
Metabolic and Physiological Responses of Shiraz and Cabernet Sauvignon (Vitis vinifera L.) to Near Optimal Temperatures of 25 and 35 °C

Abstract
:
1. Introduction
2. Results
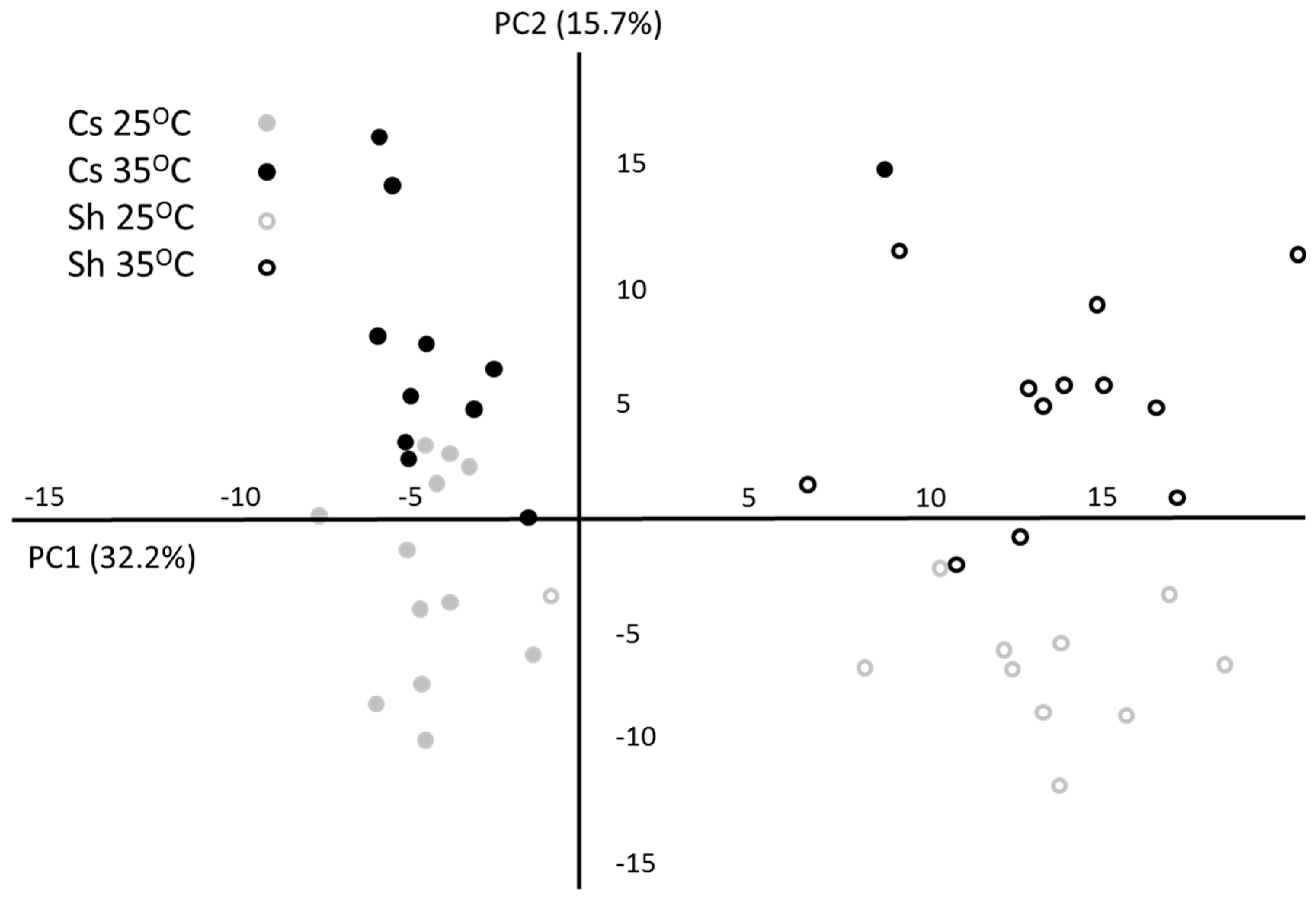
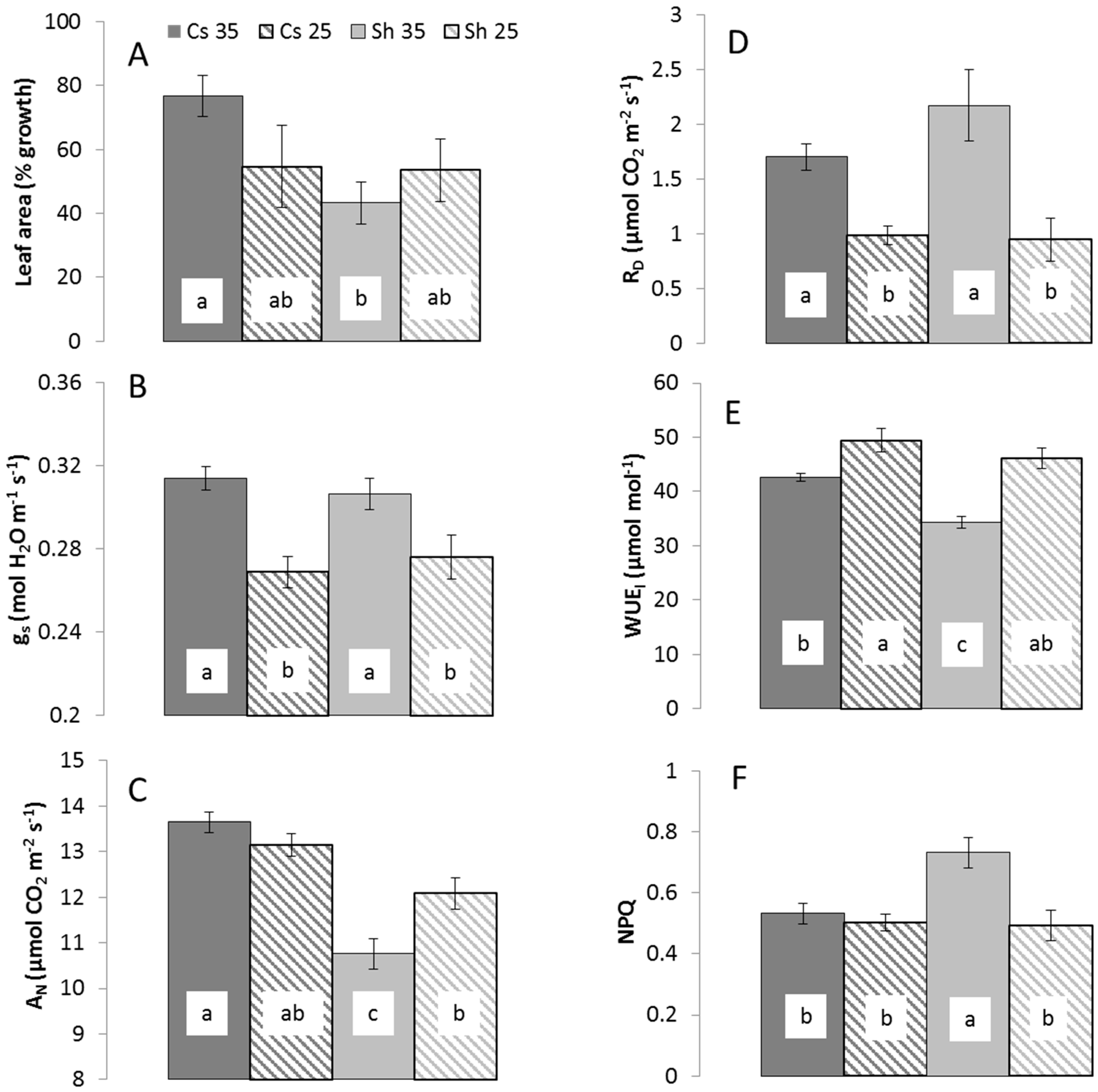
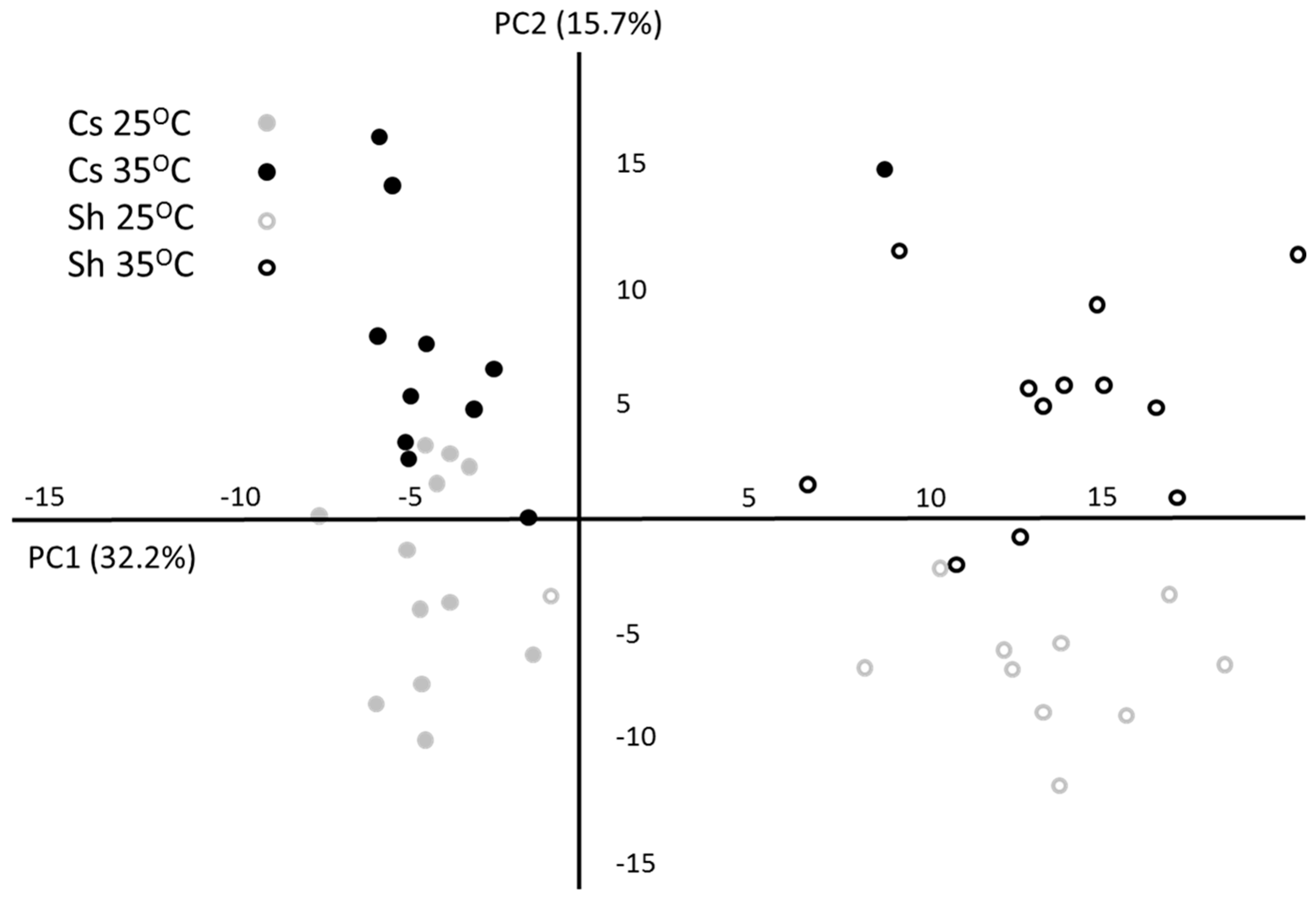
2.1. Metabolic Response to Temperature
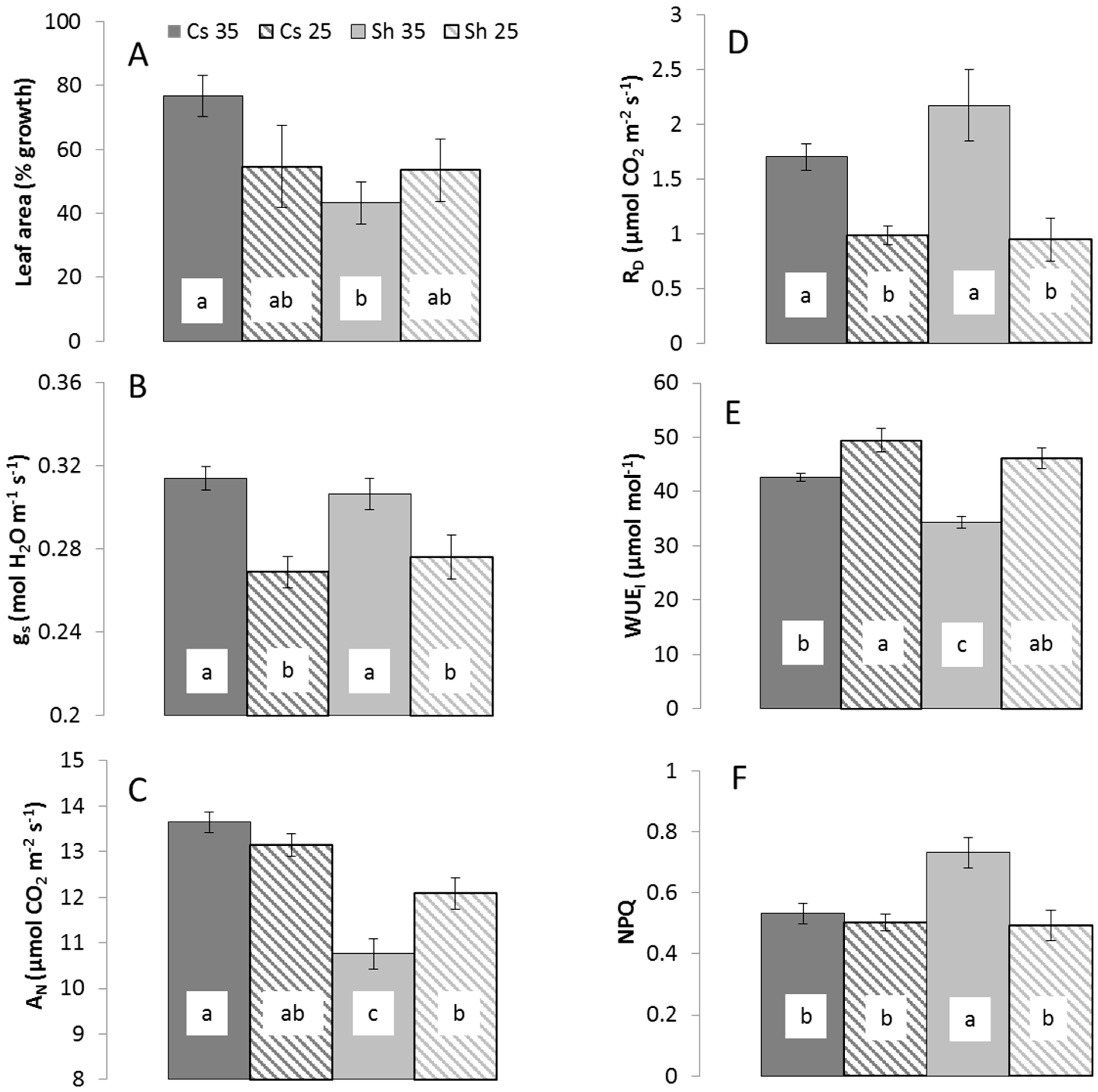

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Cs 35/25 | Sh 35/25 |
|---|---|---|
| Amino acids | ||
| Ala | 1.38 | 1.60 |
| Gly | 0.66 | 0.69 |
| Ser | 0.41 | 0.54 |
| Glu | 0.47 | 0.64 |
| Asp | 1.67 | 1.71 |
| Organic acids | ||
| Citrate | 1.39 | 1.64 |
| Malate | 0.36 | 0.57 |
| Maleate | 0.72 | 0.77 |
| Succinate | 0.48 | 0.52 |
| Threonate | 0.67 | 0.55 |
| Threonate-1,4-lactone | 0.71 | 0.74 |
| Glycerate | 0.56 | 0.51 |
| Malonate | 1.26 | 1.19 |
| Phosphorate | 1.25 | 1.53 |
| Erythronate | 1.53 | 2.08 |
| Glutarate 2-oxo | 0.71 | 0.96 |
| Arabinonic acid | 1.38 | 1.61 |
| 4-Hydroxy trans-cinnamic acid | 1.79 | 1.52 |
| 4-Hydroxy cis-cinnamic acid | 1.27 | 1.11 |
| Shikimate | 0.74 | 0.85 |
| Dehydroascorbate | 0.74 | 0.71 |
| Ascorbate | 1.28 | 1.15 |
| Glucuronate | 1.36 | 1.45 |
| cis-Caffeaet | 1.28 | 1.16 |
| Galactonate | 1.54 | 1.64 |
| trans-Caffeate | 1.32 | 1.09 |
| Sugars | ||
| Fructose | 0.90 | 0.53 |
| Glucose | 0.61 | 0.40 |
| Glucose-6-phosphate | 0.62 | 0.62 |
| Sucrose | 0.68 | 0.63 |
| Xylose | 0.84 | 0.82 |
| Fucose | 1.53 | 1.82 |
| Ribulose | 1.70 | 1.44 |
| Raffinose | 1.81 | 3.13 |
| Others | ||
| Putrescine | 0.28 | 0.34 |
| Epicatechin | 0.55 | 0.85 |
| Catechin | 0.79 | 1.37 |
| Epigallocatechin | 0.99 | 1.57 |
2.2. Cultivar Metabolic Differences
| Metabolite | Cs/Sh 35 | Cs/Sh 25 |
|---|---|---|
| Amino acids | ||
| Butanoate 2,4-dihydroxy | 1.34 | 1.38 |
| Glu | 1.25 | 1.45 |
| Organic acid | ||
| Maleate | 0.63 | 0.67 |
| Malate | 0.32 | 0.50 |
| Phosphorate | 1.23 | 1.51 |
| Glutarate | 0.81 | 1.11 |
| 4-hydroxy cis-cinnamic acid | 1.43 | 1.26 |
| Quinate | 0.58 | 0.33 |
| 5-caffeoyl trans-quinate | 33.83 | 13.65 |
| Threonate | 0.45 | 0.37 |
| Ascorbate | 1.24 | 1.11 |
| Glucuronate | 0.52 | 0.41 |
| Galactonate | 0.70 | 0.75 |
| Caffeate | 0.99 | 0.82 |
| Others | ||
| catechin | 0.50 | 0.87 |
| Epigallocatechin | 1.49 | 2.37 |
| Epicatechin | 0.60 | 0.93 |
| Sugars | ||
| Glucose | 0.71 | 0.68 |
| Raffinose | 0.56 | 0.97 |
| Xylose | 0.74 | 0.72 |
| Lyxose | 0.84 | 0.81 |
| Ribulose | 1.92 | 1.61 |
| Variety | # of Nodes | # of Edges | Common Edges | Ø | Transitivity | Modularity | ||
|---|---|---|---|---|---|---|---|---|
| Biochemical | Walk-Trap | Edge Betweenness | ||||||
| Cs | 48 | 112 | 28 (25%) | 10 | 0.4 | 6 (−0.01) | 9 (0.5) | 7 (0.5) |
| Sh | 48 | 137 | 28 (20%) | 9 | 0.5 | 6 (0.1) | 11 (0.4) | 11 (0.4) |
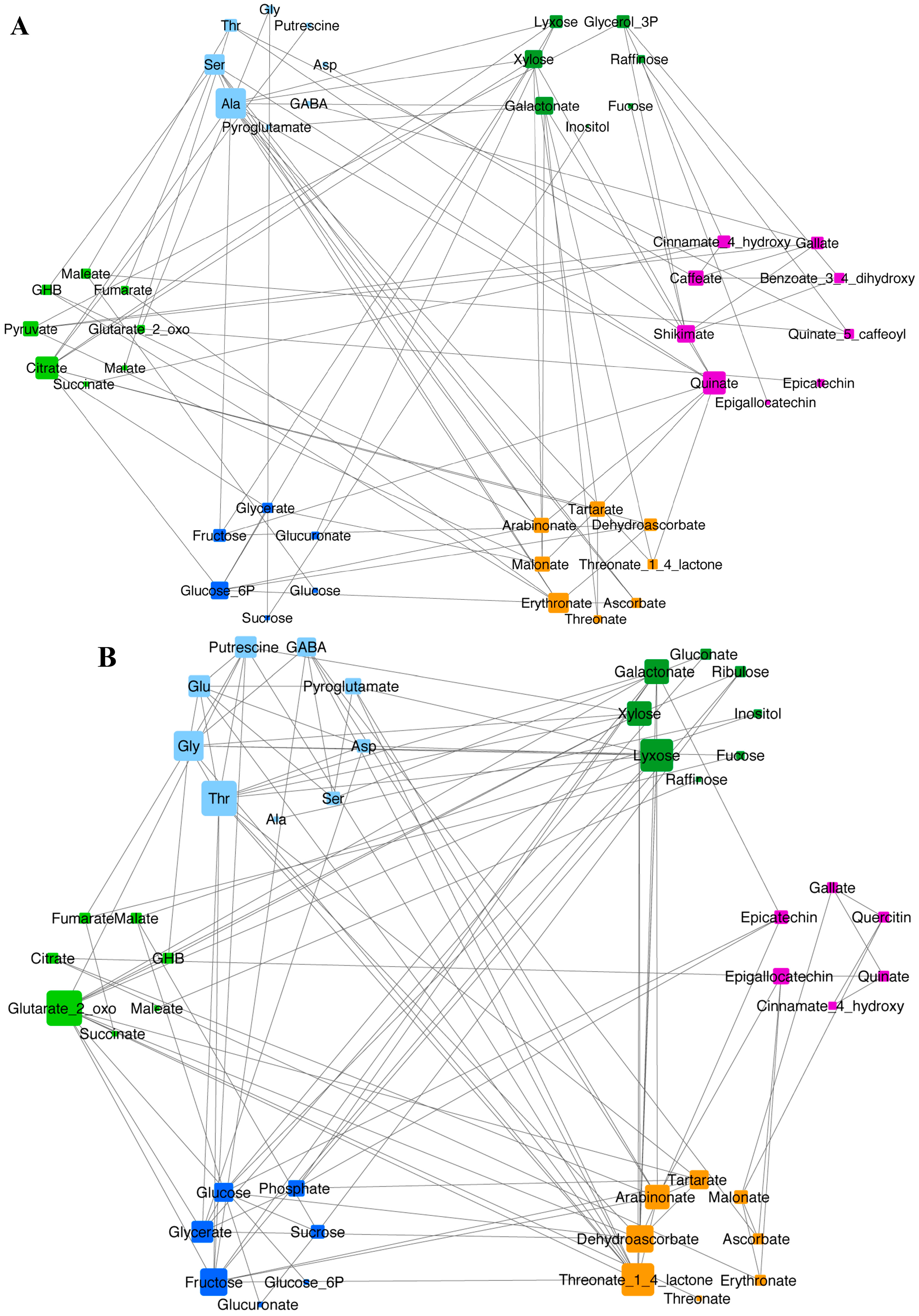
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Treatments and Experimental Designs
4.3. Leaf Area
4.4. Gas Exchange and Chlorophyll Fluorescence Analysis
4.5. Leaf Sampling and Extraction for Metabolite Profiling
4.6. GC–MS Derivatization and Data Processing
4.7. Network Analysis
4.8. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
References
- Arroyo-Garcia, R.; Ruiz-Garcia, L.; Bolling, L.; Ocete, R.; Lopez, M.; Arnold, C.; Ergul, A.; Uzun, H.; Cabello, F.; Ibáñez, J. Multiple origins of cultivated grapevine (Vitis vinifera L. ssp. sativa) based on chloroplast DNA polymorphisms. Mol. Ecol. 2006, 15, 3707–3714. [Google Scholar] [CrossRef] [PubMed]
- Webb, L.; Whetton, P.; Barlow, E. Modelled impact of future climate change on the phenology of winegrapes in Australia. Aust. J. Grape Wine Res. 2007, 13, 165–175. [Google Scholar] [CrossRef]
- Jones, G.V.; White, M.A.; Cooper, O.R.; Storchmann, K. Climate change and global wine quality. Clim. Chang. 2005, 73, 319–343. [Google Scholar] [CrossRef]
- Matsui, S.; Ryugo, K.; Kliewer, W. Growth inhibition of Thompson Seedless and Napa Gamay berries by heat stress and its partial reversibility by applications of growth regulators. Am. J. Enol. Vitic. 1986, 37, 67–71. [Google Scholar]
- Mori, K.; Sugaya, S.; Gemma, H. Decreased anthocyanin biosynthesis in grape berries grown under elevated night temperature condition. Sci. Hortic. 2005, 105, 319–330. [Google Scholar] [CrossRef]
- Mori, K.; Goto-Yamamoto, N.; Kitayama, M.; Hashizume, K. Loss of anthocyanins in red-wine grape under high temperature. J. Exp. Bot. 2007, 58, 1935–1945. [Google Scholar] [CrossRef] [PubMed]
- Sadras, V.O. Does partial root-zone drying improve irrigation water productivity in the field? A meta-analysis. Irrig. Sci. 2009, 27, 183–190. [Google Scholar] [CrossRef]
- Soar, C.J.; Collins, M.J.; Sadras, V.O. Irrigated Shiraz vines (Vitis vinifera) upregulate gas exchange and maintain berry growth in response to short spells of high maximum temperature in the field. Funct. Plant Biol. 2009, 36, 801–814. [Google Scholar] [CrossRef]
- Hendrickson, L.; Ball, M.; Wood, J.; Chow, W.; Furbank, R. Low temperature effects on photosynthesis and growth of grapevine. Plant Cell Environ. 2004, 27, 795–809. [Google Scholar] [CrossRef]
- Kadir, S.; von Weihe, M.; Al-Khatib, K. Photochemical efficiency and recovery of photosystem II in grapes after exposure to sudden and gradual heat stress. J. Am. Soc. Hortic. Sci. 2007, 132, 764–769. [Google Scholar]
- Luo, H.B.; Ma, L.; Xi, H.F.; Duan, W.; Li, S.H.; Loescher, W.; Wang, J.F.; Wang, L.J. Photosynthetic responses to heat treatments at different temperatures and following recovery in grapevine (Vitis amurensis L.) leaves. PLoS ONE 2011, 6, e23033. [Google Scholar] [CrossRef] [PubMed]
- Schultz, H. Climate change and viticulture: A European perspective on climatology, carbon dioxide and UV-B effects. Aust. J. Grape Wine Res. 2008, 6, 2–12. [Google Scholar] [CrossRef]
- Salem-Fnayou, A.B.; Bouamama, B.; Ghorbel, A.; Mliki, A. Investigations on the leaf anatomy and ultrastructure of grapevine (Vitis vinifera) under heat stress. Microsc. Res. Tech. 2011, 74, 756–762. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, L.C.; Vidigal, P.; Amâncio, S. Oxidative stress homeostasis in grapevine (Vitis vinifera L.). Front. Environ. Sci. 2015, 3, 20. [Google Scholar] [CrossRef]
- Greer, D.H.; Weston, C.; Weedon, M. Shoot architecture, growth and development dynamics of Vitis vinifera cv. Semillon vines grown in an irrigated vineyard with and without shade covering. Funct. Plant Biol. 2010, 37, 1061–1070. [Google Scholar] [CrossRef]
- Chaves, M.M.; Harley, P.C.; Tenhunen, J.D.; Lange, O.L. Gas exchange studies in two Portuguese grapevine cultivars. Physiol. Plant. 1987, 70, 639–647. [Google Scholar] [CrossRef]
- Kadir, S. Thermostability of photosynthesis of Vitis aestivalis and V. vinifera. J. Am. Soc. Hortic. Sci. 2006, 131, 476–483. [Google Scholar]
- Atkin, O.K.; Bruhn, D.; Hurry, V.M.; Tjoelker, M.G. Evans Review No. 2: The hot and the cold: Unravelling the variable response of plant respiration to temperature. Funct. Plant Biol. 2005, 32, 87–105. [Google Scholar] [CrossRef]
- Huang, B.; Rachmilevitch, S.; Xu, J. Root carbon and protein metabolism associated with heat tolerance. J. Exp. Bot. 2012, 63, 3455–3465. [Google Scholar] [CrossRef] [PubMed]
- Budyko, M.I. The heat balance of the earth’s surface. In Izvestya Akademii Nauk SSSR. Seriya Geograficheskaya; Department of Commerce, Weather Bureau: Washington, WA, USA, 1962. [Google Scholar]
- Hochberg, U.; Degu, A.; Cramer, G.R.; Rachmilevitch, S.; Fait, A. Cultivar specific metabolic changes in grapevines berry skins in relation to deficit irrigation and hydraulic behavior. Plant Physiol. Biochem. 2015, 88, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Degu, A.; Hochberg, U.; Sikron, N.; Venturini, L.; Buson, G.; Ghan, R.; Plaschkes, I.; Batushansky, A.; Chalifa-Caspi, V.; Mattivi, F. Metabolite and transcript profiling of berry skin during fruit development elucidates differential regulation between Cabernet Sauvignon and Shiraz cultivars at branching points in the polyphenol pathway. BMC Plant Biol. 2014, 14, 188. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, U.; Degu, A.; Toubiana, D.; Gendler, T.; Nikoloski, Z.; Rachmilevitch, S.; Fait, A. Metabolite profiling and network analysis reveal coordinated changes in grapevine water stress response. BMC Plant Biol. 2013, 13, 184. [Google Scholar] [CrossRef] [PubMed]
- Guy, C.; Kaplan, F.; Kopka, J.; Selbig, J.; Hincha, D.K. Metabolomics of temperature stress. Physiol. Plant. 2007, 132, 220–235. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Gerstein, M.; Snyder, M. Getting connected: Analysis and principles of biological networks. Genes Dev. 2007, 21, 1010–1024. [Google Scholar] [CrossRef]
- Bota, J.; Flexas, J.; Medrano, H. Genetic variability of photosynthesis and water use in Balearic grapevine cultivars. Ann. Appl. Biol. 2001, 138, 353–361. [Google Scholar] [CrossRef]
- Santesteban, L.; Miranda, C.; Royo, J. Effect of water deficit and rewatering on leaf gas exchange and transpiration decline of excised leaves of four grapevine (Vitis vinifera L.) cultivars. Sci. Hortic. 2009, 121, 434–439. [Google Scholar] [CrossRef]
- Lobell, D.B.; Field, C.B.; Cahill, K.N.; Bonfils, C. Impacts of future climate change on California perennial crop yields: Model projections with climate and crop uncertainties. Agric. For. Meteorol. 2006, 141, 208–218. [Google Scholar] [CrossRef]
- Hannah, L.; Roehrdanz, P.R.; Ikegami, M.; Shepard, A.V.; Shaw, M.R.; Tabor, G.; Zhi, L.; Marquet, P.A.; Hijmans, R.J. Climate change, wine, and conservation. Proc. Natl. Acad. Sci. USA 2013, 110, 6907–6912. [Google Scholar] [CrossRef] [PubMed]
- Cramer, G.R.; Ergül, A.; Grimplet, J.; Tillett, R.L.; Tattersall, E.A.R.; Bohlman, M.C.; Vincent, D.; Sonderegger, J.; Evans, J.; Osborne, C. Water and salinity stress in grapevines: Early and late changes in transcript and metabolite profiles. Funct. Integr. Genom. 2007, 7, 111–134. [Google Scholar] [CrossRef] [PubMed]
- Rachmilevitch, S.; Cohen, I.; Huang, B. Carbon allocation patterns into proteins and lipids associated with superior tolerance of perennial grass to high soil temperature. Crop Sci. 2015. [Google Scholar] [CrossRef]
- Kaplan, F.; Kopka, J.; Haskell, D.W.; Zhao, W.; Schiller, K.C.; Gatzke, N.; Sung, D.Y.; Guy, C.L. Exploring the temperature-stress metabolome of Arabidopsis. Plant Physiol. 2004, 136, 4159–4168. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Wang, Z.; Yu, W.; Liu, Y.; Huang, B. Differential metabolic responses of perennial grass Cynodon transvaalensis × Cynodon dactylon (C4) and Poa pratensis (C3) to heat stress. Physiol. Plant. 2011, 141, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Araújo, W.L.; Nunes-Nesi, A.; Trenkamp, S.; Bunik, V.I.; Fernie, A.R. Inhibition of 2-oxoglutarate dehydrogenase in potato tuber suggests the enzyme is limiting for respiration and confirms its importance in nitrogen assimilation. Plant Physiol. 2008, 148, 1782–1796. [Google Scholar] [CrossRef] [PubMed]
- Gibson, S.I. Control of plant development and gene expression by sugar signaling. Curr. Opin. Plant Biol. 2005, 8, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Couée, I.; Sulmon, C.; Gouesbet, G.; el Amrani, A. Involvement of soluble sugars in reactive oxygen species balance and responses to oxidative stress in plants. J. Exp. Bot. 2006, 57, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Sepúlveda, G.; Kliewer, W.M. Effect of high temperature on grapevines (Vitis vinifera L.). II. Distribution of soluble sugars. Am. J. Enol. Vitic. 1986, 37, 20–25. [Google Scholar]
- Quick, W.; Chaves, M.; Wendler, R.; David, M.; Rodrigues, M.; Passaharinho, J.; Pereira, J.; Adcock, M.; Leegood, R.; Stitt, M. The effect of water stress on photosynthetic carbon metabolism in four species grown under field conditions. Plant Cell Environ. 1992, 15, 25–35. [Google Scholar] [CrossRef]
- Katja, H.; Irina, B.; Hiie, I.; Olav, K.; Tiit, P.; Bahtijor, R.; Mari, T.; Ülo, N. Temperature responses of dark respiration in relation to leaf sugar concentration. Physiol. Plant. 2012, 144, 320–334. [Google Scholar] [CrossRef] [PubMed]
- Duff, D.T.; Beard, J.B. Supraoptimal temperature effects upon Agrostis palustris. Physiol. Plant. 1974, 32, 18–22. [Google Scholar] [CrossRef]
- Yamakawa, H.; Hakata, M. Atlas of rice grain filling-related metabolism under high temperature: Joint analysis of metabolome and transcriptome demonstrated inhibition of starch accumulation and induction of amino acid accumulation. Plant Cell Physiol. 2010, 51, 795–809. [Google Scholar] [CrossRef] [PubMed]
- Rizhsky, L.; Liang, H.; Shuman, J.; Shulaev, V.; Davletova, S.; Mittler, R. When defense pathways collide. The response of Arabidopsis to a combination of drought and heat stress. Plant Physiol. 2004, 134, 1683–1696. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.; Chaves, M.; Wendler, R.; David, M.; Quick, W.; Leegood, R.; Stitt, M.; Pereira, J. Osmotic adjustment in water stressed grapevine leaves in relation to carbon assimilation. Funct. Plant Biol. 1993, 20, 309–321. [Google Scholar] [CrossRef]
- Patakas, A.; Nikolaou, N.; Zioziou, E.; Radoglou, K.; Noitsakis, B. The role of organic solute and ion accumulation in osmotic adjustment in drought-stressed grapevines. Plant Sci. 2002, 163, 361–367. [Google Scholar] [CrossRef]
- Urano, K.; Kurihara, Y.; Seki, M.; Shinozaki, K. “Omics” analyses of regulatory networks in plant abiotic stress responses. Curr. Opin. Plant Biol. 2010, 13, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Hernández, I.; Alegre, L.; van Breusegem, F.; Munné-Bosch, S. How relevant are flavonoids as antioxidants in plants? Trends Plant Sci. 2009, 14, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Locato, V.; Gadaleta, C.; de Gara, L.; de Pinto, M.C. Production of reactive species and modulation of antioxidant network in response to heat shock: A critical balance for cell fate. Plant Cell Environ. 2008, 31, 1606–1619. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, J.C.; López-Cobollo, R.; Alcázar, R.; Zarza, X.; Koncz, C.; Altabella, T.; Salinas, J.; Tiburcio, A.F.; Ferrando, A. Putrescine is involved in Arabidopsis freezing tolerance and cold acclimation by regulating abscisic acid levels in response to low temperature. Plant Physiol. 2008, 148, 1094–1105. [Google Scholar] [CrossRef]
- Ioannidis, N.E.; Cruz, J.A.; Kotzabasis, K.; Kramer, D.M. Evidence that putrescine modulates the higher plant photosynthetic proton circuit. PLoS ONE 2012, 7, e29864. [Google Scholar] [CrossRef] [PubMed]
- Olivo, N.; Girona, J.; Marsal, J. Seasonal sensitivity of stem water potential to vapour pressure deficit in grapevine. Irrig. Sci. 2009, 27, 175–182. [Google Scholar] [CrossRef]
- Rogiers, S.Y.; Greer, D.H.; Hatfield, J.M.; Hutton, R.J.; Clarke, S.J.; Hutchinson, P.A.; Somers, A. Stomatal response of an anisohydric grapevine cultivar to evaporative demand, available soil moisture and abscisic acid. Tree Physiol. 2012, 32, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Feller, U. Stomatal opening at elevated temperature: An underestimated regulatory mechanism. Gen. Appl. Plant Physiol. 2006, 32, 19–31. [Google Scholar]
- Nonami, H.; Schulze, E.; Ziegler, H. Mechanisms of stomatal movement in response to air humidity, irradiance and xylem water potential. Planta 1991, 183, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Sepúlveda, G.; Kliewer, W.M. Stomatal response of three grapevine cultivars (Vitis vinifera L.) to high temperature. Am. J. Enol. Vitic. 1986, 37, 44–52. [Google Scholar]
- Hochberg, U.; Degu, A.; Fait, A.; Rachmilevitch, S. Near isohydric grapevine cultivar displays higher photosynthetic efficiency and photorespiration rates under drought stress as compared with near anisohydric grapevine cultivar. Physiol. Plant. 2013, 147, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Rapaport, T.; Hochberg, U.; Rachmilevitch, S.; Karnieli, A. The effect of differential growth rates across plants on spectral predictions of physiological parameters. PLoS ONE 2014, 9, e88930. [Google Scholar] [CrossRef] [PubMed]
- Weckwerth, W.; Wenzel, K.; Fiehn, O. Process for the integrated extraction, identification and quantification of metabolites, proteins and RNA to reveal their co-regulation in biochemical networks. Proteomics 2004, 4, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Lisec, J.; Schauer, N.; Kopka, J.; Willmitzer, L.; Fernie, A.R. Gas chromatography mass spectrometry-based metabolite profiling in plants. Nat. Protoc. 2006, 1, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Kopka, J.; Fernie, A.; Weckwerth, W.; Gibon, Y.; Stitt, M. Metabolite profiling in plant biology: Platforms and destinations. Genom. Biol. 2004, 5, 109. [Google Scholar] [CrossRef] [PubMed]
- Golm Metabolom Databases. Available online: http://gmd.mpimp-golm.mpg.de/ (accessed on 25 July 2015).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Saito, R.; Smoot, M.E.; Ono, K.; Ruscheinski, J.; Wang, P.; Lotia, S.; Pico, A.R.; Bader, G.D.; Ideker, T. A travel guide to Cytoscape plugins. Nat. Methods 2012, 9, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Igraph. Available online: http://igraph.org/r/ (accessed on 25 July 2015).
- Saeed, A.I.; Bhagabati, N.K.; Braisted, J.C.; Liang, W.; Sharov, V.; Howe, E.A.; Li, J.; Thiagarajan, M.; White, J.A.; Quackenbush, J. [9] TM4 Microarray Software Suite. Methods Enzymol. 2006, 411, 134–193. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hochberg, U.; Batushansky, A.; Degu, A.; Rachmilevitch, S.; Fait, A. Metabolic and Physiological Responses of Shiraz and Cabernet Sauvignon (Vitis vinifera L.) to Near Optimal Temperatures of 25 and 35 °C. Int. J. Mol. Sci. 2015, 16, 24276-24294. https://doi.org/10.3390/ijms161024276
Hochberg U, Batushansky A, Degu A, Rachmilevitch S, Fait A. Metabolic and Physiological Responses of Shiraz and Cabernet Sauvignon (Vitis vinifera L.) to Near Optimal Temperatures of 25 and 35 °C. International Journal of Molecular Sciences. 2015; 16(10):24276-24294. https://doi.org/10.3390/ijms161024276
Chicago/Turabian StyleHochberg, Uri, Albert Batushansky, Asfaw Degu, Shimon Rachmilevitch, and Aaron Fait. 2015. "Metabolic and Physiological Responses of Shiraz and Cabernet Sauvignon (Vitis vinifera L.) to Near Optimal Temperatures of 25 and 35 °C" International Journal of Molecular Sciences 16, no. 10: 24276-24294. https://doi.org/10.3390/ijms161024276
APA StyleHochberg, U., Batushansky, A., Degu, A., Rachmilevitch, S., & Fait, A. (2015). Metabolic and Physiological Responses of Shiraz and Cabernet Sauvignon (Vitis vinifera L.) to Near Optimal Temperatures of 25 and 35 °C. International Journal of Molecular Sciences, 16(10), 24276-24294. https://doi.org/10.3390/ijms161024276