Cholesterol Increases Lipid Binding Rate and Changes Binding Behavior of Bacillus thuringiensis Cytolytic Protein



Abstract
1. Introduction
2. Results
2.1. Changes in Lipid Bilayer Properties Upon Increasing of Cholesterol
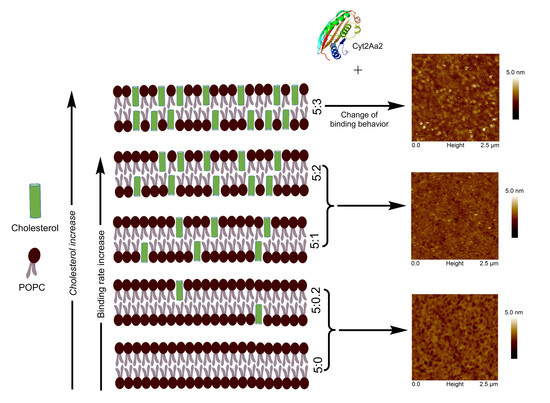
2.2. Increasing of Protein Adsorption Rate and Lipid Binding Behavior as Function of Cholesterol Content
2.3. Nanostructure of the Cyt2Aa2/Lipid Bilayer
3. Discussion
3.1. The Role of Cholesterol on Lipid Bilayer
3.2. Protein Binding as a Function of Cholesterol Content in Lipid Bilayer
4. Materials and Methods
4.1. Reagents and Buffer
4.2. Protein Preparation
4.3. Liposome Preparation and Characterization
4.4. Quartz Crystal Microbalance with Dissipation (QCM-D) Measurement
4.5. Atomic Force Microscope (AFM) Imaging
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Bt | Bacillus thuringiensis |
| Cyt protein | Cytolytic protein |
| Cyt2Aa2 protein | Cytolytic protein from Bacillus thuringiensis subsp. darmstadiensis |
| QCM-D | Quartz crystal microbalance with dissipation |
| AFM | Atomic force microscope |
| POPC | 1-palmitoyl,2-oleoyl-snglycero-3-phosphocholine |
| Chol | cholesterol |
References
- Hofte, H.; Whiteley, H.R. Insecticidal crystal proteins of Bacillus thuringiensis. Microbiol. Rev. 1989, 53, 242–255. [Google Scholar] [PubMed]
- Li, J.D.; Carroll, J.; Ellar, D.J. Crystal structure of insecticidal delta-endotoxin from Bacillus thuringiensis at 2.5 Å resolution. Nature 1991, 353, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Koni, P.A.; Ellar, D.J. Structure of the mosquitocidal delta-endotoxin CytB from Bacillus thuringiensis sp. kyushuensis and implications for membrane pore formation. J. Mol. Biol. 1996, 257, 129–152. [Google Scholar] [CrossRef] [PubMed]
- Estruch, J.J.; Warren, G.W.; Mullins, M.A.; Nye, G.J.; Craig, J.A.; Koziel, M.G. Vip3A, a novel Bacillus thuringiensis vegetative insecticidal protein with a wide spectrum of activities against lepidopteran insects. Proc. Natl. Acad. Sci. USA 1996, 93, 5389–5394. [Google Scholar] [CrossRef] [PubMed]
- Thomas, W.E.; Ellar, D.J. Bacillus thuringiensis var israelensis crystal delta-endotoxin: Effects on insect and mammalian cells in vitro and in vivo. J. Cell Sci. 1983, 60, 181–197. [Google Scholar] [PubMed]
- Cahan, R.; Friman, H.; Nitzan, Y. Antibacterial activity of Cyt1Aa from Bacillus thuringiensis subsp. israelensis. Microbiology 2008, 154, 3529–3536. [Google Scholar] [CrossRef] [PubMed]
- Maddrell, S.H.; Overton, J.A.; Ellar, D.J.; Knowles, B.H. Action of activated 27,000 Mr toxin from Bacillus thuringiensis var. israelensis on Malpighian tubules of the insect, Rhodnius prolixus. J. Cell Sci. 1989, 94 Pt 3, 601–608. [Google Scholar]
- Knowles, B.H.; White, P.J.; Nicholls, C.N.; Ellar, D.J. A broad-spectrum cytolytic toxin from Bacillus thuringiensis var. kyushuensis. Proc. Biol. Sci. 1992, 248, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Crickmore, N.; Zeigler, D.R.; Feitelson, J.; Schnepf, E.; Van Rie, J.; Lereclus, D.; Baum, J.; Dean, D.H. Revision of the nomenclature for the Bacillus thuringiensis pesticidal crystal proteins. Microbiol. Mol. Biol. Rev. 1998, 62, 807–813. [Google Scholar] [PubMed]
- Nisnevitch, M.; Cohen, S.; Ben-Dov, E.; Zaritsky, A.; Sofer, Y.; Cahan, R. Cyt2Ba of Bacillus thuringiensis israelensis: Activation by putative endogenous protease. Biochem. Biophys. Res. Commun. 2006, 344, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Dym, O.; Albeck, S.; Ben-Dov, E.; Cahan, R.; Firer, M.; Zaritsky, A. High-resolution crystal structure of activated Cyt2Ba monomer from Bacillus thuringiensis subsp. israelensis. J. Mol. Biol. 2008, 380, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Albeck, S.; Ben-Dov, E.; Cahan, R.; Firer, M.; Zaritsky, A.; Dym, O. Cyt1Aa toxin: Crystal structure reveals implications for its membrane-perforating function. J. Mol. Biol. 2011, 413, 804–814. [Google Scholar] [CrossRef] [PubMed]
- Thomas, W.E.; Ellar, D.J. Mechanism of action of Bacillus thuringiensis var israelensis insecticidal delta-endotoxin. FEBS Lett. 1983, 154, 362–368. [Google Scholar] [CrossRef]
- Gill, S.S.; Cowles, E.A.; Pietrantonio, P.V. The mode of action of Bacillus thuringiensis endotoxins. Annu. Rev. Entomol. 1992, 37, 615–636. [Google Scholar] [CrossRef] [PubMed]
- Bravo, A.; Gill, S.S.; Soberon, M. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon 2007, 49, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Pigott, C.R.; Ellar, D.J. Role of receptors in Bacillus thuringiensis crystal toxin activity. Microbiol. Mol. Biol. Rev. 2007, 71, 255–281. [Google Scholar] [CrossRef] [PubMed]
- Luckey, M. Membrane Structural Biology with Biochemical and Biophysical Foundations, 2nd ed.; Cambridge University Press: New York, NY, USA, 2014; p. 423. [Google Scholar]
- Silvius, J.R. Role of cholesterol in lipid raft formation: Lessons from lipid model systems. Biochim. Biophys. Acta 2003, 1610, 174–183. [Google Scholar] [CrossRef]
- Khan, M.S.; Dosoky, N.S.; Berdiev, B.K.; Williams, J.D. Electrochemical impedance spectroscopy for black lipid membranes fused with channel protein supported on solid-state nanopore. Eur. Biophys. J. 2016, 45, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Marheineke, K.; Grunewald, S.; Christie, W.; Reilander, H. Lipid composition of Spodoptera frugiperda (Sf9) and Trichoplusia ni (Tn) insect cells used for baculovirus infection. FEBS Lett. 1998, 441, 49–52. [Google Scholar] [CrossRef]
- Van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Suktham, K.; Pathaichindachote, W.; Promdonkoy, B.; Krittanai, C. Essential role of amino acids in alphaD-beta4 loop of a Bacillus thuringiensis Cyt2Aa2 toxin in binding and complex formation on lipid membrane. Toxicon 2013, 74, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Yandrapati, R.K. Effect of Lipid Composition on the Physical Properties of Liposomes: A Light Scattering Study. Master’s Thesis, Missouri University of Science and Technology, Rolla, MO, USA, 2012. [Google Scholar]
- Lange, Y.; Dolde, J.; Steck, T.L. The rate of transmembrane movement of cholesterol in the human erythrocyte. J. Biol. Chem. 1981, 256, 5321–5323. [Google Scholar] [PubMed]
- Steck, T.L.; Ye, J.; Lange, Y. Probing red cell membrane cholesterol movement with cyclodextrin. Biophys. J. 2002, 83, 2118–2125. [Google Scholar] [CrossRef]
- Jo, S.; Rui, H.; Lim, J.B.; Klauda, J.B.; Im, W. Cholesterol flip-flop: Insights from free energy simulation studies. J. Phys. Chem. B 2010, 114, 13342–13348. [Google Scholar] [CrossRef] [PubMed]
- Sundh, M.; Svedhem, S.; Sutherland, D.S. Influence of phase separating lipids on supported lipid bilayer formation at SiO2 surfaces. Phys. Chem. Chem. Phys. 2010, 12, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Marsh, D. Liquid-ordered phases induced by cholesterol: A compendium of binary phase diagrams. Biochim. Biophys. Acta 2010, 1798, 688–699. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.; Dosoky, N.S.; Mustafa, G.; Patel, D.; Berdiev, B.; Williams, J.D. Electrophysiology of Epithelial Sodium Channel (ENaC) Embedded in Supported Lipid Bilayer Using a Single Nanopore Chip. Langmuir 2017, 33, 13680–13688. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.; Dosoky, N.S.; Patel, D.; Weimer, J.; Williams, J.D. Lipid Bilayer Membrane in a Silicon Based Micron Sized Cavity Accessed by Atomic Force Microscopy and Electrochemical Impedance Spectroscopy. Biosensors 2017, 7, 26. [Google Scholar] [CrossRef] [PubMed]
- Crane, J.M.; Tamm, L.K. Role of Cholesterol in the Formation and Nature of Lipid Rafts in Planar and Spherical Model Membranes. Biophys. J. 2004, 86, 2965–2979. [Google Scholar] [CrossRef]
- Rabinovich, A.L.; Kornilov, V.V.; Balabaev, N.K.; Leermakers, F.A.M.; Filippov, A.V. Properties of unsaturated phospholipid bilayers: Effect of cholesterol. Biochem. Suppl. Ser. A Membr. Cell Biol. 2007, 1, 343–357. [Google Scholar] [CrossRef]
- Cathcart, K.; Patel, A.; Dies, H.; Rheinstadter, M.C.; Fradin, C. Effect of Cholesterol on the Structure of a Five-Component Mitochondria-Like Phospholipid Membrane. Membranes 2015, 5, 664–684. [Google Scholar] [CrossRef] [PubMed]
- Chalmeau, J.; Monina, N.; Shin, J.; Vieu, C.; Noireaux, V. Alpha-Hemolysin pore formation into a supported phospholipid bilayer using cell-free expression. Biochim. Biophys. Acta 2011, 1808, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.C.; Lo, Y.C.; Lin, J.Y.; Liaw, Y.C. Crystal structures and electron micrographs of fungal volvatoxin A2. J. Mol. Biol. 2004, 343, 477–491. [Google Scholar] [CrossRef] [PubMed]
- Promdonkoy, B.; Chewawiwat, N.; Tanapongpipat, S.; Luxananil, P.; Panyim, S. Cloning and characterization of a cytolytic and mosquito larvicidal delta-endotoxin from Bacillus thuringiensis subsp. darmstadiensis. Curr. Microbiol. 2003, 46, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Tharad, S.; Iturri, J.; Moreno-Cencerrado, A.; Mittendorfer, M.; Promdonkoy, B.; Krittanai, C.; Toca-Herrera, J.L. Effect of the Concentration of Cytolytic Protein Cyt2Aa2 on the Binding Mechanism on Lipid Bilayers Studied by QCM-D and AFM. Langmuir 2015, 31, 10477–10483. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| POPC:Chol Ratio (by Weight) | ∆F5 (Hz) | ∆D5 (×10−6) |
|---|---|---|
| 5:0 | −26.1 ± 1.2 | 1.6 ± 0.9 |
| 5:0.2 | −29.6 ± 2.6 | 1.6 ± 0.6 |
| 5:1 | −30.3 ± 0.5 | 1.9 ± 0.1 |
| 5:2 | −37.1 ± 1.0 | 3.6 ± 0.5 |
| 5:3 | −31.3 ± 0.6 | 2.2 ± 0.1 |
| POPC:Chol Ratio (by Weight) | ∆F5 (Hz) | ∆D5 (×10−6) | Γ (min) |
|---|---|---|---|
| 5:0 | −29 ± 2 | 2.2 ± 0.7 | 6.9 |
| 5:0.2 | −33 ± 2 | 2.2 ± 0.3 | 6.3 |
| 5:1 | −30 ± 1 | 2.7 ± 0.3 | 3.2 |
| 5:2 | −32 ± 1 | 2.5 ± 0.9 | 2.1 |
| 5:3 | −207 ± 4 | 43.7 ± 1.1 | N/A |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tharad, S.; Üzülmez, Ö.; Promdonkoy, B.; Toca-Herrera, J.L. Cholesterol Increases Lipid Binding Rate and Changes Binding Behavior of Bacillus thuringiensis Cytolytic Protein. Int. J. Mol. Sci. 2018, 19, 3819. https://doi.org/10.3390/ijms19123819
Tharad S, Üzülmez Ö, Promdonkoy B, Toca-Herrera JL. Cholesterol Increases Lipid Binding Rate and Changes Binding Behavior of Bacillus thuringiensis Cytolytic Protein. International Journal of Molecular Sciences. 2018; 19(12):3819. https://doi.org/10.3390/ijms19123819
Chicago/Turabian StyleTharad, Sudarat, Öykü Üzülmez, Boonhiang Promdonkoy, and José L. Toca-Herrera. 2018. "Cholesterol Increases Lipid Binding Rate and Changes Binding Behavior of Bacillus thuringiensis Cytolytic Protein" International Journal of Molecular Sciences 19, no. 12: 3819. https://doi.org/10.3390/ijms19123819
APA StyleTharad, S., Üzülmez, Ö., Promdonkoy, B., & Toca-Herrera, J. L. (2018). Cholesterol Increases Lipid Binding Rate and Changes Binding Behavior of Bacillus thuringiensis Cytolytic Protein. International Journal of Molecular Sciences, 19(12), 3819. https://doi.org/10.3390/ijms19123819