Expression and Functional Role of TRPV4 in Bone Marrow-Derived CD11c+ Cells

 , ,
, ,  and
and
Abstract
1. Introduction
2. Results
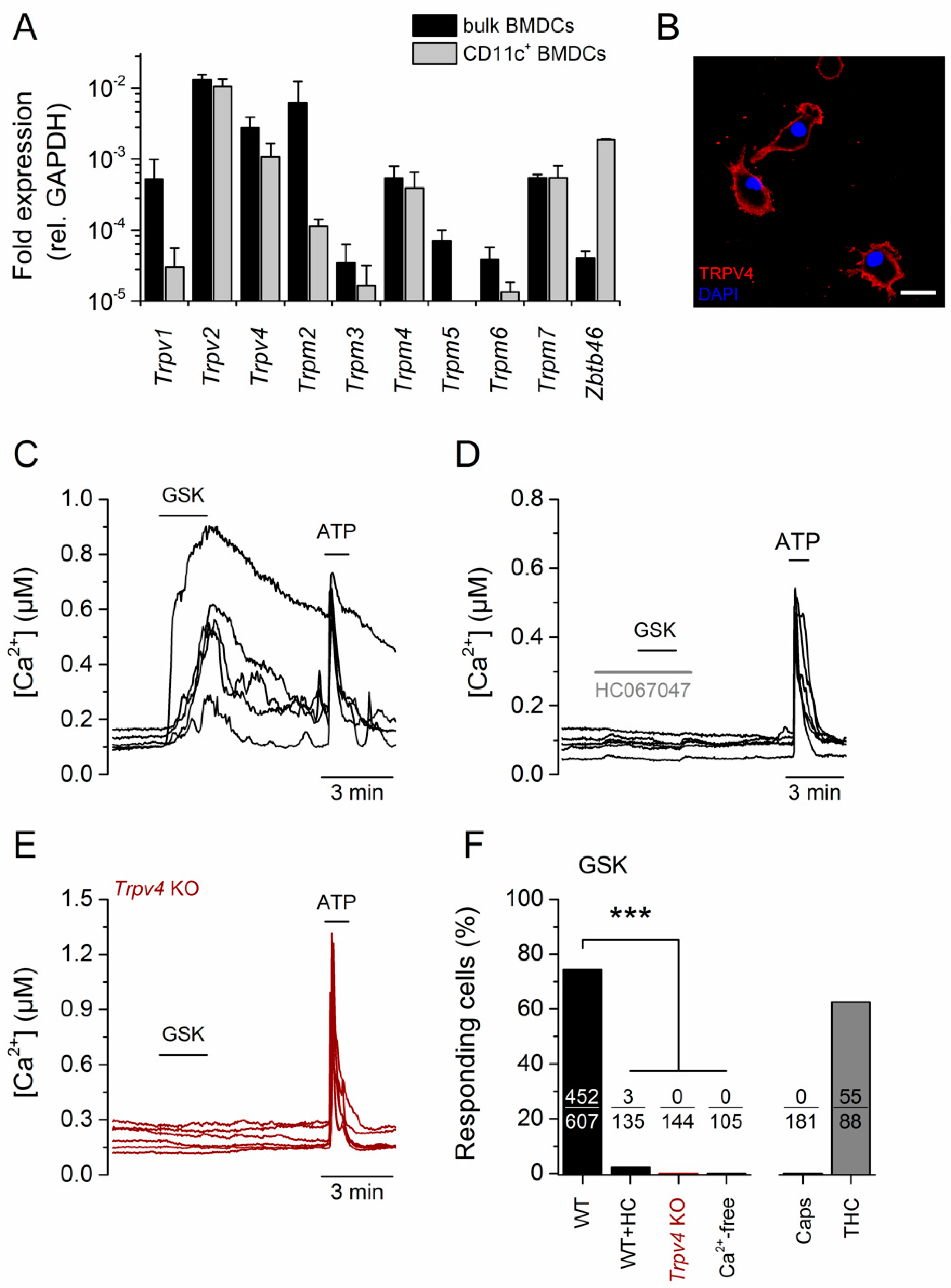
2.1. TRPV4 Was Functionally Expressed in CD11c+ BMDCs
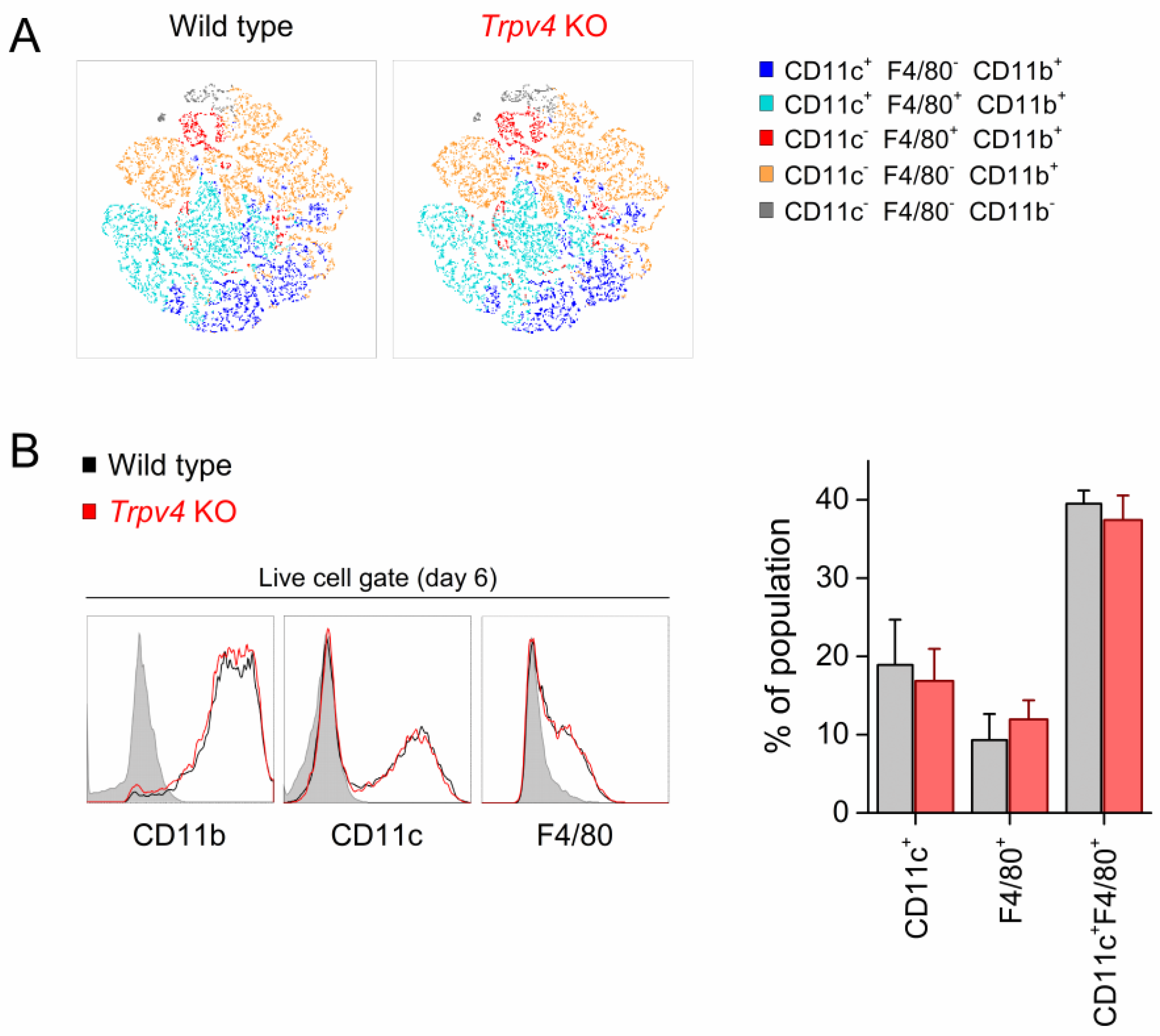
2.2. Differentiation of BMDCs Was Independent of TRPV4
2.3. TRPV4 Was Downregulated in Mature BMDCs
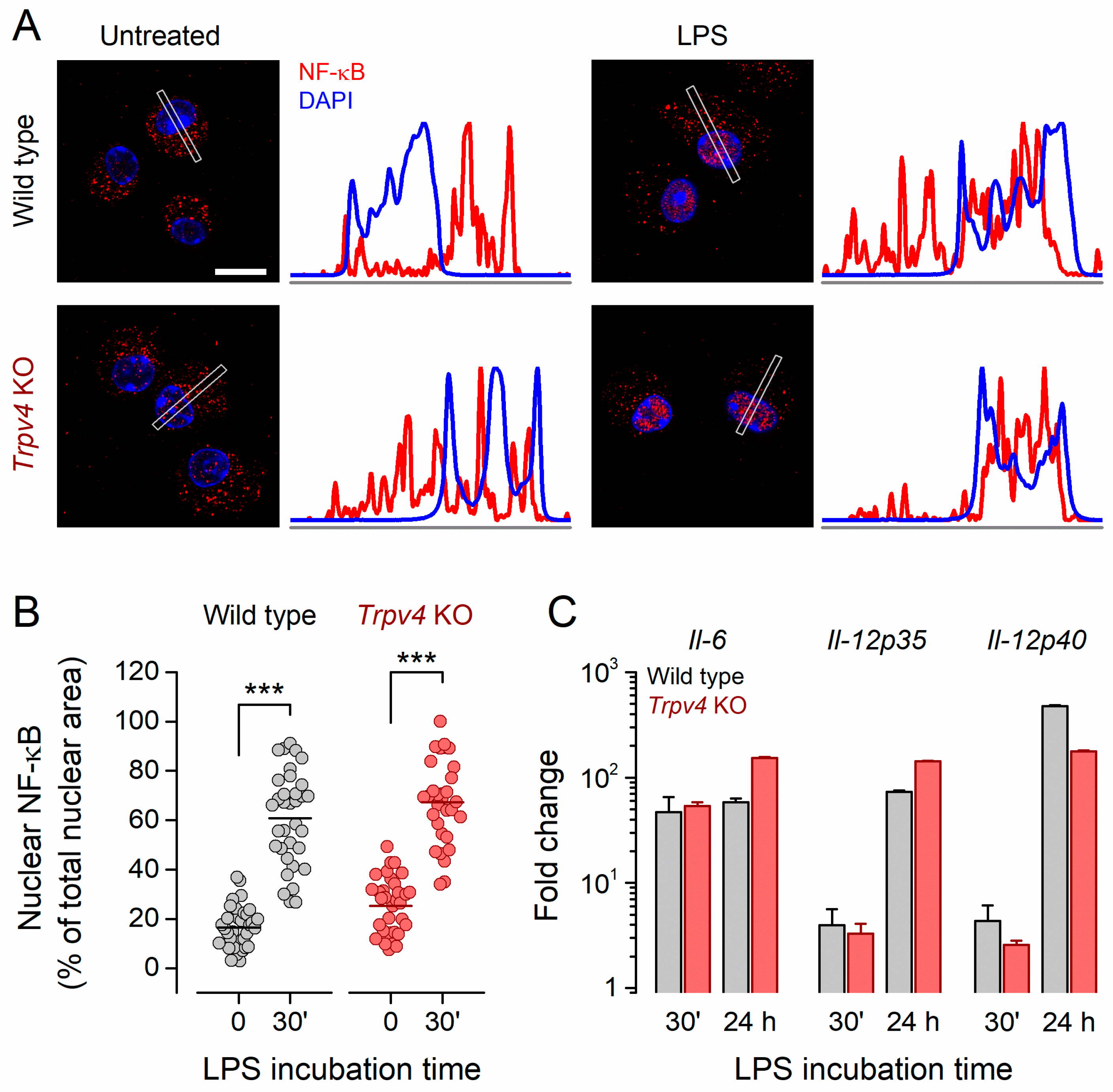
2.4. NF-κB Nuclear Translocation and Inflammatory Gene Expression Was Unaltered in Trpv4 KO CD11c+ BMDCs
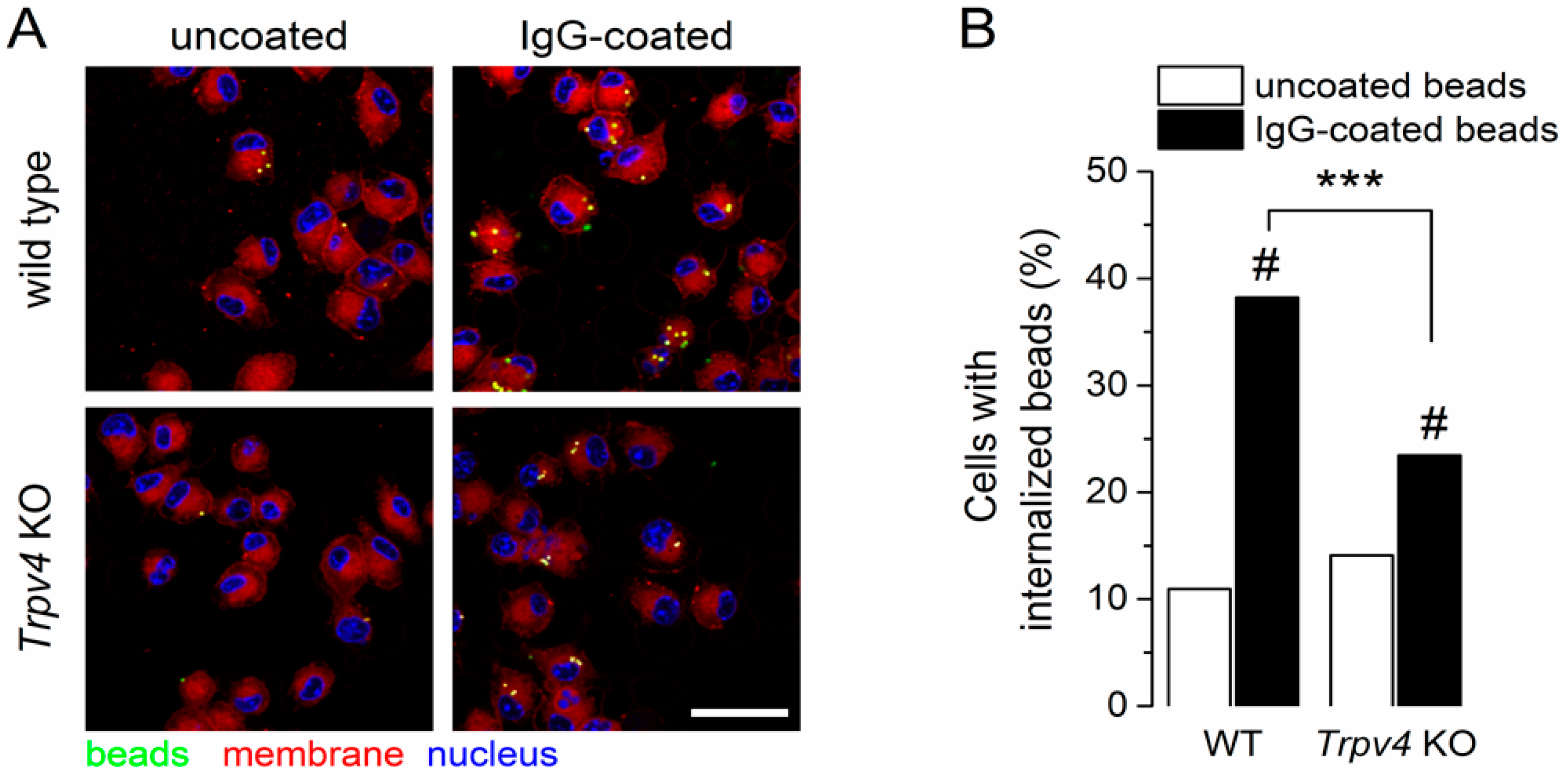
2.5. TRPV4 Contributed to the Receptor-Mediated Phagocytosis of Immature CD11c+ BMDCs
3. Discussion
4. Materials and Methods
4.1. Cells and Transfection
4.2. Animals
4.3. Isolation of Bone Marrow-Derived CD11c+ Cells
4.4. RNA Isolation and qPCR
4.5. Confocal Imaging
4.6. Intracellular Ca2+ Imaging
4.7. FACS
4.8. Nuclear Extraction and Western Blot
4.9. Phagocytosis Assay
4.10. Data and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Banchereau, J.; Briere, F.; Caux, C.; Davoust, J.; Lebecque, S.; Liu, Y.J.; Pulendran, B.; Palucka, K. Immunobiology of dendritic cells. Annu. Rev. Immunol. 2000, 18, 767–811. [Google Scholar] [CrossRef] [PubMed]
- Shumilina, E.; Huber, S.M.; Lang, F. Ca2+ signaling in the regulation of dendritic cell functions. Am. J. Physiol. Cell Physiol. 2011, 300, C1205–C1214. [Google Scholar] [CrossRef] [PubMed]
- Bagley, K.C.; Abdelwahab, S.F.; Tuskan, R.G.; Lewis, G.K. Calcium signaling through phospholipase C activates dendritic cells to mature and is necessary for the activation and maturation of dendritic cells induced by diverse agonists. Clin. Diagn. Lab. Immunol. 2004, 11, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Feske, S.; Gwack, Y.; Prakriya, M.; Srikanth, S.; Puppel, S.H.; Tanasa, B.; Hogan, P.G.; Lewis, R.S.; Daly, M.; Rao, A. A mutation in Orai1 causes immune deficiency by abrogating CRAC channel function. Nature 2006, 441, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Vig, M.; Peinelt, C.; Beck, A.; Koomoa, D.L.; Rabah, D.; Koblan-Huberson, M.; Kraft, S.; Turner, H.; Fleig, A.; Penner, R.; et al. CRACM1 is a plasma membrane protein essential for store-operated Ca2+ entry. Science 2006, 312, 1220–1223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.L.; Yeromin, A.V.; Zhang, X.H.; Yu, Y.; Safrina, O.; Penna, A.; Roos, J.; Stauderman, K.A.; Cahalan, M.D. Genome-wide RNAi screen of Ca2+ influx identifies genes that regulate Ca2+ release-activated Ca2+ channel activity. Proc. Natl. Acad. Sci. USA 2006, 103, 9357–9362. [Google Scholar] [CrossRef]
- Matzner, N.; Zemtsova, I.M.; Nguyen, T.X.; Duszenko, M.; Shumilina, E.; Lang, F. Ion channels modulating mouse dendritic cell functions. J. Immunol. 2008, 181, 6803–6809. [Google Scholar] [CrossRef]
- Vaeth, M.; Zee, I.; Concepcion, A.R.; Maus, M.; Shaw, P.; Portal-Celhay, C.; Zahra, A.; Kozhaya, L.; Weidinger, C.; Philips, J.; et al. Ca2+ Signaling but Not Store-Operated Ca2+ Entry Is Required for the Function of Macrophages and Dendritic Cells. J. Immunol. 2015, 195, 1202–1217. [Google Scholar] [CrossRef]
- Sumoza-Toledo, A.; Lange, I.; Cortado, H.; Bhagat, H.; Mori, Y.; Fleig, A.; Penner, R.; Partida-Sanchez, S. Dendritic cell maturation and chemotaxis is regulated by TRPM2-mediated lysosomal Ca2+ release. FASEB J. 2011, 25, 3529–3542. [Google Scholar] [CrossRef]
- Szollosi, A.G.; Olah, A.; Toth, I.B.; Papp, F.; Czifra, G.; Panyi, G.; Biro, T. Transient receptor potential vanilloid-2 mediates the effects of transient heat shock on endocytosis of human monocyte-derived dendritic cells. FEBS Lett. 2013, 587, 1440–1445. [Google Scholar] [CrossRef]
- Scheraga, R.G.; Abraham, S.; Niese, K.A.; Southern, B.D.; Grove, L.M.; Hite, R.D.; McDonald, C.; Hamilton, T.A.; Olman, M.A. TRPV4 Mechanosensitive Ion Channel Regulates Lipopolysaccharide-Stimulated Macrophage Phagocytosis. J. Immunol. 2016, 196, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Satpathy, A.T.; Kc, W.; Albring, J.C.; Edelson, B.T.; Kretzer, N.M.; Bhattacharya, D.; Murphy, T.L.; Murphy, K.M. Zbtb46 expression distinguishes classical dendritic cells and their committed progenitors from other immune lineages. J. Exp. Med. 2012, 209, 1135–1152. [Google Scholar] [CrossRef] [PubMed]
- Toth, B.I.; Benko, S.; Szollosi, A.G.; Kovacs, L.; Rajnavolgyi, E.; Biro, T. Transient receptor potential vanilloid-1 signaling inhibits differentiation and activation of human dendritic cells. FEBS Lett. 2009, 583, 1619–1624. [Google Scholar] [CrossRef] [PubMed]
- Thorneloe, K.S.; Sulpizio, A.C.; Lin, Z.; Figueroa, D.J.; Clouse, A.K.; McCafferty, G.P.; Chendrimada, T.P.; Lashinger, E.S.; Gordon, E.; Evans, L.; et al. N-((1S)-1-{[4-((2S)-2-{[(2,4-dichlorophenyl)sulfonyl]amino}-3-hydroxypropanoyl)-1-piperazinyl]carbonyl}-3-methylbutyl)-1-benzothiophene-2-carboxamide (GSK1016790A), a novel and potent transient receptor potential vanilloid 4 channel agonist induces urinary bladder contraction and hyperactivity: Part I. J. Pharmacol. Exp. Ther. 2008, 326, 432–442. [Google Scholar] [PubMed]
- Chiang, C.Y.; Veckman, V.; Limmer, K.; David, M. Phospholipase Cgamma-2 and intracellular calcium are required for lipopolysaccharide-induced Toll-like receptor 4 (TLR4) endocytosis and interferon regulatory factor 3 (IRF3) activation. J. Biol. Chem. 2012, 287, 3704–3709. [Google Scholar] [CrossRef] [PubMed]
- Helft, J.; Bottcher, J.; Chakravarty, P.; Zelenay, S.; Huotari, J.; Schraml, B.U.; Goubau, D.; Reis e Sousa, C. GM-CSF Mouse Bone Marrow Cultures Comprise a Heterogeneous Population of CD11c+MHCII+ Macrophages and Dendritic Cells. Immunity 2015, 42, 1197–1211. [Google Scholar] [CrossRef] [PubMed]
- Inaba, K.; Inaba, M.; Romani, N.; Aya, H.; Deguchi, M.; Ikehara, S.; Muramatsu, S.; Steinman, R.M. Generation of large numbers of dendritic cells from mouse bone marrow cultures supplemented with granulocyte/macrophage colony-stimulating factor. J. Exp. Med. 1992, 176, 1693–1702. [Google Scholar] [CrossRef] [PubMed]
- Lutz, M.B.; Kukutsch, N.; Ogilvie, A.L.; Rossner, S.; Koch, F.; Romani, N.; Schuler, G. An advanced culture method for generating large quantities of highly pure dendritic cells from mouse bone marrow. J. Immunol. Methods 1999, 223, 77–92. [Google Scholar] [CrossRef]
- Barbet, G.; Demion, M.; Moura, I.C.; Serafini, N.; Leger, T.; Vrtovsnik, F.; Monteiro, R.C.; Guinamard, R.; Kinet, J.P.; Launay, P. The calcium-activated nonselective cation channel TRPM4 is essential for the migration but not the maturation of dendritic cells. Nat. Immunol. 2008, 9, 1148–1156. [Google Scholar] [CrossRef]
- Korade-Mirnics, Z.; Corey, S.J. Src kinase-mediated signaling in leukocytes. J. Leukoc. Biol. 2000, 68, 603–613. [Google Scholar]
- Santana, C.; Noris, G.; Espinoza, B.; Ortega, E. Protein tyrosine phosphorylation in leukocyte activation through receptors for IgG. J. Leukoc. Biol. 1996, 60, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Fitzer-Attas, C.J.; Lowry, M.; Crowley, M.T.; Finn, A.J.; Meng, F.; DeFranco, A.L.; Lowell, C.A. Fcgamma receptor-mediated phagocytosis in macrophages lacking the Src family tyrosine kinases Hck, Fgr, and Lyn. J. Exp. Med. 2000, 191, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J. Inositol trisphosphate and calcium signalling. Nature 1993, 361, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.; Lorenzo, I.M.; Andrade, Y.N.; Garcia-Elias, A.; Serra, S.A.; Fernandez-Fernandez, J.M.; Valverde, M.A. IP3 sensitizes TRPV4 channel to the mechano- and osmotransducing messenger 5′-6′-epoxyeicosatrienoic acid. J. Cell Biol. 2008, 181, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Harraz, O.F.; Longden, T.A.; Hill-Eubanks, D.; Nelson, M.T. PIP2 depletion promotes TRPV4 channel activity in mouse brain capillary endothelial cells. eLife 2018, 7, e38689. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Hamada-Nakahara, S.; Itoh, Y.; Takemura, K.; Shimada, A.; Ueda, Y.; Kitamata, M.; Matsuoka, R.; Hanawa-Suetsugu, K.; Senju, Y.; et al. TRPV4 channel activity is modulated by direct interaction of the ankyrin domain to PI(4,5)P2. Nat. Commun. 2014, 5, 4994. [Google Scholar] [CrossRef]
- Sechi, A.S.; Wehland, J. The actin cytoskeleton and plasma membrane connection: PtdIns(4,5)P2 influences cytoskeletal protein activity at the plasma membrane. J. Cell Sci. 2000, 113, 3685–3695. [Google Scholar]
- Jaumouille, V.; Farkash, Y.; Jaqaman, K.; Das, R.; Lowell, C.A.; Grinstein, S. Actin cytoskeleton reorganization by Syk regulates Fcgamma receptor responsiveness by increasing its lateral mobility and clustering. Dev. Cell 2014, 29, 534–546. [Google Scholar] [CrossRef]
- Servin-Vences, M.R.; Moroni, M.; Lewin, G.R.; Poole, K. Direct measurement of TRPV4 and PIEZO1 activity reveals multiple mechanotransduction pathways in chondrocytes. eLife 2017, 6, e21074. [Google Scholar] [CrossRef]
- Defacque, H.; Bos, E.; Garvalov, B.; Barret, C.; Roy, C.; Mangeat, P.; Shin, H.W.; Rybin, V.; Griffiths, G. Phosphoinositides regulate membrane-dependent actin assembly by latex bead phagosomes. Mol. Biol. Cell 2002, 13, 1190–1202. [Google Scholar] [CrossRef][Green Version]
- Link, T.M.; Park, U.; Vonakis, B.M.; Raben, D.M.; Soloski, M.J.; Caterina, M.J. TRPV2 has a pivotal role in macrophage particle binding and phagocytosis. Nat. Immunol. 2010, 11, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Platt, C.D.; Ma, J.K.; Chalouni, C.; Ebersold, M.; Bou-Reslan, H.; Carano, R.A.; Mellman, I.; Delamarre, L. Mature dendritic cells use endocytic receptors to capture and present antigens. Proc. Natl. Acad. Sci. USA 2010, 107, 4287–4292. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Srivastava, P. Immunological role of neuronal receptor vanilloid receptor 1 expressed on dendritic cells. Proc. Natl. Acad. Sci. USA 2005, 102, 5120–5125. [Google Scholar] [CrossRef] [PubMed]
- Assas, B.M.; Wakid, M.H.; Zakai, H.A.; Miyan, J.A.; Pennock, J.L. Transient receptor potential vanilloid 1 expression and function in splenic dendritic cells: A potential role in immune homeostasis. Immunology 2016, 147, 292–304. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, P.J.; Pingle, S.C.; Ahern, G.P. Dendritic cells do not transduce inflammatory stimuli via the capsaicin receptor TRPV1. FEBS Lett. 2005, 579, 5135–5139. [Google Scholar] [CrossRef] [PubMed]
- Wissenbach, U.; Bodding, M.; Freichel, M.; Flockerzi, V. Trp12, a novel Trp related protein from kidney. FEBS Lett. 2000, 485, 127–134. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Probe ID (in Applied Biosytems) |
|---|---|
| Trpa1 | Mm00625268_m1 |
| Trpv1 | Mm01246302_m1 |
| Trpv2 | Mm00449223_m1 |
| Trpv3 | Mm00454996_m1 |
| Trpv4 | Mm00499025_m1 |
| Trpv5 | Mm01166037_m1 |
| Trpv6 | Mm00499069_m1 |
| Trpm1 | Mm00450619_m1 |
| Trpm2 | Mm00663098_m1 |
| Trpm3 | Mm00616485_m1 |
| Trpm4 | Mm00613173_m1 |
| Trpm5 | Mm01129032_m1 |
| Trpm6 | Mm00463112_m1 |
| Trpm7 | Mm00457998_m1 |
| Trpm8 | Mm00454566_m1 |
| Trpc1 | Mm00441975_m1 |
| Trpc2 | Mm00441984_m1 |
| Trpc3 | Mm0044690_m1 |
| Trpc4 | Mm00444284_m1 |
| Trpc5 | Mm00437183_m1 |
| Trpc6 | Mm01176083_m1 |
| Trpc7 | Mm00442606_m1 |
| Zbtb46 | Mm00511327_m1 |
| Il-6 | Mm00446190_m1 |
| Il-12a | Mm00434169_m1 |
| Il-12b | Mm01288989_m1 |
| Gapdh | Mm99999915_g1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naert, R.; López-Requena, A.; Voets, T.; Talavera, K.; Alpizar, Y.A. Expression and Functional Role of TRPV4 in Bone Marrow-Derived CD11c+ Cells. Int. J. Mol. Sci. 2019, 20, 3378. https://doi.org/10.3390/ijms20143378
Naert R, López-Requena A, Voets T, Talavera K, Alpizar YA. Expression and Functional Role of TRPV4 in Bone Marrow-Derived CD11c+ Cells. International Journal of Molecular Sciences. 2019; 20(14):3378. https://doi.org/10.3390/ijms20143378
Chicago/Turabian StyleNaert, Robbe, Alejandro López-Requena, Thomas Voets, Karel Talavera, and Yeranddy A. Alpizar. 2019. "Expression and Functional Role of TRPV4 in Bone Marrow-Derived CD11c+ Cells" International Journal of Molecular Sciences 20, no. 14: 3378. https://doi.org/10.3390/ijms20143378
APA StyleNaert, R., López-Requena, A., Voets, T., Talavera, K., & Alpizar, Y. A. (2019). Expression and Functional Role of TRPV4 in Bone Marrow-Derived CD11c+ Cells. International Journal of Molecular Sciences, 20(14), 3378. https://doi.org/10.3390/ijms20143378