
Xylose Fermentation by Saccharomyces cerevisiae: Challenges and Prospects

 ,
,
Abstract
:
1. Introduction
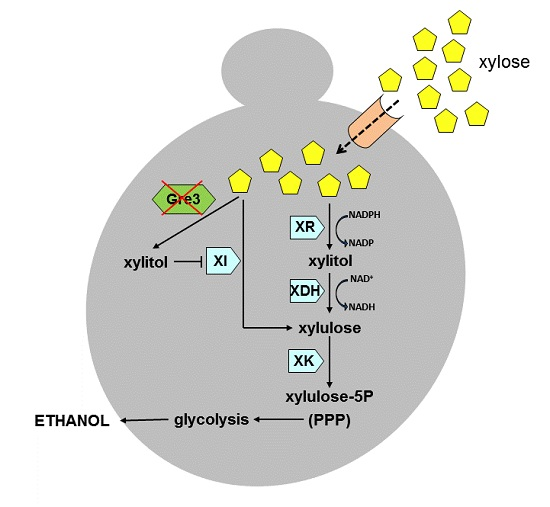
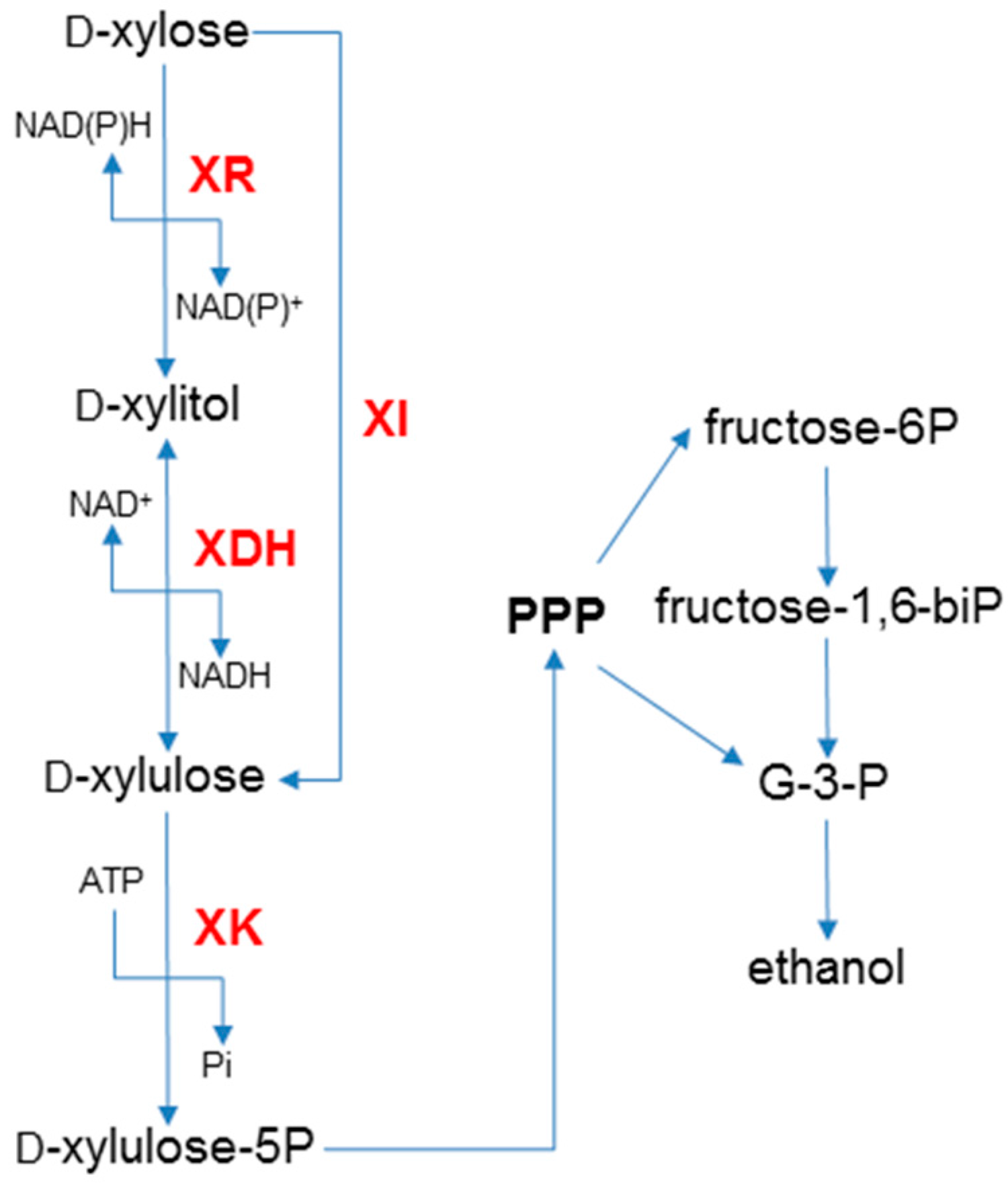
2. Xylose Metabolic Pathways
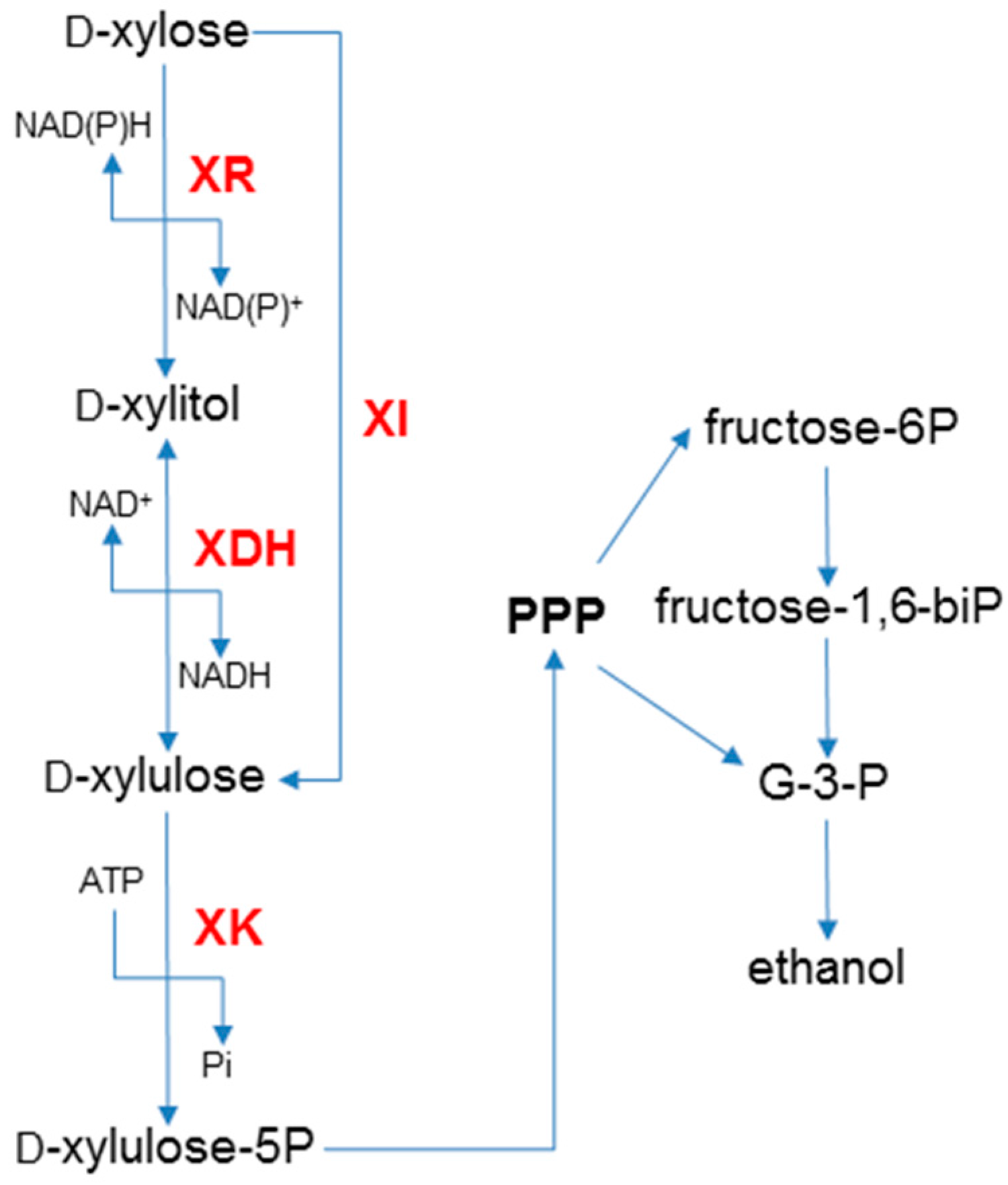
3. Engineering Saccharomyces with the XR/XDH Pathway
4. Engineering Saccharomyces with the XI Pathway
5. Additional Genetic Modifications
6. Evolutionary Engineering
7. Xylose Uptake

{kind=link}
{kind=link}
| Transporter | Strain/Plasmid Construction | Strain Characteristics | Xylose | Glucose | Reference | ||
|---|---|---|---|---|---|---|---|
| Km (mM) | Vmax (nmol/min/mg) | Km (mM) | Vmax (mmol/min/mg) | ||||
| ScHxt1 | H2219/pYX212-HXT1 | Δhxt1-7Δgal2 | 880 ± 8 | 750 ± 94 | NR | NR | [88] |
| ScHxt1 | KY73/HXT1mnX-pVT | Δhxt1-7Δgal2 | NR | NR | 46 | NR | [100] |
| ScHxt2 | H2219/pYX212-hxt2 | Δhxt1-7Δgal2 | 260 ± 130 | 340 ± 10 | NR | NR | [88] |
| ScHxt2 | KY73/HXT2mnX-pVT | Δhxt1-7Δgal2 | NR | NR | 3.3 | NR | [100] |
| ScHxt4 | H2219/pYX212-HXT4 | Δhxt1-7Δgal2 | 170 ± 120 | 190 ± 23 | NR | NR | [88] |
| ScHxt7 | H2219/pYX212-HXT7 | 130 ± 10 | 110 ± 7 | NR | NR | ||
| ScHxt7 | EBY.VW4000/pRS62N-HXT7 | Δhxt1-17 Δgal2 Δstl1 Δagt1 Δmph2 Δmph3 Δydl247w Δyjr160c | 200.3 ± 13.2 | 67 ± 2 | 0.5 ± 0.1 | 26 ± 1.1 | [94] |
| ScHxt7 (N370S) | EBY.VW4000/pRS62N-HXT7-N370S | 169.9 ± 26.3 | 24.1 ±1.6 | 10.8 ±1.0 | 47.3 ± 1.2 | ||
| ScHxt7 | KY73/HXT7mnX-pVT | Δhxt1-7Δgal2 | NR | NR | 0.67 ± 0.05 | 610 ± 60 * | [100] |
| ScHxt7 (T213V) | NR | NR | 0.40 ± 0.02 | 120 * | [101] | ||
| ScHxt7 (D340C) | NR | NR | 0.43 ± 0.05 | 430 ± 40 * | [100] | ||
| ScGal2 | EBY.VW4000/pRS62N-GAL2 | Δhxt1-17 Δgal2 Δstl1 Δagt1 Δmph2 Δmph3 Δydl247w Δyjr160c | 225.6 ± 15.8 | 91.3 ± 3.2 | 1.5 ± 0.2 | 27.2 ± 0.9 | [94] |
| ScGal2 (N376F) | EBY.VW4000/pRS62N-GAL2-N376F | 91.4 ± 8.9 | 37.3 ± 1.3 | ND | BD | [91] | |
| CiGxf1 | TBM3201/pHxt7-GXF1 | Δhxt1-17 Δgal2 Δstl1 Δagt1 Δmph2 Δmph3 | 48.6 ± 6.5 | 64.19 | 2.0 ± 0.6 | 10.5 | [87] |
| CiGxs1 | TBM3201/pHXT7-GXS1 | 0.4 ± 0.1 | 6.5 ± 1.5 | 0.012 ± 0.004 | 4.3 ± 0.3 | ||
| CiGsx1 | EBY.VW4000/p414-TEF-CiGXS1 | Δhxt1-17 Δgal2 Δstl1 Δagt1 Δmph2 Δmph3 Δydl247w Δyjr160c | 0.026 ± 0.066 | 0.0072 | NR | NR | [91] |
| CiGsx1 F38I39M40 | 0.721 ± 0.116 | 0.015 | NR | NR | |||
| SsSut1 | RE700/YEpSUT1 | Δhxt1-7 | 145 ± 1.0 | 132 ± 1.0 | 1.5 ± 0.1 | 45.0 ± 1.0 | [96] |
| SsSut4 | EBY.VW4000/pRS316-TDHp-SUT1 | Δhxt1-17 Δgal2 Δstl1 Δagt1 Δmph2 Δmph3 Δydl247w Δyjr160c | 16.6 ± 0.3 | 122 ± 2.4 | 1.3 ± 0.1 | 105 ± 4.2 | [99] |
| SsXut1 | EBY.VW4000/pRS316-TDHp-XUT1 | 0.46 ± 0.02 | 116 ± 5.8 | 0.91 ± 0.01 | 80 ± 1.0 | ||
| SsXyp29 | EBY.VW4000/pRS429-HXT7p-XYP29 | 56 ± 9.4 | 0.69 ± 0.04 | BD | BD | [89] | |
| NcAn25 | EBY.VW4000/pRS429-HXT7p-AN25 | 175.7 ± 21.4 | 0.61 ± 0.05 | BD | BD | ||
| DhXylh | W303-1A/pRS42K-XYLH | 1.4 † | 1.6 † | NR | NR | [99] | |
| ScHxt36 | DS715054/pRS313-HXT7p-HXT36 | 108 | 62.5 | 6 | 60 | [102] | |
| ScHxt36 (N367I) | Δglk1 Δhxk1 Δhxk2 Δgal1 | 40 | 23 | ND | ND | ||
| ScHxt36 (N367A) | 25 | 29 | 171 | 71 | |||
8. Effects of Lignocellulosic Hydrolysate Inhibitors on Xylose Fermentation
9. Insights on Industrial Application
10. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dien, B.S.; Cotta, M.A.; Jeffries, T.W. Bacteria engineered for fuel ethanol production: Current status. Appl. Microbiol. Biotechnol. 2003, 63, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Farrell, A.E.; Plevin, R.J.; Turner, B.T.; Jones, A.D.; O’Hare, M.; Kammen, D.M. Ethanol can contribute to energy and environmental goals. Science 2006, 311, 506–508. [Google Scholar] [CrossRef] [PubMed]
- Van Maris, A.J.A.; Abbott, D.A.; Bellissimi, E.; van den Brink, J.; Kuyper, M.; Luttik, M.A.; Wisselink, H.W.; Scheffers, W.A.; van Dijken, J.P.; Pronk, J.T. Alcoholic fermentation of carbon sources in biomass hydrolysates by Saccharomyces cerevisiae: Current status. Antonie Van Leeuwenhoek 2006, 90, 391–418. [Google Scholar] [CrossRef] [PubMed]
- Hahn-Hagerdal, B.; Galbe, M.; Gorwa-Grauslund, M.F.; Liden, G.; Zacchi, G. Bioethanol—The fuel of tomorrow from the residues of today. Trends Biotechnol. 2006, 24, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Galbe, M.; Zacchi, G. A review of the production of ethanol from softwood. Appl. Microbiol. Biotechnol. 2002, 59, 618–628. [Google Scholar] [CrossRef] [PubMed]
- Klinke, H.B.; Thomsen, A.B.; Ahring, B.K. Inhibition of ethanol-producing yeast and bacteria by degradation products produced during pre-treatment of biomass. Appl. Microbiol. Biotechnol. 2004, 66, 10–26. [Google Scholar] [CrossRef] [PubMed]
- Mussatto, S.I.; Roberto, I.C. Alternatives for detoxification of diluted-acid lignocellulosic hydrolyzates for use in fermentative processes: A review. Bioresour. Technol. 2004, 93, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Palmqvist, E.; Hahn-Hägerdal, B. Fermentation of lignocellulosic hydrolysates. II: Inhibitors and mechanisms of inhibition. Bioresour. Technol. 2000, 74, 25–33. [Google Scholar] [CrossRef]
- Barnett, J.A.; Payne, R.W.; Yarrow, D. Yeasts: Characteristics and Identification, 2nd ed.; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Olsson, L.; Hahn-Hägerdal, B. Fermentation of lignocellulosic hydrolates for ethanol production. Enzym. Microb. Technol. 1996, 18, 312–331. [Google Scholar] [CrossRef]
- Margaritis, A.; Bajpai, P. Direct fermentation of d-xylose to ethanol by Kluyveromyces marxianus strains. Appl. Environ. Microbiol. 1982, 44, 1039–1041. [Google Scholar] [PubMed]
- Wang, P.Y.; Johnson, B.F.; Schneider, H. Fermentation of d-xylose by yeasts using glucose isomerase in the medium to convert d-xylose to d-xylulose. Biotechnol. Lett. 1980, 2, 273–278. [Google Scholar] [CrossRef]
- Stephens, C.; Christen, B.; Fuchs, T.; Sundaram, V.; Watanabe, K.; Jenal, U. Genetic analysis of a novel pathway for d-xylose metabolism in Caulobacter crescentus. J. Bacteriol. 2007, 189, 2181–2185. [Google Scholar] [CrossRef] [PubMed]
- Verduyn, C.; Kleef, R.; Frank Jzn, J.; Schreuder, H.; Dijken, J.P.; Scheffers, W.A. NAD (P) H-dependent aldose reductase from the xylose-fermenting yeast Pichia stipitis. Antonie Van Leeuwenhoek 1985, 51, 562–562. [Google Scholar] [CrossRef]
- Rizzi, M.; Klein, C.; Schulze, C.; Bui-Thanh, N.A.; Dellweg, H. Xylose fermentation by yeasts. 5. Use of ATP balances for modeling oxygen-limited growth and fermentation of yeast Pichia stipitis with xylose as carbon source. Biotechnol. Bioeng. 1989, 34, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Hahn-Hagerdal, B.; Karhumaa, K.; Fonseca, C.; Spencer-Martins, I.; Gorwa-Grauslund, M.F. Towards industrial pentose-fermenting yeast strains. Appl. Microbiol. Biotechnol. 2007, 74, 937–953. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, U.; Dambeck, M.; Zaiss, H.; Fuhrer, T.; Soppa, J.; Sauer, U.; Schönheit, P. d-xylose degradation pathway in the halophilic archaeon Haloferax volcanii. J. Biol. Chem. 2009, 284, 27290–27303. [Google Scholar] [CrossRef] [PubMed]
- Chang, Q.; Griest, T.; Harter, T.; Petrash, J. Functional studies of aldo-keto reductases in Saccharomyces cerevisiae. Biochim. Biophys. Acta 2007, 1773, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Traff, K.L.; Jonsson, L.J.; Hahn-Hägerdal, B. Putative xylose and arabinose reductases in Saccharomyces cerevisiae. Yeast 2002, 19, 1233–1241. [Google Scholar] [CrossRef] [PubMed]
- Toivari, M.H.; Salusjarvi, L.; Ruohonen, L.; Penttilä, M. Endogenous xylose pathway in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2004, 70, 3681–3686. [Google Scholar] [CrossRef] [PubMed]
- Wahlbom, C.F.; Hahn-Hägerdal, B. Furfural, 5-hydroxymethyl furfural, and acetoin act as external electron acceptors during anaerobic fermentation of xylose in recombinant Saccharomyces cerevisiae. Biotechnol. Bioeng. 2002, 78, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Öhgren, K.; Bengtsson, O.; Gorwa-Grauslund, M.F.; Galbe, M.; Hahn-Hägerdal, B.; Zacchi, G. Simultaneous saccharification and co-fermentation of glucose and xylose in steam-pretreated corn stover at high fiber content with Saccharomyces cerevisiae TMB3400. J. Botechnol. 2006, 126, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Almeida, J.R.M.; Bertilsson, M.; Hahn-Hägerdal, B.; Lidén, G.; Gorwa-Grauslund, M.-F. Carbon fluxes of xylose-consuming Saccharomyces cerevisiae strains are affected differently by NADH and NADPH usage in HMF reduction. Appl. Microbiol. Biotechnol. 2009, 84, 751–761. [Google Scholar]
- Roca, C.; Nielsen, J.; Olsson, L. Metabolic engineering of ammonium assimilation in xylose-fermenting Saccharomyces cerevisiae improves ethanol production. Appl. Environ. Microbiol. 2003, 69, 4732–4736. [Google Scholar] [CrossRef] [PubMed]
- Sonderegger, M.; Schumperli, M.; Sauer, U. Metabolic engineering of a phosphoketolase pathway for pentose catabolism in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2004, 70, 2892–2897. [Google Scholar] [CrossRef] [PubMed]
- Almeida, J.R.M.; Runquist, D.; Sànchez Nogué, V.; Lidén, G.; Gorwa-Grauslund, M.F. Stress-related challenges in pentose fermentation to ethanol by the yeast Saccharomyces cerevisiae. Biotechnol. J. 2011, 6, 286–299. [Google Scholar] [CrossRef] [PubMed]
- Sarthy, A.V.; McConaughy, B.L.; Lobo, Z.; Sundstrom, J.A.; Furlong, C.E.; Hall, B.D. Expression of the Escherichia coli xylose isomerase gene in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1987, 53, 1996–2000. [Google Scholar] [PubMed]
- Amore, R.; Wilhelm, M.; Hollenberg, C.P. The fermentation of xylose—An analysis of the expression of Bacillus and Actinoplanes xylose isomerase genes in yeast. Appl. Microbiol. Biotechnol. 1989, 30, 351–357. [Google Scholar] [CrossRef]
- Moes, C.J.; Pretorius, I.S.; van Zyl, W.H. Cloning and expression of the Clostridium thermosulfurogenes d-xylose isomerase gene (xylA) in Saccharomyces cerevisiae. Biotechnol. Lett. 1996, 18, 269–274. [Google Scholar] [CrossRef]
- Gárdonyi, M.; Hahn-Hägerdal, B. The Streptomyces rubiginosus xylose isomerase is misfolded when expressed in Saccharomyces cerevisiae. Enzym. Microb. Technol. 2003, 32, 252–259. [Google Scholar] [CrossRef]
- Van Maris, A.J.; Winkler, A.A.; Kuyper, M.; de Laat, W.T.; van Dijken, J.P.; Pronk, J.T. Development of efficient xylose fermentation in Saccharomyces cerevisiae: Xylose isomerase as a key component. Adv. Biochem. Eng. Biotechnol. 2007, 108, 179–204. [Google Scholar] [PubMed]
- Walfridsson, M.; Bao, X.; Anderlund, M.; Lilius, G.; Bulow, L.; Hahn-Hagerdal, B. Ethanolic fermentation of xylose with Saccharomyces cerevisiae harboring the Thermus thermophilus xylA gene, which expresses an active xylose (glucose) isomerase. Appl. Environ. Microbiol. 1996, 62, 4648–4651. [Google Scholar] [PubMed]
- Lönn, A.; Träff-Bjerre, K.L.; Cordero Otero, R.R.; van Zyl, W.H.; Hahn-Hägerdal, B. Xylose isomerase activity influences xylose fermentation with recombinant Saccharomyces cerevisiae strains expressing mutated xylA from Thermus thermophilus. Enzym. Microb. Technol. 2003, 32, 567–573. [Google Scholar] [CrossRef]
- Ha, S.J.; Kim, S.R.; Choi, J.H.; Park, M.S.; Jin, Y.S. Xylitol does not inhibit xylose fermentation by engineered Saccharomyces cerevisiae expressing xylA as severely as it inhibits xylose isomerase reaction in vitro. Appl. Microbiol. Biotechnol. 2011, 92, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.M.; Choi, S.H.; Ko, B.S.; Jeong, G.Y.; Jang, H.B.; Han, J.G.; Jeong, K.H.; Lee, H.Y.; Won, Y.; Kim, I.C. Reduction of PDC1 expression in S. cerevisiae with xylose isomerase on xylose medium. Bioprocess. Biosyst. Eng. 2012, 35, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Aeling, K.A.; Salmon, K.A.; Laplaza, J.M.; Li, L.; Headman, J.R.; Hutagalung, A.H.; Picataggio, S. Co-fermentation of xylose and cellobiose by an engineered Saccharomyces cerevisiae. J. Ind. Microbiol. Biotechnol. 2012, 39, 1597–1604. [Google Scholar] [CrossRef] [PubMed]
- Kuyper, M.; Harhangi, H.R.; Stave, A.K.; Winkler, A.A.; Jetten, M.S.; de Laat, W.T.; den Ridder, J.J.; Op den Camp, H.J.; van Dijken, J.P.; Pronk, J.T. High-level functional expression of a fungal xylose isomerase: The key to efficient ethanolic fermentation of xylose by Saccharomyces cerevisiae? FEMS Yeast Res. 2003, 4, 69–78. [Google Scholar] [CrossRef]
- Kuyper, M.; Winkler, A.A.; van Dijken, J.P.; Pronk, J.T. Minimal metabolic engineering of Saccharomyces cerevisiae for efficient anaerobic xylose fermentation: A proof of principle. FEMS Yeast Res. 2004, 4, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Kuyper, M.; Hartog, M.M.; Toirkens, M.J.; Almering, M.J.; Winkler, A.A.; van Dijken, J.P.; Pronk, J.T. Metabolic engineering of a xylose-isomerase-expressing Saccharomyces cerevisiae strain for rapid anaerobic xylose fermentation. FEMS Yeast Res. 2005, 5, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Kuyper, M.; Toirkens, M.J.; Diderich, J.A.; Winkler, A.A.; Dijken, J.P.; Pronk, J.T. Evolutionary engineering of mixed-sugar utilization by a xylose-fermenting Saccharomyces cerevisiae strain. FEMS Yeast Res. 2005, 5, 925–934. [Google Scholar] [CrossRef] [PubMed]
- Madhavan, A.; Tamalampudi, S.; Srivastava, A.; Fukuda, H.; Bisaria, V.S.; Kondo, A. Alcoholic fermentation of xylose and mixed sugars using recombinant Saccharomyces cerevisiae engineered for xylose utilization. Appl. Microbiol. Biotechnol. 2009, 82, 1037–1047. [Google Scholar] [CrossRef] [PubMed]
- Madhavan, A.; Tamalampudi, S.; Ushida, K.; Kanai, D.; Katahira, S.; Srivastava, A.; Fukuda, H.; Bisaria, V.S.; Kondo, A. Xylose isomerase from polycentric fungus Orpinomyces: Gene sequencing, cloning, and expression in Saccharomyces cerevisiae for bioconversion of xylose to ethanol. Appl. Microbiol. Biotechnol. 2009, 82, 1067–1078. [Google Scholar] [CrossRef] [PubMed]
- Brat, D.; Boles, E.; Wiedemann, B. Functional expression of a bacterial xylose isomerase in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2009, 75, 2304–2311. [Google Scholar] [CrossRef] [PubMed]
- Hector, R.E.; Dien, B.S.; Cotta, M.A.; Mertens, J.A. Growth and fermentation of d-xylose by Saccharomyces cerevisiae expressing a novel d-xylose isomerase originating from the bacterium Prevotella ruminicola TC2-24. Biotechnol. Biofuels 2013, 6. [Google Scholar] [CrossRef] [PubMed]
- Vilela, L.F.; de Mello, V.M.; Reis, V.C.; Bon, E.P.; Goncalves, D.L.; Torres, F.A.; Neves, B.C.; Eleutherio, E.C. Functional expression of Burkholderia cenocepacia xylose isomerase in yeast increases ethanol production from a glucose-xylose blend. Bioresour. Technol. 2013, 128, 792–796. [Google Scholar] [CrossRef] [PubMed]
- Vilela, L.F.; de Araujo, V.P.; Paredes, R.S.; Bon, E.P.; Torres, F.A.; Neves, B.C.; Eleutherio, E.C. Enhanced xylose fermentation and ethanol production by engineered Saccharomyces cerevisiae strain. AMB Express 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Peng, B.; Huang, S.; Liu, T.; Geng, A. Bacterial xylose isomerases from the mammal gut Bacteroidetes cluster function in Saccharomyces cerevisiae for effective xylose fermentation. Microb. Cell Fact. 2015, 14, 70. [Google Scholar] [CrossRef] [PubMed]
- Hahn-Hägerdal, B.; Jeppsson, H.; Skoog, K.; Prior, B.A. Biochemistry and physiology of xylose fermentation by yeasts. Enzym. Microb. Technol. 1994, 16, 933–943. [Google Scholar] [CrossRef]
- Ota, M.; Sakuragi, H.; Morisaka, H.; Kuroda, K.; Miyake, H.; Tamaru, Y.; Ueda, M. Display of Clostridium cellulovorans xylose isomerase on the cell surface of Saccharomyces cerevisiae and its direct application to xylose fermentation. Biotechnol. Prog. 2013, 29, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Parachin, N.S.; Bergdahl, B.; van Niel, E.W.; Gorwa-Grauslund, M.F. Kinetic modelling reveals current limitations in the production of ethanol from xylose by recombinant Saccharomyces cerevisiae. Metab. Eng. 2011, 13, 508–517. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.S.; Ni, H.; Laplaza, J.M.; Jeffries, T.W. Optimal growth and ethanol production from xylose by recombinant Saccharomyces cerevisiae require moderate d-xylulokinase activity. Appl. Environ. Microbiol. 2003, 69, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Senac, T.; Hahn-Hagerdal, B. Effects of increased transaldolase activity on d-xylulose and d-glucose metabolism in Saccharomyces cerevisiae cell extracts. Appl. Environ. Microbiol. 1991, 57, 1701–1706. [Google Scholar] [PubMed]
- Yamanaka, K. Inhibition of d-xylose isomerase by pentitols and d-lyxose. Arch. Biochem. Biophys. 1969, 131, 502–506. [Google Scholar] [CrossRef]
- Kuhn, A.; van Zyl, C.; van Tonder, A.; Prior, B.A. Purification and partial characterization of an aldo-keto reductase from Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1995, 61, 1580–1585. [Google Scholar] [PubMed]
- Traff, K.L.; Otero Cordero, R.R.; van Zyl, W.H.; Hahn-Hagerdal, B. Deletion of the GRE3 aldose reductase gene and its influence on xylose metabolism in recombinant strains of Saccharomyces cerevisiae expressing the xylA and XKS1 genes. Appl. Environ. Microbiol. 2001, 67, 5668–5674. [Google Scholar] [CrossRef] [PubMed]
- Verho, R.; Londesborough, J.; Penttila, M.; Richard, P. Engineering redox cofactor regeneration for improved pentose fermentation in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2003, 69, 5892–5897. [Google Scholar] [CrossRef] [PubMed]
- Van Vleet, J.H.; Jeffries, T.W.; Olsson, L. Deleting the para-nitrophenyl phosphatase (pNPPase), PHO13, in recombinant Saccharomyces cerevisiae improves growth and ethanol production on d-xylose. Metab. Eng. 2008, 10, 360–369. [Google Scholar] [CrossRef] [PubMed]
- Bro, C.; Regenberg, B.; Forster, J.; Nielsen, J. In silico aided metabolic engineering of Saccharomyces cerevisiae for improved bioethanol production. Metab. Eng. 2006, 8, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Usher, J.; Balderas-Hernandez, V.; Quon, P.; Gold, N.D.; Martin, V.J.; Mahadevan, R.; Baetz, K. Chemical and synthetic genetic array analysis identifies genes that suppress xylose utilization and fermentation in Saccharomyces cerevisiae. G3 2011, 1, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Baley, J.E. Lessons from metabolic engineering for functional genomics and drug discovery. Nat. Biotechnol. 1999, 17, 616–618. [Google Scholar] [CrossRef] [PubMed]
- Petri, R.; Schmidt-Dannert, C. Dealing with complexity: Evolutionary engineering and genome shuffling. Curr. Opin. Biotechnol. 2004, 15, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, J.; Andreu, P.; Randez-Gil, F.; Prieto, J.A. Adaptive evolution of baker’s yeast in a dough-like environment enhances freeze and salinity tolerance. Microb. Biotechnol. 2010, 3, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Attfield, P.V.; Bell, P.J.L. Use of population genetics to derive nonrecombinant Saccharomyces cerevisiae strains that grow using xylose as a sole carbon source. FEMS Yeast Res. 2006, 6, 862–868. [Google Scholar] [CrossRef] [PubMed]
- Sonderegger, M.; Sauer, U. Evolutionary engineering of Saccharomyces cerevisiae for anaerobic growth on xylose. Appl. Environ. Microbiol. 2003, 69, 1990–1998. [Google Scholar] [CrossRef] [PubMed]
- Scalcinati, G.; Otero, J.M.; Vleet, J.R.; Jeffries, T.W.; Olsson, L.; Nielsen, J. Evolutionary engineering of Saccharomyces cerevisiae for efficient aerobic xylose consumption. FEMS Yeast Res. 2012, 12, 582–597. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.R.; Kwee, N.R.; Kim, H.; Jin, Y.S. Feasibility of xylose fermentation by engineered Saccharomyces cerevisiae overexpressing endogenous aldose reductase (GRE3), xylitol dehydrogenase (XYL2), and xylulokinase (XYL3) from Scheffersomyces stipitis. FEMS Yeast Res. 2013, 13, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Klimacek, M.; Kirl, E.; Krahulec, S.; Longus, K.; Novy, V.; Nidetzky, B. Stepwise metabolic adaption from pure metabolization to balanced anaerobic growth on xylose explored for recombinant Saccharomyces cerevisiae. Microb. Cell Fact. 2014, 13. [Google Scholar] [CrossRef] [PubMed]
- Demeke, M.M.; Dietz, H.; Li, Y.; Foulquie-Moreno, M.R.; Mutturi, S.; Deprez, S.; Den Abt, T.; Bonini, B.M.; Liden, G.; Dumortier, F.; et al. Development of a d-xylose fermenting and inhibitor tolerant industrial Saccharomyces cerevisiae strain with high performance in lignocellulose hydrolysates using metabolic and evolutionary engineering. Biotechnol. Biofuels 2013, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Cheng, J.S.; Wang, B.L.; Fink, G.R.; Stephanopoulos, G. Xylose isomerase overexpression along with engineering of the pentose phosphate pathway and evolutionary engineering enable rapid xylose utilization and ethanol production by Saccharomyces cerevisiae. Metab. Eng. 2012, 14, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Demeke, M.M.; Foulquie-Moreno, M.R.; Dumortier, F.; Thevelein, J.M. Rapid evolution of recombinant Saccharomyces cerevisiae for xylose fermentation through formation of extra-chromosomal circular DNA. PLoS Genet. 2015, 11, e1005010. [Google Scholar] [CrossRef] [PubMed]
- Alper, H.; Moxley, J.; Nevoigt, E.; Fink, G.R.; Stephanopoulos, G. Engineering yeast transcription machinery for improved ethanol tolerance and production. Science 2006, 314, 1565–1568. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Bae, J.Y.; Lee, Y.M.; Kwon, H.; Moon, H.Y.; Kang, H.A.; Yee, S.B.; Kim, W.; Choi, W. Construction of Saccharomyces cerevisiae strains with enhanced ethanol tolerance by mutagenesis of the TATA-binding protein gene and identification of novel genes associated with ethanol tolerance. Biotechnol. Bioeng. 2011, 108, 1776–1787. [Google Scholar] [CrossRef] [PubMed]
- Liu, E.; Hu, Y. Construction of a xylose-fermenting Saccharomyces cerevisiae strain by combined approaches of genetic engineering, chemical mutagenesis and evolutionary adaptation. Biochem. Eng. J. 2010, 48, 204–210. [Google Scholar] [CrossRef]
- Liu, H.; Liu, K.; Yan, M.; Xu, L.; Ouyang, P. gTME for improved adaptation of Saccharomyces cerevisiae to corn cob acid hydrolysate. Appl. Biochem. Biotechnol. 2011, 164, 1150–1159. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Zhang, B.; Li, Y. Engineering Saccharomyces cerevisiae for efficient anaerobic xylose fermentation: Reflections and perspectives. Biotechnol. J. 2012, 7, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Wahlbom, C.F.; Otero, R.R.C.; van Zyl, W.H.; Hahn-Hägerdal, B.; Jönsson, L.J. Molecular analysis of a Saccharomyces cerevisiae mutant with improved ability to utilize xylose shows enhanced expression of proteins involved in transport, initial xylose metabolism, and the pentose phosphate pathway. Appl. Environ. Microbiol. 2003, 69, 740–746. [Google Scholar] [CrossRef] [PubMed]
- Hector, R.E.; Qureshi, N.; Hughes, S.R.; Cotta, M.A. Expression of a heterologous xylose transporter in a Saccharomyces cerevisiae strain engineered to utilize xylose improves aerobic xylose consumption. Appl. Microbiol. Biotechnol. 2008, 80, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Runquist, D.; Hahn-Hagerdal, B.; Radstrom, P. Comparison of heterologous xylose transporters in recombinant Saccharomyces cerevisiae. Biotechnol. Biofuels. 2010, 17, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Diao, L.; Liu, Y.; Qian, F.; Yang, J.; Jiang, Y.; Yang, S. Construction of fast xylose-fermenting yeast based on industrial ethanol-producing diploid Saccharomyces cerevisiae by rational design and adaptive evolution. BMC Biotechnol. 2013, 13. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, D.L.; Matsushika, A.; Belisa, B.; Goshima, T.; Bon, E.P.; Stambuk, B.U. Xylose and xylose/glucose co-fermentation by recombinant Saccharomyces cerevisiae strains expressing individual hexose transporters. Enzym. Microb. Technol. 2014, 63, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Reifenberger, E.; Boles, E.; Ciriacy, M. Kinetic characterization of individual hexose transporters of Saccharomyces cerevisiae and their relation to the triggering mechanisms of glucose repression. Eur. J. Biochem. 1997, 245, 324–333. [Google Scholar]
- Diderich, J.A.; Schepper, M.; van Hoek, P.; Luttik, M.A.; van Dijken, J.P.; Pronk, J.T.; Klaassen, P.; Boelens, H.F.; de Mattos, M.J.; van Dam, K.; et al. Glucose uptake kinetics and transcription of HXT genes in chemostat cultures of Saccharomyces cerevisiae. J. Biol. Chem. 1999, 274, 15350–15359. [Google Scholar] [CrossRef] [PubMed]
- Sedlak, M.; Ho, N.W. Characterization of the effectiveness of hexose transporters for transporting xylose during glucose and xylose co-fermentation by a recombinant Saccharomyces yeast. Yeast 2004, 21, 671–684. [Google Scholar] [CrossRef] [PubMed]
- Hamacher, T.; Becker, J.; Gárdonyi, M.; Hahn-Hägerdal, B.; Boles, E. Characterization of the xylose-transporting properties of yeast hexose transporters and their influence on xylose utilization. Microbiology 2002, 148, 2783–2788. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Kim, M.D.; Ryu, Y.W.; Bisson, L.F.; Seo, J.H. Kinetic studies on glucose and xylose transport in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2002, 60, 186–191. [Google Scholar] [PubMed]
- Subtil, T.; Boles, E. Competition between pentoses and glucose during uptake and catabolism in recombinant Saccharomyces cerevisiae. Biotechnol. Biofuels 2012, 5. [Google Scholar] [CrossRef] [PubMed]
- Leandro, M.; Gonçalves, P.; Spencer-Martins, I. Two glucose/xylose transporter genes from the yeast Candida intermedia: First molecular characterization of a yeast xylose-H+ symporter. Biochem. J. 2006, 395, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Saloheimo, A.; Rauta, J.; Stasyk, O.V.; Sibirny, A.A.; Penttilä, M.; Ruohonen, L. Xylose transport studies with xylose-utilizing Saccharomyces cerevisiae strains expressing heterologous and homologous permeases. Appl. Microbiol. Biotechnol. 2007, 74, 1041–1052. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Li, S.; Zhao, H. Discovery and characterization of novel d-xylose-specific transporters from Neurospora crassa and Pichia stipitis. Mol. Biosyst. 2010, 6, 2150–2156. [Google Scholar] [CrossRef] [PubMed]
- Young, E.; Poucher, A.; Comer, A.; Bailey, A.; Alper, H. Functional survey for heterologous sugar transport proteins, using Saccharomyces cerevisiae as a host. Appl. Environ. Microbiol. 2011, 77, 3311–3319. [Google Scholar] [CrossRef] [PubMed]
- Young, E.M.; Tong, A.; Bui, H.; Spofford, C.; Alper, H.S. Rewiring yeast sugar transporter preference through modifying a conserved protein motif. Proc. Natl. Acad. Sci. USA 2013, 111, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Colabardini, A.C.; Ries, L.N.A.; Brown, N.A.; Dos Reis, T.F.; Savoldi, M.; Goldman, M.H.S.; Menino, J.F.; Rodrigues, F.; Goldman, G.H. Functional characterization of a xylose transporter in Aspergillus nidulans. Biotechnol. Biofuels 2014, 7. [Google Scholar] [CrossRef] [PubMed]
- Wieczorke, R.; Krampe, S.; Weierstall, T.; Freidel, K.; Hollenberg, C.P.; Boles, E. Concurrent knock-out of at least 20 transporter genes is required to block uptake of hexoses in Saccharomyces cerevisiae. FEBS Lett. 1999, 464, 123–128. [Google Scholar] [CrossRef]
- Farwick, A.; Bruder, S.; Schadeweg, V.; Oreb, M.; Boles, E. Engineering of yeast hexose transporters to transport d-xylose without inhibition by d-glucose. Proc. Natl. Acad. Sci. USA 2014, 111, 5159–5164. [Google Scholar] [CrossRef] [PubMed]
- Oreb, M.; Dietz, H.; Farwick, A.; Boles, E. Novel strategies to improve co-fermentation of pentoses with d-glucose by recombinant yeast strains in lignocellulosic hydrolysates. Bioengineered 2012, 3, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Weierstall, T.; Hollenberg, C.P.; Boles, E. Cloning and characterization of three genes (SUT1-3) encoding glucose transporters of the yeast Pichia stipitis. Mol. Microbiol. 1999, 31, 871–883. [Google Scholar] [CrossRef] [PubMed]
- Jeffries, T.W.; Bae, J.; Lin, B.C.Y.; van Vleet, J.R.H. Sugar Transport Sequences, Yeast Strains Having Improved Sugar Uptake, and Methods of Use. U.S. Patent Application Publication US 2009/0325241 A1, 31 December 2009. [Google Scholar]
- Wang, C.; Bao, X.; Li, Y.; Jiao, C.; Hou, J.; Zhang, Q.; Zhang, W.; Liu, W.; Shen, Y. Cloning and characterization of heterologous transporters in Saccharomyces cerevisiae and identification of important amino acids for xylose utilization. Metabol. Eng. 2015, 30, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, D.; Nobre, A.; Silva, M.L.; Faria-Oliveira, F.; Tulha, J.; Ferreira, C.; Lucas, C. XYLH encodes a xylose/H+ symporter from the highly related yeast species Debaryomyces fabryi and Debaryomyces hansenii. FEMS Yeast Res. 2013, 13, 585–596. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kasahara, T.; Kasahara, M. A two dimensional comprehensive study: Identification of a key residue determining substrate affinity in the yeast glucose transporter Hxt7. J. Biol. Chem. 2010, 285, 26263–26268. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, T.; Shimogawara, K.; Kasahara, M. Crucial effects of amino acid side chain length in transmembrane segment 5 on substrate affinity in yeast glucose transporter Hxt7. Biochemistry 2011, 50, 8674–8681. [Google Scholar] [CrossRef] [PubMed]
- Nijland, J.G.; Shin, H.Y.; de Jong, R.M.; de Waal, P.P.; Klaassen, P.; Driessen, A.J. Engineering of an endogenous hexose transporter into a specific d-xylose transporter facilitates glucose-xylose co-consumption in Saccharomyces cerevisiae. Biotechnol. Biofuels 2014, 7. [Google Scholar] [CrossRef] [PubMed]
- Almeida, J.R.M.; Modig, T.; Petersson, A.; Hahn-Hägerdal, B.; Lidén, G.; Gorwa-Grauslund, M.F. Increased tolerance and conversion of inhibitors in lignocellulosic hydrolysates by Saccharomyces cerevisiae. J. Chem. Technol. Biotechnol. 2007, 82, 340–349. [Google Scholar] [CrossRef]
- Ask, M.; Bettiga, M.; Duraiswamy, V.; Olsson, L. Pulsed addition of HMF and furfural to batch-grown xylose-utilizing Saccharomyces cerevisiae results in different physiological responses in glucose and xylose consumption phase. Biotechnol. Biofuels 2013, 6. [Google Scholar] [CrossRef] [PubMed]
- Demeke, M.M.; Dumortier, F.; Li, Y.; Broeckx, T.; Foulquie-Moreno, M.R.; Thevelein, J.M. Combining inhibitor tolerance and d-xylose fermentation in industrial Saccharomyces cerevisiae for efficient lignocellulose-based bioethanol production. Biotechnol. Biofuels 2013, 6. [Google Scholar] [CrossRef] [PubMed]
- Wallace-Salinas, V.; Signori, L.; Li, Y.-Y.; Ask, M.; Bettiga, M.; Porro, D.; Thevelein, J.M.; Branduardi, P.; Foulquié-Moreno, M.R.; Gorwa-Grauslund, M. Re-assessment of YAP1 and MCR1 contributions to inhibitor tolerance in robust engineered Saccharomyces cerevisiae fermenting undetoxified lignocellulosic hydrolysate. AMB Express 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jin, M.; Balan, V.; Jones, A.D.; Li, X.; Li, B.-Z.; Dale, B.E.; Yuan, Y.-J. Comparative metabolic profiling revealed limitations in xylose-fermenting yeast during co-fermentation of glucose and xylose in the presence of inhibitors. Biotechnol. Bioeng. 2014, 111, 152–164. [Google Scholar] [CrossRef] [PubMed]
- Bergdahl, B.; Heer, D.; Sauer, U.; Hahn-Hägerdal, B.; van Niel, E.W. Dynamic metabolomics differentiates between carbon and energy starvation in recombinant Saccharomyces cerevisiae fermenting xylose. Biotechnol. Biofuels 2012, 5. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.; van Rensburg, E.; Görgens, J.F. Simultaneously improving xylose fermentation and tolerance to lignocellulosic inhibitors through evolutionary engineering of recombinant Saccharomyces cerevisiae harbouring xylose isomerase. BMC Biotechnol. 2014, 14. [Google Scholar] [CrossRef] [PubMed]
- Bellissimi, E.; van Dijken, J.P.; Pronk, J.T.; van Maris, A.J.A. Effects of acetic acid on the kinetics of xylose fermentation by an engineered, xylose-isomerase-based Saccharomyces cerevisiae strain. FEMS Yeast Res. 2009, 9, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Alkasrawi, M.; Rudolf, A.; Lidén, G.; Zacchi, G. Influence of strain and cultivation procedure on the performance of simultaneous saccharification and fermentation of steam pretreated spruce. Enzym. Microb. Technol. 2006, 38, 279–286. [Google Scholar] [CrossRef]
- Tomás-Pejó, E.; Olsson, L. Influence of the propagation strategy for obtaining robust Saccharomyces cerevisiae cells that efficiently co-ferment xylose and glucose in lignocellulosic hydrolysates. Microb. Biotechnol. 2015, 8, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Xia, L. Ethanol production from corn stover hemicellulosic hydrolysate using immobilized recombinant yeast cells. Biochem. Eng. J. 2010, 49, 28–32. [Google Scholar] [CrossRef]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moysés, D.N.; Reis, V.C.B.; Almeida, J.R.M.d.; Moraes, L.M.P.d.; Torres, F.A.G. Xylose Fermentation by Saccharomyces cerevisiae: Challenges and Prospects. Int. J. Mol. Sci. 2016, 17, 207. https://doi.org/10.3390/ijms17030207
Moysés DN, Reis VCB, Almeida JRMd, Moraes LMPd, Torres FAG. Xylose Fermentation by Saccharomyces cerevisiae: Challenges and Prospects. International Journal of Molecular Sciences. 2016; 17(3):207. https://doi.org/10.3390/ijms17030207
Chicago/Turabian StyleMoysés, Danuza Nogueira, Viviane Castelo Branco Reis, João Ricardo Moreira de Almeida, Lidia Maria Pepe de Moraes, and Fernando Araripe Gonçalves Torres. 2016. "Xylose Fermentation by Saccharomyces cerevisiae: Challenges and Prospects" International Journal of Molecular Sciences 17, no. 3: 207. https://doi.org/10.3390/ijms17030207
APA StyleMoysés, D. N., Reis, V. C. B., Almeida, J. R. M. d., Moraes, L. M. P. d., & Torres, F. A. G. (2016). Xylose Fermentation by Saccharomyces cerevisiae: Challenges and Prospects. International Journal of Molecular Sciences, 17(3), 207. https://doi.org/10.3390/ijms17030207