Treatment of Periodontal Ligament Stem Cells with MOR and CBD Promotes Cell Survival and Neuronal Differentiation via the PI3K/Akt/mTOR Pathway

 , , , and
, , , and
Abstract
1. Introduction
2. Results
2.1. Cytofluorimetric and Morphological Analysis of hPDLSCs
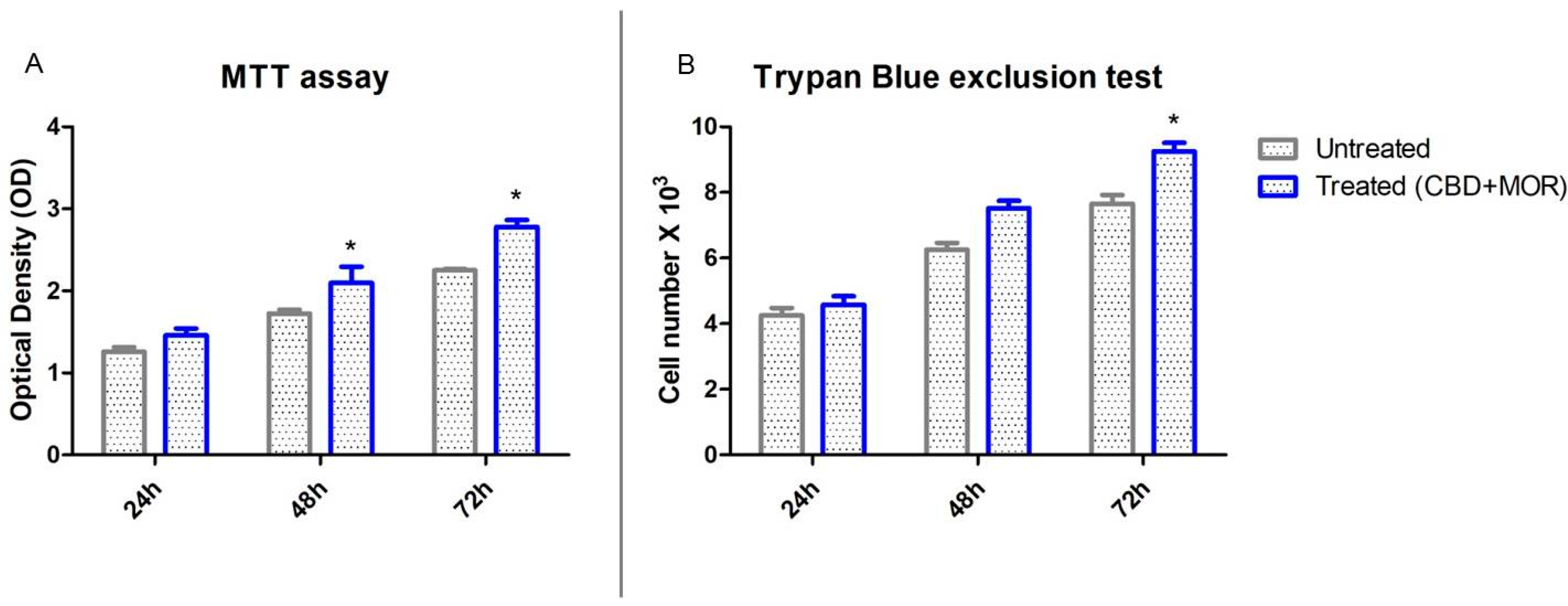
2.2. Cell Viability Assays
2.3. Immunofluorescence Analysis
2.4. NGS Analysis
3. Discussion
3.1. Inhibition of Apoptosis
3.2. PI3K/Akt/mTOR Pathway
3.3. Neuronal Differentiation
4. Materials and Methods
4.1. Ethic Statement
4.2. CBD and Moringin Extraction
4.3. hPDLSCs Culture
4.4. Cytofluorimetric Analysis of hPDLSCs
4.5. Cell Viability Assays
4.6. Immunofluorescence Detection
4.7. RNA Libraries Preparation for NGS Analysis
4.8. Data Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| hPDLSCs | Human Periodontal Ligament Stem Cells |
| MOR | Moringin |
| CBD | Cannabidiol |
| NGS | Next Generation Sequencing |
| MSCs | Mesenchymal Stem Cells |
| CTR | Control |
| MTT | 3-(4,5-dimethylthiazolyl-2)-2,5-diphenyltetrazolium bromide |
References
- Hass, R.; Kasper, C.; Bohm, S.; Jacobs, R. Different populations and sources of human mesenchymal stem cells (MSC): A comparison of adult and neonatal tissue-derived MSC. Cell Commun. Signal. 2011, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Sonoyama, W.; Liu, Y.; Yamaza, T.; Tuan, R.S.; Wang, S.; Shi, S.; Huang, G.T. Characterization of the apical papilla and its residing stem cells from human immature permanent teeth: A pilot study. J. Endod. 2008, 34, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Gronthos, S.; Brahim, J.; Li, W.; Fisher, L.W.; Cherman, N.; Boyde, A.; DenBesten, P.; Robey, P.G.; Shi, S. Stem cell properties of human dental pulp stem cells. J. Dent. Res. 2002, 81, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Miura, M.; Gronthos, S.; Zhao, M.; Lu, B.; Fisher, L.W.; Robey, P.G.; Shi, S. Shed: Stem cells from human exfoliated deciduous teeth. Proc. Natl. Acad. Sci. USA 2003, 100, 5807–5812. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.T.; Gronthos, S.; Shi, S. Mesenchymal stem cells derived from dental tissues vs. Those from other sources: Their biology and role in regenerative medicine. J. Dent. Res. 2009, 88, 792–806. [Google Scholar] [CrossRef] [PubMed]
- Trubiani, O.; Giacoppo, S.; Ballerini, P.; Diomede, F.; Piattelli, A.; Bramanti, P.; Mazzon, E. Alternative source of stem cells derived from human periodontal ligament: A new treatment for experimental autoimmune encephalomyelitis. Stem Cell Res. Ther. 2016, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, A.; Diomede, F.; Cardelli, P.; Bramanti, A.; Scionti, D.; Bramanti, P.; Trubiani, O.; Mazzon, E. Transcriptomic analysis of gingival mesenchymal stem cells cultured on 3D bioprinted scaffold: A promising strategy for neuroregeneration. J. Biomed. Mater. Res. Part A 2018, 106, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Diomede, F.; Caputi, S.; Merciaro, I.; Frisone, S.; D’Arcangelo, C.; Piattelli, A.; Trubiani, O. Pro-inflammatory cytokine release and cell growth inhibition in primary human oral cells after exposure to endodontic sealer. Int. Endod. J. 2014, 47, 864–872. [Google Scholar] [CrossRef] [PubMed]
- Diomede, F.; Rajan, T.S.; Gatta, V.; D’Aurora, M.; Merciaro, I.; Marchisio, M.; Muttini, A.; Caputi, S.; Bramanti, P.; Mazzon, E.; et al. Stemness maintenance properties in human oral stem cells after long-term passage. Stem Cells Int. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Granero-Molto, F.; Weis, J.A.; Longobardi, L.; Spagnoli, A. Role of mesenchymal stem cells in regenerative medicine: Application to bone and cartilage repair. Expert Opin. Biol. Ther. 2008, 8, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Richardson, S.M.; Kalamegam, G.; Pushparaj, P.N.; Matta, C.; Memic, A.; Khademhosseini, A.; Mobasheri, R.; Poletti, F.L.; Hoyland, J.A.; Mobasheri, A. Mesenchymal stem cells in regenerative medicine: Focus on articular cartilage and intervertebral disc regeneration. Methods 2016, 99, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Ballerini, P.; Diomede, F.; Petragnani, N.; Cicchitti, S.; Merciaro, I.; Cavalcanti, M.; Trubiani, O. Conditioned medium from relapsing-remitting multiple sclerosis patients reduces the expression and release of inflammatory cytokines induced by lps-gingivalis in THP-1 and MO3.13 cell lines. Cytokine 2017, 96, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Giacoppo, S.; Thangavelu, S.R.; Diomede, F.; Bramanti, P.; Conti, P.; Trubiani, O.; Mazzon, E. Anti-inflammatory effects of hypoxia-preconditioned human periodontal ligament cell secretome in an experimental model of multiple sclerosis: A key role of IL-37. FASEB J. 2017, 31, 5592–5608. [Google Scholar] [CrossRef] [PubMed]
- Galuppo, M.; Giacoppo, S.; De Nicola, G.R.; Iori, R.; Navarra, M.; Lombardo, G.E.; Bramanti, P.; Mazzon, E. Antiinflammatory activity of glucomoringin isothiocyanate in a mouse model of experimental autoimmune encephalomyelitis. Fitoterapia 2014, 95, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Giacoppo, S.; Iori, R.; Bramanti, P.; Mazzon, E. Topical moringin-cream relieves neuropathic pain by suppression of inflammatory pathway and voltage-gated ion channels in murine model of multiple sclerosis. Mol. Pain 2017, 13. [Google Scholar] [CrossRef] [PubMed]
- Giacoppo, S.; Rajan, T.S.; Iori, R.; Rollin, P.; Bramanti, P.; Mazzon, E. The alpha-cyclodextrin complex of the moringa isothiocyanate suppresses lipopolysaccharide-induced inflammation in raw 264.7 macrophage cells through AKT and p38 inhibition. Inflamm. Res. 2017, 66, 487–503. [Google Scholar] [CrossRef] [PubMed]
- Romeo, L.; Diomede, F.; Gugliandolo, A.; Scionti, D.; Lo Giudice, F.; Lanza Cariccio, V.; Iori, R.; Bramanti, P.; Trubiani, O.; Mazzon, E. Moringin induces neural differentiation in the stem cell of the human periodontal ligament. Sci. Rep. 2018, 8, 9153. [Google Scholar] [CrossRef] [PubMed]
- Rajan, T.S.; Giacoppo, S.; Iori, R.; De Nicola, G.R.; Grassi, G.; Pollastro, F.; Bramanti, P.; Mazzon, E. Anti-inflammatory and antioxidant effects of a combination of cannabidiol and moringin in lps-stimulated macrophages. Fitoterapia 2016, 112, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Libro, R.; Scionti, D.; Diomede, F.; Marchisio, M.; Grassi, G.; Pollastro, F.; Piattelli, A.; Bramanti, P.; Mazzon, E.; Trubiani, O. Cannabidiol modulates the immunophenotype and inhibits the activation of the inflammasome in human gingival mesenchymal stem cells. Front. Physiol. 2016, 7, 559. [Google Scholar] [CrossRef] [PubMed]
- Rajan, T.S.; Giacoppo, S.; Diomede, F.; Ballerini, P.; Paolantonio, M.; Marchisio, M.; Piattelli, A.; Bramanti, P.; Mazzon, E.; Trubiani, O. The secretome of periodontal ligament stem cells from MS patients protects against EAE. Sci. Rep. 2016, 6, 38743. [Google Scholar] [CrossRef] [PubMed]
- Cianci, E.; Recchiuti, A.; Trubiani, O.; Diomede, F.; Marchisio, M.; Miscia, S.; Colas, R.A.; Dalli, J.; Serhan, C.N.; Romano, M. Human periodontal stem cells release specialized proresolving mediators and carry immunomodulatory and prohealing properties regulated by lipoxins. Stem Cells Transl. Med. 2016, 5, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Diomede, F.; Zini, N.; Gatta, V.; Fulle, S.; Merciaro, I.; D’Aurora, M.; La Rovere, R.M.L.; Traini, T.; Pizzicannella, J.; Ballerini, P.; et al. Human periodontal ligament stem cells cultured onto cortico-cancellous scaffold drive bone regenerative process. Eur. Cells Mater. 2016, 32, 181–201. [Google Scholar] [CrossRef]
- Rajan, T.S.; De Nicola, G.R.; Iori, R.; Rollin, P.; Bramanti, P.; Mazzon, E. Anticancer activity of glucomoringin isothiocyanate in human malignant astrocytoma cells. Fitoterapia 2016, 110, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Deveraux, Q.L.; Reed, J.C. Iap family proteins—Suppressors of apoptosis. Genes Dev. 1999, 13, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Verhagen, A.M.; Ekert, P.G.; Pakusch, M.; Silke, J.; Connolly, L.M.; Reid, G.E.; Moritz, R.L.; Simpson, R.J.; Vaux, D.L. Identification of diablo, a mammalian protein that promotes apoptosis by binding to and antagonizing iap proteins. Cell 2000, 102, 43–53. [Google Scholar] [CrossRef]
- Kim, J.W.; Choi, E.J.; Joe, C.O. Activation of death-inducing signaling complex (disc) by pro-apoptotic c-terminal fragment of rip. Oncogene 2000, 19, 4491–4499. [Google Scholar] [CrossRef] [PubMed]
- Wolff, S.; Erster, S.; Palacios, G.; Moll, U.M. P53’s mitochondrial translocation and momp action is independent of puma and bax and severely disrupts mitochondrial membrane integrity. Cell Res. 2008, 18, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Beurel, E.; Jope, R.S. The paradoxical pro- and anti-apoptotic actions of gsk3 in the intrinsic and extrinsic apoptosis signaling pathways. Prog. Neurobiol. 2006, 79, 173–189. [Google Scholar] [CrossRef] [PubMed]
- Giacoppo, S.; Pollastro, F.; Grassi, G.; Bramanti, P.; Mazzon, E. Target regulation of PI3K/AKT/mTOR pathway by cannabidiol in treatment of experimental multiple sclerosis. Fitoterapia 2017, 116, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Ozaita, A.; Puighermanal, E.; Maldonado, R. Regulation of PI3K/AKT/GSK-3 pathway by cannabinoids in the brain. J. Neurochem. 2007, 102, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Brunet, A.; Datta, S.R.; Greenberg, M.E. Transcription-dependent and -independent control of neuronal survival by the pi3k-akt signaling pathway. Cur. Opin. Neurobiol. 2001, 11, 297–305. [Google Scholar] [CrossRef]
- Dudek, H.; Datta, S.R.; Franke, T.F.; Birnbaum, M.J.; Yao, R.; Cooper, G.M.; Segal, R.A.; Kaplan, D.R.; Greenberg, M.E. Regulation of neuronal survival by the serine-threonine protein kinase AKT. Science 1997, 275, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Kuruvilla, R.; Ye, H.; Ginty, D.D. Spatially and functionally distinct roles of the pi3-k effector pathway during ngf signaling in sympathetic neurons. Neuron 2000, 27, 499–512. [Google Scholar] [CrossRef]
- Lim, J.Y.; Park, S.I.; Oh, J.H.; Kim, S.M.; Jeong, C.H.; Jun, J.A.; Lee, K.S.; Oh, W.; Lee, J.K.; Jeun, S.S. Brain-derived neurotrophic factor stimulates the neural differentiation of human umbilical cord blood-derived mesenchymal stem cells and survival of differentiated cells through MAPK/ERK and PI3K/AKT-dependent signaling pathways. J. Neurosci. Res. 2008, 86, 2168–2178. [Google Scholar] [CrossRef] [PubMed]
- Ishizuka, T.; Ozawa, A.; Katsuura, M.; Nomura, S.; Satoh, Y. Effects of muscarinic acetylcholine receptor stimulation on the differentiation of mouse induced pluripotent stem cells into neural progenitor cells. Clin. Exp. Pharmacol. Physiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Van Inzen, W.G.; Peppelenbosch, M.P.; Van Den Brand, M.W.; Tertoolen, L.G.; De Laat, S.W. Neuronal differentiation of embryonic stem cells. Biochim. Biophys. Acta 1996, 1312, 21–26. [Google Scholar] [CrossRef]
- Zemelko, V.I.; Kozhucharova, I.V.; Kovaleva, Z.V.; Domnina, A.P.; Pugovkina, N.A.; Fridlyanskaya, I.I.; Puzanov, M.V.; Anisimov, S.V.; Grinchuk, T.M.; Nikolsky, N.N. Brain-derived neurotrofic factor (BDNF) secretion of human mesenchymal stem cells isolated from bone marrow, endometrium and adipose tissue. Cell Tissue Biol. 2014, 8, 283–291. [Google Scholar] [CrossRef]
- Rajan, T.S.; Scionti, D.; Diomede, F.; Grassi, G.; Pollastro, F.; Piattelli, A.; Cocco, L.; Bramanti, P.; Mazzon, E.; Trubiani, O. Gingival stromal cells as an in vitro model: Cannabidiol modulates genes linked with amyotrophic lateral sclerosis. J. Cell Biochem. 2017, 118, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Taglialatela-Scafati, O.; Pagani, A.; Scala, F.; De Petrocellis, L.; Di Marzo, V.; Grassi, G.; Appendino, G. Cannabimovone, a cannabinoid with a rearranged terpenoid skeleton from hemp. Eur. J. Org. Chem. 2010, 2010, 2067–2072. [Google Scholar] [CrossRef]
- Muller, C.; van Loon, J.; Ruschioni, S.; De Nicola, G.R.; Olsen, C.E.; Iori, R.; Agerbirk, N. Taste detection of the non-volatile isothiocyanate moringin results in deterrence to glucosinolate-adapted insect larvae. Phytochemistry 2015, 118, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Diomede, F.; Merciaro, I.; Martinotti, S.; Cavalcanti, M.F.; Caputi, S.; Mazzon, E.; Trubiani, O. Mir-2861 is involved in osteogenic commitment of human periodontal ligament stem cells grown onto 3D scaffold. J. Biol. Regul. Homeost. Agents 2016, 30, 1009–1018. [Google Scholar] [PubMed]
- Cavalcanti, M.F.; Maria, D.A.; de Isla, N.; Leal-Junior, E.C.; Joensen, J.; Bjordal, J.M.; Lopes-Martins, R.A.; Diomede, F.; Trubiani, O.; Frigo, L. Evaluation of the proliferative effects induced by low-level laser therapy in bone marrow stem cell culture. Photomed. Laser Surg. 2015, 33, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Diomede, F.; Zingariello, M.; Cavalcanti, M.; Merciaro, I.; Pizzicannella, J.; De Isla, N.; Caputi, S.; Ballerini, P.; Trubiani, O. MyD88/ERK/NFkB pathways and pro-inflammatory cytokines release in periodontal ligament stem cells stimulated by porphyromonas gingivalis. Eur. J. Histochem. 2017, 61, 2791. [Google Scholar] [CrossRef] [PubMed]
- Orciani, M.; Trubiani, O.; Guarnieri, S.; Ferrero, E.; Di Primio, R. Cd38 is constitutively expressed in the nucleus of human hematopoietic cells. J. Cell. Biochem. 2008, 105, 905–912. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lineage | Markers | % Of Expression |
|---|---|---|
| Stem cells | Oct3/4 | 94.3 ± 2.2% |
| Sox-2 | 96.4 ± 0.7% | |
| SSEA-4 | 96.4 ± 2.5% | |
| MSCs | CD29 | 97.8 ± 2.4% |
| CD44 | 95.2 ± 3.3% | |
| CD73 | 96.3 ± 1.1% | |
| CD90 | 95.2 ± 3.1% | |
| CD105 | 96.3 ± 3.2% | |
| Hematopoietic | CD14 | ND |
| Cd34 | ND | |
| CD45 | ND |
| Gene | Name | CTR | MOR+CBD | Log2-FoldChange | Q Value |
|---|---|---|---|---|---|
| BCL2A1 | BCL2 related protein A1 | 0.0001 | 2.7356 | 14.739 | 0.000108528 |
| PRPS1 | phosphoribosyl pyrophosphate synthetase 1 | 12.773 | 82.505 | 2.691 | 0.000108528 |
| CDK1 | cyclin dependent kinase 1 | 1.601 | 9.994 | 2.642 | 0.000108528 |
| PRKAG2 | protein kinase AMP-activated non-catalytic subunit gamma 2 | 3.731 | 16.269 | 2.124 | 0.000108528 |
| CYCS | cytochrome c, somatic | 4.446 | 18.7684 | 2.077 | 0.000108528 |
| TNFAIP3 | TNF alpha induced protein 3 | 0.81318 | 3.40641 | 2.066 | 0.000108528 |
| BCL2L1 | BCL2 like 1 | 2.45308 | 9.41279 | 1.940 | 0.000108528 |
| CDC37 | cell division cycle 37 | 51.8182 | 194.202 | 1.906 | 0.000108528 |
| CRADD | CASP2 and RIPK1 domain containing adaptor with death domain | 4.94457 | 11.2444 | 1.185 | 0.000679197 |
| HSPB1 | heat shock protein family B (small) member 1 | 279.982 | 688.126 | 1.297 | 0.000108528 |
| HSP90AA1 | heat shock protein 90 alpha family class A member 1 | 271.592 | 621.726 | 1.195 | 0.000108528 |
| PMAIP1 | phorbol-12-myristate-13-acetate-induced protein 1 | 1.755 | 3.424 | 0.964 | 0.0248542 |
| MAPK1 | mitogen-activated protein kinase 1 | 37.1635 | 61.9931 | 0.738 | 0.000108528 |
| XIAP | X-linked inhibitor of apoptosis | 6.18581 | 10.1329 | 0.712 | 0.000108528 |
| CYC1 | cytochrome c1 | 27.7517 | 43.9185 | 0.662 | 0.000108528 |
| HSPA4 | heat shock protein family A (Hsp70) member 4 | 59.0696 | 88.7283 | 0.586 | 0.000108528 |
| CYLD | CYLD lysine 63 deubiquitinase | 11.8515 | 17.1827 | 0.536 | 0.000108528 |
| MAPK3 | mitogen-activated protein kinase 3 | 43.4258 | 61.3592 | 0.499 | 0.000108528 |
| CASP2 | Caspase 2 | 6.65398 | 9.33578 | 0.488 | 0.000679197 |
| ROCK1 | Rho associated coiled-coil containing protein kinase 1 | 15.711 | 20.961 | 0.416 | 0.000108528 |
| MAP3K5 | mitogen-activated protein kinase kinase kinase 5 | 4.264 | 5.599 | 0.393 | 0.0185603 |
| NFKBIA | NFKB inhibitor alpha | 30.3404 | 38.8837 | 0.358 | 0.00254092 |
| AIFM1 | apoptosis inducing factor mitochondria associated 1 | 22.697 | 28.454 | 0.3579 | 0.00342361 |
| GSK3B | glycogen synthase kinase 3 beta | 7.29297 | 9.04263 | 0.310 | 0.00350034 |
| PRKAG1 | protein kinase AMP-activated non-catalytic subunit gamma 1 | 22.7303 | 27.4994 | 0.275 | 0.0360551 |
| GSK3A | glycogen synthase kinase 3 alpha | 24.6326 | 29.3496 | 0.253 | 0.0231286 |
| Gene | Name | CTR | MOR+CBD | Log2-FoldChange | Q Value |
|---|---|---|---|---|---|
| MCL1 | MCL1, BCL2 family apoptosis regulator | 15.5899 | 13.3424 | −0.224 | 0.0365845 |
| PRKACA | protein kinase cAMP-activated catalytic subunit alpha | 36.0344 | 29.1389 | −0.306 | 0.000769756 |
| JUN | Jun proto-oncogene, AP-1 transcription factor subunit | 45.231 | 35.688 | −0.341 | 0.000108528 |
| BRAF | B-Raf proto-oncogene, serine/threonine kinase | 18.1853 | 14.3317 | −0.343 | 0.00374229 |
| DIABLO | diablo IAP-binding mitochondrial protein | 24.9695 | 19.2303 | −0.377 | 0.0018832 |
| APAF1 | apoptotic peptidase activating factor 1 | 5.49731 | 4.023 | −0.450 | 0.00146168 |
| NRAS | proto-oncogene, GTPase | 9.22853 | 6.45952 | −0.514 | 0.000209655 |
| PIK3CA | phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit alpha | 17.6961 | 12.3847 | −0.515 | 0.000108528 |
| PIK3CB | phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit beta | 3.9361 | 2.71759 | −0.534 | 0.00600177 |
| IKBKB | inhibitor of nuclear factor kappa B kinase subunit beta | 9.88765 | 6.66404 | −0.569 | 0.000108528 |
| PIK3CD | phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit delta | 11.4445 | 7.6164 | −0.587 | 0.000108528 |
| APP | amyloid beta precursor protein | 812.971 | 538.076 | −0.595 | 0.000108528 |
| NFKB1 | nuclear factor kappa B subunit 1 | 9.63897 | 6.14856 | −0.648 | 0.000108528 |
| RIPK1 | receptor interacting serine/threonine kinase 1 | 8.97176 | 5.47512 | −0.712 | 0.000108528 |
| JAK2 | Janus kinase 2 | 3.17143 | 1.92764 | −0.718 | 0.00382528 |
| ROCK2 | Rho associated coiled-coil containing protein kinase 2 | 25.266 | 15.313 | −0.722 | 0.000108528 |
| TRADD | TNFRSF1A associated via death domain | 6.42414 | 3.88442 | −0.726 | 0.0318132 |
| BAD | BCL2 associated agonist of cell death | 12.1113 | 7.0611 | −0.778 | 0.0138987 |
| STAT1 | signal transducer and activator of transcription 1 | 220.682 | 120.078 | −0.878 | 0.000108528 |
| STAT3 | signal transducer and activator of transcription 3 | 60.0522 | 32.1874 | −0.900 | 0.000108528 |
| CASP8 | caspase 8 | 14.8563 | 7.94494 | −0.903 | 0.000108528 |
| BIRC2 | baculoviral IAP repeat containing 2 | 23.7492 | 11.4038 | −1.058 | 0.000108528 |
| CHEK2 | checkpoint kinase 2 | 9.375 | 4.444 | −1.077 | 0.000209655 |
| PIDD1 | p53-induced death domain protein 1 | 2.819 | 1.274 | −1.145 | 0.00453657 |
| IKBKG | inhibitor of nuclear factor kappa B kinase subunit gamma | 20.3793 | 9.20785 | −1.146 | 0.000108528 |
| HTRA2 | HtrA serine peptidase 2 | 16.330 | 7.278 | −1.166 | 0.000108528 |
| BAX | BCL2 associated X, apoptosis regulator | 72.4174 | 31.9437 | −1.181 | 0.000108528 |
| ATM | ATM serine/threonine kinase | 17.969 | 7.731 | −1.216 | 0.000108528 |
| TP53 | tumor protein p53 | 21.8171 | 8.8547 | −1.301 | 0.000108528 |
| TNFRSF1A | TNF receptor superfamily member 1A | 130.525 | 52.1762 | −1.323 | 0.000108528 |
| CASP6 | Caspase 6 | 5.42611 | 2.11284 | −1.361 | 0.00318817 |
| CASP10 | Caspase 10 | 2.91088 | 0.99445 | −1.5495 | 0.00245968 |
| CASP9 | Caspase 9 | 3.91636 | 1.29947 | −1.591 | 0.000209655 |
| CFLAR | CASP8 and FADD like apoptosis regulator | 35.1508 | 11.6351 | −1.595 | 0.000108528 |
| PTEN | phosphatase and tensin homolog | 50.3299 | 15.3705 | −1.711 | 0.000108528 |
| CASP7 | Caspase 7 | 11.4138 | 2.89071 | −1.981 | 0.000108528 |
| Gene | Name | CTR | MOR+CBD | Log2-Fold Change | Q Value |
|---|---|---|---|---|---|
| TNFRSF10A | TNF receptor superfamily member 10a | 3.73692 | 10.4229 | 1.47983 | 0.000108528 |
| MAP2K1 | mitogen-activated protein kinase kinase 1 | 25.1462 | 50.4431 | 1.00432 | 0.000108528 |
| XIAP | X-linked inhibitor of apoptosis | 6.18581 | 10.1329 | 0.71202 | 0.000108528 |
| CYLD | CYLD lysine 63 deubiquitinase | 11.8515 | 17.1827 | 0.53589 | 0.000108528 |
| CHUK | conserved helix-loop-helix ubiquitous kinase | 11.3559 | 15.2974 | 0.42985 | 0.000494671 |
| ROCK1 | Rho associated coiled-coil containing protein kinase 1 | 15.7113 | 20.9609 | 0.4159 | 0.000108528 |
| RELA | RELA proto-oncogene, NF-kB subunit | 18.2587 | 24.1675 | 0.40449 | 0.00146168 |
| MAP3K5 | mitogen-activated protein kinase kinase kinase 5 | 4.26421 | 5.59967 | 0.39306 | 0.0185603 |
| NFKBIA | NFKB inhibitor alpha | 30.3404 | 38.8837 | 0.35793 | 0.00254092 |
| MPRIP | myosin phosphatase Rho interacting protein | 21.5258 | 18.5193 | −0.217 | 0.000108528 |
| PARP1 | poly(ADP-ribose) polymerase 1 | 45.8356 | 39.1245 | −0.2284 | 0.000494671 |
| ACIN1 | apoptotic chromatin condensation inducer 1 | 30.3285 | 25.0361 | −0.2767 | 0.000494671 |
| DIABLO | diablo IAP-binding mitochondrial protein | 24.9695 | 19.2303 | −0.3768 | 0.0018832 |
| APAF1 | apoptotic peptidase activating factor 1 | 5.49731 | 4.023 | −0.4505 | 0.00146168 |
| RPAIN | RPA interacting protein | 12.2724 | 8.85997 | −0.47 | 0.0253385 |
| FAS | Fas cell surface death receptor | 48.1152 | 33.0363 | −0.5424 | 0.000108528 |
| IKBKB | inhibitor of nuclear factor kappa B kinase subunit beta | 9.88765 | 6.66404 | −0.5692 | 0.000108528 |
| RIPK1 | receptor interacting serine/threonine kinase 1 | 8.97176 | 5.47512 | −0.7125 | 0.000108528 |
| TRADD | TNFRSF1A associated via death domain | 6.42414 | 3.88442 | −0.7258 | 0.0318132 |
| MAP4K4 | mitogen-activated protein kinase kinase kinase kinase 4 | 128.313 | 75.8702 | −0.7581 | 0.000108528 |
| CASP8 | caspase 8 | 14.8563 | 7.94494 | −0.903 | 0.000108528 |
| BIRC2 | baculoviral IAP repeat containing 2 | 23.7492 | 11.4038 | −1.0584 | 0.000108528 |
| IKBKG | inhibitor of nuclear factor kappa B kinase subunit gamma | 20.3793 | 9.20785 | −1.1462 | 0.000108528 |
| TNFRSF10B | TNF receptor superfamily member 10b | 94.0163 | 40.7967 | −1.2045 | 0.000108528 |
| LMNA | lamin A/C | 1373.63 | 587.557 | −1.2252 | 0.000108528 |
| TNFRSF1A | TNF receptor superfamily member 1A | 130.525 | 52.1762 | −1.3229 | 0.000108528 |
| CASP6 | Caspase 6 | 5.42611 | 2.11284 | −1.3607 | 0.00318817 |
| CASP10 | Caspase 10 | 2.91088 | 0.99445 | −1.5495 | 0.00245968 |
| CASP9 | Caspase 9 | 3.91636 | 1.29947 | −1.5916 | 0.000209655 |
| CFLAR | CASP8 and FADD like apoptosis regulator | 35.1508 | 11.6351 | −1.5951 | 0.000108528 |
| CASP7 | Caspase 7 | 11.4138 | 2.89071 | −1.9813 | 0.000108528 |
| Gene | Name | CTR | MOR+CBD | Log2-Fold Change | Q Value |
|---|---|---|---|---|---|
| SGK1 | serum/glucocorticoid regulated kinase 1 | 4.69263 | 106.74 | 4.50757 | 0.000108528 |
| CDC37 | cell division cycle 37 | 51.8182 | 194.202 | 1.90603 | 0.000108528 |
| TMX2 | thioredoxin related transmembrane protein 2 | 25.587 | 84.3633 | 1.72121 | 0.000108528 |
| BIRC5 | Baculoviral IAP repeat containing 5 | 0.876953 | 2.84342 | 1.69706 | 0.00508584 |
| RHEB | Ras homolog, mTORC1 binding | 15.1136 | 46.575 | 1.62371 | 0.000108528 |
| GK | glycerol kinase | 5.78668 | 13.9776 | 1.27231 | 0.000108528 |
| YWHAH | tyrosine3-monooxygenase/tryptophan 5-monooxygenase activation protein beta | 15.6913 | 35.0565 | 1.15972 | 0.000108528 |
| NCL | nucleolin | 178.099 | 377.828 | 1.08505 | 0.000108528 |
| UBE2D1 | ubiquitin conjugating enzyme E2 D1 | 4.46588 | 8.44974 | 0.919964 | 0.000108528 |
| AKT1S1 | AKT1 substrate 1 | 5.66925 | 9.57204 | 0.755668 | 0.00262175 |
| TSC1 | TSC complex subunit 1 | 7.62739 | 12.8277 | 0.75 | 0.000108528 |
| MAPK1 | mitogen-activated protein kinase 1 | 37.1635 | 61.9931 | 0.73822 | 0.000108528 |
| GRB2 | growth factor receptor bound protein 2 | 17.0713 | 28.1494 | 0.721528 | 0.000108528 |
| XIAP | X-linked inhibitor of apoptosis | 6.18581 | 10.1329 | 0.712017 | 0.000108528 |
| EIF4EBP1 | eukaryotic translation initiation factor 4E binding protein 1 | 14.858 | 24.2207 | 0.705002 | 0.00318817 |
| CSK | C-terminal Src kinase | 6.02989 | 9.54202 | 0.662163 | 0.00262175 |
| NDUFA6 | NADH:ubiquinone oxidoreductase subunit A6 | 44.3725 | 67.7367 | 0.610273 | 0.000108528 |
| VRK1 | vaccinia related kinase 1 | 11.1566 | 16.9088 | 0.599877 | 0.00229935 |
| IDH3A | isocitrate dehydrogenase 3 (NAD(+)) alpha | 6.79514 | 10.2507 | 0.593141 | 0.00245968 |
| RICTOR | RPTOR independent companion of MTOR complex 2 | 4.49855 | 6.77801 | 0.591403 | 0.000108528 |
| IDH3G | isocitrate dehydrogenase 3 (NAD(+)) gamma | 24.8055 | 36.6474 | 0.56305 | 0.000108528 |
| RPS7 | ribosomal protein S7 | 1220.66 | 1745.62 | 0.516083 | 0.000108528 |
| MAPK3 | mitogen-activated protein kinase 3 | 43.4258 | 61.3592 | 0.498727 | 0.000108528 |
| NAA10 | N(alpha)-acetyltransferase 10, NatA catalytic subunit | 65.211 | 90.1417 | 0.467079 | 0.000108528 |
| MAPKAP1 | mitogen-activated protein kinase associated protein 1 | 24.7763 | 34.1371 | 0.462382 | 0.000108528 |
| PPP1CA | protein phosphatase 1 catalytic subunit alpha | 69.6542 | 95.9494 | 0.462064 | 0.000108528 |
| UBC | ubiquitin C | 643.861 | 871.181 | 0.436224 | 0.000108528 |
| CHUK | conserved helix-loop-helix ubiquitous kinase | 11.3559 | 15.2974 | 0.429848 | 0.000494671 |
| RELA | RELA proto-oncogene, NF-kB subunit | 18.2587 | 24.1675 | 0.404486 | 0.00146168 |
| ATP1A1 | ATPase Na+/K+ transporting subunit alpha 1 | 107.718 | 138.718 | 0.364897 | 0.000108528 |
| MYL12B | myosin light chain 12B | 225.236 | 289.645 | 0.362849 | 0.000108528 |
| NFKBIA | NFKB inhibitor alpha | 30.3404 | 38.8837 | 0.357928 | 0.00254092 |
| GSK3B | glycogen synthase kinase 3 beta | 7.29297 | 9.04263 | 0.310236 | 0.00350034 |
| PRKAA1 | protein kinase AMP-activated catalytic subunit alpha 1 | 10.8385 | 12.9494 | 0.256722 | 0.000769756 |
| SDCBP | Syndecan binding protein | 98.2519 | 112.121 | 0.190497 | 0.000494671 |
| MTOR | mechanistic target of rapamycin kinase | 10.8825 | 12.325 | 0.179576 | 0.0270992 |
| CDKN1A | cyclin dependent kinase inhibitor 1A | 189.675 | 171.028 | −0.14929 | |
| YWHAB | tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein beta | 36.9941 | 31.2586 | −0.24304 | 0.00270215 |
| IRS1 | insulin receptor substrate 1 | 18.1379 | 15.1972 | −0.25521 | 0.000108528 |
| BRAF | B-Raf proto-oncogene, serine/threonine kinase | 18.1853 | 14.3317 | −0.34356 | 0.00374229 |
| ITGB1 | integrin subunit beta 1 | 659.512 | 498.137 | −0.40486 | 0.000108528 |
| SOS2 | SOS Ras/Rho guanine nucleotide exchange factor 2 | 7.55781 | 5.65783 | −0.41772 | 0.00294683 |
| APAF1 | apoptotic peptidase activating factor 1 | 5.49731 | 4.023 | −0.45045 | 0.00146168 |
| ULK1 | unc-51 like autophagy activating kinase 1 | 4.66539 | 3.33227 | −0.48549 | 0.0102621 |
| PIK3CA | phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit alpha | 17.6961 | 12.3847 | −0.5149 | 0.000108528 |
| NRAS | NRAS proto-oncogene, GTPase | 9.22853 | 6.45952 | −0.51467 | 0.000209655 |
| TSC2 | TSC complex subunit 2 | 37.2339 | 25.8014 | −0.52917 | 0.000108528 |
| RPS6KB1 | ribosomal protein S6 kinase B1 | 7.68103 | 5.28824 | −0.53851 | 0.000306871 |
| IKBKB | inhibitor of nuclear factor kappa B kinase subunit beta | 9.88765 | 6.66404 | −0.56923 | 0.000108528 |
| RPS6KA3 | ribosomal protein S6 kinase A3 | 12.1111 | 7.90192 | −0.61606 | 0.000108528 |
| PRDX4 | peroxiredoxin 4 | 141.666 | 91.3201 | −0.63349 | 0.000108528 |
| GRB10 | growth factor receptor bound protein 10 | 5.92727 | 3.55434 | −0.73779 | 0.000209655 |
| RHOD | ras homolog family member D | 8.51073 | 4.80959 | −0.82337 | 0.0175082 |
| CLIP1 | CAP-Gly domain containing linker protein 1 | 24.6133 | 13.7533 | −0.83966 | 0.000108528 |
| MAP2K7 | mitogen-activated protein kinase kinase 7 | 4.97253 | 2.44847 | −1.0221 | 0.000402551 |
| SOS1 | SOS Ras/Rac guanine nucleotide exchange factor | 26.454 | 12.3817 | −1.09527 | 0.000108528 |
| IKBKG | inhibitor of nuclear factor kappa B kinase subunit gamma | 20.3793 | 9.20785 | −1.14617 | 0.000108528 |
| APBB3 | amyloid beta precursor protein binding family B member 3 | 3.31305 | 1.35579 | −1.28903 | 0.00893961 |
| TP53 | tumor protein p53 | 21.8171 | 8.8547 | −1.30095 | 0.000108528 |
| NFIL3 | nuclear factor, interleukin 3 regulated | 7.77916 | 3.03723 | −1.35686 | 0.000108528 |
| PTEN | phosphatase and tensin homolog | 50.3299 | 15.3705 | −1.71126 | 0.000108528 |
| DDIT4 | DNA damage inducible transcript 4 | 18.464 | 3.98013 | −2.21383 | 0.000108528 |
| WNT5A | Wnt family member 5A | 69.3887 | 8.13031 | −3.09332 | 0.000108528 |
| Gene | Name | CTR | MOR+CBD | Log2-Fold Change | Q Value |
|---|---|---|---|---|---|
| BCL2A1 | BCL2 related protein A1 | 0.0001 | 2.73565 | 14.73959 | 0.000108528 |
| SGK1 | serum/glucocorticoid regulated kinase 1 | 4.69263 | 106.74 | 4.50757 | 0.000108528 |
| CDK1 | cyclin dependent kinase 1 | 1.60127 | 9.99442 | 2.64191 | 0.000108528 |
| CCND1 | cyclin D1 | 57.5334 | 289.455 | 2.33087 | 0.000108528 |
| BCL2L1 | BCL2 like 1 | 2.45308 | 9.41279 | 1.94003 | 0.000108528 |
| MYC | MYC proto-oncogene, bHLH transcription factor | 5.20354 | 19.3543 | 1.89509 | 0.000108528 |
| RHEB | Ras homolog, mTORC1 binding | 15.1136 | 46.575 | 1.62371 | 0.000108528 |
| BRCA1 | BRCA1, DNA repair associated | 1.39172 | 3.87502 | 1.47734 | 0.000108528 |
| GYS1 | glycogen synthase 1 | 20.1343 | 40.8951 | 1.02227 | 0.000108528 |
| CRTC2 | CREB regulated transcription coactivator 2 | 7.74262 | 14.2376 | 0.87881 | 0.000108528 |
| AKT1S1 | AKT1 substrate 1 | 5.66925 | 9.57204 | 0.755668 | 0.00262175 |
| TSC1 | TSC complex subunit 1 | 7.62739 | 12.8277 | 0.75 | 0.000108528 |
| MAPK1 | mitogen-activated protein kinase 1 | 37.1635 | 61.9931 | 0.73822 | 0.000108528 |
| EIF4EBP1 | eukaryotic translation initiation factor 4E binding protein 1 | 14.858 | 24.2207 | 0.705002 | 0.00318817 |
| RPS6KB2 | ribosomal protein S6 kinase B2 | 10.6792 | 16.799 | 0.65357 | 0.00129168 |
| CREB3 | cAMP responsive element binding protein 3 | 15.8771 | 24.3326 | 0.615947 | 0.000108528 |
| EIF4E | eukaryotic translation initiation factor 4E | 13.0091 | 17.7561 | 0.448797 | 0.000108528 |
| NFKBIA | NFKB inhibitor alpha | 30.3404 | 38.8837 | 0.357928 | 0.00254092 |
| GSK3B | glycogen synthase kinase 3 beta | 7.29297 | 9.04263 | 0.310236 | 0.00350034 |
| PRKAA1 | protein kinase AMP-activated catalytic subunit alpha 1 | 10.8385 | 12.9494 | 0.256722 | 0.0156024 |
| GSK3A | glycogen synthase kinase 3 alpha | 24.6326 | 29.3496 | 0.252774 | 0.0231286 |
| MTOR | mechanistic target of rapamycin kinase | 10.8825 | 12.325 | 0.179576 | 0.0270992 |
| RPS6 | ribosomal protein S6 | 759.592 | 800.781 | 0.0761832 | 0.0354641 |
| MCL1 | MCL1, BCL2 family apoptosis regulator | 15.5899 | 13.3424 | −0.224596 | 0.0365845 |
| BRAF | B-Raf proto-oncogene, serine/threonine kinase | 18.1853 | 14.3317 | −0.343559 | 0.00374229 |
| EIF4B | eukaryotic translation initiation factor 4B | 108.227 | 84.7943 | −0.352014 | 0.000108528 |
| PIK3CA | phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit alpha | 17.6961 | 12.3847 | −0.514879 | 0.000108528 |
| TSC2 | TSC complex subunit 2 | 37.2339 | 25.8014 | −0.529167 | 0.000108528 |
| PIK3CB | phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit beta | 3.9361 | 2.71759 | −0.534435 | 0.00600177 |
| RPS6KB1 | ribosomal protein S6 kinase B1 | 7.68103 | 5.28824 | −0.538514 | 0.000306871 |
| IKBKB | inhibitor of nuclear factor kappa B kinase subunit beta | 9.88765 | 6.66404 | −0.569232 | 0.000108528 |
| NFKB1 | nuclear factor kappa B subunit 1 | 9.63897 | 6.14856 | −0.64863 | 0.000108528 |
| RBL2 | RB transcriptional corepressor like 2 | 12.9876 | 7.44702 | −0.802397 | 0.000108528 |
| MAP2K7 | mitogen-activated protein kinase kinase 7 | 4.97253 | 2.44847 | −1.0221 | 0.000402551 |
| TP53 | tumor protein p53 | 21.8171 | 8.8547 | −1.30095 | 0.000108528 |
| PCK2 | phosphoenolpyruvate carboxykinase 2, mitochondrial | 10.9312 | 4.17877 | −1.38731 | 0.000108528 |
| CASP9 | Caspase 9 | 3.91636 | 1.29947 | −1.59159 | 0.000209655 |
| PTEN | phosphatase and tensin homolog | 50.3299 | 15.3705 | −1.71126 | 0.000108528 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lanza Cariccio, V.; Scionti, D.; Raffa, A.; Iori, R.; Pollastro, F.; Diomede, F.; Bramanti, P.; Trubiani, O.; Mazzon, E. Treatment of Periodontal Ligament Stem Cells with MOR and CBD Promotes Cell Survival and Neuronal Differentiation via the PI3K/Akt/mTOR Pathway. Int. J. Mol. Sci. 2018, 19, 2341. https://doi.org/10.3390/ijms19082341
Lanza Cariccio V, Scionti D, Raffa A, Iori R, Pollastro F, Diomede F, Bramanti P, Trubiani O, Mazzon E. Treatment of Periodontal Ligament Stem Cells with MOR and CBD Promotes Cell Survival and Neuronal Differentiation via the PI3K/Akt/mTOR Pathway. International Journal of Molecular Sciences. 2018; 19(8):2341. https://doi.org/10.3390/ijms19082341
Chicago/Turabian StyleLanza Cariccio, Veronica, Domenico Scionti, Antonio Raffa, Renato Iori, Federica Pollastro, Francesca Diomede, Placido Bramanti, Oriana Trubiani, and Emanuela Mazzon. 2018. "Treatment of Periodontal Ligament Stem Cells with MOR and CBD Promotes Cell Survival and Neuronal Differentiation via the PI3K/Akt/mTOR Pathway" International Journal of Molecular Sciences 19, no. 8: 2341. https://doi.org/10.3390/ijms19082341
APA StyleLanza Cariccio, V., Scionti, D., Raffa, A., Iori, R., Pollastro, F., Diomede, F., Bramanti, P., Trubiani, O., & Mazzon, E. (2018). Treatment of Periodontal Ligament Stem Cells with MOR and CBD Promotes Cell Survival and Neuronal Differentiation via the PI3K/Akt/mTOR Pathway. International Journal of Molecular Sciences, 19(8), 2341. https://doi.org/10.3390/ijms19082341