Experimental Model of Colitis in the Rat via Activation of Nrf2-Dependent Mechanisms. J. Pharmacol. Exp. Ther. 2018, 366, 332–340. [Google Scholar] [CrossRef]
- Li, Y.; Shen, L.; Luo, H. Luteolin ameliorates dextran sulfate sodium-induced colitis in mice possibly through activation of the Nrf2 signaling pathway. Int. Immunopharmacol. 2016, 40, 24–31. [Google Scholar] [CrossRef]
- Kim, K.J.; Park, J.M.; Lee, J.S.; Kim, Y.S.; Kangwan, N.; Han, Y.M.; Kang, E.A.; An, J.M.; Park, Y.K.; Hahm, K.B. Oligonol prevented the relapse of dextran sulfate sodium-ulcerative colitis through enhancing NRF2-mediated antioxidative defense mechanism. J. Physiol. Pharmacol. 2018, 69, 359–371. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, H.; Song, J.; Cao, L.; Tang, L.; Qi, C. Sinomenine alleviates dextran sulfate sodium-induced colitis via the Nrf2/NQO-1 signaling pathway. Mol. Med. Rep. 2018, 18, 3691–3698. [Google Scholar] [CrossRef]
- Gao, Y.; Bai, D.; Zhao, Y.; Zhu, Q.; Zhou, Y.; Li, Z.; Lu, N. LL202 ameliorates colitis against oxidative stress of macrophage by activation of the Nrf2/HO-1 pathway. J. Cell. Physiol. 2019, 234, 10625–10639. [Google Scholar] [CrossRef]
- Saber, S.; Khalil, R.M.; Abdo, W.S.; Nassif, D.; El-Ahwany, E. Olmesartan ameliorates chemically-induced ulcerative colitis in rats via modulating NFκB and Nrf-2/HO-1 signaling crosstalk. Toxicol. Appl. Pharmacol. 2019, 364, 120–132. [Google Scholar] [CrossRef] [PubMed]
- Cheon, H.J.; Nam, S.H.; Kim, J.K. Tussilagone, a major active component in Tussilago farfara, ameliorates inflammatory responses in dextran sulphate sodium-induced murine colitis. Chem. Biol. Interact. 2018, 294, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Vemu, B.; Veenstra, J.; Petiwala, S.M.; Johnson, J.J. Carnosol, a dietary diterpene from rosemary (Rosmarinus officinalis) activates Nrf2 leading to sestrin 2 induction in colon cells. Integr. Mol. Med. 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Xia, Z.; Shao, N.; Li, B.; Xue, L.; Peng, Y.; Zhi, F.; Yang, Y. Carnosic acid prevents dextran sulfate sodium-induced acute colitis associated with the regulation of the Keap1/Nrf2 pathway. Sci. Rep. 2017, 7, 11036. [Google Scholar] [CrossRef] [PubMed]
- Sangaraju, R.; Nalban, N.; Alavala, S.; Rajendran, V.; Jerald, M.K.; Sistla, R. Protective effect of galangin against dextran sulfate sodium (DSS)-induced ulcerative colitis in Balb/c mice. Inflamm. Res. 2019, 68, 1–14. [Google Scholar] [CrossRef]
- Hwang, Y.J.; Nam, S.J.; Chun, W.; Kim, S.I.; Park, S.C.; Kang, C.D.; Lee, S.J. Anti-inflammatory effects of apocynin on dextran sulfate sodium-induced mouse colitis model. PLoS ONE 2019, 14, e0217642. [Google Scholar] [CrossRef]
- Liu, G.; Yu, L.; Fang, J.; Hu, C.A.; Yin, J.; Ni, H.; Ren, W.; Duraipandiyan, V.; Chen, S.; Al-Dhabi, N.A.; et al. Methionine restriction on oxidative stress and immune response in DSS-induced colitis mice. Oncotarget 2017, 8, 44511–44520. [Google Scholar] [CrossRef]
- Fan, F.Y.; Sang, L.X.; Jiang, M. Catechins and Their Therapeutic Benefits to Inflammatory Bowel Disease. Molecules 2017, 22, 484. [Google Scholar] [CrossRef]
- Liu, D.; Huo, X.; Gao, L.; Zhang, J.; Ni, H.; Cao, L. NF-κB and Nrf2 pathways contribute to the protective effect of Licochalcone A on dextran sulfate sodium-induced ulcerative colitis in mice. Biomed. Pharmacother. 2018, 102, 922–929. [Google Scholar] [CrossRef]
- Li, J.; Chen, H.; Wang, B.; Cai, C.; Yang, X.; Chai, Z.; Feng, W. ZnO nanoparticles act as supportive therapy in DSS-induced ulcerative colitis in mice by maintaining gut homeostasis and activating Nrf2 signaling. Sci. Rep. 2017, 7, 43126. [Google Scholar] [CrossRef]
- Ramos-Gomez, M.; Kwak, M.K.; Dolan, P.M.; Itho, K.; Yamamoto, M.; Talalay, P.; Kensler, T.W. Interactive effects of nrf2 genotype and oltipraz on benzo[a]pyrene-DNA adducts and tumor yield in mice. Carcinogenesis 2003, 24, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Osburn, W.O.; Karim, B.; Dolan, P.M.; Liu, G.; Yamamoto, M.; Huso, D.L.; Kensler, T.W. Increased colonic inflammatory injury and formation of aberrant crypt foci in Nrf2-deficient mice upon dextran sulfate treatment. Int. J. Cancer 2007, 121, 1883–1891. [Google Scholar] [CrossRef] [PubMed]
- Fahey, J.W.; Haristoy, X.; Dolan, P.M.; Kensler, T.W.; Scholtus, I.; Stephenson, K.K.; Talalay, P.; Lozniewski, A. Sulforaphane inhibits extracellular, intracellular, and antibiotic-resistant strains of Helicobacter pylori and prevents benzo[a]pyrene-induced stomach tumors. Proc. Natl. Acad. Sci. USA 2002, 99, 7610–7615. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Huang, M.T.; Shen, G.; Yuan, X.; Lin, W.; Khor, T.O.; Conney, A.H.; Kong, A.N. Inhibition of 7,12-dimethylbenz(a)anthracene-induced skin tumorigenesis in C57BL/6 mice by sulforaphane is mediated by nuclear factor E2-related factor 2. Cancer Res. 2006, 66, 8293–8296. [Google Scholar] [CrossRef] [PubMed]
- Iida, K.; Itoh, K.; Kumagai, Y.; Oyasu, R.; Hattori, K.; Kawai, K.; Shimazui, T.; Akaza, H.; Yamamoto, M. Nrf2 is essential for the chemopreventive efficacy of oltipraz against urinary bladder carcinogenesis. Cancer Res. 2004, 64, 6424–6431. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.R.; Oh, J.E.; Kim, M.S.; Kang, M.R.; Park, S.W.; Han, J.Y.; Eom, H.S.; Yoo, N.J.; Lee, S.H. Oncogenic NRF2 mutations in squamous cell carcinomas of oesophagus and skin. J. Pathol. 2010, 220, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Venkannagari, S.; Oh, K.H.; Zhang, Y.Q.; Rohde, J.M.; Liu, M.J.; Wang, A.; Xu, X.; Shahane, S.A.; Xia, M.; et al. Small molecule inhibitor of NRF2 selectively intervenes therapeutic resistance in KEAP1-deficient NSCLC tumors. ACS Chem. Biol. 2016, 11, 3214–3225. [Google Scholar] [CrossRef]
- Mitsuishi, Y.; Taguchi, K.; Kawatani, Y.; Shibata, T.; Nukiwa, T.; Aburatani, H.; Yamamoto, M.; Motohashi, H. Nrf2 redirects glucose and glutamine into anabolic pathways in metabolic reprogramming. Cancer Cell 2012, 22, 66–79. [Google Scholar] [CrossRef]
- Satoh, H.; Moriguchi, T.; Takai, J.; Ebina, M.; Yamamoto, M. Nrf2 prevents initiation but accelerates progression through the Kras signaling pathway during lung carcinogenesis. Cancer Res. 2013, 73, 4158–4168. [Google Scholar] [CrossRef]
- Satoh, H.; Moriguchi, T.; Saigusa, D.; Baird, L.; Yu, L.; Rokutan, H.; Igarashi, K.; Ebina, M.; Shibata, T.; Yamamoto, M. NRF2 Intensifies Host Defense Systems to Prevent Lung Carcinogenesis, but After Tumor Initiation Accelerates Malignant Cell Growth. Cancer Res. 2016, 76, 3088–3096. [Google Scholar] [CrossRef]
- Shim, G.S.; Manandhar, S.; Shin, D.H.; Kim, T.H.; Kwak, M.K. Acquisition of doxorubicin resistance in ovarian carcinoma cells accompanies activation of the NRF2 pathway. Free Radic. Biol. Med. 2009, 47, 1619–1631. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo, M.C.; Zhang, D.D. The emerging role of the Nrf2–Keap1 signaling pathway in cancer. Genes Dev. 2013, 27, 2179–2191. [Google Scholar] [CrossRef] [PubMed]
- Ballinger, A.B.; Anggiansah, C. Colorectal cancer. BMJ 2007, 335, 715–718. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.; Naishadham, D.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2013, 63, 11–30. [Google Scholar] [CrossRef] [PubMed]
- Arnold, M.; Sierra, M.S.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Brai, F. Global patterns and trends in colorectal cancer incidence and mortality. Gut 2016, 66, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef]
- Rubenstein, J.H.; Enns, R.; Heidelbaugh, J.; Barkun, A. Clinical Guidelines Committee. American Gastroenterological Association Institute Guideline on the Diagnosis and Management of Lynch Syndrome. Gastroenterology 2015, 149, 777–782. [Google Scholar] [CrossRef]
- Cannon, J. Colorectal neoplasia and inflammatory Bowel disease. Surg. Clin. 2015, 95, 1261–1269. [Google Scholar] [CrossRef]
- Sasso, A.; Latella, G. Role of Heme Iron in the Association between Red Meat Consumption and Colorectal Cancer. Nutr. Cancer 2018, 70, 1173–1183. [Google Scholar] [CrossRef]
- Sasso, A.; Latella, G. Dietary components that counteract the increased risk of colorectal cancer related to red meat consumption. Int. J. Food Sci. Nutr. 2018, 69, 536–548. [Google Scholar] [CrossRef]
- Triantafillidis, J.K.; Nasioulas, G.; Kosmidis, P.A. Colorectal cancer and inflammatory bowel disease: Epidemiology, risk factors, mechanisms of carcinogenesis and prevention strategies. Anticancer Res. 2009, 29, 2727–2737. [Google Scholar] [PubMed]
- Ullman, T.A.; Itzkowitz, S.H. Intestinal inflammation and cancer. Gastroenterology 2011, 140, 1807–1816. [Google Scholar] [CrossRef] [PubMed]
- Azer, S.A. Overview of molecular pathways in inflammatory bowel disease associated with colorectal cancer development. Eur. J. Gastroenterol. Hepatol. 2013, 25, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Ekbom, A.; Helmick, C.; Zack, M.; Adami, H.O. Ulcerative colitis and colorectal cancer. A population-based study. N. Engl. J. Med. 1990, 323, 1228–1233. [Google Scholar] [CrossRef] [PubMed]
- Choi, P.M.; Zelig, M.P. Similarity of colorectal cancer in Crohn’s disease and ulcerative colitis: Implications for carcinogenesis and prevention. Gut 1994, 35, 950–954. [Google Scholar] [CrossRef] [PubMed]
- Eaden, J.A.; Abrams, K.R.; Mayberry, J.F. The risk of colorectal cancer in ulcerative colitis: A meta-analysis. Gut 2001, 48, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.; Rubin, P.H.; Bodian, C.; Harpaz, N.; Present, D.H. Screening and surveillance colonoscopy in chronic Crohn’s colitis: Results of a surveillance program spanning 25 years. Gastroenterology 2001, 120, 820–826. [Google Scholar] [CrossRef]
- Itzkowitz, S.H.; Yio, X. Inflammation and cancer IV. Colorectal cancer in inflammatory bowel disease: The role of inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, G7–G17. [Google Scholar] [CrossRef]
- Jess, T.; Loftus, E.V.; Velayos, F.S.; Harmsen, W.S.; Zinsmeister, A.R.; Smyrk, T.C.; Schleck, C.D.; Tremaine, W.J.; Melton, L.J.; Munkholm, P.; et al. Risk of intestinal cancer in inflammatory bowel disease: A population-based study from Olmsted County, Minnesota. Gastroenterology 2006, 130, 1039–1046. [Google Scholar] [CrossRef]
- Rachmilewitz, D.; Stamler, J.S.; Bachwich, D.; Karmeli, F.; Ackerman, Z.; Podolsky, D.K. Enhanced colonic nitric oxide generation and nitric oxide synthase activity in ulcerative colitis and Crohn’s disease. Gut 1995, 36, 718–723. [Google Scholar] [CrossRef]
- Kimura, H.; Hokari, R.; Miura, S.; Shigematsu, T.; Hirokawa, M.; Akiba, Y.; Kurose, I.; Higuchi, H.; Fujimori, H.; Tsuzuki, Y.; et al. Increased expression of an inducible isoform of nitric oxide synthase and the formation of peroxynitrite in colonic mucosa of patients with active ulcerative colitis. Gut 1998, 42, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.P.; Amstad, P.; Raja, K.; Ambs, S.; Nagashima, M.; Bennett, W.P.; Shields, P.G.; Ham, A.J.; Swenberg, J.A.; Marrogi, A.J.; et al. Increased p53 mutation load in noncancerous colon tissue from ulcerative colitis: A cancer-prone chronic inflammatory disease. Cancer Res. 2000, 60, 3333–3337. [Google Scholar] [PubMed]
- Sadeghi, M.R.; Jeddi, F.; Soozangar, N.; Somi, M.H.; Samadi, N. The role of Nrf2-Keap1 axis in colorectal cancer, progression, and chemoresistance. Tumor Biol. 2017, 39, 1010428317705510. [Google Scholar] [CrossRef] [PubMed]
- Song, C.H.; Kim, N.; Lee, S.M.; Nam, R.H.; Choi, S.I.; Kang, S.R.; Shin, E.; Lee, D.H.; Lee, H.N.; Surh, Y.J. Effects of 17β-estradiol on colorectal cancer development after azoxymethane/dextran sulfate sodium treatment of ovariectomized mice. Biochem. Pharmacol. 2019, 164, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Spitz, M.R.; Durphorne, C.M.; Detry, M.A.; Pillow, P.C.; Amos, C.I.; Lei, L.; de Andrade, M.; Gu, X.; Hong, W.K.; Wu, X. Dietary intake of isothiocyanates: Evidence of joint effect with glutathione S-transferase polymorphisms in lung cancer risk. Cancer Epidemiol. Biomark. Prev. 2000, 9, 1017–1020. [Google Scholar]
- Joseph, M.A.; Moysich, K.B.; Freudenheim, J.L.; Shields, P.G.; Bowman, E.D.; Zhang, Y.; Marshall, J.R.; Ambrosone, C.B. Cruciferous vegetables, genetic polymorphisms in glutathione S-transferases M1 and T1, and prostate cancer risk. Nutr. Cancer 2004, 50, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Arisawa, T.; Tahara, T.; Shibata, T.; Nagasaka, M.; Nakamura, M.; Kamiya, Y.; Fujita, H.; Yoshioka, D.; Okubo, M.; Sakata, M.; et al. Nrf2 gene promoter polymorphism is associated with ulcerative colitis in a Japanese population. Hepatogastroenterology 2008, 55, 394–397. [Google Scholar]
- Marzec, J.M.; Christie, J.D.; Reddy, S.P.; Jedlicka, A.E.; Vuong, H.; Lanken, P.N.; Aplenc, R.; Yamamoto, T.; Yamamoto, M.; Cho, H.Y.; et al. Functional polymorphisms in the transcription factor NRF2 in humans increase the risk of acute lung injury. FASEB J. 2007, 21, 2237–2246. [Google Scholar] [CrossRef]
- Yokoo, Y.; Kijima, A.; Ishii, Y.; Takasu, S.; Tsuchiya, T.; Umemura, T. Effects of Nrf2 silencing on oxidative stress-associated intestinal carcinogenesis in mice. Cancer Medicine. 2016, 5, 1228–1238. [Google Scholar] [CrossRef]
- Jang, H.J.; Hong, E.M.; Kim, M.; Kim, J.H.; Jang, J.; Park, S.W.; Byun, H.W.; Koh, D.H.; Choi, M.H.; Kae, S.H.; et al. Simvastatin induces heme oxygenase-1 via NF-E2-related factor 2 (Nrf2) activation through ERK and PI3K/Akt pathway in colon cancer. Oncotarget 2016, 7, 46219–46229. [Google Scholar] [CrossRef]
- Kou, X.; Kirberger, M.; Yang, Y.; Chen, N. Natural products for cancer prevention associated with Nrf2-ARE pathway. Food Sci. Hum. Wellness 2013, 2, 22–28. [Google Scholar] [CrossRef]
- Trivedi, P.P.; Jena, G.B.; Tikoo, K.B.; Kumar, V. Melatonin modulated autophagy and Nrf2 signaling pathways in mice with colitis-associated colon carcinogenesis. Mol. Carcinog. 2016, 55, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Q.; Wu, R.; Xiao, X.; Yang, C.; Yang, Y.; Wang, C.; Lin, L.; Kong, A.N. The dietary flavone luteolin epigenetically activates the Nrf2 pathway and blocks cell transformation in human colorectal cancer HCT116 cells. J. Cell. Biochem. 2018, 119, 9573–9582. [Google Scholar] [CrossRef] [PubMed]
- Kensler, T.W.; Chen, J.G.; Egner, P.A.; Fahey, J.W.; Jacobson, L.P.; Stephenson, K.K.; Ye, L.; Coady, J.L.; Wang, J.B.; Wu, Y.; et al. Effects of glucosinolate-rich broccoli sprouts on urinary levels of aflatoxin-DNA adducts and phenanthrene tetraols in a randomized clinical trial in He Zuo township, Qidong, People’s Republic of China. Cancer Epidemiol. Biomark. Prev. 2005, 4, 2605–2613. [Google Scholar] [CrossRef] [PubMed]
- Seow, A.; Yuan, J.M.; Sun, C.L.; Van Der Berg, D.; Lee, H.P.; Yu, M.C. Dietary isothiocyanates, glutathione S-transferase polymorphisms and colorectal cancer risk in the Singapore Chinese Health Study. Carcinogenesis 2002, 23, 2055–2061. [Google Scholar] [CrossRef] [PubMed]
- Ambrosone, C.B.; McCann, S.E.; Freudenheim, J.L.; Marshall, J.R.; Zhang, Y.; Shields, P.G. Breast cancer risk in premenopausal women is inversely associated with consumption of broccoli, a source of isothiocyanates, but is not modified by GST genotype. J. Nutr. 2004, 134, 1134–1138. [Google Scholar] [CrossRef] [PubMed]
- Rao, C.V.; Rivenson, A.; Zang, E.; Steele, V.; Kelloff, G.; Reddy, B.S. Inhibition of 2-amino-1-methyl-6-phenylimidazo [4,5]-pyridine-induced lymphoma formation by oltipraz. Cancer Res. 1996, 56, 3395–3398. [Google Scholar]
- Moon, R.; Rao, K.; Detrisac, C.; Kelloff, G.; Steele, V.; Doody, L. Chemoprevention of respiratory-tract neoplasia in the hamster by oltipraz, alone and in combination. Int. J. Oncol. 1994, 4, 661–667. [Google Scholar] [CrossRef]
- Nishikawa, A.; Tanakamura, Z.; Furukawa, F.; Lee, I.S.; Kasahara, K.; Ikezaki, S.; Takahashi, M. Chemopreventive activity of oltipraz against induction of glandular stomach carcinogenesis in rats by N-methyl-N-nitro-N-nitrosoguanidine. Carcinogenesis 1998, 19, 365–368. [Google Scholar] [CrossRef]
- Szarka, C.E.; Yao, K.S.; Pfeiffer, G.R.; Balshem, A.M.; Litwin, S.; Frucht, H.; Goosenberg, E.B.; Engstrom, P.F.; Clapper, M.L.; O’Dwyer, P.J. Chronic dosing of oltipraz in people at increased risk for colorectal cancer. Cancer Detect. Prev. 2001, 24, 352–361. [Google Scholar]
- Roebuck, B.D.; Liu, X.L.; Rogers, A.E.; Groopmen, J.D.; Kensler, T.W. Protection against aflatoxin B1-induced hepatocarcinogenesis in F344 rats by 5-(2-pyrazynil)-4-methyl-1,2-dithiole-3thione (oltipraz): Predictive role of short-term moleculary dosimetry. Cancer Res. 1991, 51, 5501–5506. [Google Scholar] [PubMed]
- Kang, K.A.; Piao, M.J.; Hyun, Y.J.; Zhen, A.X.; Cho, S.J.; Ahn, M.J.; Yi, J.M.; Hyun, J.W. Luteolin promotes apoptotic cell death via upregulation of Nrf2 expression by DNA demethylase and the interaction of Nrf2 with p53 in human colon cancer cells. Exp. Mol. Med. 2019, 51, 40. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Saud, S.M.; Zhang, X.; Li, W.; Hua, B. Protective effect of Shaoyao Decoction against colorectal cancer via the Keap1-Nrf2-ARE signaling pathway. J. Ethnopharmacol. 2019, 241, 111981. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.N.; Park, G.H.; Park, S.B.; Kim, J.D.; Eo, H.J.; Son, H.J.; Song, J.H.; Jeong, J.B. Sageretia thea Inhibits Inflammation through Suppression of NF-κ B and MAPK and Activation of Nrf2/HO-1 Signaling Pathways in RAW264.7 Cells. Am. J. Chin. Med. 2019, 47, 385–403. [Google Scholar] [CrossRef] [PubMed]
- Rao, C.V.; Rivenson, A.; Katiwalla, M.; Kelloff, G.J.; Reddy, B.S. Chemopreventive effect of oltipraz during different stages of experimental colon carcinogenesis induced by azoxymethane in male F344 rats. Cancer Res. 1993, 53, 2502–2506. [Google Scholar]
- Clapper, M.L.; Wood, M.; Leahy, K.; Lang, D.; Miknyoczki, S.; Ruggeri, B.A. Chemopreventive activity of Oltipraz against N-nitrosobis (2-oxopropyl) amine (BOP)-induced ductal pancreatic carcinoma development and effects on survival of Syrian golden hamsters. Carcinogenesis 1995, 16, 2159–2165. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Gao, P.; Steele, V.E. The chemopreventive efficacy of inhaled oltipraz particulates in the B[a]P-induced A/J mouse lung adenoma model. Carcinogenesis 2006, 27, 1721–1727. [Google Scholar] [CrossRef][Green Version]
- Kensler, T.W.; Egner, P.A.; Dolan, P.M.; Groopman, J.D.; Roebuck, B.D. Mechanism of protection against aflatoxin tumorigenicity in rats fed 5-(2-pyrazinyl)-4-methyl-1,2-dithiol-3-thione (oltipraz) and related 1,2-dithiol-3-thiones and 1,2-dithiol-3-ones. Cancer Res. 1987, 47, 4271–4277. [Google Scholar]
- Kensler, T.W.; Curphey, T.J.; Maxiutenko, Y.; Roebuck, B.D. Chemoprotection by organosulfur inducers of phase 2 enzymes: Dithiolethiones and dithiins. Drug Metabol. Drug Interact. 2000, 17, 3–22. [Google Scholar] [CrossRef]
- Hu, T.; Hua, T.; Yu, Y.; Yub, S.; Guoa, H.; Hana, L.; Wanga, W.; Tiana, T.; Haoc, Y.; Liud, Z.; et al. Clinicopathologic significance of CXCR4 and Nrf2 in colorectal cancer. J. Biomed. Res. 2013, 27, 283–290. [Google Scholar]
- Zhao, X.Q.; Zhang, Y.F.; Xia, Y.F.; Zhou, Z.M.; Cao, Y.Q. Promoter demethylation of nuclear factor-erythroid 2-related factor 2 gene in drug-resistant colon cancer cells. Oncol. Lett. 2015, 10, 1287–1292. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; He, L.J.; Ye, H.Z.; Liu, D.F.; Zhu, Y.B.; Miao, D.D.; Zhang, S.P.; Chen, Y.Y.; Jia, Y.W.; Shen, J.; et al. Nrf2 is a key factor in the reversal effect of curcumin on multidrug resistance in the HCT-8/5-Fu human colorectal cancer cell line. Mol. Med. Rep. 2018, 18, 5409–5416. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, Z.Y.; Du, H.; Li, S.Z.; Tu, R.; Jia, Y.F.; Zheng, Z.; Song, X.M.; Du, R.L.; Zhang, X.D. DUB3 deubiquitinates and stabilizes NRF2 in chemotherapy resistance of colorectal cancer. Cell Death Differ. 2019. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.O.; Chang, J.T.; Wu, D.W.; Huang, C.C.; Lee, H. Cytoplasmatic localization of Nrf2 promotes colorectal cancer with more aggressive tumors via upregulation of PSMD4. Free Radic. Biol. Med. 2016, 95, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.M.; Lin, P.L.; Wu, D.W.; Wang, L.; Huang, C.C.; Lee, H. PSMD4 is a novel therapeutic target in chemoresistant colorectal cancer activated by cytoplasmic localization of Nrf2. Oncotarget 2018, 9, 26342–26352. [Google Scholar] [CrossRef] [PubMed]
- Tajmohammadi, I.; Mohammadian, J.; Sabzichi, M.; Mahmuodi, S.; Ramezani, M.; Aghajani, M.; Ramezani, F. Identification of Nrf2/STAT3 axis in induction of apoptosis through sub-G 1 cell cycle arrest mechanism in HT-29 colon cancer cells. J. Cell. Biochem. 2019, 120, 14035–14043. [Google Scholar] [CrossRef] [PubMed]
- Cernigliaro, C.; D’Anneo, A.; Carlisi, D.; Giuliano, M.; Marino Gammazza, A.; Barone, R.; Longhitano, L.; Cappello, F.; Emanuele, S.; Distefano, A.; et al. Ethanol-Mediated Stress Promotes Autophagic Survival and Aggressiveness of Colon Cancer Cells via Activation of Nrf2/HO-1 Pathway. Cancers 2019, 11, 505. [Google Scholar] [CrossRef]
- Jeong, G.; Oh, J.; Kim, J.S. Glyceollins Modulate Tumor Development and Growth in a Mouse Xenograft Model of Human Colon Cancer in a p53-Dependent Manner. J. Med. Food 2019, 22, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Taira, J.; Miyazato, H.; Ueda, K. Marine Peroxy Sesquiterpenoids Induce Apoptosis by Modulation of Nrf2-ARE Signaling in HCT116 Colon Cancer Cells. Mar. Drugs 2018, 23, 16. [Google Scholar] [CrossRef]
- Sadeghi, M.R.; Jeddi, F.; Soozanger, N.; Somi, M.H.; Shirmohamadi, M.; Khaze, V.; Samadi, N. Nrf2/P-glycoprotein axis is associated with clinicopathological characteristics in colorectal cancer. Biomed. Pharmacother. 2018, 104, 458–464. [Google Scholar] [CrossRef]
- Ballong, M.J.; Yun, H.; Sherwood, L.; Woods, A.K.; Lairson, L.L.; Schultz, P.G. A small molecule inhibits deregulated NRF2 transcriptional activity in cancer. ACS Chem. Biol. 2015, 10, 2193–2198. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Wang, H.; Chen, F.; Fu, J.; Xu, Y.; Hou, Y.; Kou, H.H.; Zhai, C.; Nelson, M.B.; Zhang, Q.; et al. An overview of chemical inhibitors of the Nrf2-ARE signaling pathway and their potential applications in cancer therapy. Free Radic. Biol. Med. 2016, 99, 544–556. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Zhao, L.; Zhao, Q.; Sun, Y.; Zhang, Y.; Miao, H.; You, Q.; Hu, R.; Guo, Q.L. NF-Kappa B and Nrf2 signaling pathways contribute to Wogonin-mediated inhibition of inflammation-associated colorectal carcinogenesis. J. Cell Death Dis. 2014, 5, e1283. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.P.; Winiarski, B.K.; Sutton, P.A.; Jones, R.P.; Ressel, L.; Duckworth, C.A.; Pritchard, D.M.; Lin, Z.X.; Vicky, F.L.; Tweedle, M.E.; et al. The Nrf2 inhibitor brusatol is a potent antitumour agent in an orthotopic mouse model of colorectal cancer. Oncotarget 2018, 9, 27104–27116. [Google Scholar] [CrossRef] [PubMed]
- Hendersen, D.; Frieson, D.; Zuber, J.; Solomon, S.S. Metformin has positive therapeutic effects in colon cancer and lung cancer. Am. J. Med. Sci. 2017, 354, 246–251. [Google Scholar] [CrossRef]
- Sena, P.; Mancini, S.; Benincasa, M.; Mariani, F.; Palumbo, C.; Roncucci, L. Metformin Induces Apoptosis and Alters Cellular Responses to Oxidative Stresses in Ht29 Colon Cancer Cells: Preliminary Findings. Int. J. Mol. Sci. 2018, 19, 1478. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).