ω-3 Long Chain Polyunsaturated Fatty Acids as Sensitizing Agents and Multidrug Resistance Revertants in Cancer Therapy

 ,
,  and
and 
Abstract
:
{kind=link}
{kind=link}
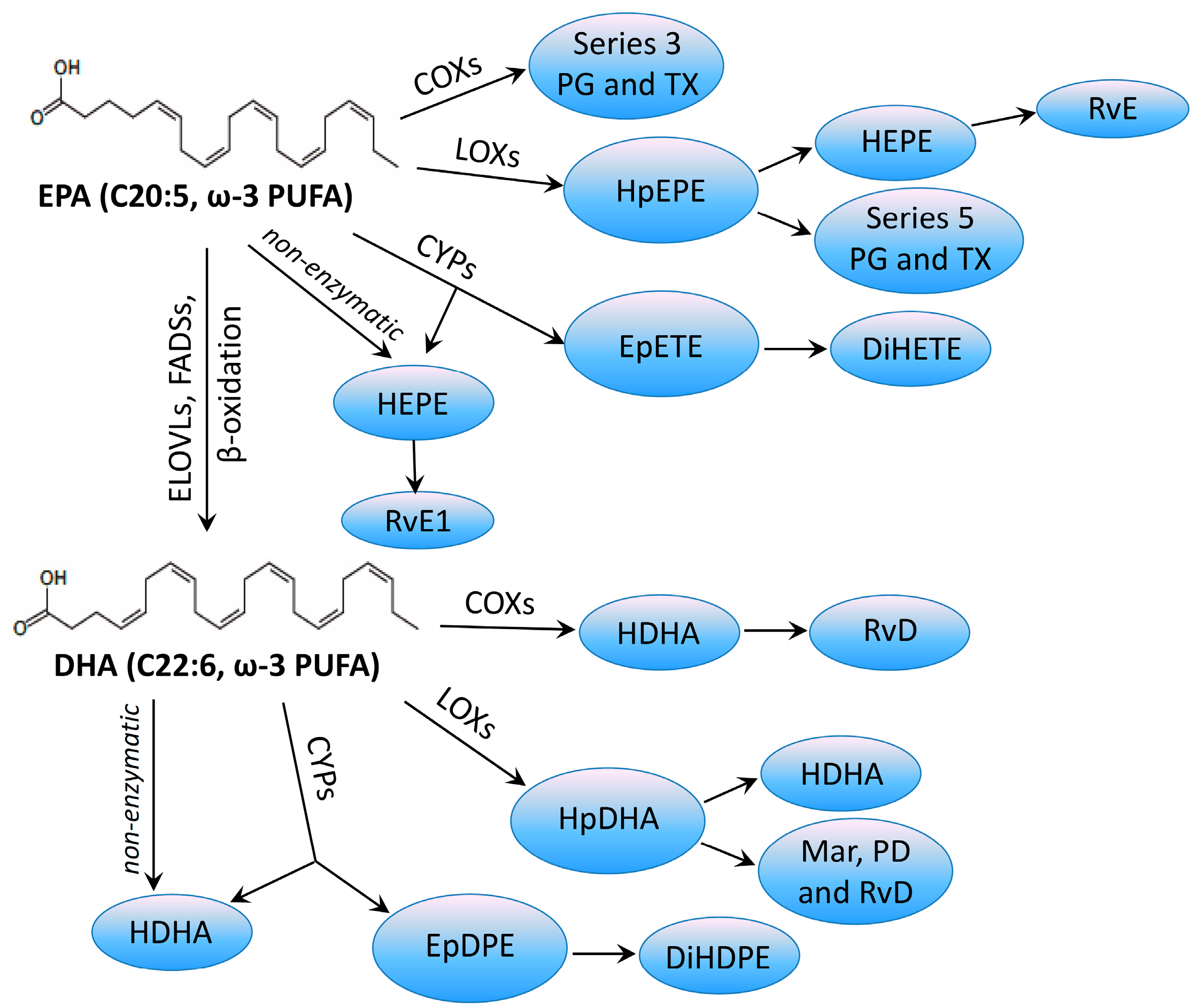{kind=link}
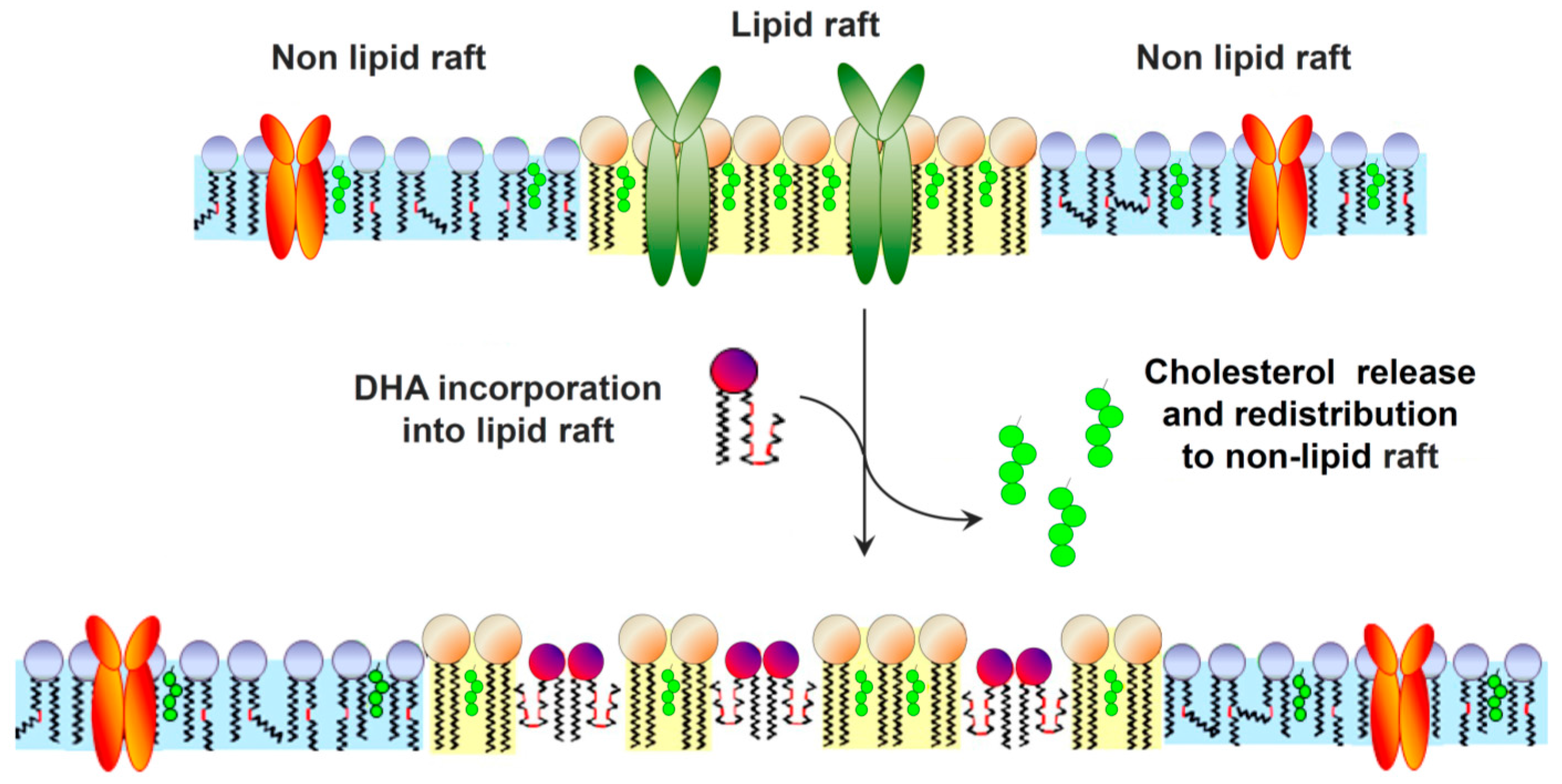{kind=link}
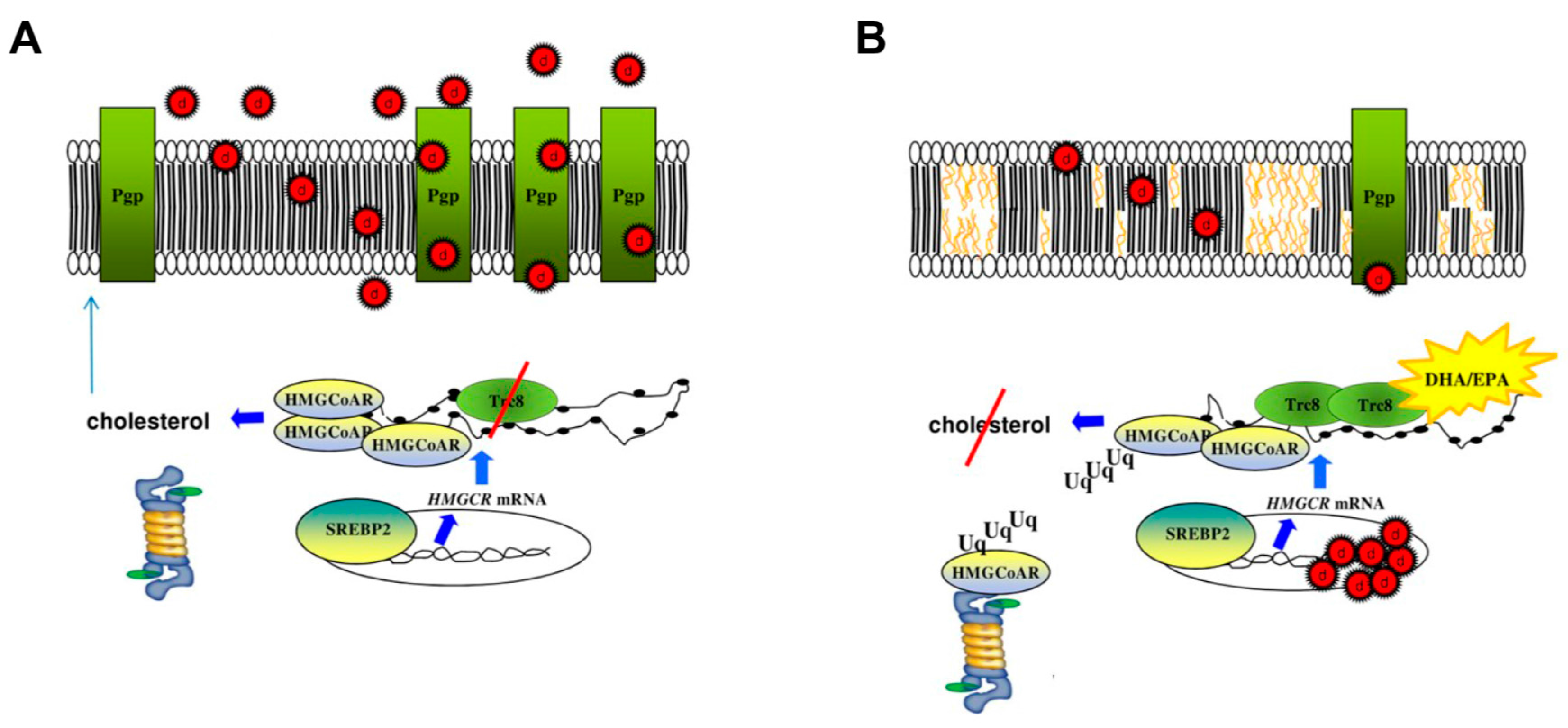{kind=link}
1. Introduction
2. In Vitro Evidences of ω-3 LCPUFA’s Positive Effects on Chemosensitization
3. Mechanisms Proposed for Chemosensitizing Effects of ω-3 LCPUFAs
4. ω-3 LCPUFA Impact on Cell Membrane Function and Lipid Raft Organization
5. ω-3 LCPUFAs as Revertants of Multidrug Resistance: In Vitro Evidences
6. ω-3 LCPUFAs as Revertants of Multidrug Resistance: Preclinical and Clinical Studies
7. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Straka, S.; Lester, J.L.; Cole, R.M.; Andridge, R.R.; Puchala, S.; Rose, A.M.; Clinton, S.K.; Belury, M.A.; Yee, L.D. Incorporation of eicosapentaenioic and docosahexaenoic acids into breast adipose tissue of women at high risk of breast cancer: A randomized clinical trial of dietary fish and n-3 fatty acid capsules. Mol. Nutr. Food Res. 2015, 59, 1780–1790. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Rani, I.; Bhatnagar, A.; Agnihotri, N. Apoptosis-Mediated Chemoprevention by Different Ratios of Fish Oil in Experimental Colon Carcinogenesis. Cancer Investig. 2016, 34, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.M.; Fujino, Y.; Kubo, T.; Ide, R.; Tokui, N.; Mizoue, T.; Ogimoto, I.; Matsuda, S.; Yoshimura, T. Fish intake and the risk of fatal prostate cancer: Findings from a cohort study in Japan. Public Health Nutr. 2009, 12, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Reynolds, L.; Mulik, R.S.; Kim, S.Y.; Van Treuren, T.; Nguyen, L.H.; Zhu, H.; Corbin, I.R. Hepatic Arterial Infusion of Low-Density Lipoprotein Docosahexaenoic Acid Nanoparticles Selectively Disrupts Redox Balance in Hepatoma Cells and Reduces Growth of Orthotopic Liver Tumors in Rats. Gastroenterology 2016, 150, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Haqq, J.; Howells, L.M.; Garcea, G.; Dennison, A.R. Targeting pancreatic cancer using a combination of gemcitabine with the omega-3 polyunsaturated fatty acid emulsion, Lipidem™. Mol. Nutr. Food Res. 2016, 60, 1437–1447. [Google Scholar] [CrossRef] [PubMed]
- Hardman, W.E.; Moyer, M.P.; Cameron, I.L. Dietary fish oil sensitizes A549 lung xenografts to doxorubicin chemotherapy. Cancer Lett. 2000, 151, 145–151. [Google Scholar] [CrossRef]
- Mackie, S.J.; Sharma, D.M.; Cooper, A.J.; Harris, N.M.; Lwaleed, B. Meglumine Eicosapentaenoic acid (MeEPA) a new soluble omega-3 fatty acid formulation: In vitro bladder cancer cytotoxicity tests in combination with epirubicin and mitomycin. Prostaglandins Leukot. Essent. Fatty Acids 2006, 75, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Qi, J.; Yang, X.; Wu, E.; Qian, S.Y. Free radical derivatives formed from cyclooxygenase-catalyzed dihomo-γ-linolenic acid peroxidation can attenuate colon cancer cell growth and enhance 5-fluorouracil’s cytotoxicity. Redox Biol. 2014, 2, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Fang, S.; Zhang, H.X.; Xu, L.X.; Zhang, Z.Q.; Yuan, K.T.; Xue, C.L.; Yu, H.L.; Zhang, S.; Li, Y.F.; et al. N-3 PUFAs have antiproliferative and apoptotic effects on human colorectal cancer stem-like cells in vitro. J. Nutr. Biochem. 2013, 24, 744–753. [Google Scholar] [CrossRef] [PubMed]
- Cha, M.C.; Meckling, K.A.; Stewart, C. Dietary docosahexaenoic acid levels influence the outcome of arabinosylcytosine chemotherapy in L1210 leukemic mice. Nutr. Cancer 2002, 44, 176–181. [Google Scholar] [CrossRef] [PubMed]
- DeGraffenried, L.A.; Friedrichs, W.E.; Fulcher, L.; Fernandes, G.; Silva, J.M.; Peralba, J.M.; Hidalgo, M. Eicosapentaenoic acid restores tamoxifen sensitivity in breast cancer cells with high Akt activity. Ann. Oncol. 2003, 14, 969–970. [Google Scholar] [CrossRef]
- Hardman, W.E.; Moyer, M.P.; Cameron, I.L. Fish oil supplementation enhanced CPT-11 (Irinotecan) efficacy against MCF7 breast carcinoma xenografts and ameliorated intestinal side effects. Br. J. Cancer 1999, 81, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Colas, S.; Paon, L.; Denis, F.; Prat, M.; Louisot, P.; Hoinard, C.; Le Floch, O.; Ogilvie, G.; Bougnoux, P. Enhanced radiosensitivity of rat autochthonous mammary tumors by dietary docosahexaenoic acid. Int. J. Cancer 2004, 109, 449–454. [Google Scholar] [CrossRef] [PubMed]
- De Aguiar Pastore Silva, J.; Emilia de Souza Fabre, M.; Waitzberg, D.L. Omega-3 supplements for patients in chemotherapy and/or radiotherapy: A systematic review. Clin. Nutr. 2015, 34, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.C. The molecular mechanisms of chemoresistance in cancers. Oncotarget 2017, 8, 59950–59964. [Google Scholar] [CrossRef] [PubMed]
- De Carlo, F.; Witte, T.R.; Hardman, W.E.; Claudio, P.P. Omega-3 eicosapentaenoic acid decreases CD133 colon cancer stem-like cell marker expression while increasing sensitivity to chemotherapy. PLoS ONE 2013, 16, e69760. [Google Scholar] [CrossRef] [PubMed]
- Rudra, P.K.; Krokan, H.E. Cell-specific enhancement of doxorubicin toxicity in human tumour cells by docosahexaenoic acid. Anticancer Res. 2001, 21, 29–38. [Google Scholar] [PubMed]
- Hardman, W.E.; Avula, C.P.; Fernandes, G.; Cameron, I.L. Three percent dietary fish oil concentrate increased efficacy of doxorubicin against MDA-MB 231 breast cancer xenografts. Clin. Cancer Res. 2001, 7, 2041–2049. [Google Scholar] [PubMed]
- Burns, C.P.; Luttenegger, D.G.; Dudley, D.T.; Buettner, G.R.; Spector, A.A. Effect of modification of plasma membrane fatty acid composition on fluidity and methotrexate transport in L1210 murine leukemia cells. Cancer Res. 1979, 39, 1726–1732. [Google Scholar] [PubMed]
- Guffy, M.M.; North, J.A.; Burns, C.P. Effect of cellular fatty acid alteration on adriamycin sensitivity in cultured L1210 murine leukemia cells. Cancer Res. 1984, 44, 1863–1866. [Google Scholar] [PubMed]
- Zijlstra, J.G.; de Vries, E.G.; Muskiet, F.A.; Martini, I.A.; Timmer-Bosscha, H.; Mulder, N.H. Influence of docosahexaenoic acid in vitro on intracellular adriamycin concentration in lymphocytes and human adriamycin-sensitive and -resistant small-cell lung cancer cell lines, and on cytotoxicity in the tumor cell lines. Int. J. Cancer 1987, 40, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Ikushima, S.; Fujiwara, F.; Todo, S.; Imashuku, S. Effects of polyunsaturated fatty acids on vincristine-resistance in human neuroblastoma cells. Anticancer Res. 1991, 11, 1215–1220. [Google Scholar] [PubMed]
- Das, U.N.; Madhavi, N.; Sravan Kumar, G.; Padma, M.; Sangeetha, P. Can tumour cell drug resistance be reversed by essential fatty acids and their metabolites? Prostaglandins Leukot. Essent. Fatty Acids 1998, 58, 39–54. [Google Scholar] [CrossRef] [PubMed]
- Sturlan, S.; Baumgartner, M.; Roth, E.; Bachleitner-Hofmann, T. Docosahexaenoic acid enhances arsenic trioxide-mediated apoptosis in arsenic trioxide-resistant HL-60 cells. Blood 2003, 101, 4990–4997. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.H.; Schipper, H.M.; Lee, J.S.; Singer, J.; Waxman, S. Mechanisms of action of arsenic trioxide. Cancer Res. 2002, 62, 3893–3903. [Google Scholar] [PubMed]
- Baumgartner, M.; Sturlan, S.; Roth, E.; Wessner, B.; Bachleitner-Hofmann, T. Enhancement of arsenic trioxide-mediated apoptosis using docosahexaenoic acid in arsenic trioxide-resistant solid tumor cells. Int. J. Cancer 2004, 112, 707–712. [Google Scholar] [CrossRef] [PubMed]
- Menendez, J.A.; Lupu, R.; Colomer, R. Exogenous supplementation with omega -3 polyunsaturated fatty acid docosahexaenoic acid (DHA; 22:6n-3) synergistically enhances taxane cytotoxicity and downregulates Her-2/neu (c-erbB-2) oncogene expression in human breast cancer cells. Eur. J. Cancer Prev. 2005, 14, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Timmer-Bosscha, H.; Hospers, G.A.; Meijer, C.; Mulder, N.H.; Muskiet, F.A.; Martini, I.A.; Uges, D.R.; de Vries, E.G. Influence of docosahexaenoic acid on cisplatin resistance in a human small cell lung carcinoma cell line. J. Natl. Cancer Inst. 1989, 81, 1069–1075. [Google Scholar] [CrossRef] [PubMed]
- Allam-Ndoul, B.; Guénard, F.; Barbier, O.; Vohl, M.C. A Study of the Differential Effects of Eicosapentaenoic Acid (EPA) and Docosahexaenoic Acid (DHA) on Gene Expression Profiles of Stimulated Thp-1 Macrophages. Nutrients 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zhu, S.L.; Wu, Y.K.; He, Z.; Chen, Y.Q. Omega-3 free fatty acids attenuate insulin-promoted breast cancer cell proliferation. Nutr. Res. 2017, 42, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Bianchini, F.; Giannoni, E.; Serni, S.; Chiarugi, P.; Calorini, L. 22:6n-3 DHA inhibits differentiation of prostate fibroblasts into myofibroblasts and tumorigenesis. Br. J. Nutr. 2012, 108, 2129–2137. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Jia, X.; Hou, L.; Liu, X.; Gao, Q. Involvement of apoptotic pathways in docosahexaenoic acid-induced benefit in prostate cancer: Pathway-focused gene expression analysis using RT2 Profile PCR Array System. Lipids Health Dis. 2017, 16, 59. [Google Scholar] [CrossRef] [PubMed]
- Quiles, J.L.; Huertas, J.R.; Battino, M.; Mataix, J.; Ramírez-Tortosa, M.C. Antioxidant nutrients and adriamycin toxicity. Toxicology 2002, 180, 79–95. [Google Scholar] [CrossRef] [PubMed]
- Takemura, G.; Fujiwara, H. Doxorubicin-induced cardiomyopathy from the cardiotoxic mechanisms to management. Prog. Cardiovasc. Dis. 2007, 49, 330–352. [Google Scholar] [CrossRef] [PubMed]
- Germain, E.; Bonnet, P.; Aubourg, L.; Grangeponte, M.C.; Chajès, V.; Bougnoux, P. Anthracycline-induced cardiac toxicity is not increased by dietary omega-3 fatty acids. Pharmacol. Res. 2003, 47, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Vibet, S.; Goupille, C.; Bougnoux, P.; Steghens, J.P.; Goré, J.; Mahéo, K. Sensitization by docosahexaenoic acid (DHA) of breast cancer cells to anthracyclines through loss of glutathione peroxidase (GP×1) response. Free Radic. Biol. Med. 2008, 44, 1483–1491. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, Y.; Koh, Y.H.; Park, Y.S.; Fujiwara, N.; Sakiyama, H.; Misonou, Y.; Ookawara, T.; Suzuki, K.; Honke, K.; Taniguchi, N. Oxidative stress caused by inactivation of glutathione peroxidase and adaptive responses. Biol. Chem. 2003, 384, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Dannenberg, A.J.; Altorki, N.K.; Boyle, J.O.; Dang, C.; Howe, L.R.; Weksler, B.B.; Subbaramaiah, K. Cyclo-oxygenase 2: A pharmacological target for the prevention of cancer. Lancet Oncol. 2001, 2, 544–551. [Google Scholar] [CrossRef]
- Hawk, E.T.; Viner, J.L.; Dannenberg, A.; DuBois, R.N. COX-2 in cancer—A player that’s defining the rules. J. Natl. Cancer Inst. 2002, 94, 545–546. [Google Scholar] [CrossRef] [PubMed]
- Negi, A.K.; Kansal, S.; Bhatnagar, A.; Agnihotri, N. Alteration in apoptosis and cell cycle by celecoxib and/or fish oil in 7,12-dimethyl benzene (α) anthracene-induced mammary carcinogenesis. Tumour Biol. 2013, 34, 3753–3764. [Google Scholar] [CrossRef] [PubMed]
- Reddy, B.S.; Patlolla, J.M.; Simi, B.; Wang, S.H.; Rao, C.V. Prevention of colon cancer by low doses of celecoxib, a cyclooxygenase inhibitor, administered in diet rich in omega-3 polyunsaturated fatty acids. Cancer Res. 2005, 65, 8022–8027. [Google Scholar] [CrossRef] [PubMed]
- Patterson, W.L.; Georgel, P.T. Breaking the cycle: The role of ω-3 polyunsaturated fatty acids in inflammation-driven cancers. Biochem. Cell Biol. 2014, 92, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, N.K.; Narayanan, B.A.; Reddy, B.S. A combination of docosahexaenoic acid and celecoxib prevents prostate cancer cell growth in vitro and is associated with modulation of nuclear factor-kappaB, and steroid hormone receptors. Int. J. Oncol. 2005, 26, 785–792. [Google Scholar] [CrossRef] [PubMed]
- Swinnen, J.V.; Verhoeven, G.J. Androgens and the control of lipid metabolism in human prostate cancer cells. Steroid Biochem. Mol. Biol. 1998, 65, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Swinnen, J.V.; Roskams, T.; Joniau, S.; Van Poppel, H.; Oyen, R.; Baert, L.; Heyns, W.; Verhoeven, G. Overexpression of fatty acid synthase is an early and common event in the development of prostate cancer. Int. J. Cancer 2002, 98, 19–22. [Google Scholar] [CrossRef] [PubMed]
- Nakatani, T.; Kim, H.J.; Kaburagi, Y.; Yasuda, K.; Ezaki, O. A low fish oil inhibits SREBP-1 proteolytic cascade, while a high-fish-oil feeding decreases SREBP-1 mRNA in mice liver: Relationship to anti-obesity. J. Lipid Res. 2003, 44, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Schønberg, S.A.; Lundemo, A.G.; Fladvad, T.; Holmgren, K.; Bremseth, H.; Nilsen, A.; Gederaas, O.; Tvedt, K.E.; Egeberg, K.W.; Krokan, H.E. Closely related colon cancer cell lines display different sensitivity to polyunsaturated fatty acids, accumulate different lipid classes and downregulate sterol regulatory element-binding protein 1. FEBS J. 2006, 273, 2749–2765. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.L.; Wilson, I.D.; Teare, J.; Marchesi, J.R.; Nicholson, J.K.; Kinross, J.M. Gut microbiota modulation of chemotherapy efficacy and toxicity. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.N.; Zhu, J.; Pan, W.S.; Shen, S.R.; Shan, W.G.; Das, U.N. Effects of fish oil with a high content of n-3 polyunsaturated fatty acids on mouse gut microbiota. Arch. Med. Res. 2014, 45, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Morland, S.L.; Martins, K.J.B.; Mazurak, V.C. n-3 polyunsaturated fatty acid supplementation during cancer chemotherapy. J. Nutr. Intermed. Metab. 2016, 5, 107–116. [Google Scholar] [CrossRef]
- Corsetto, P.A.; Cremona, A.; Montorfano, G.; Jovenitti, I.E.; Orsini, F.; Arosio, P.; Rizzo, A.M. Chemical-physical changes in cell membrane microdomains of breast cancer cells after omega-3 PUFA incorporation. Cell Biochem. Biophys. 2012, 64, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Guixà-González, R.; Javanainen, M.; Gómez-Soler, M.; Cordobilla, B.; Domingo, J.C.; Sanz, F.; Pastor, M.; Ciruela, F.; Martinez-Seara, H.; Selent, J. Membrane omega-3 fatty acids modulate the oligomerisation kinetics of adenosine A2A and dopamine D2 receptors. Sci. Rep. 2016, 6, 19839. [Google Scholar] [CrossRef] [PubMed]
- Escribá, P.V. Membrane-lipid therapy: A historical perspective of membrane-targeted therapies—From lipid bilayer structure to the pathophysiological regulation of cells. Biochim. Biophys. Acta 2017, 1859 Pt B, 1493–1506. [Google Scholar] [CrossRef]
- Torgersen, M.L.; Klokk, T.I.; Kavaliauskiene, S.; Klose, C.; Simons, K.; Skotland, T.; Sandvig, K. The anti-tumor drug 2-hydroxyoleic acid (Minerval) stimulates signaling and retrograde transport. Oncotarget 2016, 7, 86871–86888. [Google Scholar] [CrossRef] [PubMed]
- Van Meer, G. Dynamic Transbilayer Lipid Asymmetry. Cold Spring Harb. Perspect. Biol. 2011, 3. [Google Scholar] [CrossRef] [PubMed]
- Coskun, U.; Simons, K. Membrane rafting: From apical sorting to phase segregation. FEBS Lett. 2010, 584, 1685–1693. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Harshbarger, W.D.; Robles, O.; Krysiak, J.; Hull, K.G.; Cho, S.W.; Richardson, R.D.; Yang, Y.; Garcia, A.; Spiegelman, L.; et al. A strategy for dual inhibition of the proteasome and fatty acid synthase with belactosin C-orlistat hybrids. Bioorg. Med. Chem. 2017, 25, 2901–2916. [Google Scholar] [CrossRef] [PubMed]
- Hilvo, M.; Denkert, C.; Lehtinen, L.; Müller, B.; Brockmöller, S.; Seppänen-Laakso, T.; Budczies, J.; Bucher, E.; Yetukuri, L.; Castillo, S.; et al. Novel theranostic opportunities offered by characterization of altered membrane lipid metabolism in breast cancer progression. Cancer Res. 2011, 71, 3236–3245. [Google Scholar] [CrossRef] [PubMed]
- Ollila, S.; Hyvonen, M.T.; Vattulainen, I. Polyunsaturation in lipid membranes: Dynamic properties and lateral pressure profiles. J. Phys. Chem. B 2007, 111, 3139–3150. [Google Scholar] [CrossRef] [PubMed]
- Rysman, E.; Brusselmans, K.; Scheys, K.; Timmermans, L.; Derua, R.; Munck, S.; Van Veldhoven, P.P.; Waltregny, D.; Daniëls, VW.; Machiels, J.; et al. De novo lipogenesis protects cancer cells from free radicals and chemotherapeutics by promoting membrane lipid saturation. Cancer Res. 2010, 70, 8117–8126. [Google Scholar] [CrossRef] [PubMed]
- Feller, S.E.; Gawrisch, K. Properties of docosahexaenoic-acid-containing lipids and their influence on the function of rhodopsin. Curr. Opin. Struct. Biol. 2005, 15, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Mereghetti, P.; Corsetto, P.A.; Cremona, A.; Rizzo, A.M.; Doglia, S.M.; Ami, D. A Fourier transform infrared spectroscopy study of cell membrane domain modifications induced by docosahexaenoic acid. Biochim. Biophys. Acta 2014, 1840, 3115–3122. [Google Scholar] [CrossRef] [PubMed]
- Eldho, N.V.; Feller, S.E.; Tristram-Nagle, S.; Polozov, I.V.; Gawrisch, K. Polyunsaturated docosahexaenoic vs. docosapentaenoic acid-differences in lipid matrix properties from the loss of one double bond. J. Am. Chem. Soc. 2003, 125, 6409–6421. [Google Scholar] [CrossRef] [PubMed]
- Rajamoorthi, K.; Petrache, H.I.; McIntosh, T.J.; Brown, M.F. Packing and viscoelasticity of polyunsaturated omega-3 and omega-6 lipid bilayers as seen by 2H NMR and X-ray diffraction. J. Am. Chem. Soc. 2005, 127, 1576–1588. [Google Scholar] [CrossRef] [PubMed]
- Orsini, F.; Cremona, A.; Arosio, P.; Corsetto, PA.; Montorfano, G.; Lascialfari, A.; Rizzo, A.M. Atomic force microscopy imaging of lipid rafts of human breast cancer cells. Biochim. Biophys. Acta 2012, 1818, 2943–2949. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Toomre, D. Lipid rafts and signal transduction. Nat. Rev. Mol. Cell Biol. 2001, 1, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Mollinedo, F.; Gajate, C. Lipid rafts as major platforms for signaling regulation in cancer. Adv. Biol. Regul. 2015, 57, 130–146. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.C.; Park, M.J.; Ye, S.K.; Kim, C.W.; Kim, Y.N. Elevated levels of cholesterol-rich lipid rafts in cancer cells are correlated with apoptosis sensitivity induced by cholesterol-depleting agents. Am. J. Pathol. 2006, 168, 1107–1118. [Google Scholar] [CrossRef] [PubMed]
- Corsetto, P.A.; Montorfano, G.; Zava, S.; Jovenitti, I.E.; Cremona, A.; Berra, B.; Rizzo, A.M. Effects of n-3 PUFAs on breast cancer cells through their incorporation in plasma membrane. Lipids Health Dis. 2011, 10, 73. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, S.R.; Rockett, B.D.; Salameh, M.; Carraway, K. Docosahexaenoic acid modifies the clustering and size of lipid rafts and the lateral organization and surface expression of MHC class I of EL4 cells. J. Nutr. 2009, 139, 1632–1639. [Google Scholar] [CrossRef] [PubMed]
- Gottesman, M.M.; Fojo, T.; Bates, S.E. Multidrug resistance in cancer: Role of ATP-dependent transporters. Nat. Rev. Cancer 2002, 2, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.; Sane, R.; Oberoi, R.; Ohlfest, J.R.; Elmquist, W.F. Delivery of molecularly targeted therapy to malignant glioma, a disease of the whole brain. Expert Rev. Mol. Med. 2011, 13, e17. [Google Scholar] [CrossRef] [PubMed]
- Gelsomino, G.; Corsetto, P.A.; Campia, I.; Montorfano, G.; Kopecka, J.; Castella, B.; Gazzano, E.; Ghigo, D.; Rizzo, A.M.; Riganti, C. Omega-3 fatty acids chemosensitize multidrug resistant colon cancer cells by down-regulating cholesterol synthesis and altering detergent resistant membranes composition. Mol. Cancer 2013, 12, 137–157. [Google Scholar] [CrossRef] [PubMed]
- Wirtitsch, M.; Roth, E.; Bachleitner-Hofmann, T.; Wessner, B.; Sturlan, S. Omega-3 and omega-6 polyunsaturated fatty acids enhance arsenic trioxide efficacy in arsenic trioxide-resistant leukemic and solid tumor cells. Oncol. Res. 2009, 18, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Habermann, N.; Christian, B.; Luckas, B.; Pool-Zobel, B.L.; Lund, E.K.; Glei, M. Effects of fatty acids on metabolism and cell growth of human colon cell lines of different transformation state. Biofactors 2009, 35, 460–467. [Google Scholar] [CrossRef] [PubMed]
- Lavie, Y.; Liscovitch, M. Changes in lipid and protein constituents of rafts and caveolae in multidrug resistant cancer cells and their functional consequences. Glycoconj. J. 2000, 17, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Burns, C.P.; North, J.A. Adriamycin transport and sensitivity in fatty acid-modified leukemia cells. Biochim. Biophys. Acta 1986, 888, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.; Meckling-Gill, K.A. Omega-3 polyunsaturated fatty acids increase purine but not pyrimidine transport in L1210 leukaemia cells. Biochem. J. 1996, 315, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Abulrob, A.N.; Mason, M.; Bryce, R.; Gumbleton, M. The effect of fatty acids and analogues upon intracellular levels of doxorubicin in cells displaying P-glycoprotein mediated multidrug resistance. J. Drug Target. 2000, 8, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Riganti, C.; Voena, C.; Kopecka, J.; Corsetto, P.A.; Montorfano, G.; Enrico, E.; Costamagna, C.; Rizzo, A.M.; Ghigo, D.; Bosia, A. Liposome-encapsulated doxorubicin reverses drug resistance by inhibiting P-glycoprotein in human cancer cells. Mol. Pharm. 2011, 8, 683–700. [Google Scholar] [CrossRef] [PubMed]
- Lavie, Y.; Fiucci, G.; Liscovitch, M. Up-regulation of caveolae and caveolar constituents in multidrug-resistant cancer cells. J. Biol. Chem. 1998, 273, 32380–32383. [Google Scholar] [CrossRef] [PubMed]
- Bacso, Z.; Nagy, H.; Goda, K.; Bene, L.; Fenyvesi, F.; Matkó, J.; Szabó, G. Raft and cytoskeleton associations of an ABC transporter: P-glycoprotein. Cytom. A 2004, 61, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Klappe, K.; Hinrichs, J.W.; Kroesen, B.J.; Sietsma, H.; Kok, K.W. MRP1 and glucosylceramide are coordinately over expressed and enriched in rafts during multidrug resistance acquisition in colon cancer cells. Int. J. Cancer 2004, 110, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Kopecka, J.; Campia, I.; Olivero, P.; Pescarmona, G.; Ghigo, D.; Bosia, A.; Riganti, C. A LDL-masked liposomal-doxorubicin reverses drug resistance in human cancer cells. J. Control. Release 2011, 149, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Meyer dos Santos, S.; Weber, C.C.; Franke, C.; Müller, W.E.; Eckert, G.P. Cholesterol: Coupling between membrane microenvironment and ABC transporter activity. Biochem. Biophys. Res. Commun. 2007, 354, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Fenyvesi, F.; Fenyvesi, E.; Szente, L.; Goda, K.; Bacsó, Z.; Bácskay, I.; Váradi, J.; Kiss, T.; Molnár, E.; Janáky, T.; et al. P-glycoprotein inhibition by membrane cholesterol modulation. Eur. J. Pharm. Sci. 2008, 34, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Riganti, C.; Castella, B.; Kopecka, J.; Campia, I.; Coscia, M.; Pescarmona, G.; Bosia, A.; Ghigo, D.; Massaia, M. Zoledronic acid restores doxorubicin chemosensitivity and immunogenic cell death in multidrug-resistant human cancer cells. PLoS ONE 2013, 8, 60975–66989. [Google Scholar] [CrossRef] [PubMed]
- Duncan, R.E.; El-Sohemy, A.; Archer, M.C. Regulation of HMG-CoA reductase in MCF-7 cells by genistein, EPA, and DHA, alone and in combination with mevastatin. Cancer Lett. 2005, 224, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Jakobsen, C.H.; Størvold, G.L.; Bremseth, H.; Follestad, T.; Sand, K.; Mack, M.; Olsen, K.S.; Lundemo, A.G.; Iversen, J.G.; Krokan, H.E.; et al. DHA induces ER stress and growth arrest in human colon cancer cells: Associations with cholesterol and calcium homeostasis. J. Lipid Res. 2008, 49, 2089–2100. [Google Scholar] [CrossRef] [PubMed]
- Størvold, G.L.; Fleten, K.G.; Olsen, C.G.; Follestad, T.; Krokan, H.E.; Schønberg, S.A. Docosahexaenoic acid activates some SREBP-2 targets independent of cholesterol and ER stress in SW620 colon cancer cells. Lipids 2009, 44, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Kuan, C.Y.; Walker, T.H.; Luo, P.G.; Chen, C.F. Long-chain polyunsaturated fatty acids promote paclitaxel cytotoxicity via inhibition of the MDR1 gene in the human colon cancer Caco-2 cell line. J. Am. Coll. Nutr. 2011, 30, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Haslam, I.S.; Jones, K.; Coleman, T.; Simmons, N.L. Induction of P-glycoprotein expression and function in human intestinal epithelial cells (T84). Biochem. Pharmacol. 2008, 76, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Bentires-Alj, M.; Barbu, V.; Fillet, M.; Chariot, A.; Relic, B.; Jacobs, N.; Gielen, J.; Merville, M.P.; Bours, V. NF-kappaB transcription factor induces drug resistance through MDR1 expression in cancer cells. Oncogene 2003, 22, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Biondo, P.D.; Brindley, D.N.; Sawyer, M.B.; Field, C.J. The potential for treatment with dietary long-chain polyunsaturated n-3 fatty acids during chemotherapy. J. Nutr. Biochem. 2008, 19, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Morrow, C.S.; Peklak-Scott, C.; Bishwokarma, B.; Kute, T.E.; Smitherman, P.K.; Townsend, A.J. Multidrug resistance protein 1 (MRP1, ABCC1) mediates resistance to mitoxantrone via glutathione-dependent drug efflux. Mol. Pharmacol. 2006, 69, 1499–1505. [Google Scholar] [CrossRef] [PubMed]
- Habermann, N.; Lund, E.K.; Pool-Zobel, B.L.; Glei, M. Modulation of gene expression in eicosapentaenoic acid and docosahexaenoic acid treated human colon adenoma cells. Genes Nutr. 2009, 4, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Polimeni, M.; Voena, C.; Kopecka, J.; Riganti, C.; Pescarmona, G.; Bosia, A.; Ghigo, D. Modulation of doxorubicin resistance by the glucose-6-phosphate dehydrogenase activity. Biochem. J. 2011, 439, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Zajdel, A.; Paduszyński, P.; Gruchlik, A.; Głogowska-Ligus, J.; Wilczok, A.; Dzierzewicz, A. Polyunsaturated fatty acids alter expression of genes encoding antioxidant enzymes in A549 cells exposed to doxorubicin. Acta Pol. Pharm. 2010, 67, 696–700. [Google Scholar] [PubMed]
- Hering, J.; Garrean, S.; Dekoj, T.R.; Razzak, A.; Saied, A.; Trevino, J.; Babcock, T.A.; Espat, N.J. Inhibition of proliferation by omega-3 fatty acids in chemoresistant pancreatic cancer cells. Ann. Surg. Oncol. 2007, 14, 3620–3628. [Google Scholar] [CrossRef] [PubMed]
- Kornfeld, S.; Goupille, C.; Vibet, S.; Chevalier, S.; Pinet, A.; Lebeau, J.; Tranquart, F.; Bougnoux, P.; Martel, E.; Maurin, A.; et al. Reducing endothelial NOS activation and interstitial fluid pressure with n-3 PUFA offset tumor chemoresistance. Carcinogenesis 2012, 33, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Colas, S.; Mahéo, K.; Denis, F.; Goupille, C.; Hoinard, C.; Champeroux, P.; Tranquart, F.; Bougnoux, P. Sensitization by dietary docosahexaenoic acid of rat mammary carcinoma to anthracycline: A role for tumor vascularization. Clin. Cancer Res. 2006, 12, 5879–5886. [Google Scholar] [CrossRef] [PubMed]
- Barnés, C.M.; Prox, D.; Christison-Lagay, E.A.; Le, H.D.; Short, S.; Cassiola, F.; Panigrahy, D.; Chaponis, D.; Butterfield, C.; Nehra, D.; et al. Inhibition of neuroblastoma cell proliferation with omega-3 fatty acids and treatment of a murine model of human neuroblastoma using a diet enriched with omega-3 fatty acids in combination with sunitinib. Pediatr. Res. 2012, 71, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Sawyer, M.B.; Field, C.J.; Dieleman, L.A.; Baracos, V.E. Nutritional modulation of antitumor efficacy and diarrhea toxicity related to irinotecan chemotherapy in rats bearing the ward colon tumor. Clin. Cancer Res. 2007, 13, 7146–7154. [Google Scholar] [CrossRef] [PubMed]
- Ogilvie, G.K.; Fettman, M.J.; Mallinckrodt, C.H.; Walton, J.A.; Hansen, R.A.; Davenport, D.J.; Gross, K.L.; Richardson, K.L.; Rogers, Q.; Hand, M.S. Effect of fish oil, arginine, and doxorubicin chemotherapy on remission and survival time for dogs with lymphoma: A double-blind, randomized placebo-controlled study. Cancer 2000, 88, 1916–1928. [Google Scholar] [CrossRef] [PubMed]
- Wynter, M.P.; Russell, S.T.; Tisdale, M.J. Effect of n-3 fatty acids on the antitumour effects of cytotoxic drugs. In Vivo 2004, 18, 543–547. [Google Scholar] [PubMed]
- Xue, H.; Le Roy, S.; Sawyer, M.B.; Field, C.J.; Dieleman, L.A.; Baracos, V.E. Single and combined supplementation of glutamine and n-3 polyunsaturated fatty acids on host tolerance and tumour response to 7-ethyl-10-[4-(1-piperidino)-1-piperidino]carbonyloxy-camptothecin (CPT-11)/5-fluorouracil chemotherapy in rats bearing Ward colon tumour. Br. J. Nutr. 2009, 102, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Takaku, T.; Nakajima, S.; Okuda, H. Effects of carp and tuna oils on 5-fluorouracil-induced antitumor activity and side effects in sarcoma 180-bearing mice. Lipids 2001, 36, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Pardini, L.; Pardini, R.S. Intervention of transplantable human mammary carcinoma MX-1 chemotherapy with dietary menhaden oil in athymic mice: Increased therapeutic effects and decreased toxicity of cyclophosphamide. Nutr. Cancer 1997, 28, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Bougnoux, P.; Germain, E.; Chajès, V.; Hubert, B.; Lhuillery, C.; Le Floch, O.; Body, G.; Calais, G. Cytotoxic drugs efficacy correlates with adipose tissue docosahexaenoic acid level in locally advanced breast carcinoma. Br. J. Cancer 1999, 79, 1765–1769. [Google Scholar] [CrossRef] [PubMed]
- Bougnoux, P.; Hajjaji, N.; Ferrasson, M.N.; Giraudeau, B.; Couet, C.; Le Floch, O. Improving outcome of chemotherapy of metastatic breast cancer by docosahexaenoic acid: A phase II trial. Br. J. Cancer 2009, 101, 1978–1985. [Google Scholar] [CrossRef] [PubMed]
- Murphy, R.A.; Mourtzakis, M.; Chu, Q.S.; Baracos, V.E.; Reiman, T.; Mazurak, V.C. Supplementation with fish oil increases first-line chemotherapy efficacy in patients with advanced nonsmall cell lung cancer. Cancer 2011, 117, 3774–3780. [Google Scholar] [CrossRef] [PubMed]
- Patterson, A.C.; Stark, K.D. Direct determinations ofthe fatty acid composition of daily dietary intakes incorporating nutraceuticals and functional food strategies to increase n-3 highly unsaturated fatty acids. J. Am. Coll. Nutr. 2008, 27, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Wolff, A.C.; Donehower, R.C.; Carducci, M.K.; Brahmer, J.R.; Zabelina, Y.; Bradley, M.O.; Anthony, F.H.; Swindell, C.S.; Witman, P.A.; Webb, N.L.; et al. Phase I study of docosahexaenoic acid-paclitaxel: A taxane-fatty acid conjugate with a unique pharmacology and toxicity profile. Clin. Cancer Res. 2003, 9, 3589–3597. [Google Scholar] [PubMed]
- Fracasso, P.M.; Picus, J.; Wildi, J.D.; Goodner, S.A.; Creekmore, A.N.; Gao, F.; Govindan, R.; Ellis, M.J.; Tan, B.R.; Linette, G.P.; et al. Phase 1 and pharmacokinetic study of weekly docosahexaenoic acid-paclitaxel, Taxoprexin, in resistant solid tumor malignancies. Cancer Chemother. Pharmacol. 2009, 63, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Bradley, M.O.; Swindell, C.S.; Anthony, F.H.; Witman, P.A.; Devanesan, P.; Webb, N.L.; Baker, S.D.; Wolff, A.C.; Donehower, R.C. Tumor targeting by conjugation of DHA to paclitaxel. J. Control. Release 2001, 74, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Raghu Nadhanan, R.; Skinner, J.; Chung, R.; Su, Y.W.; Howe, P.R.; Xian, C.J. Supplementation with fish oil and genistein, individually or in combination, protects bone against the adverse effects of methotrexate chemotherapy in rats. PLoS ONE 2013, 8, 71592–71604. [Google Scholar] [CrossRef] [PubMed]
- Schjøtt, J.; Brurok, H.; Jynge, P.; Bjerve, K.S. Effects of eicosapentaenoic acid and docosahexaenoic acid diet supplement on tolerance to the cardiotoxicity of epirubicin and to ischaemia reperfusion in the isolated rat heart. Pharmacol. Toxicol. 1996, 79, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Hajjaji, N.; Bougnoux, P. Selective sensitization of tumors to chemotherapy by marine-derived lipids: A review. Cancer Treat. Rev. 2013, 39, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.S.; Nagawa, H.; Muto, T. Differential effects of polyunsaturated fatty acids on chemosensitivity of NIH3T3 cells and its transformants. Int. J. Cancer 1997, 70, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Pluchino, K.; Hall, M.D.; Goldsborough, A.S.; Callaghan, R.; Gottesman, M.M. Collateral sensitivity as a strategy against cancer multidrug resistance. Drug Resist. Updates 2012, 15, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Roodhart, J.M.; Daenen, L.G.; Stigter, E.C.; Prins, H.J.; Gerrits, J.; Houthuijzen, J.M.; Gerritsen, M.G.; Schipper, H.S.; Backer, M.J.; van Amersfoort, M.; et al. Mesenchymal stem cells induce resistance to chemotherapy through the release of platinum-induced fatty acids. Cancer Cell 2011, 20, 370–383. [Google Scholar] [CrossRef] [PubMed]
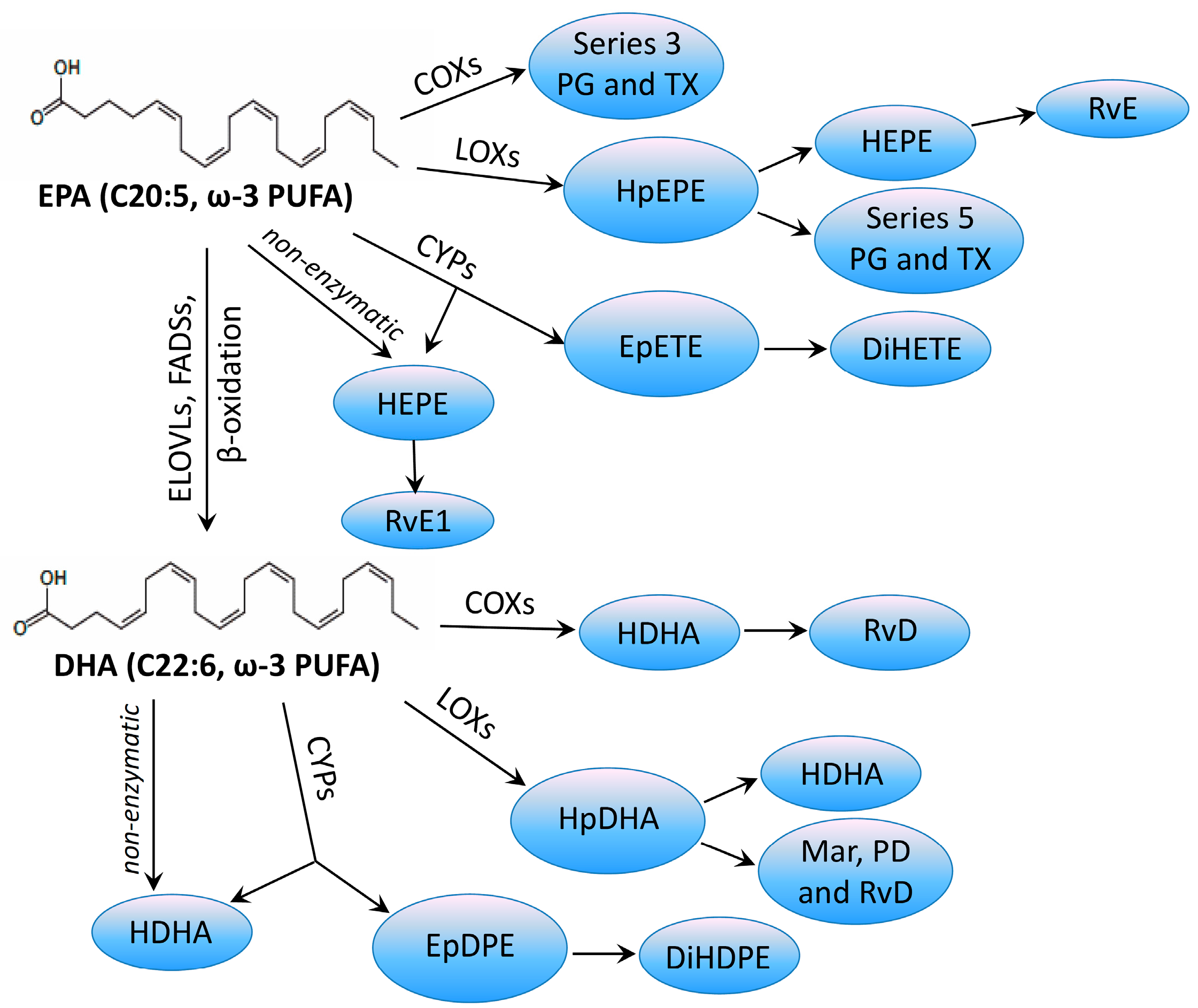
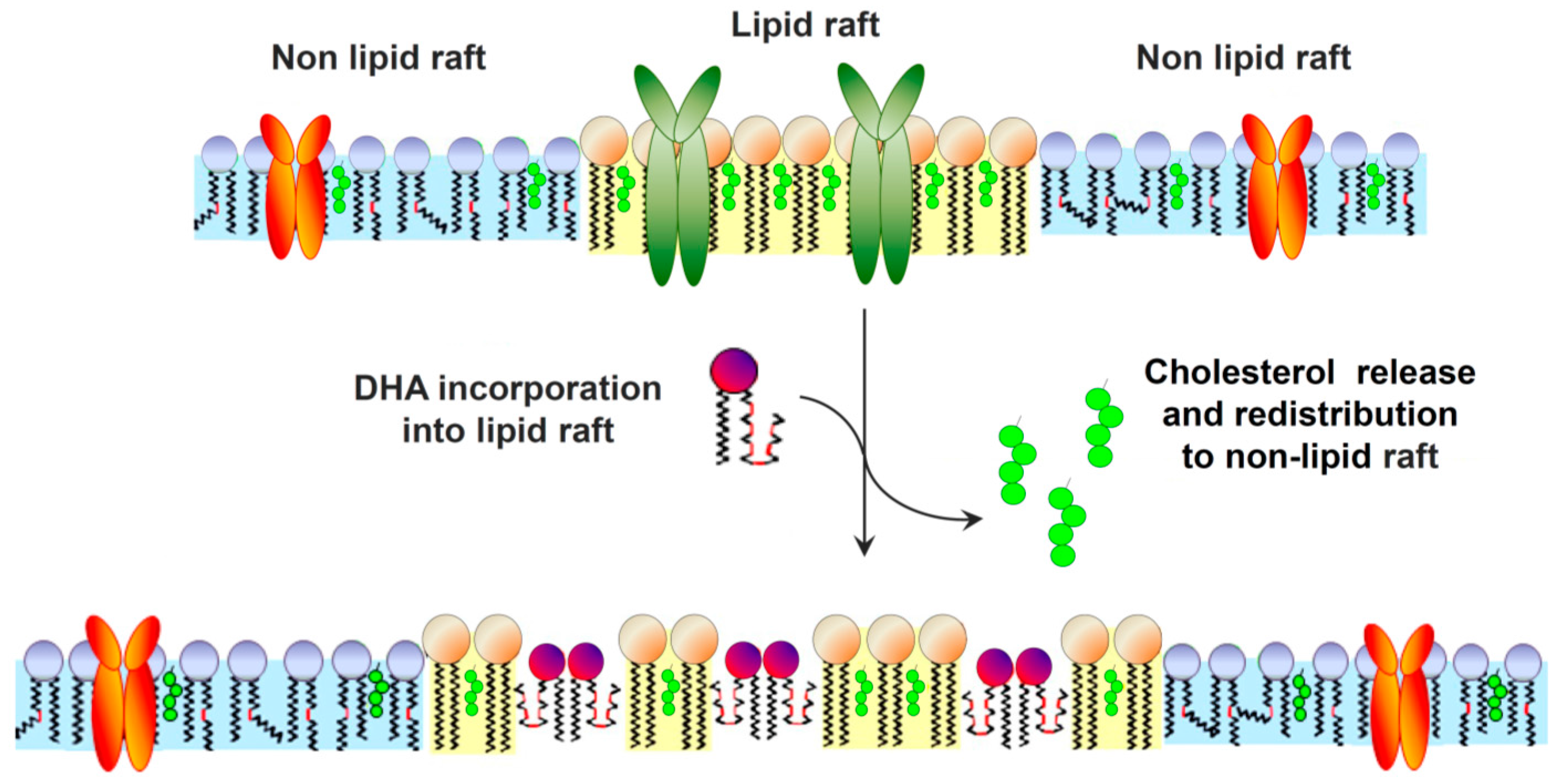
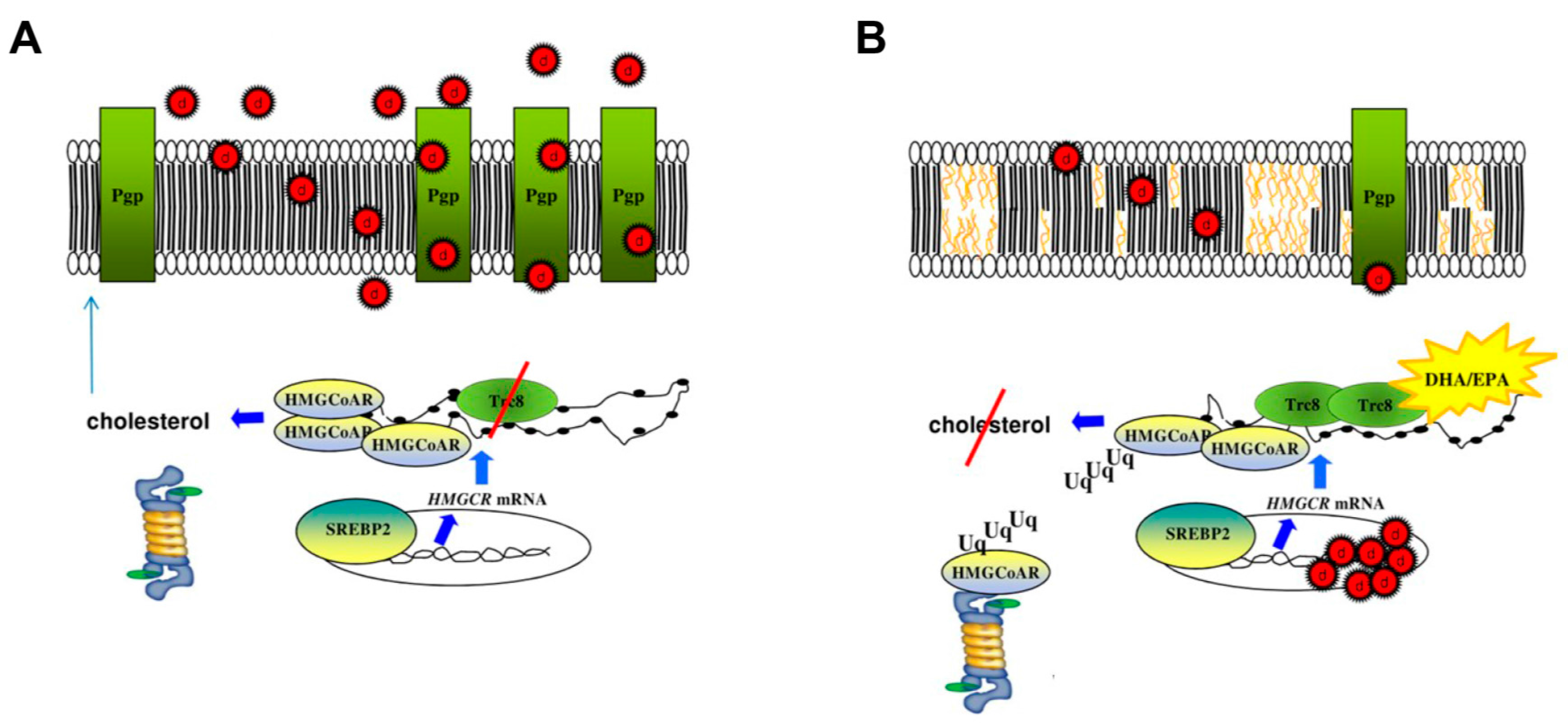
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corsetto, P.A.; Colombo, I.; Kopecka, J.; Rizzo, A.M.; Riganti, C. ω-3 Long Chain Polyunsaturated Fatty Acids as Sensitizing Agents and Multidrug Resistance Revertants in Cancer Therapy. Int. J. Mol. Sci. 2017, 18, 2770. https://doi.org/10.3390/ijms18122770
Corsetto PA, Colombo I, Kopecka J, Rizzo AM, Riganti C. ω-3 Long Chain Polyunsaturated Fatty Acids as Sensitizing Agents and Multidrug Resistance Revertants in Cancer Therapy. International Journal of Molecular Sciences. 2017; 18(12):2770. https://doi.org/10.3390/ijms18122770
Chicago/Turabian StyleCorsetto, Paola Antonia, Irma Colombo, Joanna Kopecka, Angela Maria Rizzo, and Chiara Riganti. 2017. "ω-3 Long Chain Polyunsaturated Fatty Acids as Sensitizing Agents and Multidrug Resistance Revertants in Cancer Therapy" International Journal of Molecular Sciences 18, no. 12: 2770. https://doi.org/10.3390/ijms18122770
APA StyleCorsetto, P. A., Colombo, I., Kopecka, J., Rizzo, A. M., & Riganti, C. (2017). ω-3 Long Chain Polyunsaturated Fatty Acids as Sensitizing Agents and Multidrug Resistance Revertants in Cancer Therapy. International Journal of Molecular Sciences, 18(12), 2770. https://doi.org/10.3390/ijms18122770