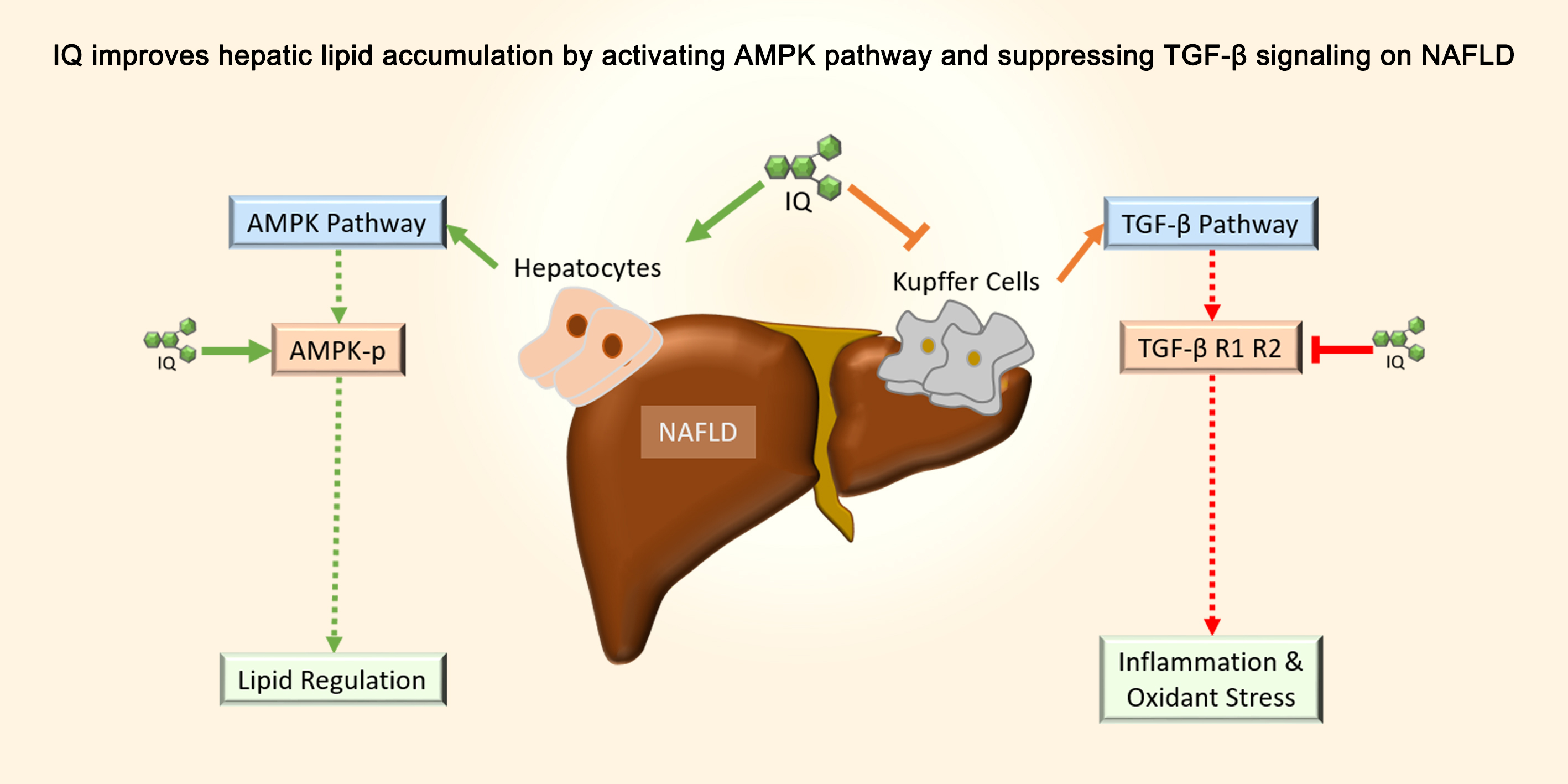
Isoquercetin Improves Hepatic Lipid Accumulation by Activating AMPK Pathway and Suppressing TGF-β Signaling on an HFD-Induced Nonalcoholic Fatty Liver Disease Rat Model

Abstract
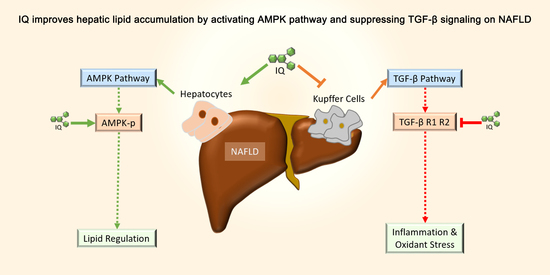
1. Introduction
2. Results
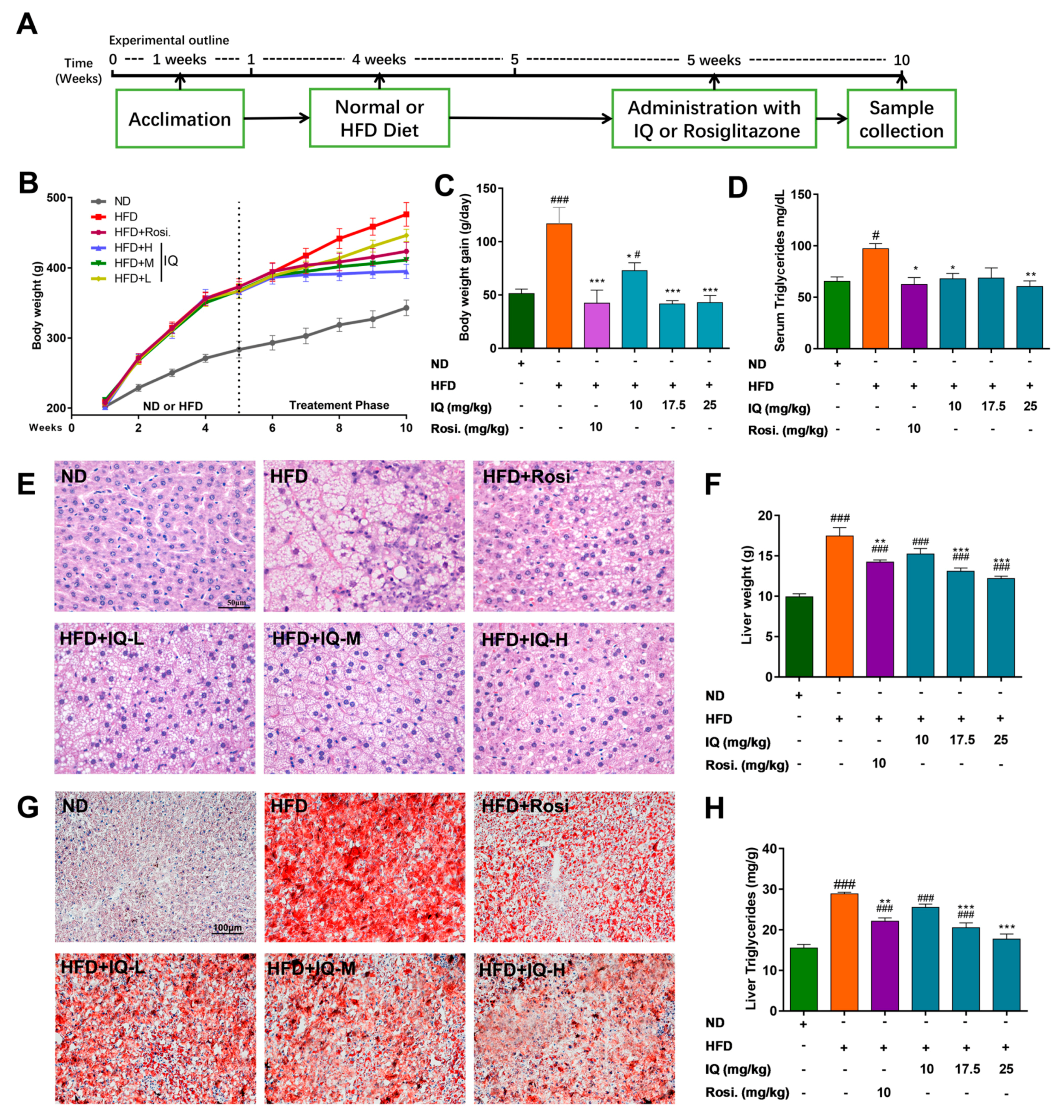
2.1. IQ Improves Lipid Accumulation on HFD Induced NAFLD Rats
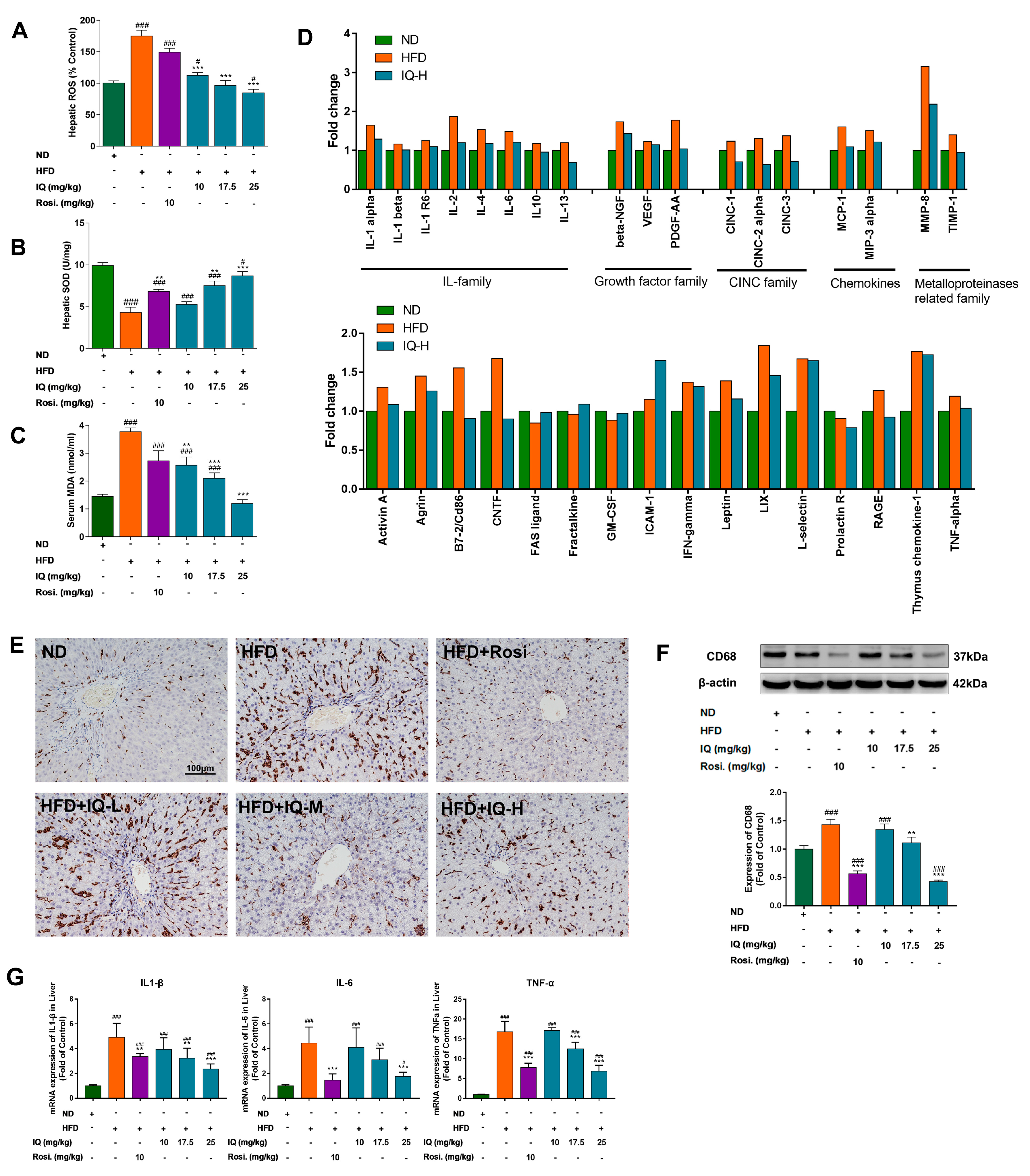
2.2. Effects of IQ on Oxidative Stress, Serum Cytokine Array, Inflammation and Kupffer Cell Markers
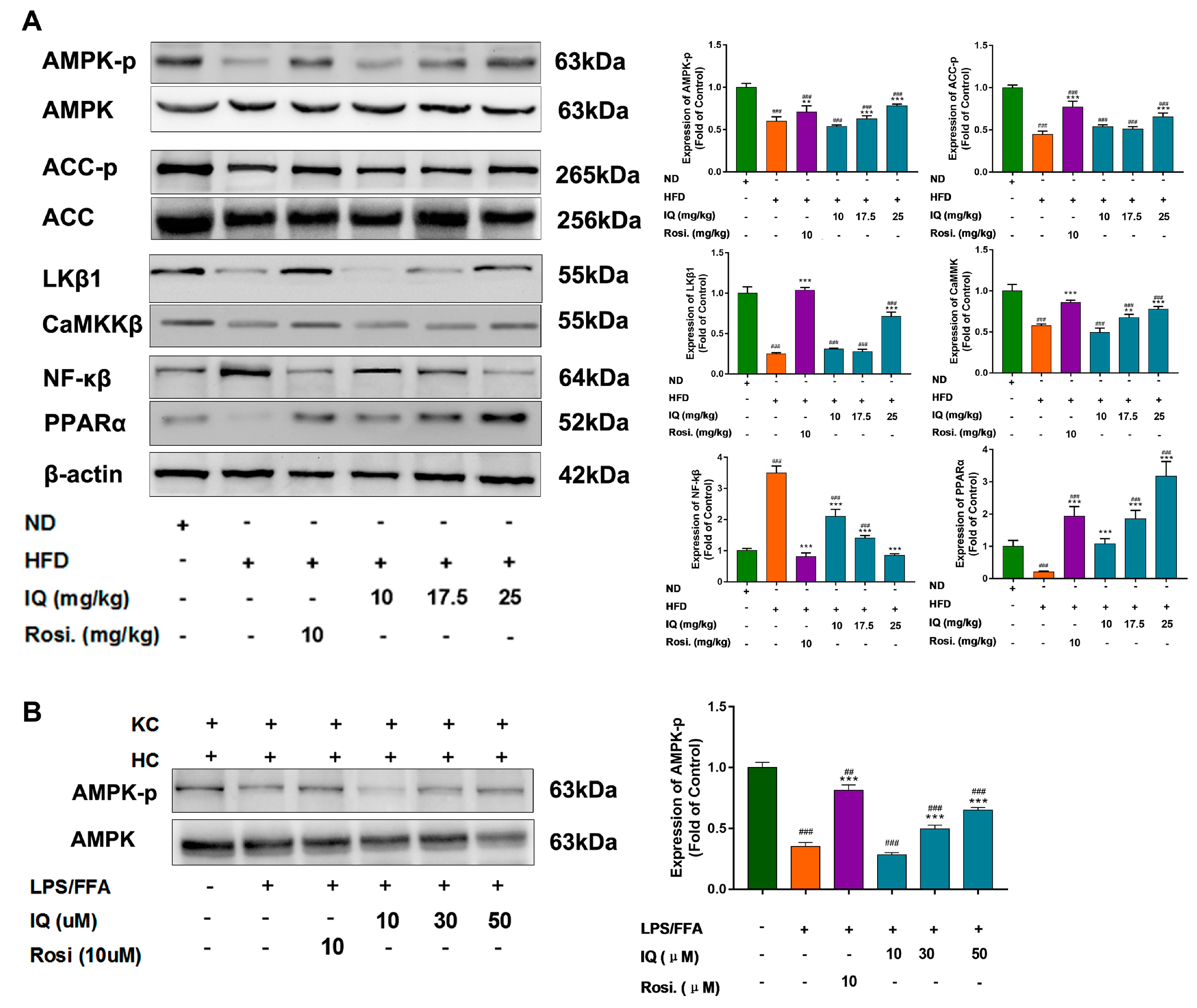
2.3. IQ Enhances the AMPK Phosphorylation via LKβ1 Upstream
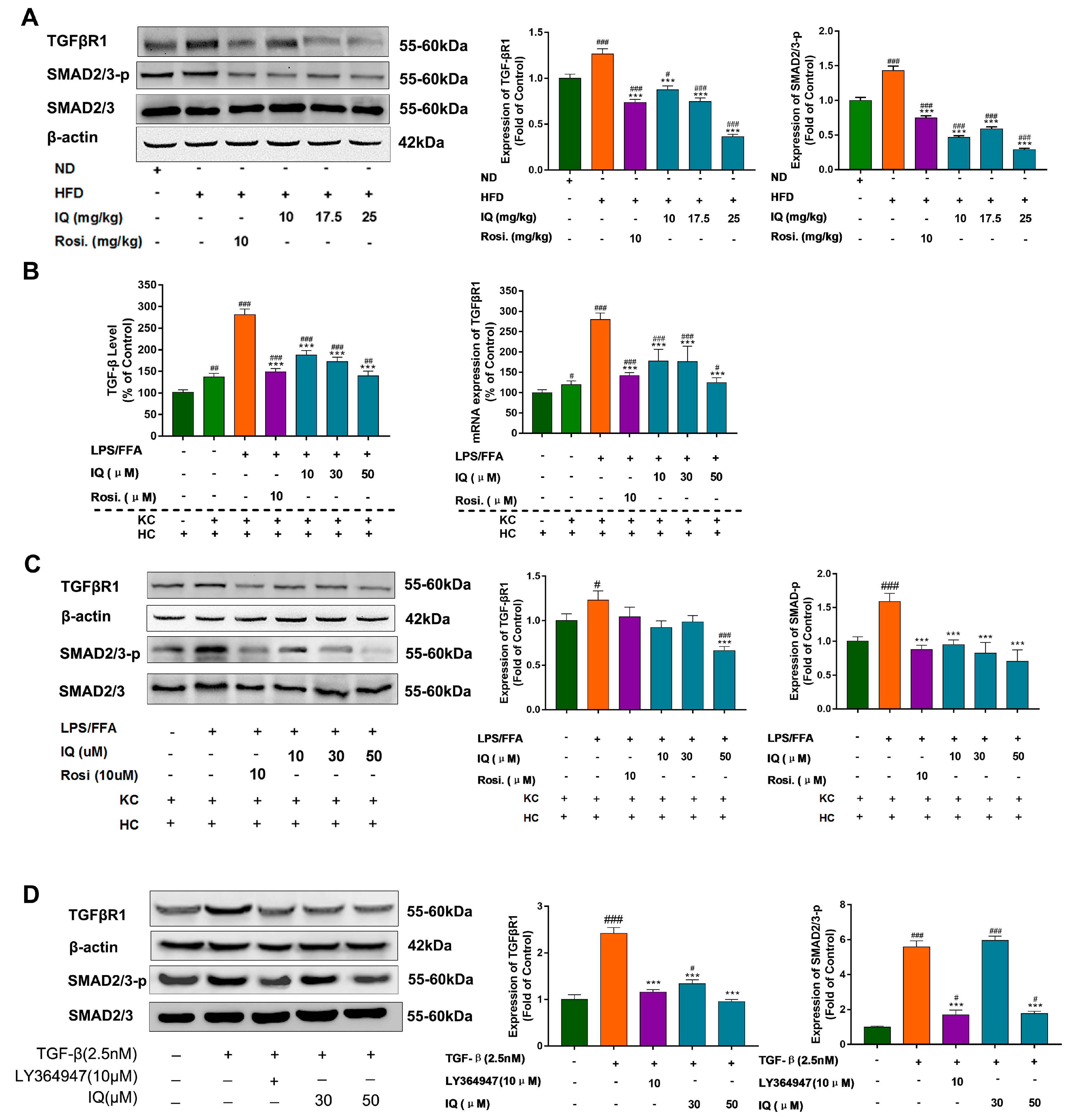
2.4. IQ Suppresses TGF-β Release, Downregulate TGF-βR1 in HFD Induced Rats and Co-Culture Model
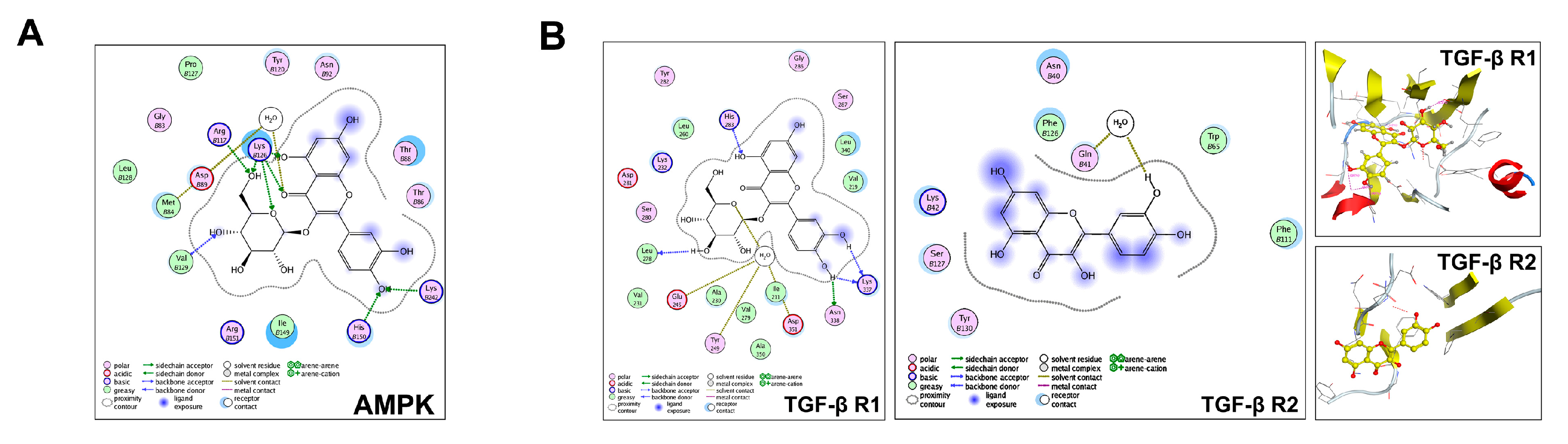
2.5. Predict the Potential Mechanism of IQ by Molecular Docking
3. Discussion
4. Materials and Methods
4.1. Animal Model and Experimental Design
4.2. Physical and Biochemical Analysis
4.3. Immunohistopathological Analysis
4.4. Measurement of ROS
4.5. Cytokine Array
4.6. Western Blotting
4.7. Isolation of Primary Hepatocytes and Kupffer Cells
4.8. Co-Culture Model
4.9. Cell Culture
4.10. ELISA
4.11. Quantitative Real-Time PCR
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global Epidemiology of Nonalcoholic Fatty Liver Disease—Meta-Analytic Assessment of Prevalence, Incidence, and Outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Maurice, J.; Manousou, P. Non-Alcoholic Fatty Liver Disease. Clin. Med. 2018, 18, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Ballestri, S.; Nascimbeni, F.; Baldelli, E.; Marrazzo, A.; Romagnoli, D.; Lonardo, A. NAFLD as a Sexual Dimorphic Disease: Role of Gender and Reproductive Status in the Development and Progression of Nonalcoholic Fatty Liver Disease and Inherent Cardiovascular Risk. Adv. Ther. 2017, 34, 1291–1326. [Google Scholar] [CrossRef] [PubMed]
- Lonardo, A.; Nascimbeni, F.; Maurantonio, M.; Marrazzo, A.; Rinaldi, L.; Adinolfi, L.E. Nonalcoholic Fatty Liver Disease: Evolving Paradigms. World J. Gastroenterol. 2017, 23, 6571–6592. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gao, J.; Peng, M.; Meng, H.; Ma, H.; Cai, P.; Xu, Y.; Zhao, Q.; Si, G. A Review on Central Nervous System Effects of Gastrodin. Front. Pharmacol. 2018, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Anstee, Q.M.; Giovanni, T.; Day, C.P. Progression of NAFLD to Diabetes Mellitus, Cardiovascular Disease or Cirrhosis. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 330. [Google Scholar] [CrossRef] [PubMed]
- Naik, A.; Košir, R.; Rozman, D. Genomic Aspects of NAFLD Pathogenesis. Genomics 2013, 102, 84–95. [Google Scholar] [CrossRef]
- Rotman, Y.; Sanyal, A.J. Current and Upcoming Pharmacotherapy for Non-Alcoholic Fatty Liver Disease. Gut 2017, 66, 180–190. [Google Scholar] [CrossRef]
- Ma, J.; Li, M.; Kalavagunta, P.K.; Li, J.; He, Q.; Zhang, Y.; Ahmad, O.; Yin, H.; Wang, T.; Shang, J. Protective Effects of Cichoric Acid on H2O2-Induced Oxidative Injury in Hepatocytes and Larval Zebrafish Models. Biomed. Pharmacother. 2018, 104, 679–685. [Google Scholar] [CrossRef]
- Day, E.A.; Ford, R.J.; Steinberg, G.R. AMPK as a Therapeutic Target for Treating Metabolic Diseases. Trends Endocrinol. Metabol. 2017, 28, 545–600. [Google Scholar] [CrossRef]
- Smith, B.K.; Marcinko, K.; Desjardins, E.M.; Lally, J.S.; Ford, R.J.; Steinberg, G.R. Treatment of Nonalcoholic Fatty Liver Disease: Role of AMPK. Am. J. Physiol. Endocrinol. Metabol. 2016, 311, E730–E740. [Google Scholar] [CrossRef]
- Galic, S.; Fullerton, M.D.; Schertzer, J.D.; Sikkema, S.; Marcinko, K.; Walkley, C.R.; Izon, D.; Honeyman, J.; Chen, Z.P.; van Denderen, B.J.; et al. Hematopoietic AMPK β1 Reduces Mouse Adipose Tissue Macrophage Inflammation and Insulin Resistance in Obesity. J. Clin. Investig. 2011, 121, 4903–4915. [Google Scholar] [CrossRef]
- Steinberg, G.R.; Michell, B.J.; Van Denderen, B.J.W.; Watt, M.J.; Carey, A.L.; Fam, B.C.; Andrikopoulos, S.; Proietto, J.; Görgün, C.Z.; Carling, D.; et al. Tumor Necrosis Factor A-Induced Skeletal Muscle Insulin Resistance Involves Suppression of AMP-Kinase Signaling. Cell Metabol. 2006, 4, 465–474. [Google Scholar] [CrossRef]
- Hsieh, C.S.; Chuang, J.H.; Chou, M.H.; Kao, Y.H. Dexamethasone Restores Transforming Growth Factor-β Activated Kinase 1 Expression and Phagocytosis Activity of Kupffer Cells in Cholestatic Liver Injury. Int. Immunopharmacol. 2018, 56, 310–319. [Google Scholar] [CrossRef]
- Kanzler, S.; Lohse, A.W.; Keil, A.; Henninger, J.; Dienes, H.P.; Schirmacher, P.; Rose-John, S.; Meyer Zum Büschenfelde, K.H.; Blessing, M. TGF-β1 in Liver Fibrosis: An Inducible Transgenic Mouse Model to Study Liver Fibrogenesis. Am. J. Physiol. 1999, 276, G1059–G1068. [Google Scholar] [CrossRef]
- Yadav, H.; Quijano, C.; Kamaraju, A.K.; Gavrilova, O.; Malek, R.; Chen, W.; Zerfas, P.; Duan, Z.; Wright, E.C.; Stuelten, C.; et al. Protection from Obesity and Diabetes by Blockade of TGF-β/Smad3 Signaling. Cell Metabol. 2011, 14, 67–79. [Google Scholar] [CrossRef]
- Tan, C.K.; Leuenberger, N.; Tan, M.J.; Yan, Y.W.; Chen, Y.; Kambadur, R.; Wahli, W.; Tan, N.S. Smad3 Deficiency in Mice Protects against Insulin Resistance and Obesity Induced by a High-Fat Diet. Diabetes 2011, 4, 464–476. [Google Scholar] [CrossRef]
- Hart, K.M.; Fabre, T.; Sciurba, J.C.; Gieseck, R.L., III; Borthwick, L.A.; Vannella, K.M.; Acciani, T.H.; De Queiroz Prado, R.; Thompson, R.W.; White, S.; et al. Type 2 Immunity Is Protective in Metabolic Disease but Exacerbates NAFLD Collaboratively with TGF-β. Sci. Translat. Med. 2017, 9. [Google Scholar] [CrossRef]
- Li, R.; Yuan, C.; Dong, C.; Shuang, S.; Choi, M.M. In Vivo Antioxidative Effect of Isoquercitrin on Cadmium-Induced Oxidative Damage to Mouse Liver and Kidney. Naunyn Schmiedebergs Arch. Pharmacol. 2011, 383, 437–445. [Google Scholar] [CrossRef]
- Jung, S.H.; Kim, B.J.; Lee, E.H.; Osborne, N.N. Isoquercitrin Is the Most Effective Antioxidant in the Plant Thuja Orientalis and Able to Counteract Oxidative-Induced Damage to a Transformed Cell Line (RGC-5 Cells). Neurochem. Int. 2010, 57, 713–721. [Google Scholar] [CrossRef]
- Boots, A.W.; Drent, M.; de Boer, V.C.; Bast, A.; Haenen, G.R. Quercetin Reduces Markers of Oxidative Stress and Inflammation in Sarcoidosis. Clin. Nutr. 2011, 30, 506–512. [Google Scholar] [CrossRef]
- Hoek-van den Hil, E.F.; Keijer, J.; Bunschoten, A.; Vervoort, J.J.; Stankova, B.; Bekkenkamp, M.; Herreman, L.; Venema, D.; Hollman, P.C.; Tvrzicka, E.; et al. Quercetin Induces Hepatic Lipid Omega-Oxidation and Lowers Serum Lipid Levels in Mice. PLoS ONE 2013, 8, e51588. [Google Scholar] [CrossRef]
- Guo, X.D.; Zhang, D.Y.; Gao, X.J.; Parry, J.; Liu, K.; Liu, B.L.; Wang, M. Quercetin and Quercetin-3-O-Glucuronide Are Equally Effective in Ameliorating Endothelial Insulin Resistance through Inhibition of Reactive Oxygen Species-Associated Inflammation. Mol. Nutr. Food Res. 2013, 57, 1037–1045. [Google Scholar] [CrossRef]
- Ding, L.; Liu, J.L.; Hassan, W.; Wang, L.L.; Yan, F.R.; Shang, J. Lipid Modulatory Activities of Cichorium Glandulosum Boiss Et Huet Are Mediated by Multiple Components within Hepatocytes. Sci. Rep. 2014, 4, 4715. [Google Scholar] [CrossRef]
- Motoyama, K.; Koyama, H.; Moriwaki, M.; Emura, K.; Okuyama, S.; Sato, E.; Inoue, M.; Shioi, A.; Nishizawa, Y. Atheroprotective and Plaque-Stabilizing Effects of Enzymatically Modified Isoquercitrin in Atherogenic Apoe-Deficient Mice. Nutrition 2009, 25, 421–427. [Google Scholar] [CrossRef]
- Lee, S.; Park, H.S.; Notsu, Y.; Ban, H.S.; Kim, Y.P.; Ishihara, K.; Hirasawa, N.; Jung, S.H.; Lee, Y.S.; Lim, S.S.; et al. Effects of Hyperin, Isoquercitrin and Quercetin on Lipopolysaccharide-Induced Nitrite Production in Rat Peritoneal Macrophages. Phytother. Res. 2008, 22, 1552–1556. [Google Scholar] [CrossRef]
- Li, H.; Zhang, M.; Ma, G. Hypolipidemic Effect of the Polysaccharide from Pholiota Nameko. Nutrition 2010, 26, 556–562. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, X.H.; Ebersole, B.; Ribnicky, D.; Wang, Z.Q. Bitter Melon Extract Attenuating Hepatic Steatosis May Be Mediated by FGF21 and AMPK/Sirt1 Signaling in Mice. Sci. Rep. 2013, 3, 3142. [Google Scholar] [CrossRef]
- Li, Y.; Xu, S.; Mihaylova, M.M.; Zheng, B.; Hou, X.; Jiang, B.; Park, O.; Luo, Z.; Lefai, E.; Shyy, J.Y.; et al. AMPK Phosphorylates and Inhibits SREBP Activity to Attenuate Hepatic Steatosis and Atherosclerosis in Diet-Induced Insulin-Resistant Mice. Cell Metabol. 2011, 13, 376–388. [Google Scholar] [CrossRef]
- Shaw, R.J.; Lamia, K.A.; Vasquez, D.; Koo, S.H.; Bardeesy, N.; Depinho, R.A.; Montminy, M.; Cantley, L.C. The Kinase LKB1 Mediates Glucose Homeostasis in Liver and Therapeutic Effects of Metformin. Science 2005, 310, 1642–1646. [Google Scholar] [CrossRef]
- Woods, A.; Azzout-Marniche, D.; Foretz, M.; Stein, S.C.; Lemarchand, P.; Ferre, P.; Foufelle, F.; Carling, D. Characterization of the Role of Amp-Activated Protein Kinase in the Regulation of Glucose-Activated Gene Expression Using Constitutively Active and Dominant Negative Forms of the Kinase. Mol. Cell. Biol. 2000, 20, 6704–6711. [Google Scholar] [CrossRef]
- Ruderman, N.B.; Park, H.; Kaushik, V.K.; Dean, D.; Constant, S.; Prentki, M.; Saha, A.K. AMPK as a Metabolic Switch in Rat Muscle, Liver and Adipose Tissue after Exercise. Acta Physiol. Scand. 2003, 178, 435–442. [Google Scholar] [CrossRef]
- Horton, J.D.; Goldstein, J.L.; Brown, M.S. SREBPs: Activators of the Complete Program of Cholesterol and Fatty Acid Synthesis in the Liver. J. Clin. Investig. 2002, 109, 1125–1131. [Google Scholar] [CrossRef]
- Guo, X.; Li, H.; Xu, H.; Halim, V.; Zhang, W.; Wang, H.; Ong, K.T.; Woo, S.L.; Walzem, R.L.; Mashek, D.G.; et al. Palmitoleate Induces Hepatic Steatosis but Suppresses Liver Inflammatory Response in Mice. PLoS ONE 2012, 7, e39286. [Google Scholar] [CrossRef]
- Burri, L.; Thoresen, G.H.; Berge, R.K. The Role of Pparalpha Activation in Liver and Muscle. PPAR Res. 2010, 1, 542359. [Google Scholar] [CrossRef]
- Cho, H.K.; Kim, S.Y.; Yoo, S.K.; Choi, Y.H.; Cheong, J. Fatty Acids Increase Hepatitis B Virus X Protein Stabilization and HBx-Induced Inflammatory Gene Expression. FEBS J. 2014, 281, 2228–2239. [Google Scholar] [CrossRef]
- Huang, W.; Metlakunta, A.; Dedousis, N.; Zhang, P.; Sipula, I.; Dube, J.J.; Scott, D.K.; O’Doherty, R.M. Depletion of Liver Kupffer Cells Prevents the Development of Diet-Induced Hepatic Steatosis and Insulin Resistance. Diabetes 2010, 59, 347–357. [Google Scholar] [CrossRef]
- Li, H.; Zheng, H.W.; Chen, H.; Xing, Z.Z.; You, H.; Cong, M.; Jia, J.D. Hepatitis B Virus Particles Preferably Induce Kupffer Cells to Produce TGF-β1 over Pro-Inflammatory Cytokines. Dig. Liver Dis. 2012, 44, 328–333. [Google Scholar] [CrossRef]
- Stewart, A.G.; Thomas, B.; Koff, J. TGF-β: Master Regulator of Inflammation and Fibrosis. Respirology 2018, 23, 1096–1097. [Google Scholar] [CrossRef]
- Meng, X.M.; Nikolic-Paterson, D.J.; Lan, H.Y. TGF-β: The Master Regulator of Fibrosis. Nat. Rev. Nephrol. 2016, 12, 325–338. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, Y.; Wang, L.; Fan, F.; Zhu, L.; Li, Z.; Ruan, X.; Huang, H.; Wang, Z.; Huang, Z.; et al. Amelioration of High Fat Diet Induced Liver Lipogenesis and Hepatic Steatosis by Interleukin-22. J. Hepatol. 2010, 53, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Rashid, K.; Bhattacharya, S.; Sil, P.C. Protective Role of D-Saccharic Acid-1,4-Lactone in Alloxan Induced Oxidative Stress in the Spleen Tissue of Diabetic Rats Is Mediated by Suppressing Mitochondria Dependent Apoptotic Pathway. Free Radic. Res. 2012, 46, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Monem, N.M.; Abdel-Azeem, A.M.; El-Ashry, E.S.H.; Ghareeb, D.A.; Nabil-Adam, A. Pretreatment Hepatoprotective Effect of the Marine Fungus Derived from Sponge on Hepatic Toxicity Induced by Heavy Metals in Rats. Biomed. Res. Int. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Yang, S.; Koo, D.J.; Ornan, D.A.; Chaudry, I.H.; Wang, P. The Role of Kupffer Cell Alpha(2)-Adrenoceptors in Norepinephrine-Induced TNF-Alpha Production. Biochim. Biophys. Acta 2001, 1537, 49–57. [Google Scholar] [CrossRef]
- Hassan, W.; Rongyin, G.; Daoud, A.; Ding, L.; Wang, L.; Liu, J.; Shang, J. Reduced Oxidative Stress Contributes to the Lipid Lowering Effects of Isoquercitrin in Free Fatty Acids Induced Hepatocytes. Oxid. Med. Cell. Longev. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | HFD (g/100g) |
|---|---|
| lard | 10 |
| bile salt | 0.5 |
| cholesterol | 1 |
| powdered milk | 5 |
| egg yolk powder | 3 |
| sugar | 5 |
| methylthiouracil | 0.2 |
| basal diet * | 75.3 |
| Gene | Species | Forward (F) and Reverse (R) Primer Sequence |
|---|---|---|
| SREBP-1C | rat | F: ACAGCACAGCA ACCAG AAACTC R: TTCATGCCCTCCATAGACACAT |
| FAS | rat | F: TTGGCTTAG TGAT TGCATCTCGT R: CAGGGTCTCTGTCCTCCTTTTGT |
| PPAR-γ | rat | F: GAAGCCCTTTGGTGACTTTATG R: AGGTTGTCTTGGATGTCCTCGA |
| CPT-1 | rat | F: TCAGAGGATGGACACTGTAAAGGAG R: CCGAAAGAGTCAAATGGGAAGG |
| TGF-βR1 | rat | F: CCACAGAGTAGGCACTAAAAGGTAT R: ACAAGATCATAGTAAGGCAACTGGT |
| TNF-α | rat | F: CACCATGAGCACGGAAAGCATGA R: CGCCTCACAGAGCAATGACTCCA |
| IL-6 | rat | F: CACTTCACAAGTCGGAGGCT R: AGCACACTAGGTTTGCCGAG |
| IL1 β | rat | F: AGGAGAGACAAGCAACGACA R: CTTTTCCATCTTCTTCTTTGGGTAT |
| GAPDH | rat | F: CAACGGGAAACCCATCACCA R: ACGCCAGTAGACTCCACGACAT |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, G.; Ma, J.; Huang, Q.; Yin, H.; Han, J.; Li, M.; Deng, Y.; Wang, B.; Hassan, W.; Shang, J. Isoquercetin Improves Hepatic Lipid Accumulation by Activating AMPK Pathway and Suppressing TGF-β Signaling on an HFD-Induced Nonalcoholic Fatty Liver Disease Rat Model. Int. J. Mol. Sci. 2018, 19, 4126. https://doi.org/10.3390/ijms19124126
Qin G, Ma J, Huang Q, Yin H, Han J, Li M, Deng Y, Wang B, Hassan W, Shang J. Isoquercetin Improves Hepatic Lipid Accumulation by Activating AMPK Pathway and Suppressing TGF-β Signaling on an HFD-Induced Nonalcoholic Fatty Liver Disease Rat Model. International Journal of Molecular Sciences. 2018; 19(12):4126. https://doi.org/10.3390/ijms19124126
Chicago/Turabian StyleQin, Guohong, Ji Ma, Qiongshan Huang, Hongli Yin, Jichun Han, Maoru Li, Yang Deng, Bing Wang, Waseem Hassan, and Jing Shang. 2018. "Isoquercetin Improves Hepatic Lipid Accumulation by Activating AMPK Pathway and Suppressing TGF-β Signaling on an HFD-Induced Nonalcoholic Fatty Liver Disease Rat Model" International Journal of Molecular Sciences 19, no. 12: 4126. https://doi.org/10.3390/ijms19124126
APA StyleQin, G., Ma, J., Huang, Q., Yin, H., Han, J., Li, M., Deng, Y., Wang, B., Hassan, W., & Shang, J. (2018). Isoquercetin Improves Hepatic Lipid Accumulation by Activating AMPK Pathway and Suppressing TGF-β Signaling on an HFD-Induced Nonalcoholic Fatty Liver Disease Rat Model. International Journal of Molecular Sciences, 19(12), 4126. https://doi.org/10.3390/ijms19124126