hCG: Biological Functions and Clinical Applications

 ,
, 

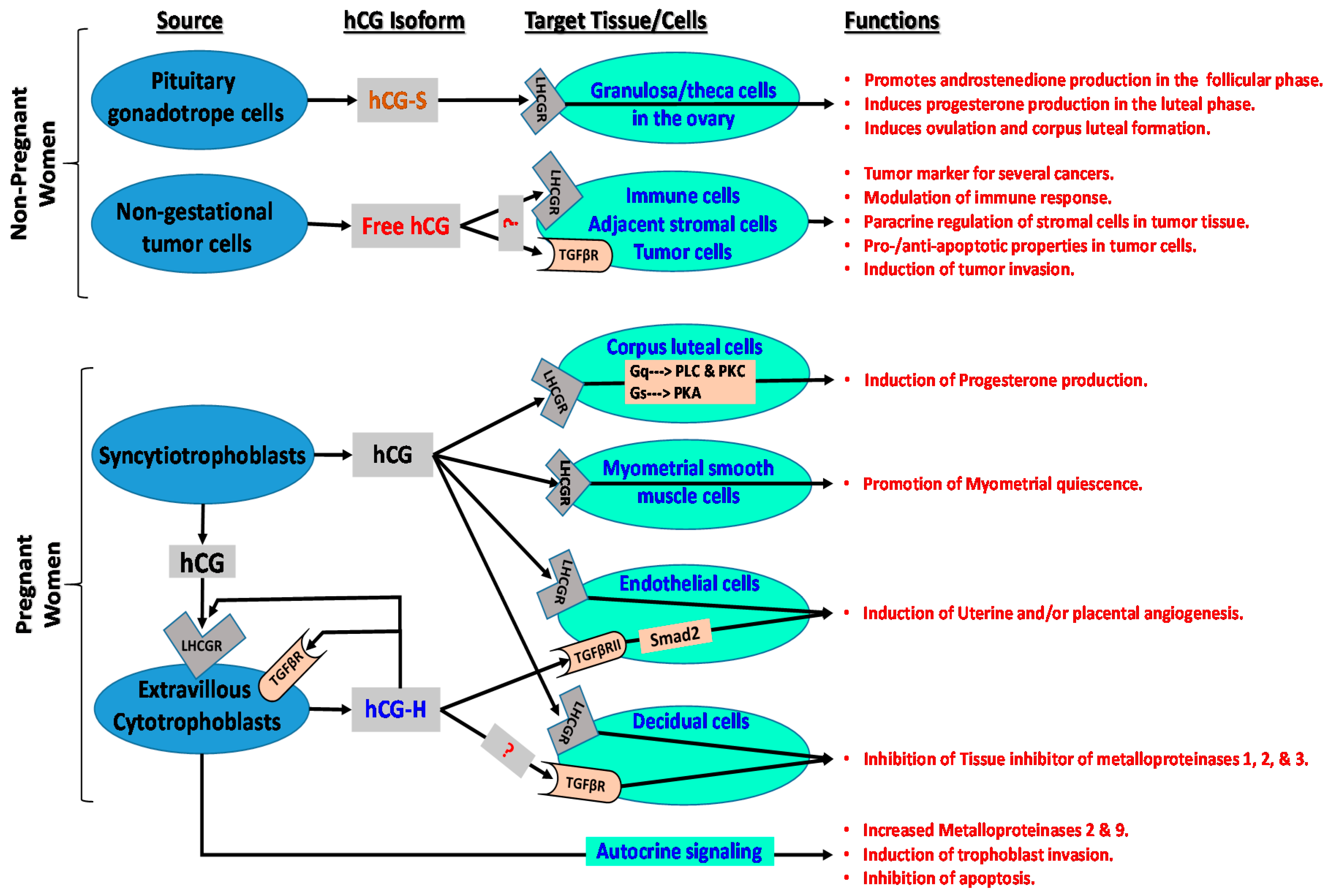{kind=link}
Abstract
1. Introduction
2. Structure, Isoforms, and Genes of Human Chorionic Gonadotropin (hCG)
3. Role of hCG in Embryo Implantation and Trophoblast Invasion
4. Metabolism of hCG
5. Measurements of hCG Levels
6. Clinical Applications of hCG
6.1. hCG Measurements in Normal and Abnormal Pregnancies
6.2. hCG, a Potential Biomarker for Preeclampsia
6.3. hCG, a Serum Marker for Down’s Syndrome Screening
6.4. hCG Is Crucial in the Diagnosis of Gestational Trophoblastic Disease
6.5. hCG Use in Assisted Reproductive Technology
7. Conclusions
Author Contributions
Conflicts of Interest
References
- Fournier, T.; Guibourdenche, J.; Evain-Brion, D. Review: hCGs: Different sources of production, different glycoforms and functions. Placenta 2015, 36, S60–S65. [Google Scholar] [CrossRef] [PubMed]
- Hay, D.L.; Lopata, A. Chorionic gonadotropin secretion by human embryos in vitro. J. Clin. Endocrinol. Metab. 1988, 67, 1322–1324. [Google Scholar] [CrossRef] [PubMed]
- Hay, D.L. Placental histology and the production of human choriogonadotrophin and its subunits in pregnancy. Br. J. Obstet. Gynaecol. 1988, 95, 1268–1275. [Google Scholar] [CrossRef] [PubMed]
- Zygmunt, M.; Herr, F.; Keller-Schoenwetter, S.; Kunzi-Rapp, K.; Munstedt, K.; Rao, C.V.; Lang, U.; Preissner, K.T. Characterization of human chorionic gonadotropin as a novel angiogenic factor. J. Clin. Endocrinol. Metab. 2002, 87, 5290–5296. [Google Scholar] [CrossRef] [PubMed]
- Ambrus, G.; Rao, C.V. Novel regulation of pregnant human myometrial smooth muscle cell gap junctions by human chorionic gonadotropin. Endocrinology 1994, 135, 2772–2779. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, A.; Heinze, K.; Witte, J.; Poloski, E.; Linzke, N.; Woidacki, K.; Zenclussen, A.C. Human chorionic gonadotropin as a central regulator of pregnancy immune tolerance. J. Immunol. 2013, 190, 2650–2658. [Google Scholar] [CrossRef] [PubMed]
- Bansal, A.S.; Bora, S.A.; Saso, S.; Smith, J.R.; Johnson, M.R.; Thum, M.Y. Mechanism of human chorionic gonadotrophin-mediated immunomodulation in pregnancy. Expert Rev. Clin. Immunol. 2012, 8, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Nisula, B.C.; Blithe, D.L.; Akar, A.; Lefort, G.; Wehmann, R.E. Metabolic fate of human choriogonadotropin. J. Steroid Biochem. 1989, 33, 733–737. [Google Scholar] [CrossRef]
- Cole, L.A. Immunoassay of human chorionic gonadotropin, its free subunits, and metabolites. Clin. Chem. 1997, 43, 2233–2243. [Google Scholar] [PubMed]
- Stenman, U.H.; Tiitinen, A.; Alfthan, H.; Valmu, L. The classification, functions and clinical use of different isoforms of HCG. Hum. Reprod. Update 2006, 12, 769–784. [Google Scholar] [CrossRef] [PubMed]
- Lapthorn, A.J.; Harris, D.C.; Littlejohn, A.; Lustbader, J.W.; Canfield, R.E.; Machin, K.J.; Morgan, F.J.; Isaacs, N.W. Crystal structure of human chorionic gonadotropin. Nature 1994, 369, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Stenman, U.H.; Alfthan, H. Determination of human chorionic gonadotropin. Best Pract. Res. Clin. Endocrinol. Metab. 2013, 27, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Pierce, J.G.; Parsons, T.F. Glycoprotein hormones: Structure and function. Annu. Rev. Biochem. 1981, 50, 465–495. [Google Scholar] [CrossRef] [PubMed]
- Cole, L.A. hCG, the wonder of today’s science. Reprod. Biol. Endocrinol. 2012, 10, 24. [Google Scholar] [CrossRef] [PubMed]
- Jameson, J.L.; Hollenberg, A.N. Regulation of chorionic gonadotropin gene expression. Endocr. Rev. 1993, 14, 203–221. [Google Scholar] [PubMed]
- Dufau, M.L. The luteinizing hormone receptor. Ann. Rev. Physiol. 1998, 60, 461–496. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, J.F.; Birken, S.; Lustbader, J.W.; Krichevsky, A.; Chen, Y.; Canfield, R.E. Recent advances in the chemistry and immunochemistry of human chorionic gonadotropin: Impact on clinical measurements. Endocr. Rev. 1994, 15, 650–683. [Google Scholar] [PubMed]
- Kovalevskaya, G.; Birken, S.; Kakuma, T.; Ozaki, N.; Sauer, M.; Lindheim, S.; Cohen, M.; Kelly, A.; Schlatterer, J.; O’Connor, J.F. Differential expression of human chorionic gonadotropin (hCG) glycosylation isoforms in failing and continuing pregnancies: Preliminary characterization of the hyperglycosylated hCG epitope. J. Endocrinol. 2002, 172, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Kovalevskaya, G.; Genbacev, O.; Fisher, S.J.; Caceres, E.; O’Connor, J.F. Trophoblast origin of hCG isoforms: Cytotrophoblasts are the primary source of choriocarcinoma-like hCG. Mol. Cell. Endocrinol. 2002, 194, 147–155. [Google Scholar] [CrossRef]
- Cole, L.A. hCG, five independent molecules. Clin. Chim. Acta Int. J. Clin. Chem. 2012, 413, 48–65. [Google Scholar] [CrossRef] [PubMed]
- Berndt, S.; Blacher, S.; Munaut, C.; Detilleux, J.; Perrier d’Hauterive, S.; Huhtaniemi, I.; Evain-Brion, D.; Noel, A.; Fournier, T.; Foidart, J.M. Hyperglycosylated human chorionic gonadotropin stimulates angiogenesis through TGF-β receptor activation. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2013, 27, 1309–1321. [Google Scholar] [CrossRef] [PubMed]
- Elliott, M.M.; Kardana, A.; Lustbader, J.W.; Cole, L.A. Carbohydrate and peptide structure of the α- and β-subunits of human chorionic gonadotropin from normal and aberrant pregnancy and choriocarcinoma. Endocrine 1997, 7, 15–32. [Google Scholar] [CrossRef] [PubMed]
- Birken, S.; Yershova, O.; Myers, R.V.; Bernard, M.P.; Moyle, W. Analysis of human choriogonadotropin core 2 o-glycan isoforms. Mol. Cell. Endocrinol. 2003, 204, 21–30. [Google Scholar] [CrossRef]
- Fiddes, J.C.; Goodman, H.M. The gene encoding the common alpha subunit of the four human glycoprotein hormones. J. Mol. Appl. Genet. 1981, 1, 3–18. [Google Scholar] [PubMed]
- Boorstein, W.R.; Vamvakopoulos, N.C.; Fiddes, J.C. Human chorionic gonadotropin β-subunit is encoded by at least eight genes arranged in tandem and inverted pairs. Nature 1982, 300, 419–422. [Google Scholar] [CrossRef] [PubMed]
- Rull, K.; Hallast, P.; Uuskula, L.; Jackson, J.; Punab, M.; Salumets, A.; Campbell, R.K.; Laan, M. Fine-scale quantification of HCG bata gene transcription in human trophoblastic and non-malignant non-trophoblastic tissues. Mol. Hum. Reprod. 2008, 14, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Hallast, P.; Rull, K.; Laan, M. The evolution and genomic landscape of CGB1 and CGB2 genes. Mol. Cell. Endocrinol. 2007, 260–262, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Knofler, M. What factors regulate HCG production in Down’s syndrome pregnancies? Regulation of HCG during normal gestation and in pregnancies affected by Down’s syndrome. Mol. Hum. Reprod. 1999, 5, 895–897. [Google Scholar] [CrossRef] [PubMed]
- Handschuh, K.; Guibourdenche, J.; Cocquebert, M.; Tsatsaris, V.; Vidaud, M.; Evain-Brion, D.; Fournier, T. Expression and regulation by PPARgamma of hCG α- and β-subunits: Comparison between villous and invasive extravillous trophoblastic cells. Placenta 2009, 30, 1016–1022. [Google Scholar] [CrossRef] [PubMed]
- Handschuh, K.; Guibourdenche, J.; Tsatsaris, V.; Guesnon, M.; Laurendeau, I.; Evain-Brion, D.; Fournier, T. Human chorionic gonadotropin produced by the invasive trophoblast but not the villous trophoblast promotes cell invasion and is down-regulated by peroxisome proliferator-activated receptor-γ. Endocrinology 2007, 148, 5011–5019. [Google Scholar] [CrossRef] [PubMed]
- Murthi, P.; Kalionis, B.; Cocquebert, M.; Rajaraman, G.; Chui, A.; Keogh, R.J.; Evain-Brion, D.; Fournier, T. Homeobox genes and down-stream transcription factor PPARgamma in normal and pathological human placental development. Placenta 2013, 34, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, A.J.; Baird, D.D.; Weinberg, C.R. Time of implantation of the conceptus and loss of pregnancy. N. Engl. J. Med. 1999, 340, 1796–1799. [Google Scholar] [CrossRef] [PubMed]
- Edelman, G.M.; Crossin, K.L. Cell adhesion molecules: Implications for a molecular histology. Annu. Rev. Biochem. 1991, 60, 155–190. [Google Scholar] [CrossRef] [PubMed]
- Norwitz, E.R.; Schust, D.J.; Fisher, S.J. Implantation and the survival of early pregnancy. N. Engl. J. Med. 2001, 345, 1400–1408. [Google Scholar] [CrossRef] [PubMed]
- Dey, S.K.; Lim, H.; Das, S.K.; Reese, J.; Paria, B.C.; Daikoku, T.; Wang, H. Molecular cues to implantation. Endocr. Rev. 2004, 25, 341–373. [Google Scholar] [CrossRef] [PubMed]
- Genbacev, O.; DiFederico, E.; McMaster, M.; Fisher, S.J. Invasive cytotrophoblast apoptosis in pre-eclampsia. Hum. Reprod. 1999, 14, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Tarrade, A.; Goffin, F.; Munaut, C.; Lai-Kuen, R.; Tricottet, V.; Foidart, J.M.; Vidaud, M.; Frankenne, F.; Evain-Brion, D. Effect of matrigel on human extravillous trophoblasts differentiation: Modulation of protease pattern gene expression. Biol. Reprod. 2002, 67, 1628–1637. [Google Scholar] [CrossRef] [PubMed]
- Gudermann, T.; Birnbaumer, M.; Birnbaumer, L. Evidence for dual coupling of the murine luteinizing hormone receptor to adenylyl cyclase and phosphoinositide breakdown and Ca2+ mobilization. Studies with the cloned murine luteinizing hormone receptor expressed in L cells. J. Biol. Chem. 1992, 267, 4479–4488. [Google Scholar] [PubMed]
- Casarini, L.; Lispi, M.; Longobardi, S.; Milosa, F.; La Marca, A.; Tagliasacchi, D.; Pignatti, E.; Simoni, M. LH and hCG action on the same receptor results in quantitatively and qualitatively different intracellular signalling. PLoS ONE 2012, 7, e46682. [Google Scholar] [CrossRef] [PubMed]
- Maymo, J.L.; Perez Perez, A.; Maskin, B.; Duenas, J.L.; Calvo, J.C.; Sanchez Margalet, V.; Varone, C.L. The alternative Epac/cAMP pathway and the MAPK pathway mediate hCG induction of leptin in placental cells. PLoS ONE 2012, 7, e46216. [Google Scholar] [CrossRef] [PubMed]
- Keryer, G.; Alsat, E.; Tasken, K.; Evain-Brion, D. Cyclic AMP-dependent protein kinases and human trophoblast cell differentiation in vitro. J. Cell Sci. 1998, 111 Pt 7, 995–1004. [Google Scholar] [PubMed]
- Riccetti, L.; Yvinec, R.; Klett, D.; Gallay, N.; Combarnous, Y.; Reiter, E.; Simoni, M.; Casarini, L.; Ayoub, M.A. Human Luteinizing Hormone and Chorionic Gonadotropin Display Biased Agonism at the LH and LH/CG Receptors. Sci. Rep. 2017, 7, 940. [Google Scholar] [CrossRef] [PubMed]
- Resnik, R.; Creasy, R.K.; Iams, J.D.; Lockwood, C.J.; MHCM; Moore, T.; Greene, M.F. Creasy & Resnik’s Maternal-Fetal Medicine: Principles and Practice, 7th ed.; Elsevier Saunders: Philadelphia, PA, USA, 2014. [Google Scholar]
- Salamonsen, L.A.; Evans, J.; Nguyen, H.P.; Edgell, T.A. The Microenvironment of Human Implantation: Determinant of Reproductive Success. Am. J. Reprod. Immunol. 2016, 75, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Paiva, P.; Hannan, N.J.; Hincks, C.; Meehan, K.L.; Pruysers, E.; Dimitriadis, E.; Salamonsen, L.A. Human chorionic gonadotrophin regulates FGF2 and other cytokines produced by human endometrial epithelial cells, providing a mechanism for enhancing endometrial receptivity. Hum. Reprod. 2011, 26, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.; Catalano, R.D.; Brown, P.; Sherwin, R.; Critchley, H.O.; Fazleabas, A.T.; Jabbour, H.N. Prokineticin 1 mediates fetal-maternal dialogue regulating endometrial leukemia inhibitory factor. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2009, 23, 2165–2175. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.L.; Kaspar, P.; Brunet, L.J.; Bhatt, H.; Gadi, I.; Kontgen, F.; Abbondanzo, S.J. Blastocyst implantation depends on maternal expression of leukaemia inhibitory factor. Nature 1992, 359, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Evans, J. Hyperglycosylated hCG: A Unique Human Implantation and Invasion Factor. Am. J. Reprod. Immunol. 2016, 75, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Lindhard, A.; Bentin-Ley, U.; Ravn, V.; Islin, H.; Hviid, T.; Rex, S.; Bangsboll, S.; Sorensen, S. Biochemical evaluation of endometrial function at the time of implantation. Fertil. Steril. 2002, 78, 221–233. [Google Scholar] [CrossRef]
- Guibourdenche, J.; Handschuh, K.; Tsatsaris, V.; Gerbaud, P.; Leguy, M.C.; Muller, F.; Brion, D.E.; Fournier, T. Hyperglycosylated hCG is a marker of early human trophoblast invasion. J. Clin. Endocrinol. Metab. 2010, 95, E240–E244. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.; Salamonsen, L.A.; Menkhorst, E.; Dimitriadis, E. Dynamic changes in hyperglycosylated human chorionic gonadotrophin throughout the first trimester of pregnancy and its role in early placentation. Hum. Reprod. 2015, 30, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Koistinen, H.; Hautala, L.; Koli, K.; Stenman, U.H. Absence of TGF-β Receptor Activation by Highly Purified hCG Preparations. Mol. Endocrinol. 2015, 29, 1787–1791. [Google Scholar] [CrossRef] [PubMed]
- Cole, L.A.; Butler, S. Hyperglycosylated hCG, hCGβ and Hyperglycosylated hCGβ: Interchangeable cancer promoters. Mol. Cell. Endocrinol. 2012, 349, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Iles, R.K.; Delves, P.J.; Butler, S.A. Does hCG or hCGβ play a role in cancer cell biology? Mol. Cell. Endocrinol. 2010, 329, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Lehnert, S.A.; Akhurst, R.J. Embryonic expression pattern of TGF beta type-1 RNA suggests both paracrine and autocrine mechanisms of action. Development 1988, 104, 263–273. [Google Scholar] [PubMed]
- Fluhr, H.; Bischof-Islami, D.; Krenzer, S.; Licht, P.; Bischof, P.; Zygmunt, M. Human chorionic gonadotropin stimulates matrix metalloproteinases-2 and -9 in cytotrophoblastic cells and decreases tissue inhibitor of metalloproteinases-1, -2, and -3 in decidualized endometrial stromal cells. Fertil. Steril. 2008, 90, 1390–1395. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, A.; Brachwitz, N.; Sohr, S.; Engeland, K.; Langwisch, S.; Dolaptchieva, M.; Alexander, T.; Taran, A.; Malfertheiner, S.F.; Costa, S.D.; et al. Human chorionic gonadotropin attracts regulatory T cells into the fetal-maternal interface during early human pregnancy. J. Immunol. 2009, 182, 5488–5497. [Google Scholar] [CrossRef] [PubMed]
- Wehmann, R.E.; Nisula, B.C. Characterization of a discrete degradation product of the human chorionic gonadotropin β-subunit in humans. J. Clin. Endocrinol. Metab. 1980, 51, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Norman, R.J.; Menabawey, M.; Lowings, C.; Buck, R.H.; Chard, T. Relationship between blood and urine concentrations of intact human chorionic gonadotropin and its free subunits in early pregnancy. Obstet. Gynecol. 1987, 69, 590–593. [Google Scholar] [PubMed]
- Wehmann, R.E.; Nisula, B.C. Metabolic and renal clearance rates of purified human chorionic gonadotropin. J. Clin. Investig. 1981, 68, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, J.; Alfthan, H.; Ylostalo, P.; Veldhuis, J.; Stenman, U.H. Disappearance of human chorionic gonadotropin and its α- and β-subunits after term pregnancy. Clin. Chem. 1997, 43, 2155–2163. [Google Scholar] [PubMed]
- Rizkallah, T.; Gurpide, E.; Vande Wiele, R.L. Metabolism of HCG in man. J. Clin. Endocrinol. Metab. 1969, 29, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Blithe, D.L.; Nisula, B.C. Similarity of the clearance rates of free α-subunit and α-subunit dissociated from intact human chorionic gonadotropin, despite differences in sialic acid contents. Endocrinology 1987, 121, 1215–1220. [Google Scholar] [CrossRef] [PubMed]
- Lambert, A.; Talbot, J.A.; Anobile, C.J.; Robertson, W.R. Gonadotrophin heterogeneity and biopotency: Implications for assisted reproduction. Mol. Hum. Reprod. 1998, 4, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Southers, J.L.; Cassels, J.W., Jr.; Banks, S.M.; Wehmann, R.E.; Blithe, D.L.; Chen, H.C.; Nisula, B.C. Structure-kinetic relationships of choriogonadotropin and related molecules. Am. J. Physiol. 1989, 256 Pt 6, E721–E724. [Google Scholar] [PubMed]
- Rosa, C.; Amr, S.; Birken, S.; Wehmann, R.; Nisula, B. Effect of desialylation of human chorionic gonadotropin on its metabolic clearance rate in humans. J. Clin. Endocrinol. Metab. 1984, 59, 1215–1219. [Google Scholar] [CrossRef] [PubMed]
- Vaitukaitis, J.L.; Braunstein, G.D.; Ross, G.T. A radioimmunoassay which specifically measures human chorionic gonadotropin in the presence of human luteinizing hormone. Am. J. Obstet. Gynecol. 1972, 113, 751–758. [Google Scholar] [CrossRef]
- Montagnana, M.; Trenti, T.; Aloe, R.; Cervellin, G.; Lippi, G. Human chorionic gonadotropin in pregnancy diagnostics. Clin. Chim. Acta Int. J. Clin. Chem. 2011, 412, 1515–1520. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, P.H.; Moustafa, Z.A.; Krichevsky, A.; Birken, S.; Armstrong, E.G.; Canfield, R.E. Characterization and relative orientation of epitopes for monoclonal antibodies and antisera to human chorionic gonadotropin. Am. J. Reprod. Immunol. Microbiol. 1985, 8, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Bidart, J.M.; Ozturk, M.; Bellet, D.H.; Jolivet, M.; Gras-Masse, H.; Troalen, F.; Bohuon, C.J.; Wands, J.R. Identification of epitopes associated with hCG and the beta hCG carboxyl terminus by monoclonal antibodies produced against a synthetic peptide. J. Immunol. 1985, 134, 457–464. [Google Scholar] [PubMed]
- Norman, R.J.; Poulton, T.; Gard, T.; Chard, T. Monoclonal antibodies to human chorionic gonadotropin: Implications for antigenic mapping, immunoradiometric assays, and clinical applications. J. Clin. Endocrinol. Metab. 1985, 61, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Berger, P.; Wick, G. The antigenic surface of human chorionic gonadotropin as mapped by murine monoclonal antibodies. Endocrinology 1986, 118, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Alfthan, H.; Haglund, C.; Dabek, J.; Stenman, U.H. Concentrations of human choriogonadotropin, its β-subunit, and the core fragment of the β-subunit in serum and urine of men and nonpregnant women. Clin. Chem. 1992, 38, 1981–1987. [Google Scholar] [PubMed]
- Berger, P.; Sturgeon, C.; Bidart, J.M.; Paus, E.; Gerth, R.; Niang, M.; Bristow, A.; Birken, S.; Stenman, U.H. The ISOBM TD-7 Workshop on hCG and related molecules. Towards user-oriented standardization of pregnancy and tumor diagnosis: Assignment of epitopes to the three-dimensional structure of diagnostically and commercially relevant monoclonal antibodies directed against human chorionic gonadotropin and derivatives. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2002, 23, 1–38. [Google Scholar]
- Cole, L.A.; Butler, S. Detection of hCG in trophoblastic disease. The USA hCG reference service experience. J. Reprod. Med. 2002, 47, 433–444. [Google Scholar] [PubMed]
- McChesney, R.; Wilcox, A.J.; O’Connor, J.F.; Weinberg, C.R.; Baird, D.D.; Schlatterer, J.P.; McConnaughey, D.R.; Birken, S.; Canfield, R.E. Intact HCG, free HCG beta subunit and HCG beta core fragment: Longitudinal patterns in urine during early pregnancy. Hum. Reprod. 2005, 20, 928–935. [Google Scholar] [CrossRef] [PubMed]
- Cole, L.A. Familial HCG syndrome. J. Reprod. Immunol. 2012, 93, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Stenman, U.H.; Alfthan, H.; Ranta, T.; Vartiainen, E.; Jalkanen, J.; Seppala, M. Serum levels of human chorionic gonadotropin in nonpregnant women and men are modulated by gonadotropin-releasing hormone and sex steroids. J. Clin. Endocrinol. Metab. 1987, 64, 730–736. [Google Scholar] [CrossRef] [PubMed]
- Cole, L.A.; Sasaki, Y.; Muller, C.Y. Normal production of human chorionic gonadotropin in menopause. N. Engl. J. Med. 2007, 356, 1184–1186. [Google Scholar] [CrossRef] [PubMed]
- Poikkeus, P.; Hiilesmaa, V.; Tiitinen, A. Serum HCG 12 days after embryo transfer in predicting pregnancy outcome. Hum. Reprod. 2002, 17, 1901–1905. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, J.F.; Ellish, N.; Kakuma, T.; Schlatterer, J.; Kovalevskaya, G. Differential urinary gonadotrophin profiles in early pregnancy and early pregnancy loss. Prenat. Diagn. 1998, 18, 1232–1240. [Google Scholar] [CrossRef]
- Wilcox, A.J.; Weinberg, C.R.; O’Connor, J.F.; Baird, D.D.; Schlatterer, J.P.; Canfield, R.E.; Armstrong, E.G.; Nisula, B.C. Incidence of early loss of pregnancy. N. Engl. J. Med. 1988, 319, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, J.; Stenman, U.H.; Ylostalo, P. Serum human chorionic gonadotropin dynamics during spontaneous resolution of ectopic pregnancy. Fertil. Steril. 1994, 61, 632–636. [Google Scholar] [CrossRef]
- Barnhart, K.T.; Sammel, M.D.; Rinaudo, P.F.; Zhou, L.; Hummel, A.C.; Guo, W. Symptomatic patients with an early viable intrauterine pregnancy: HCG curves redefined. Obstet. Gynecol. 2004, 104, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Seeber, B.E.; Sammel, M.D.; Guo, W.; Zhou, L.; Hummel, A.; Barnhart, K.T. Application of redefined human chorionic gonadotropin curves for the diagnosis of women at risk for ectopic pregnancy. Fertil. Steril. 2006, 86, 454–459. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.; Sammel, M.D.; Zhou, L.; Gracia, C.; Hummel, A.C.; Barnhart, K. Human chorionic gonadotropin profile for women with ectopic pregnancy. Obstet. Gynecol. 2006, 107, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, J.; Tiitinen, A.; Alfthan, H.; Ylostalo, P.; Stenman, U.H. Ectopic pregnancy after in vitro fertilization is characterized by delayed implantation but a normal increase of serum human chorionic gonadotrophin and its subunits. Hum. Reprod. 1996, 11, 2750–2757. [Google Scholar] [CrossRef] [PubMed]
- Norris, W.; Nevers, T.; Sharma, S.; Kalkunte, S. Review: hCG, preeclampsia and regulatory T cells. Placenta 2011, 32, S182–S185. [Google Scholar] [CrossRef] [PubMed]
- Lockwood, C.J.; Huang, S.J.; Krikun, G.; Caze, R.; Rahman, M.; Buchwalder, L.F.; Schatz, F. Decidual hemostasis, inflammation, and angiogenesis in pre-eclampsia. Semin. Thromb. Hemost. 2011, 37, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Roberge, S.; Giguere, Y.; Villa, P.; Nicolaides, K.; Vainio, M.; Forest, J.C.; von Dadelszen, P.; Vaiman, D.; Tapp, S.; Bujold, E. Early administration of low-dose aspirin for the prevention of severe and mild preeclampsia: A systematic review and meta-analysis. Am. J. Perinatol. 2012, 29, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Roberge, S.; Villa, P.; Nicolaides, K.; Giguere, Y.; Vainio, M.; Bakthi, A.; Ebrashy, A.; Bujold, E. Early administration of low-dose aspirin for the prevention of preterm and term preeclampsia: A systematic review and meta-analysis. Fetal Diagn. Ther. 2012, 31, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.P.; Piao, L.; Chen, X.; Yu, J.; Masch, R.; Schatz, F.; Lockwood, C.J.; Huang, S.J. Expression of Interferon gamma by Decidual Cells and Natural Killer Cells at the Human Implantation Site: Implications for Preeclampsia, Spontaneous Abortion, and Intrauterine Growth Restriction. Reprod. Sci. 2015, 22, 1461–1467. [Google Scholar] [CrossRef] [PubMed]
- Funghi, L.; Damiani, F.; Yen, C.F.; Lee, C.L.; Lombardi, A.; Schatz, F.; Lockwood, C.J.; Marcolongo, P.; Petraglia, F.; Arcuri, F. Expression and regulation of 11beta-hydroxysteroid dehydrogenase type 1 in first trimester human decidua cells: Implication in preeclampsia. Mol. Cell. Endocrinol. 2016, 437, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Lockwood, C.J.; Basar, M.; Kayisli, U.A.; Guzeloglu-Kayisli, O.; Murk, W.; Wang, J.; De Paz, N.; Shapiro, J.P.; Masch, R.J.; Semerci, N.; et al. Interferon-gamma protects first-trimester decidual cells against aberrant matrix metalloproteinases 1, 3, and 9 expression in preeclampsia. Am. J. Pathol. 2014, 184, 2549–2559. [Google Scholar] [CrossRef] [PubMed]
- Lockwood, C.J.; Huang, S.J.; Chen, C.P.; Huang, Y.; Xu, J.; Faramarzi, S.; Kayisli, O.; Kayisli, U.; Koopman, L.; Smedts, D.; et al. Decidual cell regulation of natural killer cell-recruiting chemokines: Implications for the pathogenesis and prediction of preeclampsia. Am. J. Pathol. 2013, 183, 841–856. [Google Scholar] [CrossRef] [PubMed]
- Schatz, F.; Guzeloglu-Kayisli, O.; Arlier, S.; Kayisli, U.A.; Lockwood, C.J. The role of decidual cells in uterine hemostasis, menstruation, inflammation, adverse pregnancy outcomes and abnormal uterine bleeding. Hum. Reprod. Update 2016, 22, 497–515. [Google Scholar] [CrossRef] [PubMed]
- Meekins, J.W.; Pijnenborg, R.; Hanssens, M.; McFadyen, I.R.; van Asshe, A. A study of placental bed spiral arteries and trophoblast invasion in normal and severe pre-eclamptic pregnancies. Br. J. Obstet. Gynaecol. 1994, 101, 669–674. [Google Scholar] [CrossRef] [PubMed]
- Huppertz, B. Placental origins of preeclampsia: Challenging the current hypothesis. Hypertension 2008, 51, 970–975. [Google Scholar] [CrossRef] [PubMed]
- Keikkala, E.; Vuorela, P.; Laivuori, H.; Romppanen, J.; Heinonen, S.; Stenman, U.H. First trimester hyperglycosylated human chorionic gonadotrophin in serum—A marker of early-onset preeclampsia. Placenta 2013, 34, 1059–1065. [Google Scholar] [CrossRef] [PubMed]
- Kalkunte, S.; Navers, T.; Norris, W.; Banerjee, P.; Fazleabas, A.; Kuhn, C.; Jeschke, U.; Sharma, S. Presence of non-functional hCG in preeclampsia and rescue of normal pregnancy by recombinant hCG. Placenta 2010, 31, A126. [Google Scholar]
- Kalkunte, S.; Boij, R.; Norris, W.; Friedman, J.; Lai, Z.; Kurtis, J.; Lim, K.H.; Padbury, J.F.; Matthiesen, L.; Sharma, S. Sera from preeclampsia patients elicit symptoms of human disease in mice and provide a basis for an in vitro predictive assay. Am. J. Pathol. 2010, 177, 2387–2398. [Google Scholar] [CrossRef] [PubMed]
- Ong, C.Y.; Liao, A.W.; Spencer, K.; Munim, S.; Nicolaides, K.H. First trimester maternal serum free beta human chorionic gonadotrophin and pregnancy associated plasma protein A as predictors of pregnancy complications. BJOG Int. J. Obstet. Gynaecol. 2000, 107, 1265–1270. [Google Scholar] [CrossRef]
- Smith, G.C.; Stenhouse, E.J.; Crossley, J.A.; Aitken, D.A.; Cameron, A.D.; Connor, J.M. Early pregnancy levels of pregnancy-associated plasma protein a and the risk of intrauterine growth restriction, premature birth, preeclampsia, and stillbirth. J. Clin. Endocrinol. Metab. 2002, 87, 1762–1767. [Google Scholar] [CrossRef] [PubMed]
- Ranta, J.K.; Raatikainen, K.; Romppanen, J.; Pulkki, K.; Heinonen, S. Decreased PAPP-A is associated with preeclampsia, premature delivery and small for gestational age infants but not with placental abruption. Eur. J. Obstet. Gynecol. Reprod. Biol. 2011, 157, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Zhu, F.; Ding, Y. Serum screening in first trimester to predict pre-eclampsia, small for gestational age and preterm delivery: Systematic review and meta-analysis. BMC Pregnancy Childbirth 2015, 15, 191. [Google Scholar] [CrossRef] [PubMed]
- Kagan, K.O.; Wright, D.; Baker, A.; Sahota, D.; Nicolaides, K.H. Screening for trisomy 21 by maternal age, fetal nuchal translucency thickness, free β-human chorionic gonadotropin and pregnancy-associated plasma protein-A. Ultrasound Obstet. Gynecol. Off. J. Int. Soc. Ultrasound Obstet. Gynecol. 2008, 31, 618–624. [Google Scholar] [CrossRef] [PubMed]
- Wright, D.; Spencer, K.; Kagan, K.K.; Torring, N.; Petersen, O.B.; Christou, A.; Kallikas, J.; Nicolaides, K.H. First-trimester combined screening for trisomy 21 at 7–14 weeks’ gestation. Ultrasound Obstet. Gynecol. Off. J. Int. Soc. Ultrasound Obstet. Gynecol. 2010, 36, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Cuckle, H. Biochemical screening for Down syndrome. Eur. J. Obstet. Gynecol. Reprod. Biol. 2000, 92, 97–101. [Google Scholar] [CrossRef]
- American College of Obstetricians and Gynecologists. Diagnosis and treatment of gestational trophoblastic disease. Obstet. Gynecol. 2004, 103, 1365–1377. [Google Scholar]
- Lurain, J.R.; Brewer, J.I.; Torok, E.E.; Halpern, B. Natural history of hydatidiform mole after primary evacuation. Am. J. Obstet. Gynecol. 1983, 145, 591–595. [Google Scholar] [CrossRef]
- Curry, S.L.; Hammond, C.B.; Tyrey, L.; Creasman, W.T.; Parker, R.T. Hydatidiform mole: Diagnosis, management, and long-term followup of 347 patients. Obstet. Gynecol. 1975, 45, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Baergen, R.N.; Rutgers, J.L.; Young, R.H.; Osann, K.; Scully, R.E. Placental site trophoblastic tumor: A study of 55 cases and review of the literature emphasizing factors of prognostic significance. Gynecol. Oncol. 2006, 100, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Khazaeli, M.B.; Hedayat, M.M.; Hatch, K.D.; To, A.C.; Soong, S.J.; Shingleton, H.M.; Boots, L.R.; LoBuglio, A.F. Radioimmunoassay of free β-subunit of human chorionic gonadotropin as a prognostic test for persistent trophoblastic disease in molar pregnancy. Am. J. Obstet. Gynecol. 1986, 155, 320–324. [Google Scholar] [CrossRef]
- Berkowitz, R.; Ozturk, M.; Goldstein, D.; Bernstein, M.; Hill, L.; Wands, J.R. Human chorionic gonadotropin and free subunits’ serum levels in patients with partial and complete hydatidiform moles. Obstet. Gynecol. 1989, 74, 212–216. [Google Scholar] [PubMed]
- Vaitukaitis, J.L.; Ebersole, E.R. Evidence for altered synthesis of human chorionic gonadotropin in gestational trophoblastic tumors. J. Clin. Endocrinol. Metab. 1976, 42, 1048–1055. [Google Scholar] [CrossRef] [PubMed]
- Vartiainen, J.; Alfthan, H.; Lehtovirta, P.; Stenman, U.H. Elevated hCG and a high proportion of hCGβ in serum preceding the diagnosis of trophoblastic disease by seven months. BJOG Int. J. Obstet. Gynaecol. 2002, 109, 589–590. [Google Scholar] [CrossRef]
- Filicori, M.; Fazleabas, A.T.; Huhtaniemi, I.; Licht, P.; Rao Ch, V.; Tesarik, J.; Zygmunt, M. Novel concepts of human chorionic gonadotropin: Reproductive system interactions and potential in the management of infertility. Fertil. Steril. 2005, 84, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Casarini, L.; Brigante, G.; Simoni, M.; Santi, D. Clinical Applications of Gonadotropins in the Female: Assisted Reproduction and Beyond. Prog. Mol. Biol. Transl. Sci. 2016, 143, 85–119. [Google Scholar] [PubMed]
- Santi, D.; Casarini, L.; Alviggi, C.; Simoni, M. Efficacy of Follicle-Stimulating Hormone (FSH) Alone, FSH + Luteinizing Hormone, Human Menopausal Gonadotropin or FSH + Human Chorionic Gonadotropin on Assisted Reproductive Technology Outcomes in the “Personalized” Medicine Era: A Meta-analysis. Front. Endocrinol. 2017, 8, 114. [Google Scholar] [CrossRef] [PubMed]
- Tesarik, J.; Hazout, A.; Mendoza, C. Luteinizing hormone affects uterine receptivity independently of ovarian function. Reprod. Biomed. Online 2003, 7, 59–64. [Google Scholar] [CrossRef]
- Chang, P.; Kenley, S.; Burns, T.; Denton, G.; Currie, K.; DeVane, G.; O’Dea, L. Recombinant human chorionic gonadotropin (rhCG) in assisted reproductive technology: Results of a clinical trial comparing two doses of rhCG (Ovidrel) to urinary hCG (Profasi) for induction of final follicular maturation in in vitro fertilization-embryo transfer. Fertil. Steril. 2001, 76, 67–74. [Google Scholar] [PubMed]
- Abdelmassih, V.; Oliveira, F.G.; Goncalves, S.P.; Varella, A.D.; Diamond, M.P.; Abdelmassih, R. A prospective, randomized and blinded comparison between 10,000 IU urinary and 250 microg recombinant human chorionic gonadotropin for oocyte maturation in in vitro fertilization cycles. J. Assist. Reprod. Genet. 2005, 22, 149–153. [Google Scholar] [CrossRef] [PubMed]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nwabuobi, C.; Arlier, S.; Schatz, F.; Guzeloglu-Kayisli, O.; Lockwood, C.J.; Kayisli, U.A. hCG: Biological Functions and Clinical Applications. Int. J. Mol. Sci. 2017, 18, 2037. https://doi.org/10.3390/ijms18102037
Nwabuobi C, Arlier S, Schatz F, Guzeloglu-Kayisli O, Lockwood CJ, Kayisli UA. hCG: Biological Functions and Clinical Applications. International Journal of Molecular Sciences. 2017; 18(10):2037. https://doi.org/10.3390/ijms18102037
Chicago/Turabian StyleNwabuobi, Chinedu, Sefa Arlier, Frederick Schatz, Ozlem Guzeloglu-Kayisli, Charles Joseph Lockwood, and Umit Ali Kayisli. 2017. "hCG: Biological Functions and Clinical Applications" International Journal of Molecular Sciences 18, no. 10: 2037. https://doi.org/10.3390/ijms18102037
APA StyleNwabuobi, C., Arlier, S., Schatz, F., Guzeloglu-Kayisli, O., Lockwood, C. J., & Kayisli, U. A. (2017). hCG: Biological Functions and Clinical Applications. International Journal of Molecular Sciences, 18(10), 2037. https://doi.org/10.3390/ijms18102037