Comprehensive Analysis of Soybean Mosaic Virus P3 Protein Interactors and Hypersensitive Response-Like Lesion-Inducing Protein Function

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
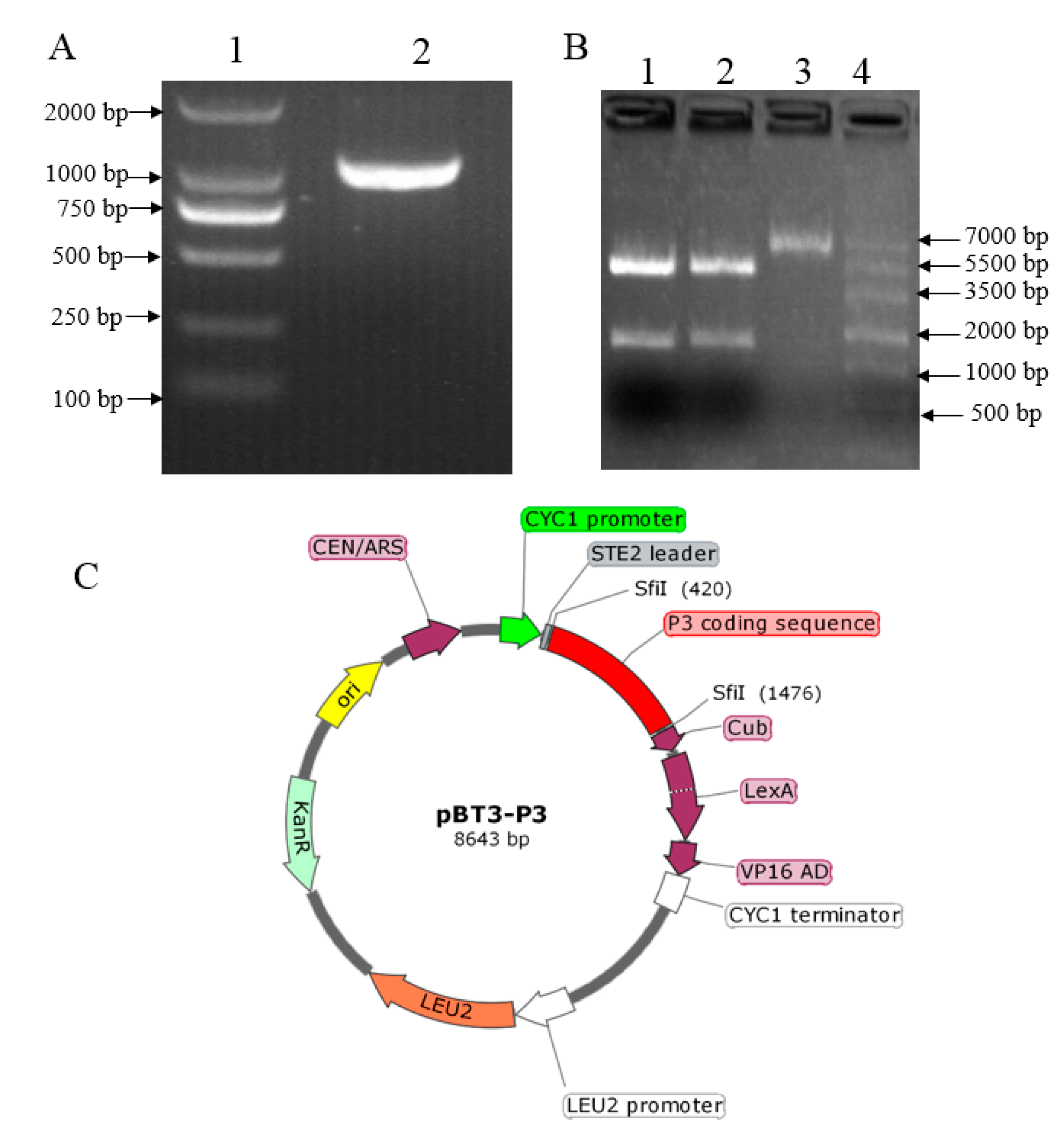
2.1. Construction, Validation, and Auto-Activation of the pBT3-P3 Bait Vector
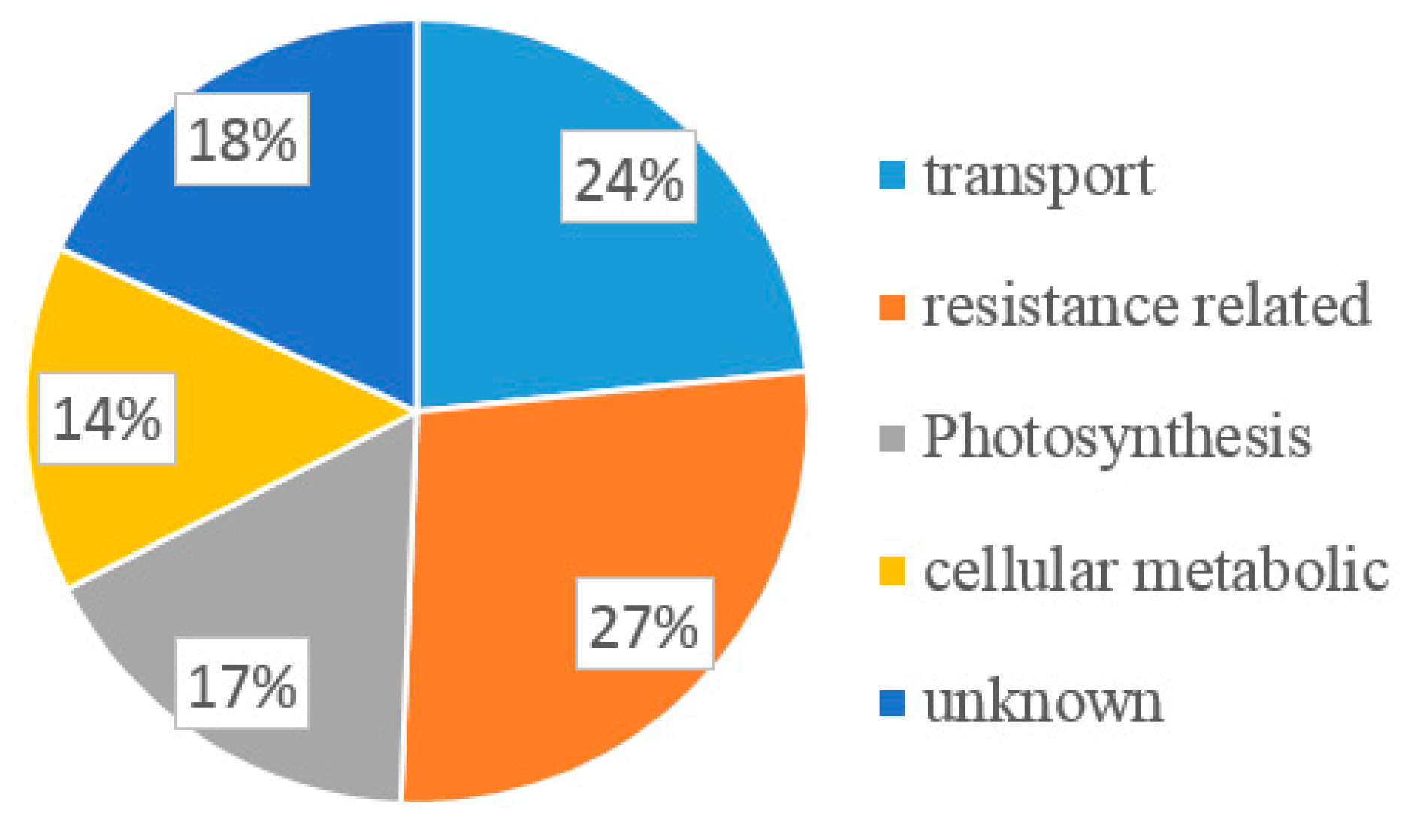
2.2. Selection of SMV P3 Interactors Using Yeast Two Hybrid Analysis
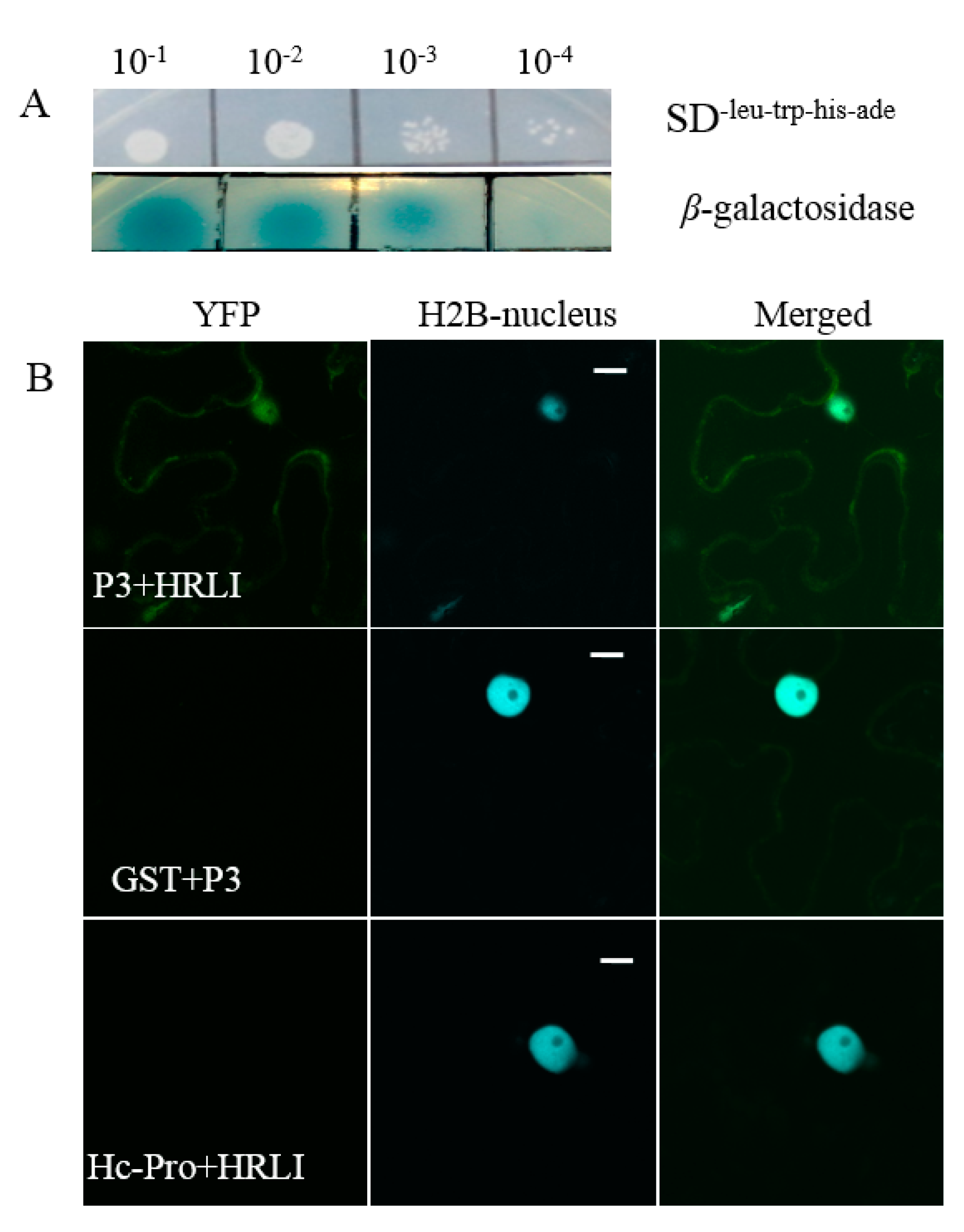
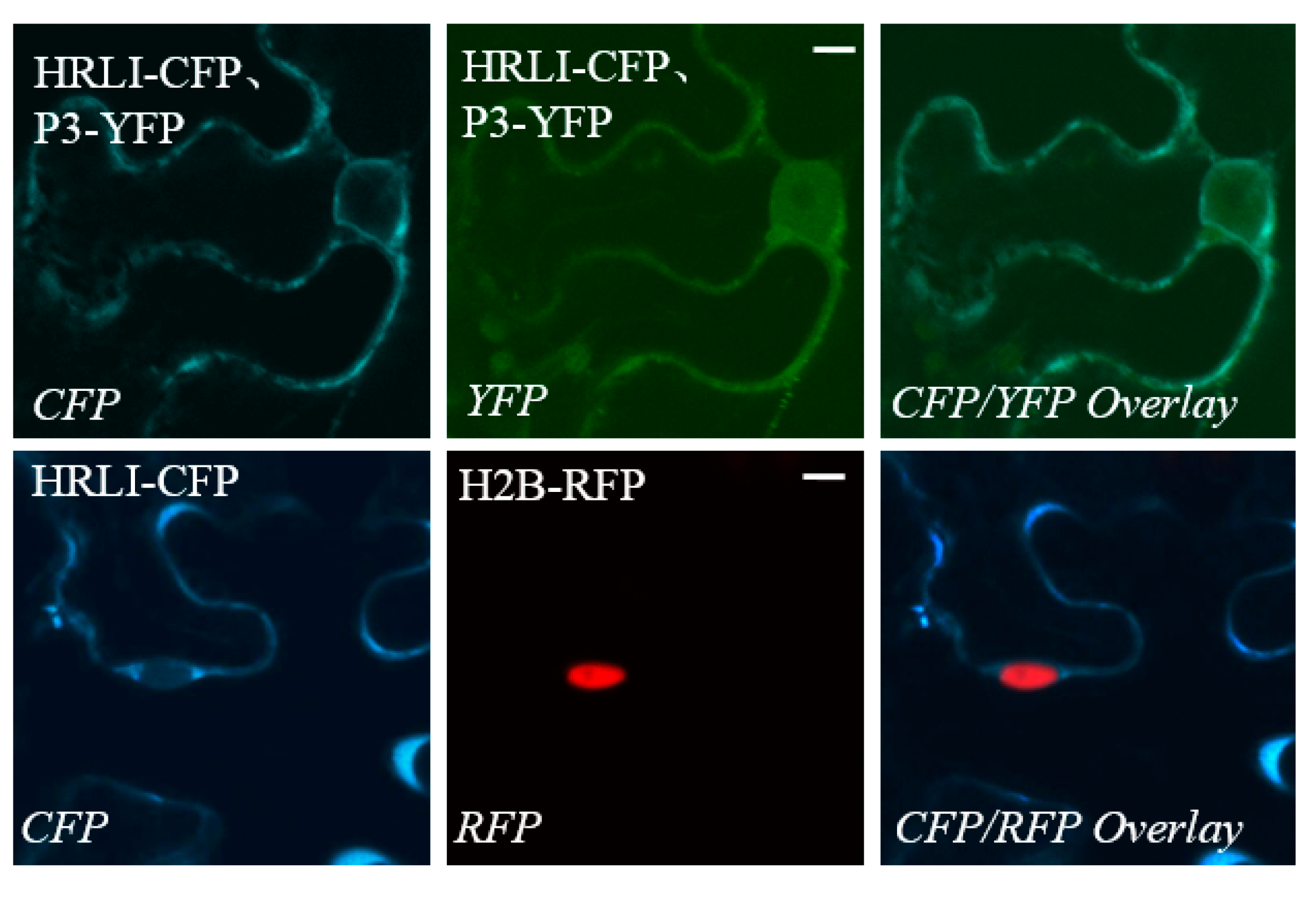
2.3. Verification of the Interaction between GmHRLI and P3 Using BiFCAssay
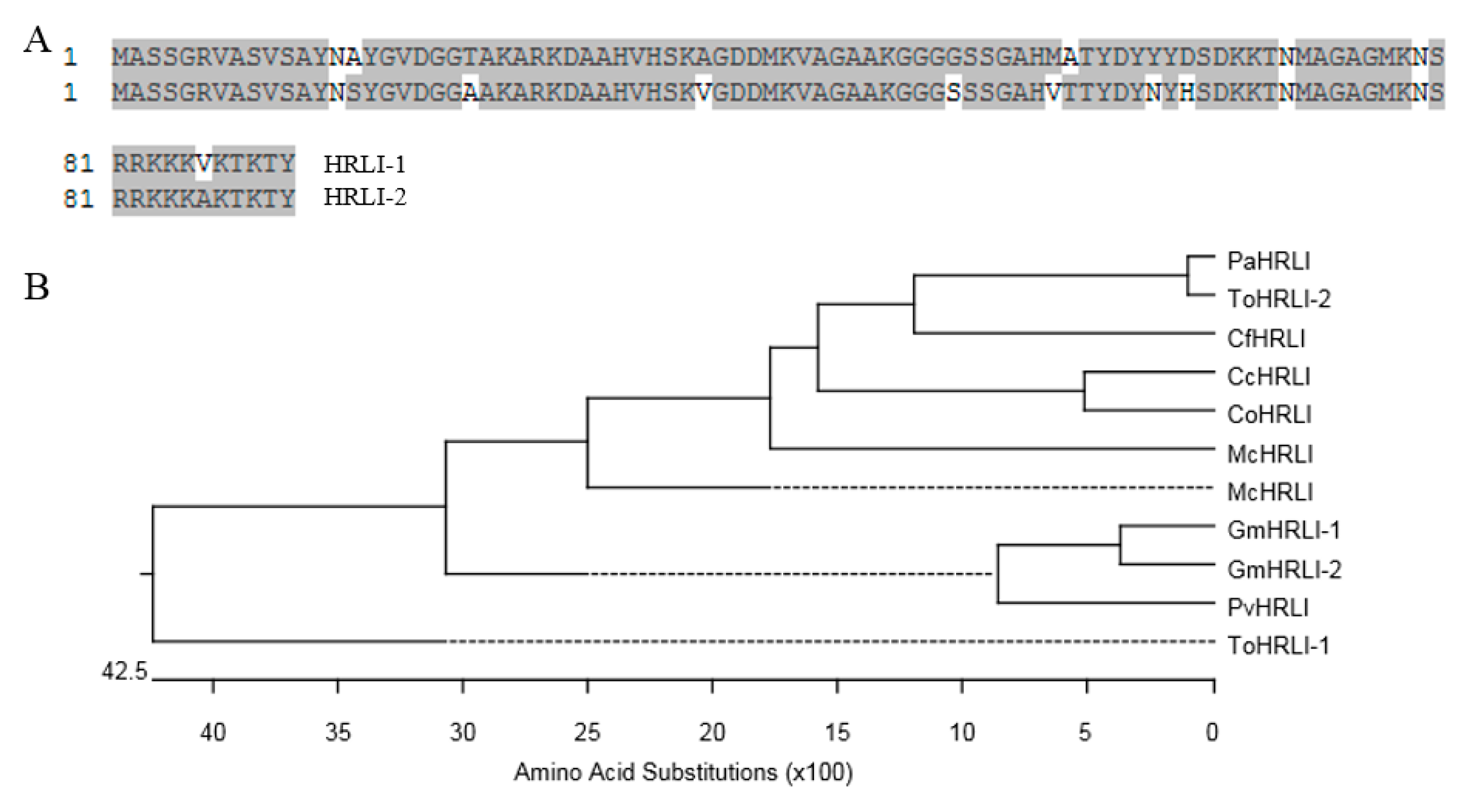
2.4. Sequence Analysis of HRLI Genes
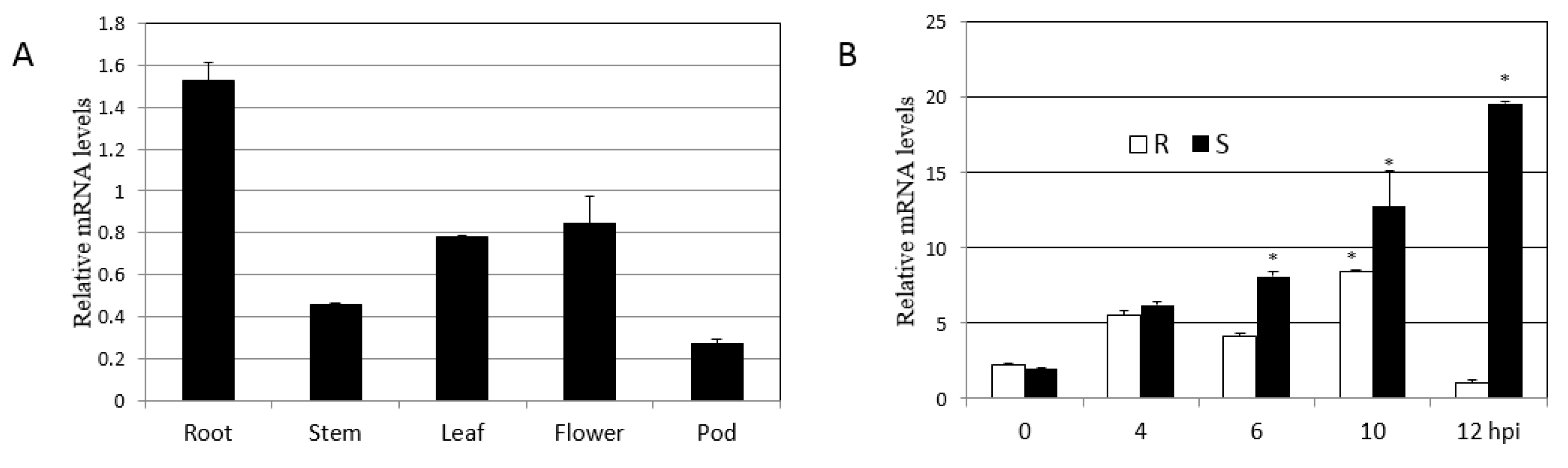
2.5. Expression Patterns of GmHRLI Gene
3. Discussion
4. Materials and Methods
4.1. Plant Growth and Virus Strains
4.2. Soybean cDNA Library and Bait Vector Construction
4.3. Yeast Two-Hybrid Assay
4.4. Bimolecular Fluorescence Complementation and Localization Assays
4.5. Sequence Accessions and Phylogenetic Analysis
4.6. Qutantitative RT-PCR Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Adams, M.J.; Antoniw, J.F.; Fauquet, C.M. Molecular criteria for genus and species discrimination within the family Potyviridae. Arch. Virol. 2005, 150, 459–479. [Google Scholar] [CrossRef] [PubMed]
- Ren, Q.; Pfeiffer, G.W.; Ghabrial, S.A. Soybean mosaic virus resistance improves productivity of double-cropped soybean. Crop Sci. 1997, 37, 1712–1718. [Google Scholar] [CrossRef]
- Abney, T.S.; Sillings, J.O.; Richards, T.L.; Broersma, D.B. Aphids and other insects as vectors of soybean mosaic virus. J. Econ. Entomol. 1976, 69, 254–256. [Google Scholar] [CrossRef]
- Grjr, B.; Goodman, R.M. Soybean mosaic virus: Infection of soybean seed parts and seed transmission. Phytopathology 1979, 69, 569–572. [Google Scholar] [CrossRef]
- Cho, E.K.; Goodman, R.M. Strains of soybean mosaic virus: Classification based on virulence in resistant soybean cultivars. Phytopathology 1979, 69, 467–470. [Google Scholar] [CrossRef]
- Cho, E.K.; Goodman, R.M. Evaluation of resistance in soybeans to soybean mosaic virus strains. Crop Sci. 1982, 22, 1133–1136. [Google Scholar] [CrossRef]
- Wang, X.Q.; Gai, J.Y.; Pu, Z.Q. Classification and distribution of strain groups of soybean mosaic virus in middle and lower Huang-Huai and Changjiang valleys. Soybean Sci. 2003, 22, 102–107. [Google Scholar]
- Li, K.; Yang, Q.H.; Zhi, H.J.; Gai, J.Y. Identification and distribution of soybean mosaic virus strains in southern China. Plant Dis. 2010, 94, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Ma, Y.; Yang, Y.; Liu, N.; Li, C.; Song, Y. Fine mapping and analyses of RSC8 resistance candidate genes to soybean mosaic virus in soybean. Theor. Appl. Genet. 2010, 122, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Ma, Y.; Liu, N.; Yang, Z.; Zheng, G.; Zhi, H. Fine mapping and identification of the soybean RSC4 resistance candidate gene to soybean mosaic virus. Plant Breed. 2011, 130, 653–659. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, D.G.; Li, H.C.; Zheng, G.J.; Yang, Y.Q.; Li, H.W.; Zhi, H.J. Fine mapping of the RSC14Q locus for resistance to soybean mosaic virus in soybean. Euphytica 2011, 181, 127–135. [Google Scholar] [CrossRef]
- Zheng, G.J.; Yang, Y.Q.; Ma, Y.; Yang, X.F.; Chen, S.Y.; Ren, R.; Wang, D.G.; Yang, Z.L.; Zhi, H.J. Fine mapping and candidate gene analysis of resistance gene RSC3Q to soybean mosaic virus in Qihuang 1. J. Integr. Agric. 2014, 13, 2608–2615. [Google Scholar] [CrossRef]
- Riechmann, J.L.; Lain, S.; García, J.A. Highlights and prospects of potyvirus molecular biology. J. Gen. Virol. 1992, 73, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Urcuqui-Inchima, S.; Haenni, A.L.; Bernardi, F. Potyvirus proteins: A wealth of functions. Virus Res. 2001, 74, 157–175. [Google Scholar] [CrossRef]
- Chung, Y.W.; Miller, W.A.; Atkins, J.F.; Firth, A.E. An overlapping essential gene in the Potyviridae. Proc. Natl. Acad. Sci. USA 2008, 105, 5897–5902. [Google Scholar] [CrossRef] [PubMed]
- Vijayapalani, P.; Maeshima, M.; Nagasaki-Takekuchi, N.; Miller, W.A. Interaction of the trans-frame potyvirus protein P3N-PIPO with host protein PCaP1 facilitates potyvirus movement. PLoS Pathog. 2012, 8, e1002639. [Google Scholar] [CrossRef]
- Dolja, V.V.; Haldeman, R.; Robertson, N.L.; Dougherty, W.G.; Carrington, J.C. Distinct functions of capsid protein in assembly and movement of tobacco etch potyvirus in plants. EMBO J. 1994, 13, 1482–1491. [Google Scholar] [CrossRef]
- Dolja, V.V.; Haldeman-Cahill, R.; Montgomery, A.E.; Vandenbosch, K.A.; Carrington, J.C. Capsid protein determinants involved in cell-to-cell and long distance movement of tobacco etch potyvirus. Virology 1995, 206, 1007–1016. [Google Scholar] [CrossRef]
- Maia, I.G.; Haenni, A.; Bernardi, F. Potyviral HC-Pro: A multifunctional protein. J. Gen. Virol. 1996, 77, 1335–1341. [Google Scholar] [CrossRef]
- Nicolas, O.; Dunnington, S.W.; Gotow, L.F.; Pirone, T.P.; Hellmann, G.M. Variations in the VPg protein allow a potyvirus to overcome va gene resistance in tobacco. Virology 1997, 237, 452–459. [Google Scholar] [CrossRef]
- Rojas, M.R.; Zerbini, F.M.; Allison, R.F.; Gilbertson, R.L.; Lucas, W.J. Capsid protein and helper component-proteinase function as potyvirus cell-to-cell movement proteins. Virology 1997, 237, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Carrington, J.C.; Whitham, S.A. Viral invasion and host defense: Strategies and counter-strategies. Curr. Opin. Plant Biol. 1998, 1, 336–341. [Google Scholar] [CrossRef]
- Wen, R.H.; Hajimorad, M.R. Mutational analysis of the putative pipo of soybean mosaic virus suggests disruption of PIPO protein impedes movement. Virology 2010, 400, 1–7. [Google Scholar] [CrossRef]
- Kasschau, K.D.; Carrington, J.C. A counter defensive strategy of plant viruses: Suppression of posttranscriptional gene silencing. Cell 1998, 95, 461–470. [Google Scholar] [CrossRef]
- Puustinen, P.; Mäkinen, K. Uridylylation of the potyvirus VPg by viral replicase NIb correlates with the nucleotide binding capacity of VPg. J. Biol. Chem. 2004, 279, 38103–38110. [Google Scholar] [CrossRef] [PubMed]
- Eagles, R.M.; Balmori-Melián, E.; Beck, D.L.; Gardner, R.C.; Forster, R.L.S. Characterization of NTPase, RNA-binding and RNA-helicase activities of the cytoplasmic inclusion protein of tamarillo mosaic potyvirus. Eur. J. Biochem. 1994, 224, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Klein, P.G.; Klein, R.R.; Rodríguez-Cerezo, E.; Hunt, A.G.; Shaw, J.G. Mutational analysis of the tobacco vein mottling virus genome. Virology 1994, 204, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Cronin, S.; Verchot, J.; Haldeman-Cahill, R.; Schaad, M.C.; Carrington, J.C. Long-distance movement factor: A transport function of the potyvirus helper component proteinase. Plant Cell. 1995, 7, 549–559. [Google Scholar] [CrossRef] [PubMed]
- LópezMoya, J.J.; Pirone, T.P. Charge changes near the N terminus of the coat protein of two potyviruses affect virus movement. J. Gen. Virol. 1998, 79, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Schaad, M.C.; Haldeman-Cahill, R.; Cronin, S.; Carrington, J.C. Analysis of the VPg-proteinase (NIa) encoded by tobacco etch potyvirus - effects of mutations on subcellular transport, proteolytic processing, and genome amplification. J. Virol. 1996, 70, 7039–7048. [Google Scholar]
- Spetz, C.; Valkonen, J.P.T. Potyviral 6K2 protein long-distance movement and symptom-induction functions are independent and host-specific. Mol. Plant Microbe Int. 2004, 17, 502–510. [Google Scholar] [CrossRef]
- Eggenberger, A.L.; Hajimorad, M.R.; Hill, J.H. Gain of virulence on Rsv1-genotype soybean by an avirulent soybean mosaic virus requires concurrent mutations in both P3 and HC-Pro. Mol. Plant Microbe Int. 2008, 21, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Hajimorad, M.R.; Eggenberger, A.L.; Hill, J.H. Adaptation of soybean mosaic virus avirulent chimeras containing P3 sequences from virulent strains to Rsv1-genotype soybeans is mediated by mutations in HC-Pro. Mol. Plant Microbe Int. 2008, 21, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Hajimorad, M.R.; Eggenberger, A.L.; Tsang, S.; Whitham, S.A.; Hill, J.H. Cytoplasmic inclusion cistron of soybean mosaic virus serves as a virulence determinant on rsv3-genotype soybean and a symptom determinant. Virology 2009, 391, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Luan, H.; Shine, M.B.; Cui, X.; Chen, X.; Ma, N.; Kachroo, P.; Kachroo, A.; Zhi, H. The potyviral P3 protein targets eukaryotic elongation factor 1A to promote the unfolded protein response and viral pathogenesis. Plant Physiol. 2016, 172, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Luo, Z.; Yan, F.; Lu, Y.; Zheng, H.; Chen, J. Interaction between potyvirus P3 and ribulose-1, 5-bisphosphate carboxylase/oxygenase (RubisCO) of host plants. Virus Genes 2011, 43, 90–92. [Google Scholar] [CrossRef]
- Lu, L.; Wu, G.; Xu, X.; Luan, H.; Zhi, H.; Cui, J.; Cui, X.; Chen, X. Soybean actin-depolymerizing factor 2 interacts with soybean mosaic virus-encoded P3 protein. Virus Genes 2015, 50, 333–339. [Google Scholar] [CrossRef]
- Merits, A.; Guo, D.; Järvekülg, L.; Saarma, M. Biochemical and genetic evidence for interactions between potato A potyvirus-encoded proteins P1 and P3 and proteins of the putative replication complex. Virology 1999, 263, 15–22. [Google Scholar] [CrossRef]
- Johansen, I.E.; Lund, O.S.; Hjulsager, C.K.; Laursen, J. Recessive resistance in Pisum sativum and potyvirus pathotype resolved in a gene-for-cistron correspondence between host and virus. J. Virol. 2001, 75, 6609–6614. [Google Scholar] [CrossRef]
- Suehiro, N.; Natsuaki, T.; Watanabe, T.; Okuda, S. An important determinant of the ability of turnip mosaic virus to infect Brassica spp. and/or Raphanus sativus is in its P3 protein. J. Gen. Virol. 2004, 85, 2087–2098. [Google Scholar] [CrossRef]
- Dallot, S.; Quiot-Douine, L.; Sáenz, P.; Cervera, M.T.; García, J.A.; Quiot, J.B. Identification of plum pox virus determinants implicated in specific interactions with different prunus spp. Phytopathology 2001, 91, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Chowda-Reddy, R.V.; Sun, H.; Chen, H.; Poysa, V.; Ling, H.; Gijzen, M.; Wang, A. Mutations in the P3 protein of soybean mosaic virus G2 isolates determine virulence on Rsv4-genotype soybean. Mol. Plant Microbe Int. 2011, 24, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Wen, R.H.; Khatabi, B.; Ashfield, T.; Saghai, M.M.A.; Hajimorad, M.R. The HC-Pro and P3 cistrons of an avirulent soybean mosaic virus are recognized by different resistance genes at the complex Rsv1 locus. Mol. Plant Microbe Int. 2013, 26, 203. [Google Scholar] [CrossRef] [PubMed]
- Wen, R.; Saghai, M.M.A.; Hajimorad, M.R. Amino acid changes in P3, and not the overlapping pipo-encoded protein, determine virulence of soybean mosaic virus on functionally immune Rsv1-genotype soybean. Mol. Plant Pathol. 2011, 12, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Devadas, S.K.; Enyedi, A.; Raina, R. The Arabidopsis hrl1 mutation reveals novel overlapping roles for salicylic acid, jasmonic acid and ethylene signalling in cell death and defence against pathogens. Plant J. 2010, 30, 467–480. [Google Scholar] [CrossRef]
- Dutta, A.; Chan, S.H.; Pauli, N.T.; Raina, R. Hypersensitive response-like lesions 1 codes for AtPPT1 and regulates accumulation of ROS and defense against bacterial pathogen Pseudomonas syringae in Arabidopsis thaliana. Antioxid. Redox Signal. 2015, 22, 785. [Google Scholar] [CrossRef]
- Melillo, M.T.; Leonetti, P.; Bongiovanni, M.; Castagnone-Sereno, P.; Bleve-Zacheo, T. Modulation of reactive oxygen species activities and H2O2 accumulation during compatible and incompatible tomato-root-knot nematode interactions. New Phytol. 2010, 170, 501–512. [Google Scholar] [CrossRef]
- Kim, B.M.; Lotter-Stark, H.C.T.; Rybicki, E.P.; Chikwamba, R.K.; Palmer, K.E. Characterization of the hypersensitive response-like cell death phenomenon induced by targeting antiviral lectin Griffithsin to the secretory pathway. Plant Biotechnol. J. 2018, 16, 1811–1821. [Google Scholar] [CrossRef]
- Hou, H.; Hu, Y.; Wang, Q.; Xu, X.; Qian, Y.; Zhou, X. Gene expression profiling shows that NbFDN1 is involved in modulating the hypersensitive response-like cell death induced by the Oat dwarf virus RepA protein. Mol. Plant Microbe Int. 2018, 31, 1006–1020. [Google Scholar] [CrossRef]
- Camagna, M.; Takemoto, D. Hypersensitive Response in Plants; Wiley: New York, NY, USA, 2018. [Google Scholar]
- Zhao, W.; Li, X.; Liu, W.H.; Zhao, J.; Jin, Y.M.; Sui, T.T. Construction of high-quality Caco-2 three-frame cDNA library and its application to yeast two-hybrid for the human astrovirus protein-protein interaction. J. Virol. Methods 2014, 205, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Chen, X.; Wu, B.; Gao, L.; Zhi, H.; Cui, X. Identification for soybean host factors interacting with P3N-PIPO protein of Soybean mosaic virus. Acta Physiol. Plant. 2016, 38, 131. [Google Scholar] [CrossRef]
- Selote, D.; Kachroo, A. RPG1-B-derived resistance to AvrB-expressing Pseudomonas syringae requires RIN4-like proteins in soybean. Plant Physiol. 2010, 153, 1199–1211. [Google Scholar] [CrossRef] [PubMed]
- Martin, K.; Kopperud, K.; Chakrabarty, R.; Banerjee, R.; Brooks, R.; Goodin, M.M. Transient expression in Nicotiana benthamiana fluorescent marker lines provides enhanced definition of protein localization, movement and interactions in planta. Plant J. 2010, 59, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Goodin, M.M.; Chakrabarty, R.; Yelton, S.; Martin, K.; Clark, A.; Brooks, R. Membrane and protein dynamics in live plant nuclei infected with Sonchus yellow net virus, a plant-adapted rhabdovirus. J. Gen. Virol. 2007, 88, 1810–1820. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP: Phylogenetic Analysis Using Parsimony and Other Methods, Software Version 4.0 b10; Sinaur Associates: Sunderland, MA, USA, 2000.






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luan, H.; Liao, W.; Niu, H.; Cui, X.; Chen, X.; Zhi, H. Comprehensive Analysis of Soybean Mosaic Virus P3 Protein Interactors and Hypersensitive Response-Like Lesion-Inducing Protein Function. Int. J. Mol. Sci. 2019, 20, 3388. https://doi.org/10.3390/ijms20143388
Luan H, Liao W, Niu H, Cui X, Chen X, Zhi H. Comprehensive Analysis of Soybean Mosaic Virus P3 Protein Interactors and Hypersensitive Response-Like Lesion-Inducing Protein Function. International Journal of Molecular Sciences. 2019; 20(14):3388. https://doi.org/10.3390/ijms20143388
Chicago/Turabian StyleLuan, Hexiang, Wenlin Liao, Haopeng Niu, Xiaoyan Cui, Xin Chen, and Haijian Zhi. 2019. "Comprehensive Analysis of Soybean Mosaic Virus P3 Protein Interactors and Hypersensitive Response-Like Lesion-Inducing Protein Function" International Journal of Molecular Sciences 20, no. 14: 3388. https://doi.org/10.3390/ijms20143388
APA StyleLuan, H., Liao, W., Niu, H., Cui, X., Chen, X., & Zhi, H. (2019). Comprehensive Analysis of Soybean Mosaic Virus P3 Protein Interactors and Hypersensitive Response-Like Lesion-Inducing Protein Function. International Journal of Molecular Sciences, 20(14), 3388. https://doi.org/10.3390/ijms20143388