Periostin Mediates Right Ventricular Failure through Induction of Inducible Nitric Oxide Synthase Expression in Right Ventricular Fibroblasts from Monocrotaline-Induced Pulmonary Arterial Hypertensive Rats

Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Expression of POSTN Was Increased in RVs of MCT-Injected Rats
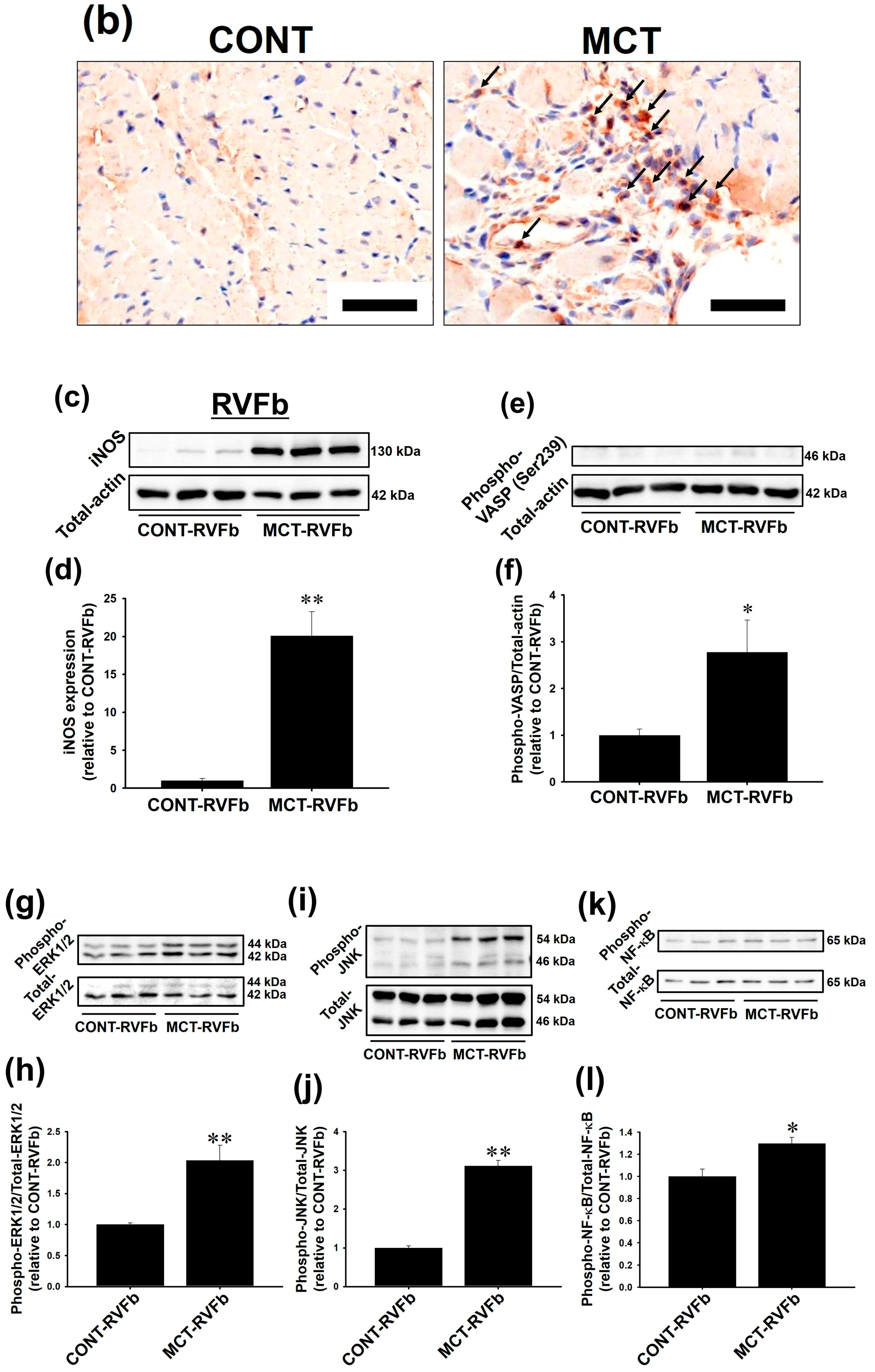
2.2. Expression of iNOS and Phosphorylation of Extracellular sSignal-Regulated Kinase 1/2 (ERK1/2), c-Jun N-Terminal Kinase (JNK) and NF-κB Were Enhanced in MCT-RVFb
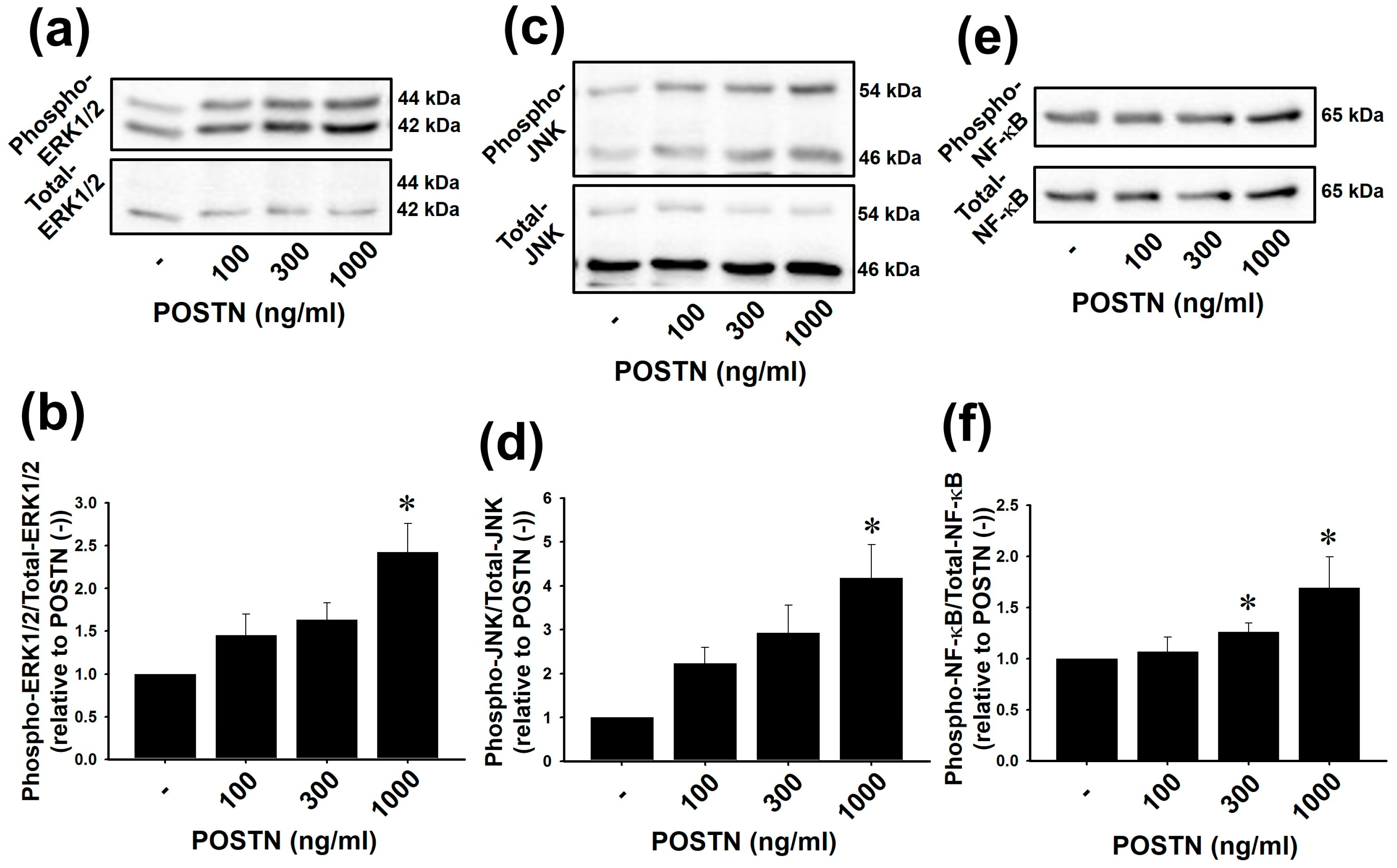
2.3. Recombinant Rat POSTN Increased Phosphorylation of ERK1/2, JNK and NF-κB in RVFbs
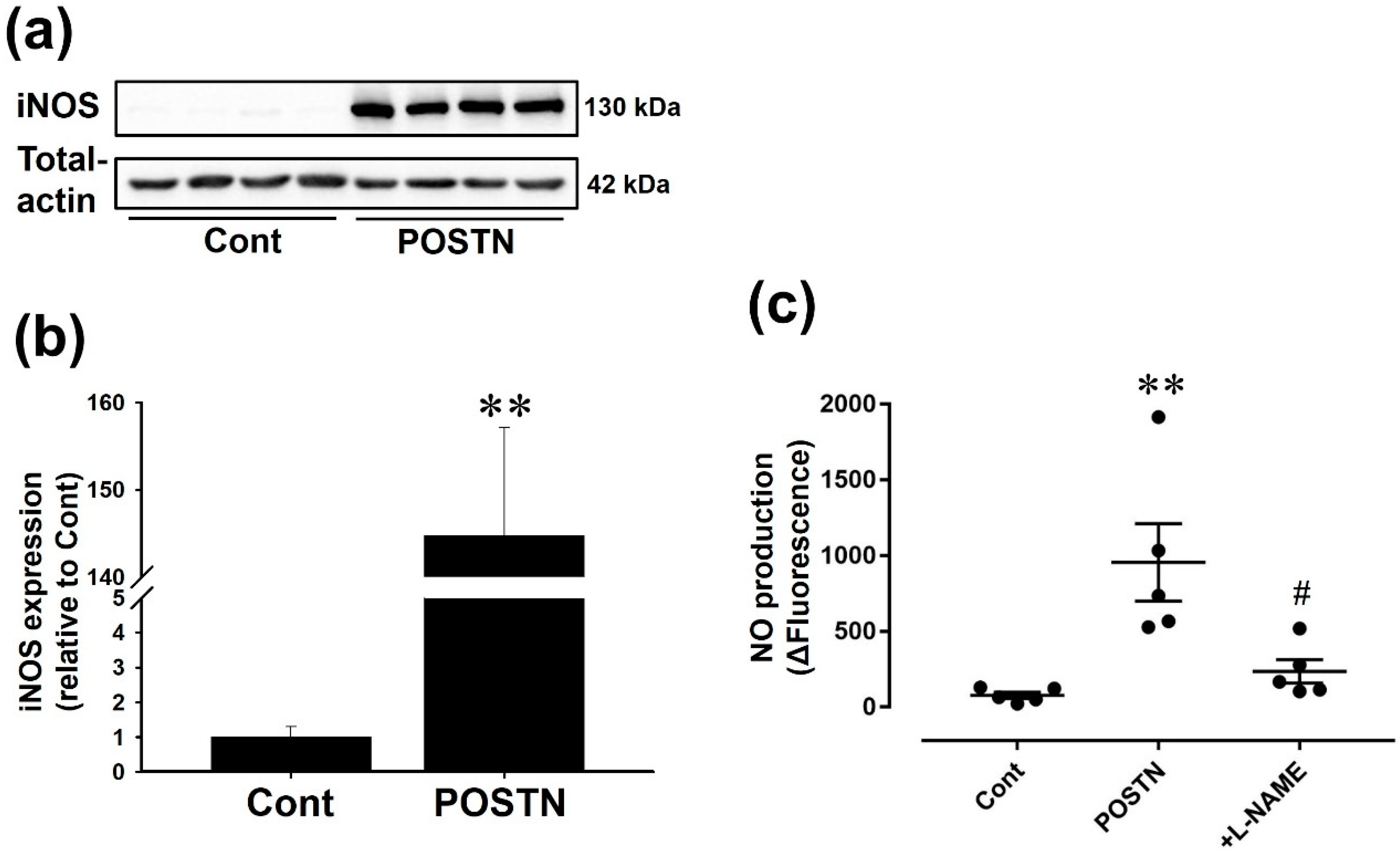
2.4. Recombinant Rat POSTN Induced iNOS Expression and NO Production in RVFbs
2.5. Pharmacological Inhibition of NF-κB, ERK1/2 or JNK Suppressed the POSTN-Induced iNOS Expression and NO Production in RVFbs
2.6. Culture Media (CM) Derived from POSTN-Stimulated RVFbs Inhibited LTCC Activity of H9c2 Cardiomyoblasts
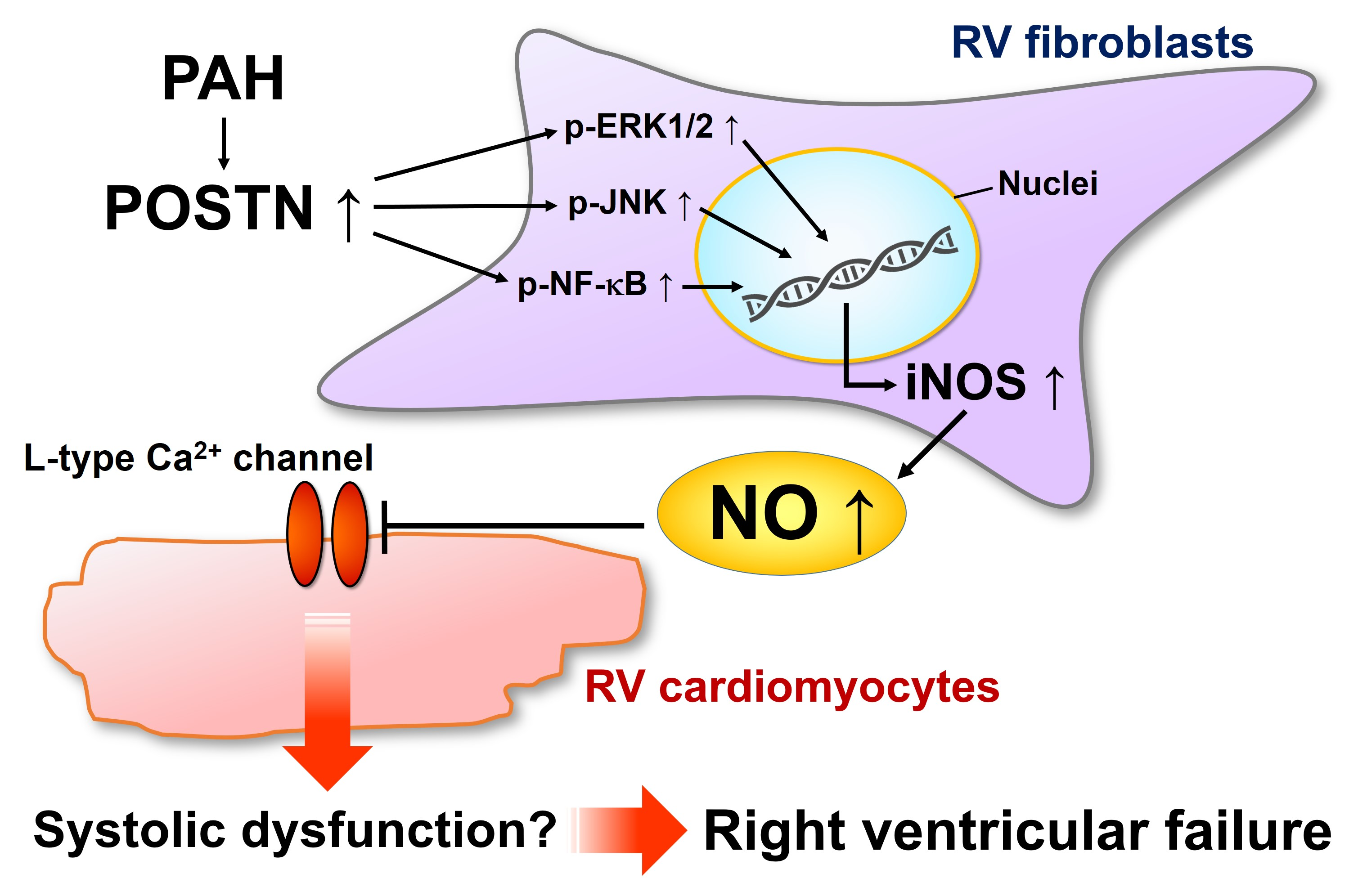
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Production of Recombinant Rat POSTN Protein in E. coli
4.3. Animals
4.4. MCT-Induced PAH Model Rats
4.5. Cell Culture
4.5.1. Isolation of RVFbs
4.5.2. Cell Culture
4.6. Western Blotting
4.7. Histology
4.8. NO Measurement
4.9. Measurement of Ca2+ Inflow through LTCC
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lan, N.; Massam, B.; Kulkarni, S.; Lang, C. Pulmonary arterial hypertension: Pathophysiology and treatment. Diseases 2018, 6, 38. [Google Scholar] [CrossRef] [PubMed]
- Humbert, M.; Sitbon, O.; Chaouat, A.; Bertocchi, M.; Habib, G.; Gressin, V.; Yaïci, A.; Weitzenblum, E.; Cordier, J.-F.; Chabot, F.; et al. Survival in patients with idiopathic, familial, and anorexigen-associated pulmonary arterial hypertension in the modern management era. Circulation 2010, 122, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Benza, R.L.; Miller, D.P.; Barst, R.J.; Badesch, D.B.; Frost, A.E.; McGoon, M.D. An evaluation of long-term survival from time of diagnosis in pulmonary arterial hypertension from the REVEAL Registry. Chest 2012, 142, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Hoeper, M.M.; Kramer, T.; Pan, Z.; Eichstaedt, C.A.; Spiesshoefer, J.; Benjamin, N.; Olsson, K.M.; Meyer, K.; Vizza, C.D.; Vonk-Noordegraaf, A.; et al. Mortality in pulmonary arterial hypertension: Prediction by the 2015 European pulmonary hypertension guidelines risk stratification model. Eur. Respir. J. 2017, 50, 1700740. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, A.; Satoh, T.; Tamura, Y.; Fukuda, K.; Matsubara, H. Survival of japanese patients with idiopathic/heritable pulmonary arterial hypertension. Am. J. Cardiol. 2017, 119, 1479–1484. [Google Scholar] [CrossRef] [PubMed]
- Rain, S.; Handoko, M.L.; Vonk Noordegraaf, A.; Bogaard, H.J.; van der Velden, J.; de Man, F.S. Pressure-overload-induced right heart failure. Pflüg. Arch. Eur. J. Physiol. 2014, 466, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Rain, S.; Handoko, M.L.; Trip, P.; Gan, C.T.-J.; Westerhof, N.; Stienen, G.J.; Paulus, W.J.; Ottenheijm, C.A.C.; Marcus, J.T.; Dorfmüller, P.; et al. Right ventricular diastolic impairment in patients with pulmonary arterial hypertension. Circulation 2013, 128, 2016–2025. [Google Scholar] [CrossRef]
- Takeshita, S.; Kikuno, R.; Tezuka, K.; Amann, E. Osteoblast-specific factor 2: Cloning of a putative bone adhesion protein with homology with the insect protein fasciclin I. Biochem. J. 1993, 294 Pt 1, 271–278. [Google Scholar] [CrossRef]
- Horiuchi, K.; Amizuka, N.; Takeshita, S.; Takamatsu, H.; Katsuura, M.; Ozawa, H.; Toyama, Y.; Bonewald, L.F.; Kudo, A. Identification and characterization of a novel protein, periostin, with restricted expression to periosteum and periodontal ligament and increased expression by transforming growth factor β. J. Bone Miner. Res. 1999, 14, 1239–1249. [Google Scholar] [CrossRef]
- Lindsley, A.; Snider, P.; Zhou, H.; Rogers, R.; Wang, J.; Olaopa, M.; Kruzynska-Frejtag, A.; Koushik, S.V.; Lilly, B.; Burch, J.B.E.; et al. Identification and characterization of a novel Schwann and outflow tract endocardial cushion lineage-restricted periostin enhancer. Dev. Biol. 2007, 307, 340–355. [Google Scholar] [CrossRef]
- Bao, S.; Ouyang, G.; Bai, X.; Huang, Z.; Ma, C.; Liu, M.; Shao, R.; Anderson, R.M.; Rich, J.N.; Wang, X.-F. Periostin potently promotes metastatic growth of colon cancer by augmenting cell survival via the Akt/PKB pathway. Cancer Cell 2004, 5, 329–339. [Google Scholar] [CrossRef]
- Tai, I.T.; Dai, M.; Chen, L.B. Periostin induction in tumor cell line explants and inhibition of in vitro cell growth by anti-periostin antibodies. Carcinogenesis 2005, 26, 908–915. [Google Scholar] [CrossRef] [PubMed]
- Hakuno, D.; Kimura, N.; Yoshioka, M.; Mukai, M.; Kimura, T.; Okada, Y.; Yozu, R.; Shukunami, C.; Hiraki, Y.; Kudo, A.; et al. Periostin advances atherosclerotic and rheumatic cardiac valve degeneration by inducing angiogenesis and MMP production in humans and rodents. J. Clin. Investig. 2010, 120, 2292–2306. [Google Scholar] [CrossRef] [PubMed]
- Lindner, V.; Wang, Q.; Conley, B.A.; Friesel, R.E.; Vary, C.P.H. Vascular injury induces expression of periostin: Implications for vascular cell differentiation and migration. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Norris, R.A.; Potts, J.D.; Yost, M.J.; Junor, L.; Brooks, T.; Tan, H.; Hoffman, S.; Hart, M.M.; Kern, M.J.; Damon, B.; et al. Periostin promotes a fibroblastic lineage pathway in atrioventricular valve progenitor cells. Dev. Dyn. 2009, 238, 1052–1063. [Google Scholar] [CrossRef]
- Ashley, S.L.; Wilke, C.A.; Kim, K.K.; Moore, B.B. Periostin regulates fibrocyte function to promote myofibroblast differentiation and lung fibrosis. Mucosal Immunol. 2017, 10, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Wu, H.; Xia, W.; Chen, X.; Zhu, S.; Zhang, S.; Shao, Y.; Ma, W.; Yang, D.; Zhang, J. Periostin expression is upregulated and associated with myocardial fibrosis in human failing hearts. J. Cardiol. 2014, 63, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Stansfield, W.E.; Andersen, N.M.; Tang, R.-H.; Selzman, C.H. Periostin is a novel factor in cardiac remodeling after experimental and clinical unloading of the failing heart. Ann. Thorac. Surg. 2009, 88, 1916–1921. [Google Scholar] [CrossRef] [PubMed]
- Shimazaki, M.; Nakamura, K.; Kii, I.; Kashima, T.; Amizuka, N.; Li, M.; Saito, M.; Fukuda, K.; Nishiyama, T.; Kitajima, S.; et al. Periostin is essential for cardiac healingafter acute myocardial infarction. J. Exp. Med. 2008, 205, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Oka, T.; Xu, J.; Kaiser, R.A.; Melendez, J.; Hambleton, M.; Sargent, M.A.; Lorts, A.; Brunskill, E.W.; Dorn, G.W.; Conway, S.J.; et al. Genetic manipulation of periostin expression reveals a role in cardiac hypertrophy and ventricular remodeling. Circ. Res. 2007, 101, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Ladage, D.; Yaniz-Galende, E.; Rapti, K.; Ishikawa, K.; Tilemann, L.; Shapiro, S.; Takewa, Y.; Muller-Ehmsen, J.; Schwarz, M.; Garcia, M.J.; et al. Stimulating myocardial regeneration with periostin peptide in large mammals improves function post-myocardial infarction but increases myocardial fibrosis. PLoS ONE 2013, 8, e59656. [Google Scholar] [CrossRef] [PubMed]
- Imoto, K.; Okada, M.; Yamawaki, H. Expression profile of matricellular proteins in hypertrophied right ventricle of monocrotaline-induced pulmonary hypertensive rats. J. Vet. Med. Sci. 2017, 79, 1096–1102. [Google Scholar] [CrossRef] [PubMed]
- Imoto, K.; Okada, M.; Yamawaki, H. Characterization of fibroblasts from hypertrophied right ventricle of pulmonary hypertensive rats. Pflüg. Arch. Eur. J. Physiol. 2018, 470, 1405–1417. [Google Scholar] [CrossRef]
- Umar, S.; van der Laarse, A. Nitric oxide and nitric oxide synthase isoforms in the normal, hypertrophic, and failing heart. Mol. Cell. Biochem. 2010, 333, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Ferreiro, C.R.; Chagas, A.C.P.; Carvalho, M.H.C.; Dantas, A.P.; Scavone, C.; Souza, L.C.B.; Buffolo, E.; da Luz, P.L. Expression of inducible nitric oxide synthase is increased in patients with heart failure due to ischemic disease. Braz. J. Med. Biol. Res. 2004, 37, 1313–1320. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dias, F.A.L.; Urboniene, D.; Yuzhakova, M.A.; Biesiadecki, B.J.; Pena, J.R.; Goldspink, P.H.; Geenen, D.L.; Wolska, B.M. Ablation of iNOS delays cardiac contractile dysfunction in chronic hypertension. Front. Biosci. (Elite Ed.) 2010, 2, 312–324. [Google Scholar] [CrossRef]
- Zhang, P.; Xu, X.; Hu, X.; van Deel, E.D.; Zhu, G.; Chen, Y. Inducible nitric oxide synthase deficiency protects the heart from systolic overload–induced ventricular hypertrophy and congestive heart failure. Circ. Res. 2007, 100, 1089–1098. [Google Scholar] [CrossRef]
- Schulz, R.; Panas, D.L.; Catena, R.; Moncada, S.; Olley, P.M.; Lopaschuk, G.D. The role of nitric oxide in cardiac depression induced by interleukin-1β and tumour necrosis factor-α. Br. J. Pharmacol. 1995, 114, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Chijimatsu, R.; Kunugiza, Y.; Taniyama, Y.; Nakamura, N.; Tomita, T.; Yoshikawa, H. Expression and pathological effects of periostin in human osteoarthritis cartilage. BMC Musculoskelet. Disord. 2015, 16, 215. [Google Scholar] [CrossRef] [PubMed]
- Litvin, J.; Zhu, S.; Norris, R.; Markwald, R. Periostin family of proteins: Therapeutic targets for heart disease. Anat. Rec. Part A Discov. Mol. Cell. Evol. Biol. 2005, 287, 1205–1212. [Google Scholar] [CrossRef]
- Campbell, D.L.; Stamler, J.S.; Strauss, H.C. Redox modulation of l-type calcium channels in ferret ventricular myocytes. Dual mechanism regulation by nitric oxide and S-nitrosothiols. J. Gen. Physiol. 1996, 108, 277–293. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.G.; Rechenmacher, C.E.; Lipsius, S.L. Nitric oxide signaling mediates stimulation of l-type Ca2+ current elicited by withdrawal of acetylcholine in cat atrial myocytes. J. Gen. Physiol. 1998, 111, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, N.; Brun, J.; Rousseau, J.-C.; Duong, L.T.; Ferrari, S.L. Cathepsin K controls cortical bone formation by degrading periostin. J. Bone Miner. Res. 2017, 32, 1432–1441. [Google Scholar] [CrossRef]
- Bonnet, N.; Biver, E.; Chevalley, T.; Rizzoli, R.; Garnero, P.; Ferrari, S.L. Serum levels of a cathepsin-K generated periostin fragment predict incident low-trauma fractures in postmenopausal women independently of BMD and FRAX. J. Bone Miner. Res. 2017, 32, 2232–2238. [Google Scholar] [CrossRef] [PubMed]
- Garnero, P.; Bonnet, N.; Ferrari, S.L. Development of a new immunoassay for human cathepsin K-generated periostin fragments as a serum biomarker for cortical bone. Calcif. Tissue Int. 2017, 101, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Xu, X.; Shi, G.-P.; Chicco, A.J.; Ren, J.; Nair, S. Cathepsin K knockout alleviates pressure overload-induced cardiac hypertrophy. Hypertension 2013, 61, 1184–1192. [Google Scholar] [CrossRef] [PubMed]
- Crawford, J.; Nygard, K.; Gan, B.S.; O’Gorman, D.B. Periostin induces fibroblast proliferation and myofibroblast persistence in hypertrophic scarring. Exp. Dermatol. 2015, 24, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, S.S.; Yuan, S.; Innes, A.L.; Kerr, S.; Woodruff, P.G.; Hou, L.; Muller, S.J.; Fahy, J.V. Roles of epithelial cell-derived periostin in TGF-β activation, collagen production, and collagen gel elasticity in asthma. Proc. Natl. Acad. Sci. USA 2010, 107, 14170–14175. [Google Scholar] [CrossRef] [PubMed]
- Kanaoka, M.; Yamaguchi, Y.; Komitsu, N.; Feghali-Bostwick, C.A.; Ogawa, M.; Arima, K.; Izuhara, K.; Aihara, M. Pro-fibrotic phenotype of human skin fibroblasts induced by periostin via modulating TGF-β signaling. J. Dermatol. Sci. 2018, 90, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Pinto, A.R.; Ilinykh, A.; Ivey, M.J.; Kuwabara, J.T.; D’Antoni, M.L.; Debuque, R.; Chandran, A.; Wang, L.; Arora, K.; Rosenthal, N.A.; et al. Revisiting Cardiac Cellular Composition. Circ. Res. 2016, 118, 400–409. [Google Scholar] [CrossRef]
- Ratajczak-Wrona, W.; Jablonska, E.; Garley, M.; Jablonski, J.; Radziwon, P.; Iwaniuk, A. Role of AP-1 family proteins in regulation of inducible nitric oxide synthase (iNOS) in human neutrophils. J. Immunotoxicol. 2013, 10, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-B.; Lee, W.S.; Shin, J.-S.; Jang, D.S.; Lee, K.T. Xanthotoxin suppresses LPS-induced expression of iNOS, COX-2, TNF-α, and IL-6 via AP-1, NF-κB, and JAK-STAT inactivation in RAW 264.7 macrophages. Int. Immunopharmacol. 2017, 49, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Kleinert, H.; Schwarz, P.M.; Förstermann, U. Regulation of the expression of inducible nitric oxide synthase. Biol. Chem. 2003, 384, 1343–1364. [Google Scholar] [CrossRef] [PubMed]
- Kudo, A. Introductory review: Periostin-gene and protein structure. Cell. Mol. Life Sci. 2017, 74, 4259–4268. [Google Scholar] [CrossRef] [PubMed]
- Masuoka, M.; Shiraishi, H.; Ohta, S.; Suzuki, S.; Arima, K.; Aoki, S.; Toda, S.; Inagaki, N.; Kurihara, Y.; Hayashida, S.; et al. Periostin promotes chronic allergic inflammation in response to Th2 cytokines. J. Clin. Investig. 2012, 122, 2590–2600. [Google Scholar] [CrossRef] [PubMed]
- Ontsuka, K.; Kotobuki, Y.; Shiraishi, H.; Serada, S.; Ohta, S.; Tanemura, A.; Yang, L.; Fujimoto, M.; Arima, K.; Suzuki, S.; et al. Periostin, a matricellular protein, accelerates cutaneous wound repair by activating dermal fibroblasts. Exp. Dermatol. 2012, 21, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Chuanyu, S.; Yuqing, Z.; Chong, X.; Guowei, X.; Xiaojun, Z. Periostin promotes migration and invasion of renal cell carcinoma through the integrin/focal adhesion kinase/c-Jun N-terminal kinase pathway. Tumor Biol. 2017, 39. [Google Scholar] [CrossRef]
- Almanza, A.; Navarrete, F.; Vega, R.; Soto, E. Modulation of voltage-gated Ca2+ current in vestibular hair cells by nitric oxide. J. Neurophysiol. 2007, 97, 1188–1195. [Google Scholar] [CrossRef]
- Hu, H.; Chiamvimonvat, N.; Yamagishi, T.; Marban, E. Direct inhibition of expressed cardiac l-type Ca2+ channels by S-nitrosothiol nitric oxide donors. Circ. Res. 1997, 81, 742–752. [Google Scholar]
- Imoto, K.; Kumatani, S.; Okada, M.; Yamawaki, H. Endostatin is protective against monocrotaline-induced right heart disease through the inhibition of T-type Ca2+ channel. Pflüg. Arch. Eur. J. Physiol. 2016, 468, 1259–1270. [Google Scholar] [CrossRef]
- Okada, M.; Yamawaki, H. Levosimendan inhibits interleukin-1β-induced apoptosis through activation of Akt and inhibition of inducible nitric oxide synthase in rat cardiac fibroblasts. Eur. J. Pharmacol. 2015, 769, 86–92. [Google Scholar] [CrossRef] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imoto, K.; Okada, M.; Yamawaki, H. Periostin Mediates Right Ventricular Failure through Induction of Inducible Nitric Oxide Synthase Expression in Right Ventricular Fibroblasts from Monocrotaline-Induced Pulmonary Arterial Hypertensive Rats. Int. J. Mol. Sci. 2019, 20, 62. https://doi.org/10.3390/ijms20010062
Imoto K, Okada M, Yamawaki H. Periostin Mediates Right Ventricular Failure through Induction of Inducible Nitric Oxide Synthase Expression in Right Ventricular Fibroblasts from Monocrotaline-Induced Pulmonary Arterial Hypertensive Rats. International Journal of Molecular Sciences. 2019; 20(1):62. https://doi.org/10.3390/ijms20010062
Chicago/Turabian StyleImoto, Keisuke, Muneyoshi Okada, and Hideyuki Yamawaki. 2019. "Periostin Mediates Right Ventricular Failure through Induction of Inducible Nitric Oxide Synthase Expression in Right Ventricular Fibroblasts from Monocrotaline-Induced Pulmonary Arterial Hypertensive Rats" International Journal of Molecular Sciences 20, no. 1: 62. https://doi.org/10.3390/ijms20010062
APA StyleImoto, K., Okada, M., & Yamawaki, H. (2019). Periostin Mediates Right Ventricular Failure through Induction of Inducible Nitric Oxide Synthase Expression in Right Ventricular Fibroblasts from Monocrotaline-Induced Pulmonary Arterial Hypertensive Rats. International Journal of Molecular Sciences, 20(1), 62. https://doi.org/10.3390/ijms20010062