Amino Acid Metabolism and Transport Mechanisms as Potential Antifungal Targets

Abstract
1. Introduction
2. Yeasts
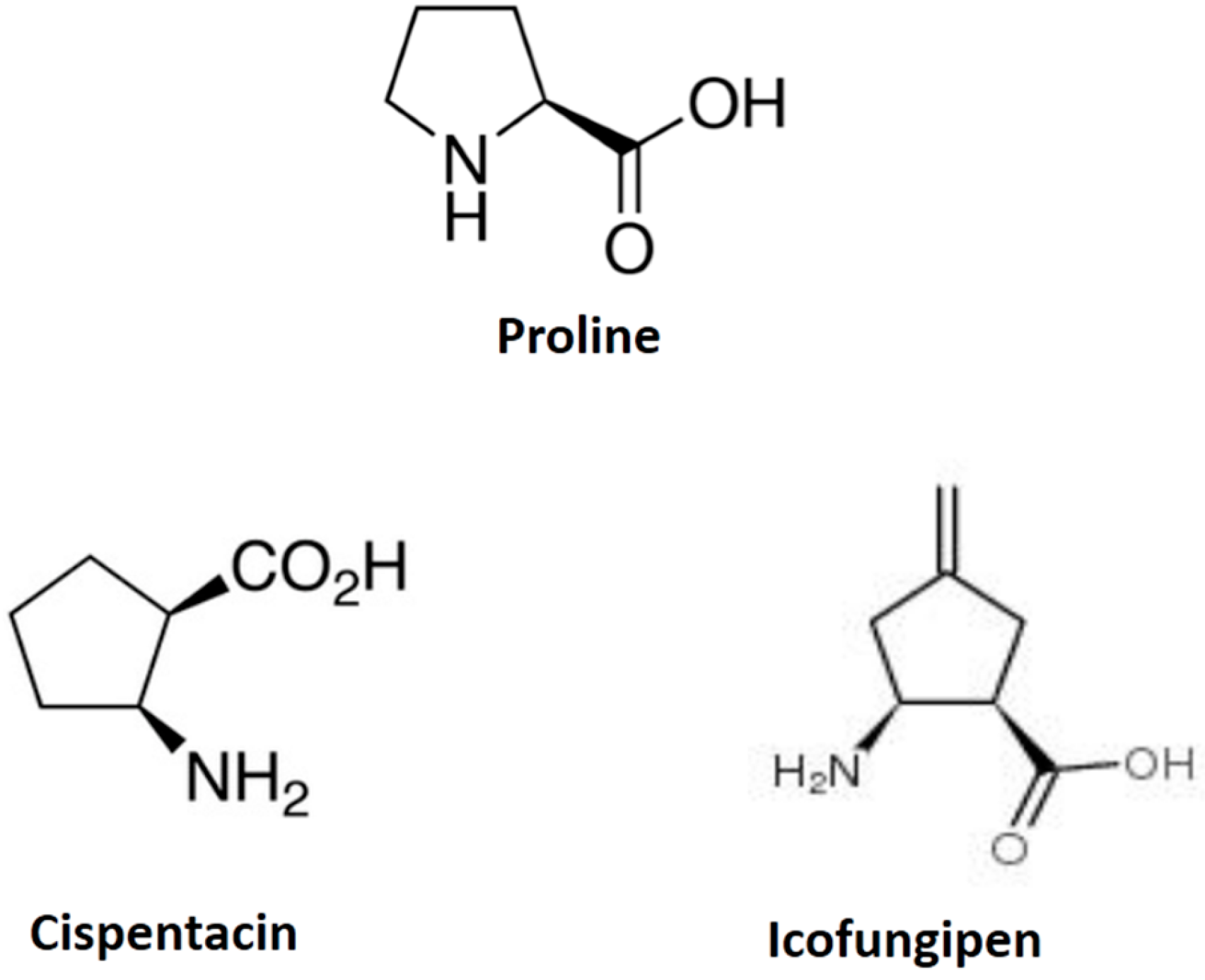
2.1. Icofungipen
2.2. Inositol Pathway
3. Molds
3.1. Histidine Pathway
3.2. Galactofuranose Pathway
4. Drug Discovery and Delivery
4.1. Drug Discovery
4.2. Drug Delivery
5. Conclusions and Future Directions
Author Contributions
Conflicts of Interest
References
- McCarthy, M.W.; Kontoyiannis, D.P.; Cornely, O.A.; Perfect, J.R.; Walsh, T.J. Novel Agents and Drug Targets to Meet the Challenges of Resistant Fungi. J. Infect. Dis. 2017, 216 (Suppl. S3), S474–S483. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, M.W.; Walsh, T.J. Drug development challenges and strategies to address emerging and resistant fungal pathogens. Expert Rev. Anti-Infect. Ther. 2017, 15, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.W.; Baker, S.E. Mycotoxins: A Fungal Genomics Perspective. Methods Mol. Biol. 2017, 1542, 367–379. [Google Scholar] [PubMed]
- Osherov, N.; Kontoyiannis, D.P. The anti-Aspergillus drug pipeline: Is the glass half full or empty? Med. Mycol. 2017, 55, 118–124. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Denning, D.W. Echinocandin antifungal drugs. Lancet 2003, 362, 1142–1151. [Google Scholar] [CrossRef]
- Ianiri, G.; Idnurm, A. Essential gene discovery in the basidiomycete Cryptococcus neoformans for antifungal drug target prioritization. mBio 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Kovanda, L.L.; Maher, R.; Hope, W.W. Isavuconazonium sulfate: A new agent for the treatment of invasive aspergillosis and invasive mucormycosis. Expert Rev. Clin. Pharmacol. 2016, 9, 887–897. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, M.; Rosengart, A.; Schuetz, A.N.; Kontoyiannis, D.P.; Walsh, T.J. Mold infections of the central nervous system. N. Engl. J. Med. 2014, 371, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Farmakiotis, D.; Kontoyiannis, D.P. Epidemiology of antifungal resistance in human pathogenic yeasts: Current viewpoint and practical recommendations for management. Int. J. Antimicrob. Agents 2017. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Lara, M.F.; Sifuentes-Osornio, J.; Ostrosky-Zeichner, L. Drugs in Clinical Development for Fungal Infections. Drugs 2017. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, M.W.; Denning, D.W.; Walsh, T.J. Future Research Priorities in Fungal Resistance. J. Infect. Dis. 2017, 216 (Suppl. S3), S484–S492. [Google Scholar] [CrossRef] [PubMed]
- Nishida, I.; Watanabe, D.; Tsolmonbaatar, A.; Kaino, T.; Ohtsu, I.; Takagi, H. Vacuolar amino acid transporters upregulated by exogenous proline and involved in cellular localization of proline in Saccharomyces cerevisiae. J. Gen. Appl. Microbiol. 2016, 62, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Uemura, S.; Mochizuki, T.; Kurosaka, G.; Hashimoto, T.; Masukawa, Y.; Abe, F. Functional analysis of human aromatic amino acid transporter MCT10/TAT1 using the yeast Saccharomyces cerevisiae. Biochim. Biophys. Acta 2017, 1859, 2076–2085. [Google Scholar] [CrossRef] [PubMed]
- Llinares, E.; Barry, A.O.; André, B. The AP-3 adaptor complex mediates sorting of yeast and mammalian PQ-loop-family basic amino acid transporters to the vacuolar/lysosomal membrane. Sci. Rep. 2015, 5, 16665. [Google Scholar] [CrossRef] [PubMed]
- Wipf, D.; Ludewig, U.; Tegeder, M.; Rentsch, D.; Koch, W.; Frommer, W.B. Conservation of amino acid transporters in fungi, plants and animals. Trends Biochem. Sci. 2002, 27, 139–147. [Google Scholar] [CrossRef]
- Tanaka, N.; Mukai, Y. Yeast Cyc8p and Tup1p proteins function as coactivators for transcription of Stp1/2p-dependent amino acid transporter genes. Biochem. Biophys. Res. Commun. 2015, 468, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Hou, Y.; Yue, L.; Liu, S.; Du, J.; Sun, S. Potential Targets for Antifungal Drug Discovery Based on Growth and Virulence in Candida albicans. Antimicrob. Agents Chemother. 2015, 59, 5885–5891. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Ma, Y.; Zhou, X.; Furuyashiki, T. Constitutive Tor2 Activity Promotes Retention of the Amino Acid Transporter Agp3 at Trans-Golgi/Endosomes in Fission Yeast. PLoS ONE 2015, 10, e0139045. [Google Scholar] [CrossRef] [PubMed]
- Nonn, M.; Kiss, L.; Hänninen, M.M.; Sillanpää, R.; Fülöp, F. Synthesis of highly functionalized fluorinated cispentacin derivatives. Chem. Biodivers. 2012, 9, 2571–2581. [Google Scholar] [CrossRef] [PubMed]
- Kiss, L.; Cherepanova, M.; Forró, E.; Fülöp, F. A new access route to functionalized cispentacins from norbornene β-amino acids. Chemistry 2013, 19, 2102–2107. [Google Scholar] [CrossRef] [PubMed]
- Konishi, M.; Nishio, M.; Saitoh, K.; Miyaki, T.; Oki, T.; Kawaguchi, H. Cispentacin, a new antifungal antibiotic. I. Production, isolation, physico-chemical properties and structure. J. Antibiot. 1989, 42, 1749–1755. [Google Scholar] [CrossRef] [PubMed]
- Ziegelbauer, K.; Babczinski, P.; Schönfeld, W. Molecular mode of action of the antifungal beta-amino acid BAY 10-8888. Antimicrob. Agents Chemother. 1998, 42, 2197–2205. [Google Scholar] [PubMed]
- Aggarwal, V.K.; Roseblade, S.J.; Barrell, J.K.; Alexander, R. Highly diastereoselective nitrone cycloaddition onto a chiral ketene equivalent: Asymmetric synthesis of cispentacin. Org. Lett. 2002, 4, 1227–1229. [Google Scholar] [CrossRef] [PubMed]
- Langer, O.; Kählig, H.; Zierler-Gould, K.; Bats, J.W.; Mulzer, J. A bicyclic cispentacin derivative as a novel reverse turn inducer in a GnRH mimetic. J. Org. Chem. 2002, 67, 6878–6883. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, V.K.; Roseblade, S.; Alexander, R. The use of enantiomerically pure ketene dithioacetal bis(sulfoxides) in highly diastereoselective intramolecular nitrone cycloadditions. Application in the total synthesis of the beta-amino acid (−)-cispentacin and the first asymmetric synthesis of cis-(3R,4R)-4-amino-pyrrolidine-3-carboxylic acid. Org. Biomol. Chem. 2003, 1, 684–691. [Google Scholar] [PubMed]
- Mierke, D.F.; Nössner, G.; Schiller, P.W.; Goodman, M. Morphiceptin analogs containing 2-aminocyclopentane carboxylic acid as a peptidomimetic for proline. Int. J. Pept. Protein Res. 1990, 35, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Capobianco, J.O.; Zakula, D.; Coen, M.L.; Goldman, R.C. Anti-Candida activity of cispentacin: The active transport by amino acid permeases and possible mechanisms of action. Biochem. Biophys. Res. Commun. 1993, 190, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Zanol, M.; Hermann, R.; Bernareggi, A.; Borgonovi, M.; Taglietti, E.; Zerilli, L.F. HPLC method for the quantitation of cispentacin enantiomers in rat urine. Boll. Chim. Farm. 1995, 134, 390–393. [Google Scholar] [PubMed]
- Jethwaney, D.; Höfer, M.; Khaware, R.K.; Prasad, R. Functional reconstitution of a purified proline permease from Candida albicans: Interaction with the antifungal cispentacin. Microbiology 1997, 143, 397–404. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Oki, T.; Hirano, M.; Tomatsu, K.; Numata, K.; Kamei, H. Cispentacin, a new antifungal antibiotic. II. In vitro and in vivo antifungal activities. J. Antibiot. 1989, 42, 1756–1762. [Google Scholar] [CrossRef] [PubMed]
- Mazu, T.K.; Bricker, B.A.; Flores-Rozas, H.; Ablordeppey, S.Y. The Mechanistic Targets of Antifungal Agents: An Overview. Mini Rev. Med. Chem. 2016, 16, 555–578. [Google Scholar] [CrossRef] [PubMed]
- Petraitiene, R.; Petraitis, V.; Kelaher, A.M.; Sarafandi, A.A.; Mickiene, D.; Groll, A.H.; Sein, T.; Bacher, J.; Walsh, T.J. Efficacy, plasma pharmacokinetics, and safety of icofungipen, an inhibitor of Candida isoleucyl-tRNA synthetase, in treatment of experimental disseminated candidiasis in persistently neutropenic rabbits. Antimicrob. Agents Chemother. 2005, 49, 2084–2092. [Google Scholar] [CrossRef] [PubMed]
- Hasenoehrl, A.; Galic, T.; Ergovic, G.; Marsic, N.; Skerlev, M.; Mittendorf, J.; Geschke, U.; Schmidt, A.; Schoenfeld, W. In vitro activity and in vivo efficacy of icofungipen (PLD-118), a novel oral antifungal agent, against the pathogenic yeast Candida albicans. Antimicrob. Agents Chemother. 2006, 50, 3011–3018. [Google Scholar] [CrossRef] [PubMed]
- Petraitis, V.; Petraitiene, R.; Kelaher, A.M.; Sarafandi, A.A.; Sein, T.; Mickiene, D.; Bacher, J.; Groll, A.H.; Walsh, T.J. Efficacy of PLD-118, a novel inhibitor of candida isoleucyl-tRNA synthetase, against experimental oropharyngeal and esophageal candidiasis caused by fluconazole-resistant C. albicans. Antimicrob. Agents Chemother. 2004, 48, 3959–3967. [Google Scholar] [CrossRef] [PubMed]
- Ziegelbauer, K. Decreased accumulation or increased isoleucyl-tRNA synthetase activity confers resistance to the cyclic beta-amino acid BAY 10-8888 in Candida albicans and Candida tropicalis. Antimicrob. Agents Chemother. 1998, 42, 1581–1586. [Google Scholar] [PubMed]
- Mittendorf, J.; Kunisch, F.; Matzke, M.; Militzer, H.C.; Schmidt, A.; Schönfeld, W. Novel antifungal beta-amino acids: Synthesis and activity against Candida albicans. Bioorg. Med. Chem. Lett. 2003, 13, 433–436. [Google Scholar] [CrossRef]
- Kiss, L.; Forró, E.; Orsy, G.; Ábrahámi, R.; Fülöp, F. Stereo- and Regiocontrolled Syntheses of Exomethylenic Cyclohexane β-Amino Acid Derivatives. Molecules 2015, 20, 21094–21102. [Google Scholar] [CrossRef] [PubMed]
- Parnham, M.J.; Bogaards, J.J.; Schrander, F.; Schut, M.W.; Oresković, K.; Mildner, B. The novel antifungal agent PLD-118 is neither metabolized by liver microsomes nor inhibits cytochrome P450 in vitro. Biopharm. Drug Dispos. 2005, 26, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Stanek, J.; Caravatti, G.; Frei, J.; Furet, P.; Mett, H.; Schneider, P.; Regenass, U. 4-Amidinoindan-1-one 2′-amidinohydrazone: A new potent and selective inhibitor of S-adenosylmethionine decarboxylase. J. Med. Chem. 1993, 36, 2168–2171. [Google Scholar] [CrossRef] [PubMed]
- Debono, M.; Gordee, R.S. Antibiotics that inhibit fungal cell wall development. Annu. Rev. Microbiol. 1994, 48, 471–497. [Google Scholar] [CrossRef] [PubMed]
- Merali, S.; Vargas, D.; Franklin, M.; Clarkson, A.B. S-adenosylmethionine and Pneumocystis carinii. J. Biol. Chem. 2000, 275, 14958–14963. [Google Scholar] [CrossRef] [PubMed]
- Nevez, G.; Totet, A.; Pautard, J.C.; Raccurt, C. Pneumocystis carinii detection using nested-PCR in nasopharyngeal aspirates of immunocompetent infants with bronchiolitis. J. Eukaryot. Microbiol. 2001, 122S–123S. [Google Scholar] [CrossRef]
- Dworkin, M.S.; Williamson, J.; Jones, J.L.; Kaplan, J.E. Prophylaxis with trimethoprim-sulfamethoxazole for human immunodeficiency virus-infected patients: Impact on risk for infectious diseases. Clin. Infect. Dis. 2001, 33, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Kaneshiro, E.S.; Rosenfeld, J.A.; Basselin-Eiweida, M.; Stringer, J.R.; Keely, S.P.; Smulian, A.G.; Giner, J.L. The Pneumocystis carinii drug target S-adenosyl-l-methionine:sterol C-24 methyl transferase has a unique substrate preference. Mol. Microbiol. 2002, 44, 989–999. [Google Scholar] [CrossRef] [PubMed]
- Porollo, A.; Sesterhenn, T.M.; Collins, M.S.; Welge, J.A.; Cushion, M.T. Comparative genomics of pneumocystis species suggests the absence of genes for myo-inositol synthesis and reliance on inositol transport and metabolism. mBio 2014, 5, e01834. [Google Scholar] [CrossRef] [PubMed]
- Xue, C. Finding the sweet spot: How human fungal pathogens acquire and turn the sugar inositol against their hosts. mBio 2015, 6, e00109. [Google Scholar] [CrossRef] [PubMed]
- Cushion, M.T.; Collins, M.S.; Sesterhenn, T.; Porollo, A.; Vadukoot, A.K.; Merino, E.J. Functional Characterization of Pneumocystis carinii Inositol Transporter 1. mBio 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Arthurs, B.; Wunderle, K.; Hsu, M.; Kim, S. Invasive aspergillosis related to ibrutinib therapy for chronic lymphocytic leukemia. Respir. Med. Case Rep. 2017, 21, 27–29. [Google Scholar] [CrossRef] [PubMed]
- Baron, M.; Zini, J.M.; Challan Belval, T.; Vignon, M.; Denis, B.; Alanio, A.; Malphettes, M. Fungal infections in patients treated with ibrutinib: Two unusual cases of invasive aspergillosis and cryptococcal meningoencephalitis. Leuk. Lymphoma 2017. [Google Scholar] [CrossRef] [PubMed]
- Jaumally, B.A.; Salem, A.M.; Sagar, A.E.; Pyrgos, G. Invasive pulmonary aspergillosis in a steroid-dependent asthmatic. Respir. Med. Case Rep. 2017, 20, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Dietl, A.M.; Amich, J.; Leal, S.; Beckmann, N.; Binder, U.; Beilhack, A.; Pearlman, E.; Haas, H. Histidine biosynthesis plays a crucial role in metal homeostasis and virulence of Aspergillus fumigatus. Virulence 2016, 7, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Wan, P.J.; Tang, Y.H.; Yuan, S.Y.; Wang, W.X.; Lai, F.X.; Yu, X.P.; Fu, Q. ATP phosphoribosyltransferase from symbiont Entomomyces delphacidicola invovled in histidine biosynthesis of Nilaparvata lugens (Stål). Amino Acids 2016, 48, 2605–2617. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Wang, S. Histidine Regulates Seed Oil Deposition through Abscisic Acid Biosynthesis and β-Oxidation. Plant Physiol. 2016, 172, 848–857. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, L.L.; Toledo, M.S.; Ferreira, F.A.; Straus, A.H.; Takahashi, H.K. Structural diversity and biological significance of glycosphingolipids in pathogenic and opportunistic fungi. Front. Cell. Infect. Microbiol. 2014, 4, 138. [Google Scholar] [CrossRef] [PubMed]
- Misra, S.; Gupta, J.; Misra-Bhattacharya, S. RNA interference mediated knockdown of Brugia malayi UDP-Galactopyranose mutase severely affects parasite viability, embryogenesis and in vivo development of infective larvae. Parasites Vectors 2017, 10, 34. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhu, Q.; Chen, Q.; Song, Y.; Huang, H.; Li, J.; Ma, J.; Li, Q.; Ju, J. Deciphering the sugar biosynthetic pathway and tailoring steps of nucleoside antibiotic A201A unveils a GDP-l-galactose mutase. Proc. Natl. Acad. Sci. USA 2017, 114, 4948–4953. [Google Scholar] [CrossRef] [PubMed]
- Li, L.X.; Rautengarten, C.; Heazlewood, J.L.; Doering, T.L. Xylose donor transport is critical for fungal virulence. PLoS Pathog. 2018, 14, e1006765. [Google Scholar] [CrossRef] [PubMed]
- Oka, T. Biosynthesis of galactomannans found in filamentous fungi belonging to Pezizomycotina. Biosci. Biotechnol. Biochem. 2018, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Geno, K.A.; Bush, C.A.; Wang, M.; Jin, C.; Nahm, M.H.; Yang, J. WciG O-Acetyltransferase Functionality Differentiates Pneumococcal Serotypes 35C and 42. J. Clin. Microbiol. 2017, 55, 2775–2784. [Google Scholar] [CrossRef] [PubMed]
- Katafuchi, Y.; Li, Q.; Tanaka, Y.; Shinozuka, S.; Kawamitsu, Y.; Izumi, M.; Ekino, K.; Mizuki, K.; Takegawa, K.; Shibata, N.; et al. GfsA is a β1,5-galactofuranosyltransferase involved in the biosynthesis of the galactofuran side chain of fungal-type galactomannan in Aspergillus fumigatus. Glycobiology 2017, 27, 568–581. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Salas, O.; Bowler, K.; Bar-Peled, M.; Sharon, A. UDP-4-Keto-6-Deoxyglucose, a Transient Antifungal Metabolite, Weakens the Fungal Cell Wall Partly by Inhibition of UDP-Galactopyranose Mutase. mBio 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, E.; Higuchi, Y.; Mori, K.; Yairo, N.; Toyota, S.; Oka, T.; Tashiro, K.; Takegawa, K. Characterization of a PA14 domain-containing galactofuranose-specific β-d-galactofuranosidase from Streptomyces sp. Biosci. Biotechnol. Biochem. 2017, 81, 1314–1319. [Google Scholar] [CrossRef] [PubMed]
- Afroz, S.; El-Ganiny, A.M.; Sanders, D.A.; Kaminskyj, S.G. Roles of the Aspergillus nidulans UDP-galactofuranose transporter, UgtA in hyphal morphogenesis, cell wall architecture, conidiation, and drug sensitivity. Fungal Genet. Biol. 2011, 48, 896–903. [Google Scholar] [CrossRef] [PubMed]
- Marino, C.; Rinflerch, A.; de Lederkremer, R.M. Galactofuranose antigens, a target for diagnosis of fungal infections in humans. Future Sci. OA 2017, 3, FSO199. [Google Scholar] [CrossRef] [PubMed]
- Stanek, J.; Caravatti, G.; Capraro, H.G.; Furet, P.; Mett, H.; Schneider, P.; Regenass, U. S-adenosylmethionine decarboxylase inhibitors: New aryl and heteroaryl analogues of methylglyoxal bis(guanylhydrazone). J. Med. Chem. 1993, 36, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Skelly, M.J.; Holzman, R.S.; Merali, S. S-adenosylmethionine levels in the diagnosis of Pneumocystis carinii pneumonia in patients with HIV infection. Clin. Infect. Dis. 2008, 46, 467–471. [Google Scholar] [CrossRef] [PubMed]
- Skelly, M.; Hoffman, J.; Fabbri, M.; Holzman, R.S.; Clarkson, A.B.; Merali, S. S-adenosylmethionine concentrations in diagnosis of Pneumocystis carinii pneumonia. Lancet 2003, 361, 1267–1268. [Google Scholar] [CrossRef]
- Shivji, M.; Burger, S.; Moncada, C.A.; Clarkson, A.B.; Merali, S. Effect of nicotine on lung S-adenosylmethionine and development of Pneumocystis pneumonia. J. Biol. Chem. 2005, 280, 15219–15228. [Google Scholar] [CrossRef] [PubMed]
- Merali, S.; Clarkson, A.B. S-adenosylmethionine and Pneumocystis. FEMS Microbiol. Lett. 2004, 237, 179–186. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Perez-Leal, O.; Moncada, C.; Clarkson, A.B.; Merali, S. Pneumocystis S-adenosylmethionine transport: A potential drug target. Am. J. Respir. Cell Mol. Biol. 2011, 45, 1142–1146. [Google Scholar] [CrossRef] [PubMed]
- Helweg-Larsen, J. S-adenosylmethionine in plasma to test for Pneumocystis carinii pneumonia. Lancet 2003, 361, 1237. [Google Scholar] [CrossRef]
- McCarthy, M.W.; Walsh, T.J. Drugs currently under investigation for the treatment of invasive candidiasis. Expert Opin. Investig. Drugs 2017, 26, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, A.A.; Sharma, M.; Bachhawat, A.K. Insights into the molecular basis for substrate binding and specificity of the fungal cystine transporter CgCYN1. Biochim. Biophys. Acta 2017, 1859, 2259–2268. [Google Scholar] [CrossRef] [PubMed]
- Popov-Čeleketić, D.; Bianchi, F.; Ruiz, S.J.; Meutiawati, F.; Poolman, B. A Plasma Membrane Association Module in Yeast Amino Acid Transporters. J. Biol. Chem. 2016, 291, 16024–16037. [Google Scholar] [CrossRef] [PubMed]
- Dutta, D.; Fliegel, L. Structure and function of yeast and fungal Na+/H+ antiporters. IUBMB Life 2018, 70, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Gao, Y.; Meng, F.; Yang, C.; Fu, J.; Li, Y. The Sphingosine 1-Phosphate Analogue FTY720 Alleviates Seizure-induced Over-Expression of P-Glycoprotein in Rat Hippocampus. Basic Clin. Pharmacol. Toxicol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Foster, A.D.; Vicente, D.; Sexton, J.J.; Johnston, L.; Clark, N.; Leonhardt, C.; Elster, E.A.; Davis, T.A.; Bradley, M.J. Administration of FTY720 during Tourniquet-Induced Limb Ischemia Reperfusion Injury Attenuates Systemic Inflammation. Mediat. Inflamm. 2017, 2017, 4594035. [Google Scholar] [CrossRef] [PubMed]
- Welsch, C.A.; Roth, L.W.; Goetschy, J.F.; Movva, N.R. Genetic, biochemical, and transcriptional responses of Saccharomyces cerevisiae to the novel immunomodulator FTY720 largely mimic those of the natural sphingolipid phytosphingosine. J. Biol. Chem. 2004, 279, 36720–36731. [Google Scholar] [CrossRef] [PubMed]
- Barthelemy, C.; Barry, A.O.; Twyffels, L.; André, B. FTY720-induced endocytosis of yeast and human amino acid transporters is preceded by reduction of their inherent activity and TORC1 inhibition. Sci. Rep. 2017, 7, 13816. [Google Scholar] [CrossRef] [PubMed]
- Hagihara, K.; Kinoshita, K.; Ishida, K.; Hojo, S.; Kameoka, Y.; Satoh, R.; Takasaki, T.; Sugiura, R. A genome-wide screen for FTY720-sensitive mutants reveals genes required for ROS homeostasis. Microb. Cell 2017, 4, 390–401. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, F.; Pan, Y.; Lin, L.; Xiong, X. Effects of FTY720 on Lung Injury Induced by Hindlimb Ischemia Reperfusion in Rats. Mediat. Inflamm. 2017, 2017, 5301312. [Google Scholar] [CrossRef] [PubMed]
- Hagihara, K.; Kita, A.; Mizukura, A.; Yao, M.; Kitai, Y.; Kunoh, T.; Masuko, T.; Matzno, S.; Chiba, K.; Sugiura, R. Fingolimod (FTY720) stimulates Ca2+/calcineurin signaling in fission yeast. PLoS ONE 2013, 8, e81907. [Google Scholar] [CrossRef] [PubMed]
- Gournas, C.; Prévost, M.; Krammer, E.M.; André, B. Function and Regulation of Fungal Amino Acid Transporters: Insights from Predicted Structure. Adv. Exp. Med. Biol. 2016, 892, 69–106. [Google Scholar] [PubMed]
- Park, J.; Tefsen, B.; Heemskerk, M.J.; Lagendijk, E.L.; van den Hondel, C.A.; van Die, I.; Ram, A.F. Identification and functional analysis of two Golgi-localized UDP-galactofuranose transporters with overlapping functions in Aspergillus niger. BMC Microbiol. 2015, 15, 253. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.K.; van Straaten, K.E.; Sanders, D.A.; Kaminskyj, S.G. Aspergillus nidulans cell wall composition and function change in response to hosting several Aspergillus fumigatus UDP-galactopyranose mutase activity mutants. PLoS ONE 2014, 9, e85735. [Google Scholar] [CrossRef] [PubMed]
- Perfect, J.R. The antifungal pipeline: A reality check. Nat. Rev. Drug Discov. 2017. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Amino Acid Transporter | Antifungal Agent | Organism | Structures |
|---|---|---|---|
| Proline | cispentacin (1R,2S)-2-aminocyclopentane-1-carboxylic acid | Candida albicans Cryptococcus neoformans | Figure 1 |
| Proline | icofungipen (1R,2S)-2-amino-4-methylidene-cyclopentane | Candida albicans Cryptococcus neoformans | Figure 1 |
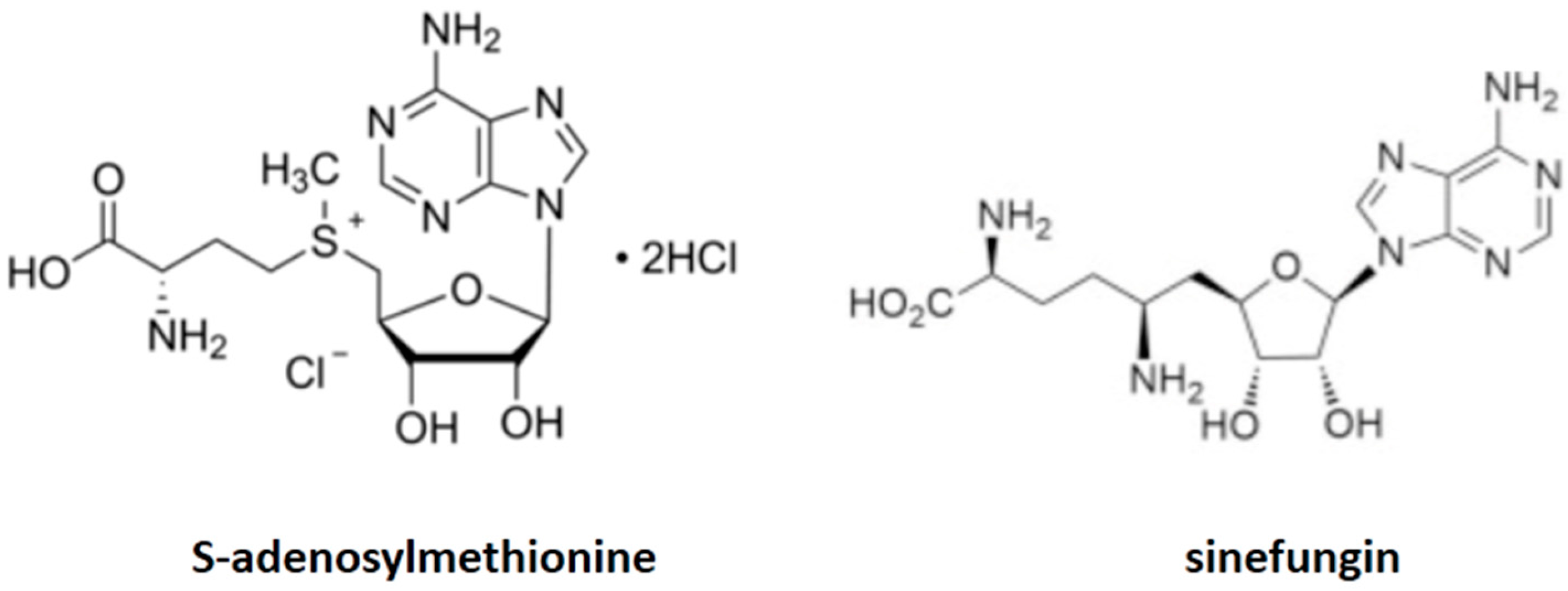
| Mitochondrial S-adenosylmethionine transporter | Sinefungin (2S,5S)-2,5-diamino-6-[(2R,3S,4R,5R)-5-(6-aminopurin-9-yl)-3,4-dihydroxyoxolan-2-yl]hexanoic acid | Pneumocystis Saccharomyces cerevisiae | Figure 2 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McCarthy, M.W.; Walsh, T.J. Amino Acid Metabolism and Transport Mechanisms as Potential Antifungal Targets. Int. J. Mol. Sci. 2018, 19, 909. https://doi.org/10.3390/ijms19030909
McCarthy MW, Walsh TJ. Amino Acid Metabolism and Transport Mechanisms as Potential Antifungal Targets. International Journal of Molecular Sciences. 2018; 19(3):909. https://doi.org/10.3390/ijms19030909
Chicago/Turabian StyleMcCarthy, Matthew W., and Thomas J. Walsh. 2018. "Amino Acid Metabolism and Transport Mechanisms as Potential Antifungal Targets" International Journal of Molecular Sciences 19, no. 3: 909. https://doi.org/10.3390/ijms19030909
APA StyleMcCarthy, M. W., & Walsh, T. J. (2018). Amino Acid Metabolism and Transport Mechanisms as Potential Antifungal Targets. International Journal of Molecular Sciences, 19(3), 909. https://doi.org/10.3390/ijms19030909