Zinc: A Necessary Ion for Mammalian Sperm Fertilization Competency




Abstract
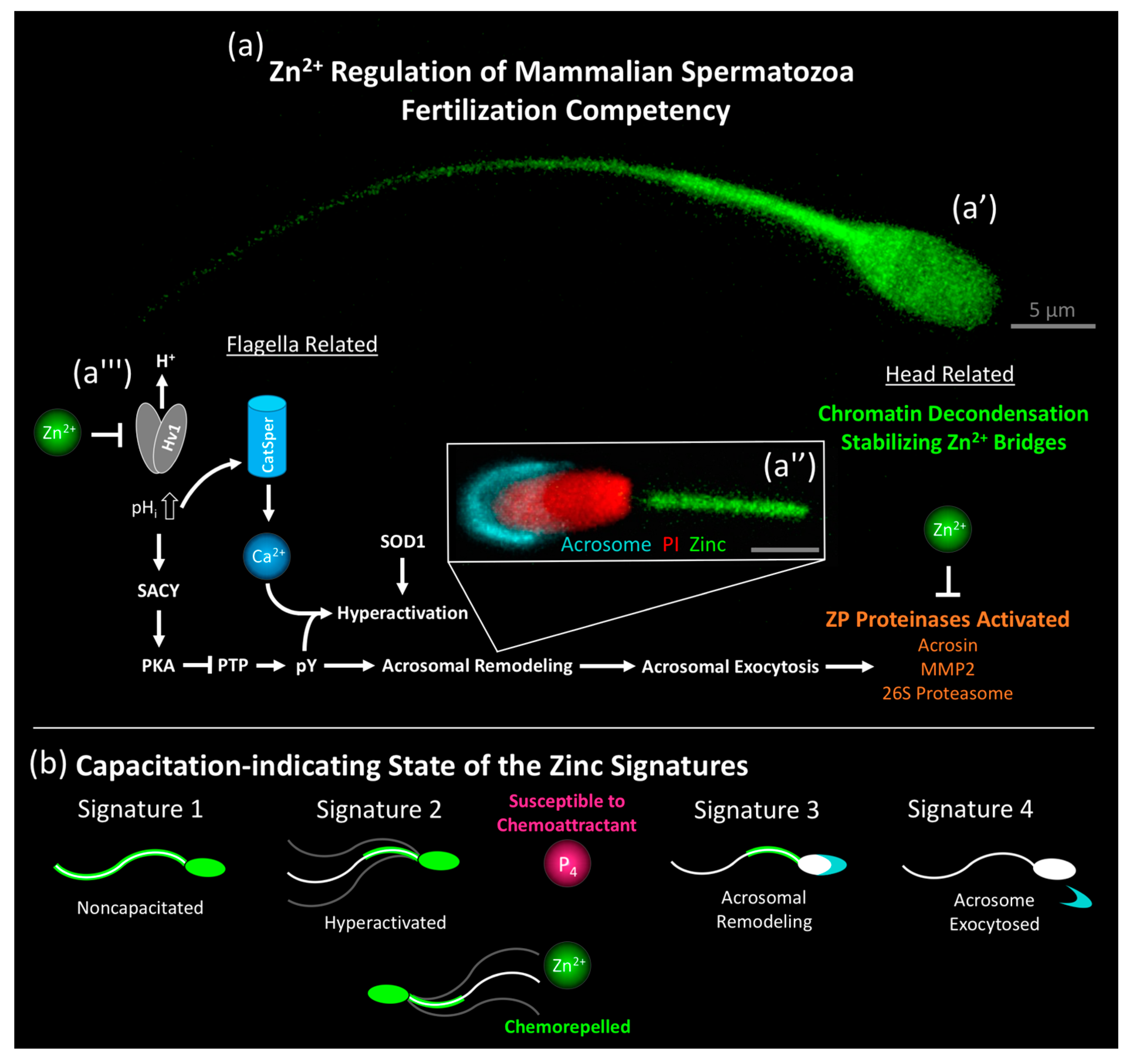
1. Introduction—Encyclopedism of Biological Zinc
2. Zinc-Interacting Sperm Proteins
3. Zinc-Containing Sperm Proteins
4. Zinc as a Regulator of Sperm Capacitation and Fertilization
5. Effect of Zinc Supplementation on Male Fertility
6. Spermatotoxicity and Reprotoxicity of High Zinc Contamination and Zinc Deficiency
7. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MDPI | Multidisciplinary Digital Publishing Institute |
| AI | Artificial insemination |
| Zn | Zinc |
| Zn2+ | Zinc ion |
| ZIP | Zrt- and Irt-like protein |
| ZnT | Zinc transporter |
| MT | Metallothionein |
| TPEN | N,N,N′,N′-tetrakis(2-pyridinylmethyl)-1,2-ethanediamine |
| ODF | Outer dense fibers |
| EDTA | 2,2′,2″,2′′′-(Ethane-1,2-diyldinitrilo)tetraacetic acid |
| ZnBP | Zinc-binding proteins |
| MMP | Matrix metalloproteinase |
| Ca2+ | Calcium ion |
| ZP | Zona pellucida |
| ICSI | Intracytoplasmic sperm injection |
| SOD | Superoxide dismutase |
| LDH | Lactate dehydrogenase |
| ACE | Angiotensin converting enzyme |
| ALP | Alkaline phosphatase |
| ADAM | A disintegrin and metalloproteinase |
| Hv1 | Hydrogen voltage-gated channel 1 |
| DMSA | (2R,3S)-2,3-Bis(sulfanyl)butanedioic acid |
| DMPS | 2,3-dimercaptopropane-1-sulfonate |
| ZnR | Zn-sensing receptor |
| GPCR | G-protein-coupled receptor |
| EGFR | Epidermal growth factor receptor |
| PI3K | Phosphoinositide 3-kinase |
| IVF | In vitro fertilization |
| pY | Protein tyrosine phosphorylation |
| SACY | Soluble sperm adenylyl cyclase |
| PKA | Protein kinase A |
| PTP | Protein tyrosine phosphatase |
| ROS | Reactive oxygen species |
| IUI | Intrauterine insemination |
References
- Levaot, N.; Hershfinkel, M. How cellular Zn2+ signaling drives physiological functions. Cell Calcium 2018, 75, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.R. Critical Role of Zinc as Either an Antioxidant or a Prooxidant in Cellular Systems. Oxid. Med. Cell. Longev. 2018, 2018, 9156285. [Google Scholar] [CrossRef] [PubMed]
- Lovell, M.A. A potential role for alterations of zinc and zinc transport proteins in the progression of Alzheimer’s disease. J. Alzheimer’s Dis. 2009, 16, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Szewczyk, B. Zinc homeostasis and neurodegenerative disorders. Front. Aging Neurosc. 2013, 5, 33. [Google Scholar] [CrossRef] [PubMed]
- Prakash, A.; Bharti, K.; Majeed, A.B. Zinc: Indications in brain disorders. Fundament. Clin. Pharmacol. 2015, 29, 131–149. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.S. Zinc: Role in immunity, oxidative stress and chronic inflammation. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Giachi, G.; Pallecchi, P.; Romualdi, A.; Ribechini, E.; Lucejko, J.J.; Colombini, M.P.; Mariotti Lippi, M. Ingredients of a 2,000-y-old medicine revealed by chemical, mineralogical, and botanical investigations. Proc. Natl. Acad. Sci. USA 2013, 110, 1193–1196. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.S. Discovery of human zinc deficiency: Its impact on human health and disease. Adv. Nutr. 2013, 4, 176–190. [Google Scholar] [CrossRef] [PubMed]
- Outten, C.E.; O’Halloran, T.V. Femtomolar sensitivity of metalloregulatory proteins controlling zinc homeostasis. Science 2001, 292, 2488–2492. [Google Scholar] [CrossRef] [PubMed]
- Beyersmann, D.; Haase, H. Functions of zinc in signaling, proliferation and differentiation of mammalian cells. Biometals 2001, 14, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Wastney, M.E.; Aamodt, R.L.; Rumble, W.F.; Henkin, R.I. Kinetic analysis of zinc metabolism and its regulation in normal humans. Am. J. Physiol. 1986, 251 Pt 2, R398–R408. [Google Scholar] [CrossRef]
- Bentley, P.J.; Grubb, B.R. Effects of a zinc-deficient diet on tissue zinc concentrations in rabbits. J. Anim. Sci. 1991, 69, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- He, L.S.; Yan, X.S.; Wu, D.C. Age-dependent variation of zinc-65 metabolism in LACA mice. Int. J. Radiat. Biol. 1991, 60, 907–916. [Google Scholar] [PubMed]
- Llobet, J.M.; Domingo, J.L.; Colomina, M.T.; Mayayo, E.; Corbella, J. Subchronic oral toxicity of zinc in rats. Bull. Environ. Contam. Toxicol. 1988, 41, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.M.; Schoenheit, J.E.; Weaver, A.D. Pretreatment and heavy metal LD50 values. Toxicol. Appl. Pharmacol. 1979, 49, 41–44. [Google Scholar] [CrossRef]
- Prasad, A.S.; Oberleas, D. Binding of zinc to amino acids and serum proteins in vitro. J. Lab. Clin. Med. 1970, 76, 416–425. [Google Scholar] [PubMed]
- Scott, B.J.; Bradwell, A.R. Identification of the serum binding proteins for iron, zinc, cadmium, nickel, and calcium. Clin. Chem. 1983, 29, 629–633. [Google Scholar] [PubMed]
- Rukgauer, M.; Klein, J.; Kruse-Jarres, J.D. Reference values for the trace elements copper, manganese, selenium, and zinc in the serum/plasma of children, adolescents, and adults. J. Trace Elem. Med. Biol. 1997, 11, 92–98. [Google Scholar] [CrossRef]
- Vallee, B.L.; Falchuk, K.H. The biochemical basis of zinc physiology. Physiol. Rev. 1993, 73, 79–118. [Google Scholar] [CrossRef] [PubMed]
- Lichten, L.A.; Cousins, R.J. Mammalian zinc transporters: Nutritional and physiologic regulation. Ann. Rev. Nutr. 2009, 29, 153–176. [Google Scholar] [CrossRef] [PubMed]
- Foresta, C.; Garolla, A.; Cosci, I.; Menegazzo, M.; Ferigo, M.; Gandin, V.; De Toni, L. Role of zinc trafficking in male fertility: From germ to sperm. Hum. Reprod. 2014, 29, 1134–1145. [Google Scholar] [CrossRef] [PubMed]
- Croxford, T.P.; McCormick, N.H.; Kelleher, S.L. Moderate zinc deficiency reduces testicular Zip6 and Zip10 abundance and impairs spermatogenesis in mice. J. Nutr. 2011, 141, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Zalewski, P.D.; Forbes, I.J.; Betts, W.H. Correlation of apoptosis with change in intracellular labile Zn(II) using zinquin [(2-methyl-8-p-toluenesulphonamido-6-quinolyloxy)acetic acid], a new specific fluorescent probe for Zn(II). Biochem. J. 1993, 296 Pt 2, 403–408. [Google Scholar] [CrossRef]
- Wellenreuther, G.; Cianci, M.; Tucoulou, R.; Meyer-Klaucke, W.; Haase, H. The ligand environment of zinc stored in vesicles. Biochem. Biophys. Res. Commun. 2009, 380, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Dittmer, P.J.; Park, J.G.; Jansen, K.B.; Palmer, A.E. Measuring steady-state and dynamic endoplasmic reticulum and Golgi Zn2+ with genetically encoded sensors. Proc. Natl. Acad. Sci. USA 2011, 108, 7351–7356. [Google Scholar] [CrossRef] [PubMed]
- Sensi, S.L.; Canzoniero, L.M.; Yu, S.P.; Ying, H.S.; Koh, J.Y.; Kerchner, G.A.; Choi, D.W. Measurement of intracellular free zinc in living cortical neurons: Routes of entry. J. Neurosci. 1997, 17, 9554–9564. [Google Scholar] [CrossRef] [PubMed]
- Vinkenborg, J.L.; Nicolson, T.J.; Bellomo, E.A.; Koay, M.S.; Rutter, G.A.; Merkx, M. Genetically encoded FRET sensors to monitor intracellular Zn2+ homeostasis. Nat. Methods 2009, 6, 737–740. [Google Scholar] [CrossRef] [PubMed]
- Tapiero, H.; Tew, K.D. Trace elements in human physiology and pathology: Zinc and metallothioneins. Biomed. Pharmacother. 2003, 57, 399–411. [Google Scholar] [CrossRef]
- Coyle, P.; Philcox, J.C.; Carey, L.C.; Rofe, A.M. Metallothionein: The multipurpose protein. Cell. Mol. Life Sci. 2002, 59, 627–647. [Google Scholar] [CrossRef] [PubMed]
- Blindauer, C.A.; Leszczyszyn, O.I. Metallothioneins: Unparalleled diversity in structures and functions for metal ion homeostasis and more. Nat. Prod. Rep. 2010, 27, 720–741. [Google Scholar] [CrossRef] [PubMed]
- Maret, W.; Krezel, A. Cellular zinc and redox buffering capacity of metallothionein/thionein in health and disease. Mol. Med. 2007, 13, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.H.; Kwon, J.W.; Liang, S.; Kim, S.H.; Li, Y.H.; Oh, J.S.; Kim, N.H.; Cui, X.S. Zinc regulates meiotic resumption in porcine oocytes via a protein kinase C-related pathway. PLoS ONE 2014, 9, e102097. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.M.; Bernhardt, M.L.; Kong, B.Y.; Ahn, R.W.; Vogt, S.; Woodruff, T.K.; O’Halloran, T.V. Zinc sparks are triggered by fertilization and facilitate cell cycle resumption in mammalian eggs. ACS Chem. Biol. 2011, 6, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.M.; Vogt, S.; O’Halloran, T.V.; Woodruff, T.K. Zinc availability regulates exit from meiosis in maturing mammalian oocytes. Nat. Chem. Biol. 2010, 6, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Que, E.L.; Duncan, F.E.; Bayer, A.R.; Philips, S.J.; Roth, E.W.; Bleher, R.; Gleber, S.C.; Vogt, S.; Woodruff, T.K.; O’Halloran, T.V. Zinc sparks induce physiochemical changes in the egg zona pellucida that prevent polyspermy. Integr. Biol. 2017, 9, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S.; Miura, C.; Kikuchi, K.; Celino, F.T.; Agusa, T.; Tanabe, S.; Miura, T. Zinc is an essential trace element for spermatogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 10859–10864. [Google Scholar] [CrossRef] [PubMed]
- Elgazar, V.; Razanov, V.; Stoltenberg, M.; Hershfinkel, M.; Huleihel, M.; Nitzan, Y.B.; Lunenfeld, E.; Sekler, I.; Silverman, W.F. Zinc-regulating proteins, ZnT-1, and metallothionein I/II are present in different cell populations in the mouse testis. J. Histochem. Cytochem. 2005, 53, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Sugihara, T.; Wadhwa, R.; Kaul, S.C.; Mitsui, Y. A novel testis-specific metallothionein-like protein, tesmin, is an early marker of male germ cell differentiation. Genomics 1999, 57, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Chi, Z.H.; Feng, W.Y.; Gao, H.L.; Zheng, W.; Huang, L.; Wang, Z.Y. ZNT7 and Zn2+ are present in different cell populations in the mouse testis. Histol. Histopathol. 2009, 24, 25–30. [Google Scholar] [PubMed]
- Shihan, M.; Chan, K.H.; Konrad, L.; Scheiner-Bobis, G. Non-classical testosterone signaling in spermatogenic GC-2 cells is mediated through ZIP9 interacting with Gnalpha11. Cell. Signal. 2015, 27, 2077–2086. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Gonzalez, I.; Trevino, C.L.; Darszon, A. Regulation of Spermatogenic Cell T-Type Ca2+ Currents by Zn2+: Implications in Male Reproductive Physiology. J. Cell. Physiol. 2016, 231, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Barney, G.H.; Orgebin-Crist, M.C.; Macapinalac, M.P. Genesis of esophageal parakeratosis and histologic changes in the testes of the zinc-deficient rat and their reversal by zinc repletion. J. Nutr. 1968, 95, 526–534. [Google Scholar] [CrossRef] [PubMed]
- Baccetti, B.; Pallini, V.; Burrini, A.G. The accessory fibers of the sperm tail. II. Their role in binding zinc in mammals and cephalopods. J. Ultrastruct. Res. 1976, 54, 261–275. [Google Scholar] [CrossRef]
- Bedford, J.M.; Calvin, H.I. Changes in -S-S- linked structures of the sperm tail during epididymal maturation, with comparative observations in sub-mammalian species. J. Exp. Zool. 1974, 187, 181–204. [Google Scholar] [CrossRef] [PubMed]
- Bjorndahl, L.; Kjellberg, S.; Roomans, G.M.; Kvist, U. The human sperm nucleus takes up zinc at ejaculation. Int. J. Androl. 1986, 9, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Porath, J.; Carlsson, J.; Olsson, I.; Belfrage, G. Metal chelate affinity chromatography, a new approach to protein fractionation. Nature 1975, 258, 598–599. [Google Scholar] [CrossRef] [PubMed]
- Bjorndahl, L.; Kvist, U. Human sperm chromatin stabilization: A proposed model including zinc bridges. Mol. Hum. Reprod. 2010, 16, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Bjorndahl, L.; Kvist, U. A model for the importance of zinc in the dynamics of human sperm chromatin stabilization after ejaculation in relation to sperm DNA vulnerability. Syst. Biol. Reprod. Med. 2011, 57, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Kvist, U. Importance of spermatozoal zinc as temporary inhibitor of sperm nuclear chromatin decondensation ability in man. Acta Physiol. Scand. 1980, 109, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Kvist, U. Sperm nuclear chromatin decondensation ability. An in vitro study on ejaculated human spermatozoa. Acta physiol. Scand. Suppl. 1980, 486, 1–24. [Google Scholar] [PubMed]
- Roomans, G.M.; Lundevall, E.; Bjorndahl, L.; Kvist, U. Removal of zinc from subcellular regions of human spermatozoa by EDTA treatment studied by X-ray microanalysis. Int. J. Androl. 1982, 5, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Henkel, R.; Baldauf, C.; Bittner, J.; Weidner, W.; Miska, W. Elimination of zinc from the flagella of spermatozoa during epididymal transit is important for motility. Reprod. Technol. 2001, 10, 6. [Google Scholar]
- Andrews, J.C.; Nolan, J.P.; Hammerstedt, R.H.; Bavister, B.D. Role of zinc during hamster sperm capacitation. Biol. Reprod. 1994, 51, 1238–1247. [Google Scholar] [CrossRef]
- Steven, F.S.; Griffin, M.M.; Chantler, E.N. Inhibition of human and bovine sperm acrosin by divalent metal ions. Possible role of zinc as a regulator of acrosin activity. Int. J. Androl. 1982, 5, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, O.; Eliasson, R.; Lofman, C.O. Inhibition of the gelatinolytic and esterolytic activity of human sperm acrosin by zinc. Acta Physiol. Scand. 1982, 114, 475–476. [Google Scholar] [CrossRef] [PubMed]
- Bettger, W.J.; O’Dell, B.L. A critical physiological role of zinc in the structure and function of biomembranes. Life Sci. 1981, 28, 1425–1438. [Google Scholar] [CrossRef]
- Mankad, M.; Sathawara, N.G.; Doshi, H.; Saiyed, H.N.; Kumar, S. Seminal plasma zinc concentration and alpha-glucosidase activity with respect to semen quality. Biol. Trace Elem. Res. 2006, 110, 97–106. [Google Scholar] [CrossRef]
- Riffo, M.; Leiva, S.; Astudillo, J. Effect of zinc on human sperm motility and the acrosome reaction. Int. J. Androl. 1992, 15, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Acott, T.S.; Carr, D.W. Inhibition of bovine spermatozoa by caudal epididymal fluid: II. Interaction of pH and a quiescence factor. Biol. Reprod. 1984, 30, 926–935. [Google Scholar] [CrossRef]
- Ho, H.C.; Granish, K.A.; Suarez, S.S. Hyperactivated motility of bull sperm is triggered at the axoneme by Ca2+ and not cAMP. Dev. Biol. 2002, 250, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Babcock, D.F.; Rufo, G.A., Jr.; Lardy, H.A. Potassium-dependent increases in cytosolic pH stimulate metabolism and motility of mammalian sperm. Proc. Natl. Acad. Sci. USA 1983, 80, 1327–1331. [Google Scholar] [CrossRef] [PubMed]
- Bray, T.M.; Bettger, W.J. The physiological role of zinc as an antioxidant. Free Radic. Biol. Med. 1990, 8, 281–291. [Google Scholar] [CrossRef]
- Narasimhaiah, M.; Arunachalam, A.; Sellappan, S.; Mayasula, V.K.; Guvvala, P.R.; Ghosh, S.K.; Chandra, V.; Ghosh, J.; Kumar, H. Organic zinc and copper supplementation on antioxidant protective mechanism and their correlation with sperm functional characteristics in goats. Reprod. Domest. Anim. 2018, 53, 644–654. [Google Scholar] [CrossRef] [PubMed]
- Arver, S. Zinc and zinc ligands in human seminal plasma. I. Methodological aspects and normal findings. Int. J. Androl. 1980, 3, 629–642. [Google Scholar] [CrossRef] [PubMed]
- Arver, S.; Eliasson, R. Zinc and zinc ligands in human seminal plasma. II. Contribution by ligands of different origin to the zinc binding properties of human seminal plasma. Acta Physiol. Scand. 1982, 115, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Arver, S. Zinc and zinc ligands in human seminal plasma. III. The principal low molecular weight zinc ligand in prostatic secretion and seminal plasma. Acta Physiol. Scand. 1982, 116, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Siciliano, L.; De Stefano, C.; Petroni, M.F.; Vivacqua, A.; Rago, V.; Carpino, A. Prostatic origin of a zinc binding high molecular weight protein complex in human seminal plasma. Mol. Hum. Reprod. 2000, 6, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Robert, M.; Gagnon, C. Semenogelin I: A coagulum forming, multifunctional seminal vesicle protein. Cell. Mol. Life Sci. 1999, 55, 944–960. [Google Scholar] [CrossRef] [PubMed]
- Vivacqua, A.; Siciliano, L.; Sabato, M.; Palma, A.; Carpino, A. Prostasomes as zinc ligands in human seminal plasma. Int. J. Androl. 2004, 27, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Mogielnicka-Brzozowska, M.; Wysocki, P.; Strzezek, J.; Kordan, W. Zinc-binding proteins from boar seminal plasma—Isolation, biochemical characteristics and influence on spermatozoa stored at 4 degrees C. Acta Biochim. Pol. 2011, 58, 171–177. [Google Scholar] [PubMed]
- Johnson, L.; Wikström, S.; Nylander, G. The Vehicle for Zinc in the Prostatic Secretion of Dogs. Scand. J. Urol. Nephrol. 1969, 3, 9–11. [Google Scholar] [CrossRef] [PubMed]
- Mogielnicka-Brzozowska, M.; Strzezek, R.; Wasilewska, K.; Kordan, W. Prostasomes of canine seminal plasma—Zinc-binding ability and effects on motility characteristics and plasma membrane integrity of spermatozoa. Reprod. Domest. Anim. 2015, 50, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Andreini, C.; Banci, L.; Bertini, I.; Rosato, A. Counting the zinc-proteins encoded in the human genome. J. Proteome Res. 2006, 5, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.E. Zinc proteins: Enzymes, storage proteins, transcription factors, and replication proteins. Ann. Rev. Biochem. 1992, 61, 897–946. [Google Scholar] [CrossRef] [PubMed]
- Vallee, B.L.; Coleman, J.E.; Auld, D.S. Zinc fingers, zinc clusters, and zinc twists in DNA-binding protein domains. Proc. Natl. Acad. Sci. USA 1991, 88, 999–1003. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, F.; Rousseaux-Prevost, R.; Sautiere, P.; Rousseaux, J. P2 protamines from human sperm are zinc -finger proteins with one CYS2/HIS2 motif. Biochem. Biophys. Res. Commun. 1992, 182, 540–547. [Google Scholar] [CrossRef]
- Seiler, H.G.; Sigel, H. Handbook on Toxicity of Inorganic Compounds; Marcel Dekker: New York, NY, USA, 1988. [Google Scholar]
- Vallee, B.L.; Auld, D.S. Zinc coordination, function, and structure of zinc enzymes and other proteins. Biochemistry 1990, 29, 5647–5659. [Google Scholar] [CrossRef] [PubMed]
- Vallee, B.L.; Auld, D.S. Active zinc binding sites of zinc metalloenzymes. In Matrix Metalloproteinases and Inhibitors; Birkedal-Hansen, H., Werb, Z., Welgus, H., Van Wart, H., Eds.; Fischer: Stuttgart, Germany, 1992; p. 5. [Google Scholar]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and Biological Attributes of Matrix Metalloproteinases. Prog. Mol. Biol. Transl. Sci. 2017, 147, 1–73. [Google Scholar] [PubMed]
- Nagase, H.; Visse, R.; Murphy, G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Shimokawa Ki, K.; Katayama, M.; Matsuda, Y.; Takahashi, H.; Hara, I.; Sato, H.; Kaneko, S. Matrix metalloproteinase (MMP)-2 and MMP-9 activities in human seminal plasma. Mol. Hum. Reprod. 2002, 8, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Tentes, I.; Asimakopoulos, B.; Mourvati, E.; Diedrich, K.; Al-Hasani, S.; Nikolettos, N. Matrix metalloproteinase (MMP)-2 and MMP-9 in seminal plasma. J. Assist. Reprod. Genet. 2007, 24, 278–281. [Google Scholar] [CrossRef] [PubMed]
- Saengsoi, W.; Shia, W.Y.; Shyu, C.L.; Wu, J.T.; Warinrak, C.; Lee, W.M.; Cheng, F.P. Detection of matrix metalloproteinase (MMP)-2 and MMP-9 in canine seminal plasma. Anim. Reprod. Sci. 2011, 127, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Warinrak, C.; Wu, J.T.; Hsu, W.L.; Liao, J.W.; Chang, S.C.; Cheng, F.P. Expression of matrix metalloproteinases (MMP-2, MMP-9) and their inhibitors (TIMP-1, TIMP-2) in canine testis, epididymis and semen. Reprod. Domest. Anim. 2015, 50, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Buchman-Shaked, O.; Kraiem, Z.; Gonen, Y.; Goldman, S. Presence of matrix metalloproteinases and tissue inhibitor of matrix metalloproteinase in human sperm. J. Androl. 2002, 23, 702–708. [Google Scholar] [PubMed]
- Ferrer, M.; Rodriguez, H.; Zara, L.; Yu, Y.; Xu, W.; Oko, R. MMP2 and acrosin are major proteinases associated with the inner acrosomal membrane and may cooperate in sperm penetration of the zona pellucida during fertilization. Cell. Tissue Res. 2012, 349, 881–895. [Google Scholar] [CrossRef] [PubMed]
- Kratz, E.M.; Kaluza, A.; Ferens-Sieczkowska, M.; Olejnik, B.; Fiutek, R.; Zimmer, M.; Piwowar, A. Gelatinases and their tissue inhibitors are associated with oxidative stress: A potential set of markers connected with male infertility. Reprod. Fertil. Dev. 2016, 28, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Atabakhsh, M.; Khodadadi, I.; Amiri, I.; Mahjub, H.; Tavilani, H. Activity of Matrix Metalloproteinase 2 and 9 in Follicular Fluid and Seminal Plasma and Its Relation to Embryo Quality and Fertilization Rate. J. Reprod. Infertil. 2018, 19, 140–145. [Google Scholar] [PubMed]
- McCord, J.M.; Fridovich, I. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar] [PubMed]
- Marklund, S.L. Extracellular superoxide dismutase and other superoxide dismutase isoenzymes in tissues from nine mammalian species. Biochem. J. 1984, 222, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, J.G.; Touchstone, J.C.; Blasco, L.; Storey, B.T. Spontaneous lipid peroxidation and production of hydrogen peroxide and superoxide in human spermatozoa. Superoxide dismutase as major enzyme protectant against oxygen toxicity. J. Androl. 1987, 8, 338–348. [Google Scholar] [CrossRef] [PubMed]
- Beconi, M.T.; Francia, C.R.; Mora, N.G.; Affranchino, M.A. Effect of natural antioxidants on frozen bovine semen preservation. Theriogenology 1993, 40, 841–851. [Google Scholar] [CrossRef]
- O’Flaherty, C.; Beconi, M.; Beorlegui, N. Effect of natural antioxidants, superoxide dismutase and hydrogen peroxide on capacitation of frozen-thawed bull spermatozoa. Andrologia 1997, 29, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Kowalowka, M.; Wysocki, P.; Fraser, L.; Strzezek, J. Extracellular superoxide dismutase of boar seminal plasma. Reprod. Domest. Anim. 2008, 43, 490–496. [Google Scholar] [CrossRef] [PubMed]
- Strzezek, R.; Fraser, L. Characteristics of spermatozoa of whole ejaculate and sperm-rich fraction of dog semen following exposure to media varying in osmolality. Reprod. Biol. 2009, 9, 113–126. [Google Scholar] [CrossRef]
- Marti, E.; Mara, L.; Marti, J.I.; Muino-Blanco, T.; Cebrian-Perez, J.A. Seasonal variations in antioxidant enzyme activity in ram seminal plasma. Theriogenology 2007, 67, 1446–1454. [Google Scholar] [CrossRef] [PubMed]
- Peeker, R.; Abramsson, L.; Marklund, S.L. Superoxide dismutase isoenzymes in human seminal plasma and spermatozoa. Mol. Hum. Reprod. 1997, 3, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Jeon, S.; Song, Y.-J.; Yi, L.S.H. Proteomic analysis of boar spermatozoa and quantity changes of superoxide dismutase 1, glutathione peroxidase, and peroxiredoxin 5 during epididymal maturation. Anim. Reprod. Sci. 2012, 135, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Sikka, S.C. Relative impact of oxidative stress on male reproductive function. Curr. Med. Chem. 2001, 8, 851–862. [Google Scholar] [CrossRef] [PubMed]
- Dhanda, O.P.; Rao, B.R.; Razdan, M.N. Sorbitol dehydrogenase & hyaluronidase activity in buffalo semen. Indian J. Exp. Biol. 1981, 19, 286. [Google Scholar] [PubMed]
- Gavella, M.; Cvitkovic, P. Semen LDH-X deficiency and male infertility. Arch. Androl. 1985, 15, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Virji, N. LDH-C4 in human seminal plasma and its relationship to testicular function. I. Methodological aspects. Int. J. Androl. 1985, 8, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Wheat, T.E.; Goldberg, E. Sperm-specific lactate dehydrogenase C4: Antigenic structure and immunosuppression of fertility. In Isozymes. Current Topics in Biological and Medical Research; Scandalios, J.G., Whitt, G.S., Eds.; Liss: New York, NY, USA, 1983; pp. 113–130. [Google Scholar]
- Duan, C.; Goldberg, E. Inhibition of lactate dehydrogenase C4 (LDH-C4) blocks capacitation of mouse sperm in vitro. Cytogenet. Genome Res. 2003, 103, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, S.W.; Yi, Y.J.; Sutovsky, M.; van Leeuwen, F.W.; Conant, G.; Sutovsky, P. Identification and characterization of RING-finger ubiquitin ligase UBR7 in mammalian spermatozoa. Cell. Tissue Res. 2014, 356, 261–278. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, S.W.; Manandhar, G.; Yi, Y.J.; Gupta, S.K.; Sutovsky, M.; Odhiambo, J.F.; Powell, M.D.; Miller, D.J.; Sutovsky, P. Sperm proteasomes degrade sperm receptor on the egg zona pellucida during mammalian fertilization. PLoS ONE 2011, 6, e17256. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, A.; Joshi, P.; Kumar, M.V.; Panda, J.N.; Singh, L.N. Angiotensin converting enzyme in the testis and epididymis of mammals. Andrologia 1984, 16, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Yotsumoto, H.; Sato, S.; Shibuya, M. Localization of angiotensin converting enzyme (dipeptidyl carboxypeptidase) in swine sperm by immunofluorescence. Life Sci. 1984, 35, 1257–1261. [Google Scholar] [CrossRef]
- Krassnigg, F.; Niederhauser, H.; Placzek, R.; Frick, J.; Schill, W.B. Investigations on the functional role of angiotensin converting enzyme (ACE) in human seminal plasma. Adv. Exp. Med. Biol. 1986, 198 Pt A, 477–485. [Google Scholar]
- Brentjens, J.R.; Matsuo, S.; Andres, G.A.; Caldwell, P.R.; Zamboni, L. Gametes contain angiotensin converting enzyme (kininase II). Experientia 1986, 42, 399–402. [Google Scholar] [CrossRef] [PubMed]
- Vivet, F.; Callard, P.; Gamoudi, A. Immunolocalization of angiotensin 1 converting enzyme in the human male genital tract by the avidin-biotin-complex method. Histochemistry 1987, 86, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Dobrinski, I.; Ignotz, G.G.; Fagnan, M.S.; Yudin, S.I.; Ball, B.A. Isolation and characterization of a protein with homology to angiotensin converting enzyme from the periacrosomal plasma membrane of equine spermatozoa. Mol. Reprod. Dev. 1997, 48, 251–260. [Google Scholar] [CrossRef]
- Gatti, J.L.; Druart, X.; Guerin, Y.; Dacheux, F.; Dacheux, J.L. A 105- to 94-kilodalton protein in the epididymal fluids of domestic mammals is angiotensin I-converting enzyme (ACE); evidence that sperm are the source of this ACE. Biol. Reprod. 1999, 60, 937–945. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P.G.; O’Dell, B.L. Zinc deficiency in rats and angiotensin-converting enzyme activity: Comparative effects on lung and testis. J. Nutr. 1988, 118, 622–626. [Google Scholar] [CrossRef] [PubMed]
- Singh, U.S.; Kumar, M.V.; Panda, J.N. Angiotensin converting enzyme in semen and its possible role in capacitation. Andrologia 1985, 17, 472–475. [Google Scholar] [CrossRef] [PubMed]
- Kondoh, G.; Tojo, H.; Nakatani, Y.; Komazawa, N.; Murata, C.; Yamagata, K.; Maeda, Y.; Kinoshita, T.; Okabe, M.; Taguchi, R.; et al. Angiotensin-converting enzyme is a GPI-anchored protein releasing factor crucial for fertilization. Nat. Med. 2005, 11, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Zigo, M.; Jonakova, V.; Sulc, M.; Manaskova-Postlerova, P. Characterization of sperm surface protein patterns of ejaculated and capacitated boar sperm, with the detection of ZP binding candidates. Int. J. Biol. Macromol. 2013, 61, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Einarsson, S.; Gustafsson, B.; Settergren, I. Alkaline phosphatase activity of epididymal contents in boars with normal or reduced spermatogenesis. Andrologia 1976, 8, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Bell, D.J.; Lake, P.E. A comparison of phosphomonesterase activities in the seminal plasmas of the domestic cock, turkey tom, boar, bull, buck rabbit and of man. J. Reprod. Fertil. 1962, 3, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Soucek, D.A.; Vary, J.C. Some properties of acid and alkaline phosphates from boar sperm plasma membranes. Biol. Reprod. 1984, 31, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Bucci, D.; Isani, G.; Giaretta, E.; Spinaci, M.; Tamanini, C.; Ferlizza, E.; Galeati, G. Alkaline phosphatase in boar sperm function. Andrology 2014, 2, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Gillis, B.A.; Tamblyn, T.M. Association of bovine sperm aldolase with sperm subcellular components. Biol. Reprod. 1984, 31, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Mildvan, A.S.; Kobes, R.D.; Rutter, W.J. Magnetic resonance studies of the role of the divalent cation in the mechanism of yeast aldolase. Biochemistry 1971, 10, 1191–1204. [Google Scholar] [CrossRef] [PubMed]
- Dafeldecker, W.P.; Vallee, B.L. Organ-specific human alcohol dehydrogenase: Isolation and characterization of isozymes from testis. Biochem. Biophys. Res. Commun. 1986, 134, 1056–1063. [Google Scholar] [CrossRef]
- Khokha, A.M.; Voronov, P.P.; Zimatkin, S.M. Immunoenzyme and immunohistochemical analysis of class III alcohol dehydrogenase from human testis. Biokhimiia 1994, 59, 997–1002. [Google Scholar] [PubMed]
- Cho, C. Testicular and epididymal ADAMs: Expression and function during fertilization. Nat. Rev. Urol. 2012, 9, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Chunghee, C. Mammalian ADAMs with Testis-Specific or -Predominant Expression. In The ADAM Family of Proteases; Hooper, N., Lendeckel, U., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 239–259. [Google Scholar]
- Loeb, J. On Some Non-Specific Factors for the Entrance of the Spermatozoon into the Egg. Science 1914, 40, 316–318. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.; Yanagimachi, R. Release of hyaluronidase from guinea-pig spermatozoa through an acrosome reaction initiated by calcium. J. Reprod. Fertil. 1975, 44, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Yanagimachi, R.; Kanoh, Y. Manner of Sperm Entry in Herring Egg, with Special Reference to the Role of Calcium Ions in Fertilization. J. Fac. Sci. Hokkaido Univ. 1953, 11, 487. [Google Scholar]
- Yanagimachi, R.; Usui, N. Calcium dependence of the acrosome reaction and activation of guinea pig spermatozoa. Exp. Cell Res. 1974, 89, 161–174. [Google Scholar] [CrossRef]
- Gervasi, M.G.; Visconti, P.E. Chang’s meaning of capacitation: A molecular perspective. Mol. Reprod. Dev. 2016, 83, 860–874. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.C. Fertilizing capacity of spermatozoa deposited into the fallopian tubes. Nature 1951, 168, 697–698. [Google Scholar] [CrossRef] [PubMed]
- Kerns, K.; Zigo, M.; Drobnis, E.Z.; Sutovsky, M.; Sutovsky, P. Zinc ion flux during mammalian sperm capacitation. Nat. Commun. 2018, 9, 2061. [Google Scholar] [CrossRef] [PubMed]
- De Jonge, C. Biological basis for human capacitation-revisited. Hum. Reprod. Update 2017, 23, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Florman, H.M.; Jungnickel, M.K.; Sutton, K.A. Shedding light on sperm pHertility. Cell 2010, 140, 310–312. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Oberdorf, J.A.; Florman, H.M. pH regulation in mouse sperm: Identification of Na(+)-, Cl(−)-, and HCO3(-)-dependent and arylaminobenzoate-dependent regulatory mechanisms and characterization of their roles in sperm capacitation. Dev. Biol. 1996, 173, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Hu, J.; Bobulescu, I.A.; Quill, T.A.; McLeroy, P.; Moe, O.W.; Garbers, D.L. A sperm-specific Na+/H+ exchanger (sNHE) is critical for expression and in vivo bicarbonate regulation of the soluble adenylyl cyclase (sAC). Proc. Natl. Acad. Sci. USA 2007, 104, 9325–9330. [Google Scholar] [CrossRef] [PubMed]
- Lishko, P.V.; Botchkina, I.L.; Fedorenko, A.; Kirichok, Y. Acid extrusion from human spermatozoa is mediated by flagellar voltage-gated proton channel. Cell 2010, 140, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Henkel, R.; Bittner, J.; Weber, R.; Hüther, F.; Miska, W. Relevance of zinc in human sperm flagella and its relation to motility. Fertil. Steril. 1999, 71, 1138–1143. [Google Scholar] [CrossRef]
- Wroblewski, N.; Schill, W.-B.; Henkel, R. Metal chelators change the human sperm motility pattern. Fertil. Steril. 2003, 79, 1584–1589. [Google Scholar] [CrossRef]
- Miller, M.R.; Kenny, S.J.; Mannowetz, N.; Mansell, S.A.; Wojcik, M.; Mendoza, S.; Zucker, R.S.; Xu, K.; Lishko, P.V. Asymmetrically Positioned Flagellar Control Units Regulate Human Sperm Rotation. Cell Rep. 2018, 24, 2606–2613. [Google Scholar] [CrossRef] [PubMed]
- Kerns, K.; Morales, P.; Sutovsky, P. Regulation of Sperm Capacitation by the 26S Proteasome: An Emerging New Paradigm in Spermatology. Biol. Reprod. 2016, 94, 117. [Google Scholar] [CrossRef] [PubMed]
- Yokota, N.; Sawada, H. Sperm proteasomes are responsible for the acrosome reaction and sperm penetration of the vitelline envelope during fertilization of the sea urchin Pseudocentrotus depressus. Dev. Biol. 2007, 308, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, R.; Deppe, M.; Schulz, M.; Bravo, P.; Villegas, J.; Morales, P.; Risopatrón, J. Participation of the sperm proteasome during in vitro fertilisation and the acrosome reaction in cattle. Andrologia 2011, 43, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Chakravarty, S.; Bansal, P.; Sutovsky, P.; Gupta, S.K. Role of proteasomal activity in the induction of acrosomal exocytosis in human spermatozoa. Reprod. Biomed. Online 2008, 16, 391–400. [Google Scholar] [CrossRef]
- Ambroggio, X.I.; Rees, D.C.; Deshaies, R.J. JAMM: A Metalloprotease-Like Zinc Site in the Proteasome and Signalosome. PLOS Biol. 2003, 2, e2. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.; Kim, C.H.; Kim, J.H.; Lee, J.; Choi, J.J.; Chen, Z.A.; Lee, M.G.; Chung, K.C.; Hsu, C.Y.; Ahn, Y.S. Pyrrolidine dithiocarbamate and zinc inhibit proteasome-dependent proteolysis. Exp. Cell Res. 2004, 298, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Clapper, D.L.; Davis, J.A.; Lamothe, P.J.; Patton, C.; Epel, D. Involvement of zinc in the regulation of pHi, motility, and acrosome reactions in sea urchin sperm. J. Cell Biol. 1985, 100, 1817–1824. [Google Scholar] [CrossRef] [PubMed]
- Michailov, Y.; Ickowicz, D.; Breitbart, H. Zn2+-stimulation of sperm capacitation and of the acrosome reaction is mediated by EGFR activation. Dev. Biol. 2014, 396, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Guidobaldi, H.A.; Cubilla, M.; Moreno, A.; Molino, M.V.; Bahamondes, L.; Giojalas, L.C. Sperm chemorepulsion, a supplementary mechanism to regulate fertilization. Hum. Reprod. 2017, 32, 1560–1573. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, J.L.; Brackett, B.G. Influences of zinc on fertilisation and development of bovine oocytes in vitro. Zygote 1999, 7, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Backstrom, J.R.; Miller, C.A.; Tokes, Z.A. Characterization of neutral proteinases from Alzheimer-affected and control brain specimens: Identification of calcium-dependent metalloproteinases from the hippocampus. J. Neurochem. 1992, 58, 983–992. [Google Scholar] [CrossRef] [PubMed]
- Beek, J.; Nauwynck, H.; Maes, D.; Van Soom, A. Inhibitors of zinc-dependent metalloproteases hinder sperm passage through the cumulus oophorus during porcine fertilization in vitro. Reproduction 2012, 144, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Andreychenko, S.V.; Klepko, A.V.; Gorban, L.V.; Motryna, O.A.; Grubska, L.V.; Trofimenko, O.V. Post-Chornobyl remote radiation effects on human sperm and seminal plasma characteristics. Exp. Oncol. 2016, 38, 245–251. [Google Scholar] [PubMed]
- Colagar, A.H.; Marzony, E.T.; Chaichi, M.J. Zinc levels in seminal plasma are associated with sperm quality in fertile and infertile men. Nutr. Res. 2009, 29, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Dissanayake, D.; Wijesinghe, P.S.; Ratnasooriya, W.D.; Wimalasena, S. Effects of zinc supplementation on sexual behavior of male rats. J. Hum. Reprod. Sci. 2009, 2, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, S.K.; Prasad, A.S.; McDonald, F.D. Sexual dysfunction in uremic male: Improvement following oral zinc supplementation. Contrib. Nephrol. 1984, 38, 103–111. [Google Scholar] [PubMed]
- Prasad, A.S.; Mantzoros, C.S.; Beck, F.W.; Hess, J.W.; Brewer, G.J. Zinc status and serum testosterone levels of healthy adults. Nutrition 1996, 12, 344–348. [Google Scholar] [CrossRef]
- Kilic, M. Effect of fatiguing bicycle exercise on thyroid hormone and testosterone levels in sedentary males supplemented with oral zinc. Neuro Endocrinol. Lett. 2007, 28, 681–685. [Google Scholar] [PubMed]
- Underwood, E.; Somers, M. Studies of zinc nutrition in sheep. I. The relation of zinc to growth, testicular development, and spermatogenesis in young rams. Aust. J. Agric. Res. 1969, 20, 889–897. [Google Scholar] [CrossRef]
- Irani, M.; Amirian, M.; Sadeghi, R.; Lez, J.L.; Latifnejad Roudsari, R. The Effect of Folate and Folate Plus Zinc Supplementation on Endocrine Parameters and Sperm Characteristics in Sub-Fertile Men: A Systematic Review and Meta-Analysis. Urol. J. 2017, 14, 4069–4078. [Google Scholar] [PubMed]
- Hadwan, M.H.; Almashhedy, L.A.; Alsalman, A.R. Oral Zinc Supplementation Restores Superoxide Radical Scavengers to Normal Levels in Spermatozoa of Iraqi Asthenospermic Patients. Int. J. Vitam. Nutr. Res. 2015, 85, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Arangasamy, A.; Venkata Krishnaiah, M.; Manohar, N.; Selvaraju, S.; Guvvala, P.R.; Soren, N.M.; Reddy, I.J.; Roy, K.S.; Ravindra, J.P. Advancement of puberty and enhancement of seminal characteristics by supplementation of trace minerals to bucks. Theriogenology 2018, 110, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Leitzmann, M.F.; Stampfer, M.J.; Wu, K.; Colditz, G.A.; Willett, W.C.; Giovannucci, E.L. Zinc Supplement Use and Risk of Prostate Cancer. J. Natl. Cancer Inst. 2003, 95, 1004–1007. [Google Scholar] [CrossRef] [PubMed]
- Plum, L.M.; Rink, L.; Haase, H. The essential toxin: Impact of zinc on human health. Int. J. Environ. Res. Public Health 2010, 7, 1342–1365. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, C.; Fournier, D.M. Reactive oxygen species and protein modifications in spermatozoa. Biol. Reprod. 2017, 97, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Powell, S.R. The antioxidant properties of zinc. J. Nutr. 2000, 130, 1447S–1454S. [Google Scholar] [CrossRef] [PubMed]
- Talevi, R.; Barbato, V.; Fiorentino, I.; Braun, S.; Longobardi, S.; Gualtieri, R. Protective effects of in vitro treatment with zinc, d-aspartate and coenzyme q10 on human sperm motility, lipid peroxidation and DNA fragmentation. Reprod. Biol. Endocrinol. 2013, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gualtieri, R.; Barbato, V.; Fiorentino, I.; Braun, S.; Rizos, D.; Longobardi, S.; Talevi, R. Treatment with zinc, d-aspartate, and coenzyme Q10 protects bull sperm against damage and improves their ability to support embryo development. Theriogenology 2014, 82, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Posthuma, L.; Baerselman, R.; Van Veen, R.P.M.; Dirven-Van Breemen, E.M. Single and Joint Toxic Effects of Copper and Zinc on Reproduction ofEnchytraeus crypticusin Relation to Sorption of Metals in Soils. Ecotoxicol. Environ. Saf. 1997, 38, 108–121. [Google Scholar] [CrossRef] [PubMed]
- Barceloux, D.G.; Barceloux, D. Zinc. J. Toxicol. Clin. Toxicol. 1999, 37, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Johnson, F.O.; Gilbreath, E.T.; Ogden, L.; Graham, T.C.; Gorham, S. Reproductive and developmental toxicities of zinc supplemented rats. Reprod. Toxicol. 2011, 31, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Manzo, S.; Schiavo, S.; Oliviero, M.; Toscano, A.; Ciaravolo, M.; Cirino, P. Immune and reproductive system impairment in adult sea urchin exposed to nanosized ZnO via food. Sci. Total Environ. 2017, 599–600, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Merrells, K.J.; Blewett, H.; Jamieson, J.A.; Taylor, C.G.; Suh, M. Relationship between abnormal sperm morphology induced by dietary zinc deficiency and lipid composition in testes of growing rats. Br. J. Nutr. 2009, 102, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Ding, B.; Zhong, Q. Zinc deficiency: An unexpected trigger for autophagy. J. Biol. Chem. 2017, 292, 8531–8532. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, B.S.; Eisenbach, M. Capacitation. In Fertilization; Hardy, D.M., Ed.; Academic Press: Cambridge, MA, USA, 2002; pp. 57–117. [Google Scholar]
- Aitken, R.J.; Baker, M.A.; Nixon, B. Are sperm capacitation and apoptosis the opposite ends of a continuum driven by oxidative stress? Asian J. Androl. 2015, 17, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.J.; Manandhar, G.; Oko, R.J.; Breed, W.G.; Sutovsky, P. Mechanism of sperm-zona pellucida penetration during mammalian fertilization: 26S proteasome as a candidate egg coat lysin. Soc. Reprod. Fertil. Suppl. 2007, 63, 385–408. [Google Scholar] [PubMed]


{kind=link}
| Protein | Localization 1 | Function 2 | Reference |
|---|---|---|---|
| Metallothionein I & II | Spermatocytes (rat) | Zinc ion binding | [37] |
| Tesmin | Spermatocytes (rat) | Developmental protein | [37] |
| Zinc transporter ZIP9 | Spermatogenic GC-2 cells (rat) | Zinc transmembrane transporter activity | [38] |
| Protamine-2 | Sperm nucleus (man) | Developmental protein, DNA-binding | [42,47,48] |
| Acrosin | Acrosome (man, bull) | Hydrolase, protease, serine protease | [54,55] |
| Semenoglins | Prostatic and vesicular fluids (man) | - | [64,65,66,67] |
| ZnBP1–6 | Seminal fluid (boar, dog) | - | [70,71,72] |
| Matrix metalloproteinase-2 | Seminal fluid, inner acrosomal membrane, flagellum (man, bull) | Hydrolase, metalloprotease, protease | [87] |
| Matrix metalloproteinase-9 | Seminal fluid and flagellum (man, bull) | Hydrolase, metalloprotease, protease | [87] |
| Superoxide dismutase 1, 2, 3 | Mitochondria and seminal fluids (man) | Antioxidant, oxidoreductase | [91,92,93,94,95,96,97,98,99] |
| l-lactate dehydrogenase | Seminal fluids (man) | Oxidoreductase | [102,103,104] |
| Putative E3 ubiquitin-protein ligase UBR7 | Sperm inner acrosomal membrane (boar) | Transferase | [105] |
| Angiotensin converting enzyme | Testis, epididymis, and spermatozoa (man, boar, stallion) | Carboxypeptidase, hydrolase, metalloprotease, protease | [107] |
| Alkaline phosphatase, germ cell type | Seminal fluid and sperm plasma membrane (man, boar) | Alkaline phosphatase | [119,120,121] |
| Putative class-II fructose-bisphosphate aldolase | Spermatozoa (bull) | Lyase | [123] |
| Alcohol dehydrogenase class-3 | Testis, spermatozoa (man) | Oxidoreductase | [125,126] |
| ADAMs | Spermatozoa | Integrin binding, metalloendopeptidase | Review [127] |
| Voltage-gated hydrogen channel 1 | Flagellum (man) | Ion channel | [140] |
| G-protein coupled receptor | Acrosome (bull) | G-protein coupled receptor activity | [151] |
| 26S proteasome non-ATPase regulatory subunit 14 | Inner acrosomal membrane | Hydrolase, metalloprotease, protease | [180] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kerns, K.; Zigo, M.; Sutovsky, P. Zinc: A Necessary Ion for Mammalian Sperm Fertilization Competency. Int. J. Mol. Sci. 2018, 19, 4097. https://doi.org/10.3390/ijms19124097
Kerns K, Zigo M, Sutovsky P. Zinc: A Necessary Ion for Mammalian Sperm Fertilization Competency. International Journal of Molecular Sciences. 2018; 19(12):4097. https://doi.org/10.3390/ijms19124097
Chicago/Turabian StyleKerns, Karl, Michal Zigo, and Peter Sutovsky. 2018. "Zinc: A Necessary Ion for Mammalian Sperm Fertilization Competency" International Journal of Molecular Sciences 19, no. 12: 4097. https://doi.org/10.3390/ijms19124097
APA StyleKerns, K., Zigo, M., & Sutovsky, P. (2018). Zinc: A Necessary Ion for Mammalian Sperm Fertilization Competency. International Journal of Molecular Sciences, 19(12), 4097. https://doi.org/10.3390/ijms19124097