

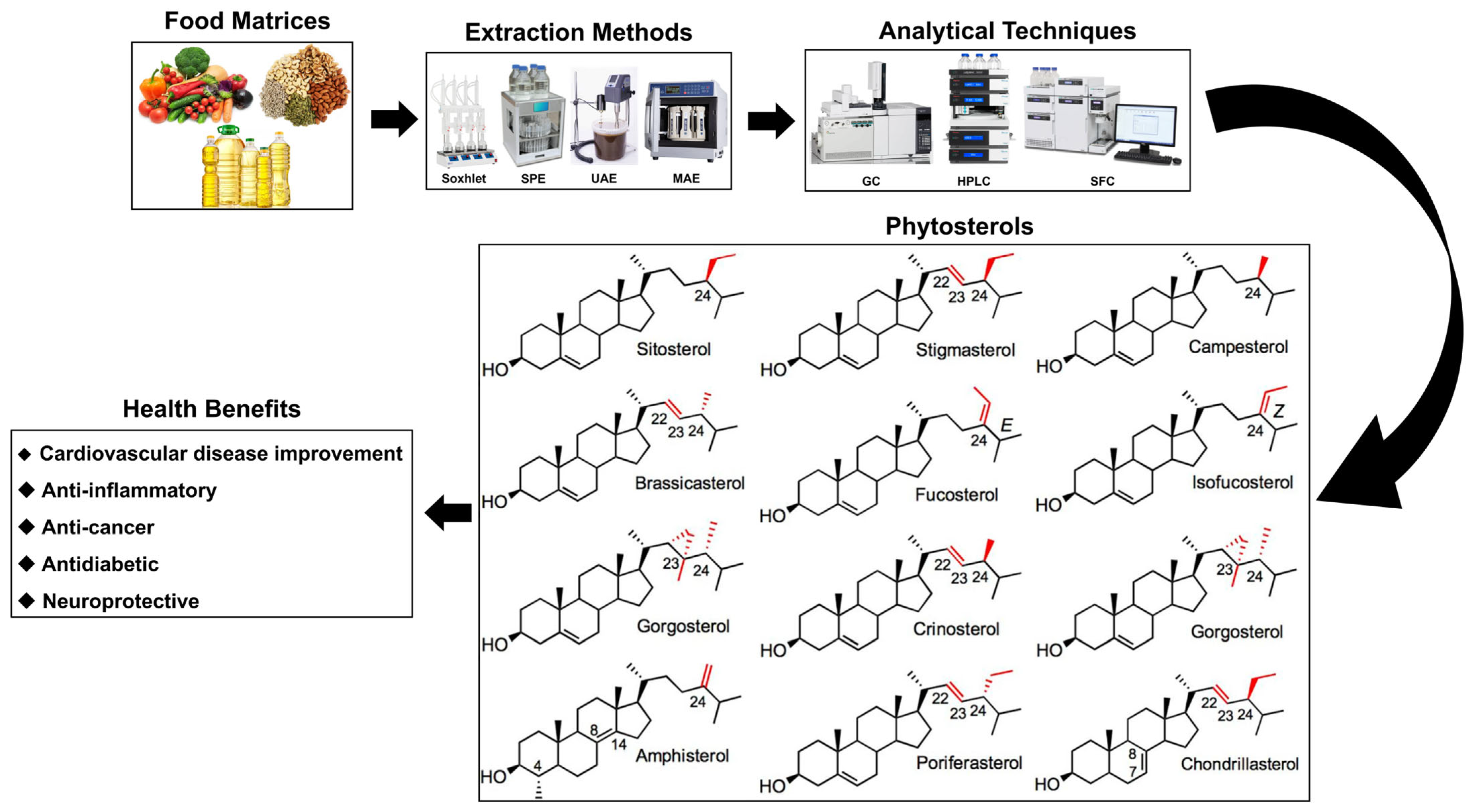
Phytosterols: Extraction Methods, Analytical Techniques, and Biological Activity

Abstract
1. Introduction
2. Extraction Methods and Analytical Techniques for Phytosterols
2.1. Conventional Extraction Methods
2.1.1. Solid–Liquid Extraction (SLE)
2.1.2. Soxhlet Extraction
2.1.3. Saponification
2.1.4. Maceration
2.2. Recent Extraction Methods
2.2.1. Ultrasonic-Assisted Extraction (UAE)
2.2.2. Microwave-Assisted Extraction (MAE)
2.2.3. Pressurized Liquid Extraction (PLE)
2.2.4. Solid-Phase Extraction (SPE)
2.2.5. Solid-Phase Microextraction (SPME)
2.2.6. Liquid-Phase Extraction
2.3. Phytosterol Analysis Techniques
2.3.1. Liquid Chromatography
2.3.2. Gas Chromatography (GC)
2.3.3. Supercritical Fluid Chromatography (SFC)

{kind=link}
{kind=link}
| Sample Matrix | Analyzed Phytosterols | Extraction Methods | Determination Methods | Extract Yield (mg/g) | References |
|---|---|---|---|---|---|
| Linseed oil | β-Sitosterol | Saponification, LLE with diethyl ether | RPLC-SALDI MS | 2.680 | [86] |
| Stigmasterol | 0.850 | ||||
| Campesterol | 1.670 | ||||
| Brassicasterol | 0.680 | ||||
| Niger seed oil | β-Sitosterol | Saponification, LLE with diethyl ether, SPE | TMS-derivatized samples were analyzed by GC-MS | 2.779 | [29] |
| Stigmasterol | 0.920, | ||||
| Campesterol | 0.870 | ||||
| Cycloartenol | 0.131 | ||||
| P. lactiflora Pall seed oils | Campesterol | Maceration with n-hexane, UAE | HPLC | 0.37345 | [87] |
| Stigmasterol | 0.04596 | ||||
| β-Sitosterol | 2.47376 | ||||
| Isofucosterol | 0.2347 | ||||
| Δ7-Avenasterol | 0.10951 | ||||
| P. lactiflora Pall seed oils | Campesterol | SFE | HPLC | 0.31535 | [87] |
| Stigmasterol | 0.02383 | ||||
| β-Sitosterol | 3.28167 | ||||
| Isofucosterol | 0.37213 | ||||
| Δ7-Avenasterol | 0.17138 | ||||
| P. lactiflora Pall seed oils | Campesterol | Pressing | HPLC | 0.32799 | [87] |
| Stigmasterol | 0.04674 | ||||
| β-Sitosterol | 2.7845 | ||||
| Isofucosterol | 0.30375 | ||||
| Δ7-Avenasterol | 0.14032 | ||||
| Olive drupes | Brassicasterol | UAE, saponification, LLE with hexane, SPE | TMS-derivatized samples were analyzed by GC-MS | 0.004 | [88] |
| Campesterol | 0.0068 | ||||
| Stigmasterol | 0.0198 | ||||
| β-Sitosterol | 2.3997 | ||||
| Canola oil (Richfood) | β-Sitosterol | Saponification, LLE with n-hexane | APCI-LC-MS/MS | 3.597 | [74] |
| Campesterol | 1.837 | ||||
| Brassicasterol | 0.4879 | ||||
| Stigmasterol | 0.034 | ||||
| Cycloartenol | 0.087 | ||||
| Lupeol | <LLOQ | ||||
| Olive oil (Richfood) | β-Sitosterol | Saponification, LLE with n-hexane | APCI-LC-MS/MS | 1.209 | [74] |
| Campesterol | 0.0542 | ||||
| Brassicasterol | <LLOQ | ||||
| Stigmasterol | 0.0236 | ||||
| Cycloartenol | 0.338 | ||||
| Lupeol | 0.0196 | ||||
| Walnut oil | Campesterol | In situ direct analysis | SFC-QTOF-MS | 0.03181 | [85] |
| β-Sitosterol | 0.86884 | ||||
| Stigmasterol | 0.04065 | ||||
| Hemp oil (Living Harvest) | β-Sitosterol | Saponification, LLE with n-hexane | APCI-LC-MS/MS | 3.16 | [74] |
| Campesterol | 0.706 | ||||
| Brassicasterol | <LLOQ | ||||
| Stigmasterol | 0.1136 | ||||
| Cycloartenol | 0.169 | ||||
| Lupeol | <LLOQ | ||||
| Almond oil (International Collection) | β-Sitosterol | Saponification, LLE with n-hexane | APCI-LC-MS/MS | 0.940 | [74] |
| Campesterol | 0.1842 | ||||
| Brassicasterol | 0.0146 | ||||
| Stigmasterol | 0.1569 | ||||
| Cycloartenol | 0.139 | ||||
| Lupeol | 0.0195 | ||||
| Hazelnut oil (International Collection) | β-Sitosterol | Saponification, LLE with n-hexane | APCI-LC-MS/MS | 0.472 | [74] |
| Campesterol | 0.301 | ||||
| Brassicasterol | 0.0015 | ||||
| Stigmasterol | 0.0097 | ||||
| Cycloartenol | 0.0054 | ||||
| Lupeol | 0.0035 | ||||
| Walnut oil (International Collection) | β-Sitosterol | Saponification, LLE with n-hexane | APCI-LC-MS/MS | 1.222 | [74] |
| Campesterol | 0.0899 | ||||
| Brassicasterol | 0.0026 | ||||
| Stigmasterol | 0.00131 | ||||
| Cycloartenol | 0.244 | ||||
| Lupeol | <LLOQ | ||||
| Almonds | β-Sitosterol | PLE with methanol | LC-DAD | 1.16 ± 15 | [48] |
| Grape seed oil (International Collection) | β-Sitosterol | Saponification, LLE with n-hexane | APCI-LC-MS/MS | 1.913 | [74] |
| Campesterol | 0.3935 | ||||
| Brassicasterol | <LLOQ | ||||
| Stigmasterol | 0.340 | ||||
| Cycloartenol | 0.2617 | ||||
| Lupeol | 0.0285 | ||||
| Sesame oil (Spectrum) | β-Sitosterol | Saponification, LLE with n-hexane | APCI-LC-MS/MS | 3.215 | [74] |
| Campesterol | 0.880 | ||||
| Brassicasterol | <LLOQ | ||||
| Stigmasterol | 0.418 | ||||
| Cycloartenol | 0.1564 | ||||
| Lupeol | 0.0128 | ||||
| Avocado oil (Spectrum) | β-Sitosterol | Saponification, LLE with n-hexane | APCI-LC-MS/MS | 2.629 | [74] |
| Campesterol | 0.4043 | ||||
| Brassicasterol | <LLOQ | ||||
| Stigmasterol | 0.1726 | ||||
| Cycloartenol | 0.3471 | ||||
| Lupeol | 0.0181 | ||||
| Sunflower oil (Richfood) | β-Sitosterol | Saponification, LLE with n-hexane | APCI-LC-MS/MS | 2.178 | [74] |
| Campesterol | 0.3178 | ||||
| Brassicasterol | 0.0135 | ||||
| Stigmasterol | 0.301 | ||||
| Cycloartenol | 0.1745 | ||||
| Lupeol | 0.0302 | ||||
| Peanut oil (Richfood) | β-Sitosterol | Saponification, LLE with n-hexane | APCI-LC-MS/MS | 1.030 | [74] |
| Campesterol | 0.278 | ||||
| Brassicasterol | 0.019 | ||||
| Stigmasterol | 0.1541 | ||||
| Cycloartenol | 0.0582 | ||||
| Lupeol | 0.0128 | ||||
| Corn oil (Richfood) | β-Sitosterol | Saponification, LLE with n-hexane | APCI-LC-MS/MS | 4.354 | [74] |
| Campesterol | 1.555 | ||||
| Brassicasterol | <LLOQ | ||||
| Stigmasterol | 0.703 | ||||
| Cycloartenol | 0.2731 | ||||
| Lupeol | <LLOQ | ||||
| Macadamia nut oil (Olivado) | β-Sitosterol | Saponification, LLE with n-hexane | APCI-LC-MS/MS | 1.915 | [74] |
| Campesterol | 0.1378 | ||||
| Brassicasterol | <LLOQ | ||||
| Stigmasterol | 0.0071 | ||||
| Cycloartenol | 0.0085 | ||||
| Lupeol | <LLOQ | ||||
| Sunflower oil (Hollywood) | β-Sitosterol | Saponification, LLE with n-hexane | APCI-LC-MS/MS | 1.349 | [74] |
| Campesterol | 0.369 | ||||
| Brassicasterol | 0.0111 | ||||
| Stigmasterol | 0.1594 | ||||
| Cycloartenol | 0.14 | ||||
| Lupeol | 0.042 | ||||
| U. pinnatifida | Fucosterol | MAE, saponification, LLE with n-hexane | HPLC-UV and GC-MS | 1.21 | [44] |
| 24-Methylenecholesterol | 0.16 | ||||
| Coconut oil | Brassicasterol | Solvent extraction with heptane, saponification | SFC-APCI-MS/MS | 9.41 ± 0.39 | [84] |
| Campesterol | 4.34 ± 0.33 | ||||
| δ-5-Acenasterol | 20.94 ± 0.60 | ||||
| Stigmasterol | 21.51 ± 0.46 | ||||
| β-Sitosterol | 32.71 ± 2.75 | ||||
| Lupeol | 18.89 ± 0.89 | ||||
| Cycloartenol | 18.97 ± 1.26 | ||||
| Palm oil | Brassicasterol | Solvent extraction with heptane, saponification | SFC-APCI-MS/MS | 4.93 ± 0.76 | [84] |
| Campesterol | 4.14 ± 0.12 | ||||
| δ-5-Acenasterol | 12.70 ± 0.82 | ||||
| Stigmasterol | 13.20 ± 0.92 | ||||
| β-Sitosterol | 31.97 ± 0.28 | ||||
| Lupeol | 16.30 ± 0.45 | ||||
| Cycloartenol | 18.49 ± 0.26 |
3. Health Benefits of Phytosterols
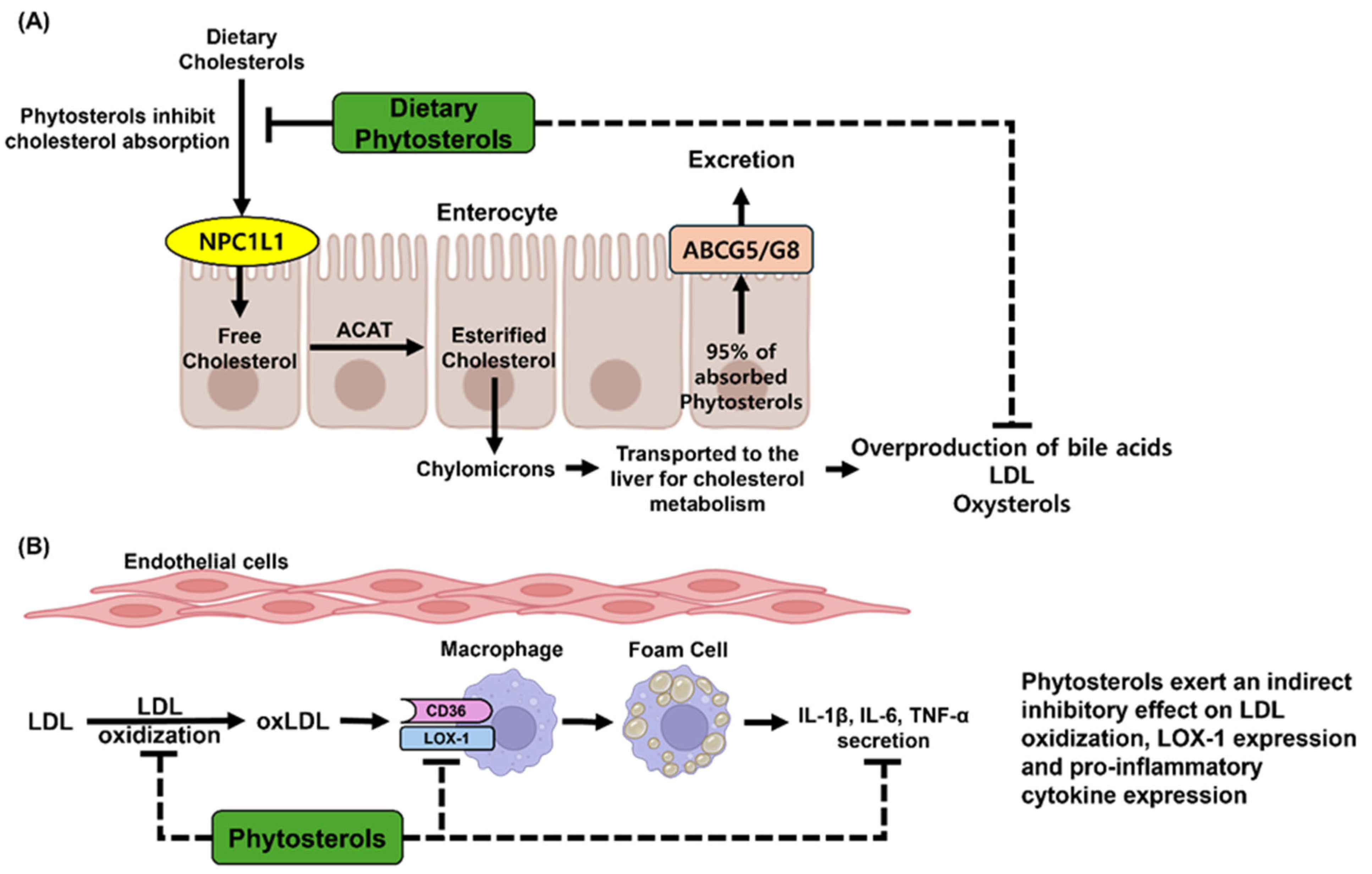
3.1. Cardiovascular Diseases (CVDs) and Phytosterols
3.2. Anti-Inflammatory Effects of Phytosterols
3.3. Anticancer Effects of Phytosterols
3.4. Antidiabetic Effects of Phytosterols
3.5. Neuroprotective Effects of Phytosterols
4. Bioavailability of Phytosterols
5. Conclusions
6. Methodology
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABCG5/G8 | ATP-binding cassette transporter G5 |
| AChE | Acetylcholinesterase |
| AD | Alzheimer’s disease |
| APP | Amyloid precursor protein |
| Aβ | Amyloid-β |
| BACE1 | β-Site amyloid precursor protein-cleaving enzyme 1 |
| BBB | Blood–brain barrier |
| BDNF | Brain-derived neurotrophic factor |
| COX-2 | Cyclooxygenase-2 |
| CVDs | Cardiovascular diseases |
| DLLME | Dispersive liquid–liquid microextraction |
| DMBA | 7,12-Dimethylbenz[a]anthracene |
| DSS | Dextran sulfate sodium |
| DSS | Dextran sodium sulfate |
| ER | Reticulum stress |
| ERK | Extracellular signal-regulated kinase |
| FPG | Fasting plasma glucose |
| GC | Gas chromatography |
| GLUT4 | Glucose transporter 4 |
| GSH | Glutathione |
| HDACs | Histone deacetylases |
| HDL | High-density lipoprotein |
| HOMA-IR | Homeostatic model assessment for insulin resistance |
| HPLC | High-performance liquid chromatography |
| IBD | Inflammatory bowel disease |
| IBD | Inflammatory bowel disease |
| iNOS | Inducible nitric oxide synthase |
| LDL-C | Low-density lipoprotein cholesterol |
| LLE | Liquid–liquid extraction |
| LLME | Liquid–liquid microextraction |
| LPS | Lipopolysaccharide |
| MAE | Microwave-assisted extraction |
| MAPK | Mitogen-activated protein kinase |
| MPO | Myeloperoxidase |
| NF-κB | Nuclear factor-κ B |
| NO | Nitric oxide |
| NPC1L1 | Niemann-Pick C1-like 1 |
| oxLDL | Oxidized low-density lipoprotein |
| PGE2 | Prostaglandin E2 |
| PLE | Pressurized liquid extraction |
| QUICKI | Quantitative insulin sensitivity check index |
| ROS | Reactive oxygen species |
| SFC | Supercritical fluid chromatography |
| SLE | Solid–liquid extraction |
| SPE | Solid-phase extraction |
| SPME | Solid-phase microextraction |
| TG | Triglyceride |
| TC | Total cholesterol |
| TNF-α | Tumor necrosis factor-α |
| TrkB-ERK1/2 | Tropomyosin receptor kinase B–extracellular signal-regulated kinase 1/2 |
| UAE | Ultrasonic-assisted extraction |
References
- Beyene, S.D. The impact of food insecurity on health outcomes: Empirical evidence from sub-Saharan African countries. BMC Public Health 2023, 23, 338. [Google Scholar] [CrossRef] [PubMed]
- Baker, P.; Machado, P.; Santos, T.; Sievert, K.; Backholer, K.; Hadjikakou, M.; Russell, C.; Huse, O.; Bell, C.; Scrinis, G.; et al. Ultra-processed foods and the nutrition transition: Global, regional and national trends, food systems transformations and political economy drivers. Obes. Rev. 2020, 21, e13126. [Google Scholar] [CrossRef] [PubMed]
- Orlien, V.; Bolumar, T. Biochemical and Nutritional Changes during Food Processing and Storage. Foods 2019, 8, 494. [Google Scholar] [CrossRef]
- Rarison, R.H.G.; Truong, V.L.; Yoon, B.H.; Park, J.W.; Jeong, W.S. Antioxidant and Anti-Inflammatory Mechanisms of Lipophilic Fractions from Polyscias fruticosa Leaves Based on Network Pharmacology, In Silico, and In Vitro Approaches. Foods 2023, 12, 3643. [Google Scholar] [CrossRef]
- Mao, Q.Q.; Xu, X.Y.; Shang, A.; Gan, R.Y.; Wu, D.T.; Atanasov, A.G.; Li, H.B. Phytochemicals for the Prevention and Treatment of Gastric Cancer: Effects and Mechanisms. Int. J. Mol. Sci. 2020, 21, 570. [Google Scholar] [CrossRef]
- Ellegård, L.H.; Andersson, S.W.; Normén, A.L.; Andersson, H.A. Dietary plant sterols and cholesterol metabolism. Nutr. Rev. 2007, 65, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.T.; Wen, B.; Shen, X.C.; Bian, Z.X. Potential of Plant-sourced Phenols for Inflammatory Bowel Disease. Curr. Med. Chem. 2018, 25, 5191–5217. [Google Scholar] [CrossRef]
- Sirotkin, A.V.; Kolesárová, A. The anti-obesity and health-promoting effects of tea and coffee. Physiol. Res. 2021, 70, 161–168. [Google Scholar] [CrossRef]
- Pagliaro, B.; Santolamazza, C.; Simonelli, F.; Rubattu, S. Phytochemical Compounds and Protection from Cardiovascular Diseases: A State of the Art. Biomed. Res. Int. 2015, 2015, 918069. [Google Scholar] [CrossRef]
- Makhmudova, U.; Schulze, P.C.; Lütjohann, D.; Weingärtner, O. Phytosterols and Cardiovascular Disease. Curr. Atheroscler. Rep. 2021, 23, 68. [Google Scholar] [CrossRef]
- Poli, A.; Marangoni, F.; Corsini, A.; Manzato, E.; Marrocco, W.; Martini, D.; Medea, G.; Visioli, F. Phytosterols, Cholesterol Control, and Cardiovascular Disease. Nutrients 2021, 13, 2810. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Quispe, C.; Sharifi-Rad, J.; Cruz-Martins, N.; Nigam, M.; Mishra, A.P.; Konovalov, D.A.; Orobinskaya, V.; Abu-Reidah, I.M.; Zam, W.; et al. Phytosterols: From Preclinical Evidence to Potential Clinical Applications. Front. Pharmacol. 2020, 11, 599959. [Google Scholar] [CrossRef] [PubMed]
- Othman, R.A.; Moghadasian, M.H. Beyond cholesterol-lowering effects of plant sterols: Clinical and experimental evidence of anti-inflammatory properties. Nutr. Rev. 2011, 69, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, N.; Khan, W.; Shadab, M.; Ali, A.; Saluja, S.S.; Sharma, S.; Al-Allaf, F.A.; Abduljaleel, Z.; Ibrahim, I.A.A.; Abdel-Wahab, A.F. Phytosterols as a natural anticancer agent: Current status and future perspective. Biomed. Pharmacother. 2017, 88, 786–794. [Google Scholar] [CrossRef]
- Yoshida, Y.; Niki, E. Antioxidant effects of phytosterol and its components. J. Nutr. Sci. Vitaminol. 2003, 49, 277–280. [Google Scholar] [CrossRef]
- Shen, M.; Yuan, L.; Zhang, J.; Wang, X.; Zhang, M.; Li, H.; Jing, Y.; Zeng, F.; Xie, J. Phytosterols: Physiological Functions and Potential Application. Foods 2024, 13, 1754. [Google Scholar] [CrossRef]
- Chang, Z.-Y.; Chen, C.-W.; Tsai, M.-J.; Chen, C.-C.; Alshetaili, A.; Hsiao, Y.-T.; Fang, J.-Y. The elucidation of structure–activity and structure-permeation relationships for the cutaneous delivery of phytosterols to attenuate psoriasiform inflammation. Int. Immunopharmacol. 2023, 119, 110202. [Google Scholar] [CrossRef]
- Salehi-Sahlabadi, A.; Varkaneh, H.K.; Shahdadian, F.; Ghaedi, E.; Nouri, M.; Singh, A.; Farhadnejad, H.; Găman, M.-A.; Hekmatdoost, A.; Mirmiran, P. Effects of Phytosterols supplementation on blood glucose, glycosylated hemoglobin (HbA1c) and insulin levels in humans: A systematic review and meta-analysis of randomized controlled trials. Diabetes Metab. J. 2020, 19, 625–632. [Google Scholar] [CrossRef]
- Dash, R.; Mitra, S.; Ali, M.C.; Oktaviani, D.F.; Hannan, M.A.; Choi, S.M.; Moon, I.S. Phytosterols: Targeting Neuroinflammation in Neurodegeneration. Curr. Pharm. Des. 2021, 27, 383–401. [Google Scholar] [CrossRef]
- Jesch, E.D.; Carr, T.P. Food Ingredients That Inhibit Cholesterol Absorption. Prev. Nutr. Food. Sci. 2017, 22, 67–80. [Google Scholar]
- Quitério, E.; Grosso, C.; Ferraz, R.; Delerue-Matos, C.; Soares, C. A Critical Comparison of the Advanced Extraction Techniques Applied to Obtain Health-Promoting Compounds from Seaweeds. Mar. Drugs 2022, 20, 677. [Google Scholar] [CrossRef] [PubMed]
- Osorio-Tobón, J.F. Recent advances and comparisons of conventional and alternative extraction techniques of phenolic compounds. J. Food Sci. Technol. 2020, 57, 4299–4315. [Google Scholar] [CrossRef] [PubMed]
- Roiaini, M.; Seyed, H.; Jinap, S.; Norhayati, H. Effect of extraction methods on yield, oxidative value, phytosterols and antioxidant content of cocoa butter. Int. Food Res. J. 2016, 23, 47. [Google Scholar]
- Nyam, K.L.; Tan, C.P.; Lai, O.M.; Long, K.; Che Man, Y.B. Optimization of supercritical CO 2 extraction of phytosterol-enriched oil from Kalahari melon seeds. Food Bioprocess Technol. 2011, 4, 1432–1441. [Google Scholar] [CrossRef]
- Lagarda, M.J.; García-Llatas, G.; Farré, R. Analysis of phytosterols in foods. J. Pharm. Biomed. Anal. 2006, 41, 1486–1496. [Google Scholar] [CrossRef]
- MS, U.; Ferdosh, S.; Haque Akanda, M.J.; Ghafoor, K.; AH, R.; Ali, M.E.; Kamaruzzaman, B.; MB, F.; Shaarani, S.; Islam Sarker, M.Z. Techniques for the extraction of phytosterols and their benefits in human health: A review. Sep. Sci. Technol 2018, 53, 2206–2223. [Google Scholar] [CrossRef]
- Chen, Z.; Shen, N.; Wu, X.; Jia, J.; Wu, Y.; Chiba, H.; Hui, S. Extraction and Quantitation of Phytosterols from Edible Brown Seaweeds: Optimization, Validation, and Application. Foods 2023, 12, 244. [Google Scholar] [CrossRef]
- Wang, S.-F.; Furuno, T.; Cheng, Z. Study of extraction of phytosterol from masson pine raw tall oil. J. Wood Sci. 2002, 48, 505–511. [Google Scholar] [CrossRef]
- Deme, T.; Haki, G.D.; Retta, N.; Woldegiorgis, A.; Geleta, M.; Mateos, H.; Lewandowski, P.A. Sterols as a biomarker in tracing niger and sesame seeds oils adulterated with palm oil. Heliyon 2021, 7, e06797. [Google Scholar] [CrossRef]
- Younas, R.; Sahar, A.; Aadil, R.M.; Faisal, M.N. Phytochemical and antioxidant screening of phytosterols extracted from Pisum sativum L. and Cicer arietinum L. through different extraction methods. Biomass Convers. Biorefin. 2024, 15, 8031–8039. [Google Scholar] [CrossRef]
- Vinatoru, M.; Mason, T.J.; Calinescu, I. Ultrasonically assisted extraction (UAE) and microwave assisted extraction (MAE) of functional compounds from plant materials. TrAC—Trends Anal. Chem 2017, 97, 159–178. [Google Scholar] [CrossRef]
- Shen, L.; Pang, S.; Zhong, M.; Sun, Y.; Qayum, A.; Liu, Y.; Rashid, A.; Xu, B.; Liang, Q.; Ma, H.; et al. A comprehensive review of ultrasonic assisted extraction (UAE) for bioactive components: Principles, advantages, equipment, and combined technologies. Ultrason. Sonochem. 2023, 101, 106646. [Google Scholar] [CrossRef] [PubMed]
- Walayat, N.; Yurdunuseven-Yıldız, A.; Kumar, M.; Goksen, G.; Öztekin, S.; Lorenzo, J.M. Oxidative stability, quality, and bioactive compounds of oils obtained by ultrasound and microwave-assisted oil extraction. Crit. Rev. Food. Sci. Nutr. 2024, 64, 9974–9991. [Google Scholar] [CrossRef]
- Barp, L.; Višnjevec, A.M.; Moret, S. Pressurized Liquid Extraction: A Powerful Tool to Implement Extraction and Purification of Food Contaminants. Foods 2023, 12, 2017. [Google Scholar] [CrossRef]
- Hennion, M.C. Solid-phase extraction: Method development, sorbents, and coupling with liquid chromatography. J. Chromatogr. A 1999, 856, 3–54. [Google Scholar] [CrossRef] [PubMed]
- Abubakar, A.R.; Haque, M. Preparation of Medicinal Plants: Basic Extraction and Fractionation Procedures for Experimental Purposes. J. Pharm. Bioallied Sci. 2020, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tavares, G.R.; Massa, T.B.; Gonçalves, J.E.; da Silva, C.; dos Santos, W.D. Assessment of ultrasound-assisted extraction of crambe seed oil for biodiesel synthesis by in situ interesterification. Renew. Energy 2017, 111, 659–665. [Google Scholar] [CrossRef]
- Hien, T.T.; Minh, N.T. Enhancing the extraction of pumpkin seed (Cucurbita pepo L) for increasing oil yield and its phytosterol content. Food Sci. Appl. Biotechnol. 2021, 4, 6–13. [Google Scholar] [CrossRef]
- Xinyue, L. Advances in the source, preparation, and application of phytosterols. Theor. Nat. Sci. 2024, 32, 221–230. [Google Scholar] [CrossRef]
- Akhtar, G.; Masoodi, F.; Muzaffar, S. Effect of ultrasonic and microwave assisted extraction on the oxidative stability, phytochemical and antioxidative properties of Himalayan walnut oil. J. Food Sci. Technol. 2025, 1–10. [Google Scholar] [CrossRef]
- Delazar, A.; Nahar, L.; Hamedeyazdan, S.; Sarker, S.D. Microwave-assisted extraction in natural products isolation. Methods Mol. Biol. 2012, 864, 89–115. [Google Scholar]
- Kaufmann, B.; Christen, P. Recent extraction techniques for natural products: Microwave-assisted extraction and pressurised solvent extraction. Phytochem. Anal. 2002, 13, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Souza, D.E.S.; Melo, J.J.C.; Santos, F.F.D.; Vasconcelos, A.; Jesus, A.D.S.; Freitas, L.D.S.; Souza, R.L.; Soares, C.M.F. Microwave-Assisted vs. Conventional Extraction of Moringa oleifera Seed Oil: Process Optimization and Efficiency Comparison. Foods 2024, 13, 3141. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.-H.; Yuan, Z.-Q.; Li, G.-K. Preparation of phytosterols and phytol from edible marine algae by microwave-assisted extraction and high-speed counter-current chromatography. Sep. Purif. Technol. 2013, 104, 284–289. [Google Scholar] [CrossRef]
- López-Salazar, H.; Camacho-Díaz, B.H.; Ocampo, M.A.; Campos-Mendiola, R.; Martínez-Velarde, R.; López-Bonilla, A.; Jiménez-Aparicio, A.R. Microwave-assisted extraction of β-Sitosterol: A by-product from Agave angustifolia Haw bagasse. BioResources 2024, 19, 568. [Google Scholar] [CrossRef]
- Loganathan, V.; Vijayan, L.; Balakrishnaraja, R.; Abdullah, S. Optimization of microwave-assisted extraction of Tamarindus indica seed oil: An in silico approach to development of potential hypolipidemic compound for reducing LDL cholesterol. Meas. Food 2024, 13, 100125. [Google Scholar] [CrossRef]
- Perez-Vazquez, A.; Carpena, M.; Barciela, P.; Cassani, L.; Simal-Gandara, J.; Prieto, M.A. Pressurized Liquid Extraction for the Recovery of Bioactive Compounds from Seaweeds for Food Industry Application: A Review. Antioxidants 2023, 12, 612. [Google Scholar] [CrossRef]
- Delgado-Zamarreno, M.; Bustamante-Rangel, M.; Martinez-Pelarda, D.; Carabias-Martinez, R. Analysis of β-sitosterol in seeds and nuts using pressurized liquid extraction and liquid chromatography. Anal. Sci. 2009, 25, 765–768. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Yu, Z.; Wang, C.; Li, S. Determination of sterols in rapeseed by pressurized liquid extraction with gas chromatography–tandem mass spectrometry. J. Food Compos. Anal. 2023, 123, 105502. [Google Scholar] [CrossRef]
- Echenique, J.V.F.; Alvarez-Rivera, G.; Luna, V.M.A.; da Cruz Antonio, A.F.V.; Mazalli, M.R.; Ibañez, E.; Cifuentes, A.; de Oliveira, A.L. Pressurized liquid extraction with ethanol in an intermittent process for rice bran oil: Evaluation of process variables on the content of β-sitosterol and phenolic compounds, antioxidant capacity, acetylcholinesterase inhibitory activity, and oil quality. LWT-Food Sci. Technol. 2024, 207, 116650. [Google Scholar] [CrossRef]
- Buszewski, B.; Szultka, M. Past, present, and future of solid phase extraction: A review. Crit. Rev. Anal. Chem. 2012, 42, 198–213. [Google Scholar] [CrossRef]
- Nawaz, M.S.; Ferdousi, F.K.; Rahman, M.A.; Alam, A.S. Reversed Phase SPE and GC-MS Study of Polycyclic Aromatic Hydrocarbons in Water Samples from the River Buriganga, Bangladesh. Int. Sch. Res. Not. 2014, 2014, 234092. [Google Scholar] [CrossRef] [PubMed]
- Walker, V.; Mills, G.A. Solid-phase extraction in clinical biochemistry. Ann. Clin. Biochem. 2002, 39, 464–477. [Google Scholar] [CrossRef]
- Dugheri, S.; Marrubini, G.; Mucci, N.; Cappelli, G.; Bonari, A.; Pompilio, I.; Trevisani, L.; Arcangeli, G. A review of micro-solid-phase extraction techniques and devices applied in sample pretreatment coupled with chromatographic analysis. Acta Chromatogr. 2021, 33, 99–111. [Google Scholar] [CrossRef]
- Wang, H.; Liu, H.; Guo, B.; Lan, D.; Pang, X.; Yan, H.; Han, D.; Guo, H.; Bai, L. A novel poly (NMA-co-DEA-co-EDMA) monolithic column as a sorbent for online solid-phase extraction and its application in the determination of β-sitosterol in plant oil samples. Food Chem. 2019, 278, 594–600. [Google Scholar] [CrossRef]
- Wu, L.; Zhao, J.; Wu, L.; Zhang, Y.; Li, J. Simultaneous determination of squalene, tocopherols and phytosterols in edible vegetable oil by SPE combined with saponification and GC-MS. LWT-Food Sci. Technol. 2022, 169, 114026. [Google Scholar] [CrossRef]
- Zacharis, C.K.; Tzanavaras, P.D. Solid-Phase Microextraction. Molecules 2020, 25, 379. [Google Scholar] [CrossRef]
- Agatonovic-Kustrin, S.; Gegechkori, V.; Kobakhidze, T.; Morton, D. Solid-Phase Microextraction Techniques and Application in Food and Horticultural Crops. Molecules 2023, 28, 6880. [Google Scholar] [CrossRef]
- Kardani, F.; Mirzajani, R.; Ramezani, Z. Direct cholesterol and β-sitosterol analysis in food samples using monolithic molecularly-imprinted solid-phase microextraction fibers coupled with high performance liquid chromatography. J. Iran. Chem. Soc. 2018, 15, 2877–2888. [Google Scholar] [CrossRef]
- Domeño, C.; Ruiz, B.; Nerín, C. Determination of sterols in biological samples by SPME with on-fiber derivatization and GC/FID. Anal. Bioanal. Chem. 2005, 381, 1576–1583. [Google Scholar] [CrossRef]
- Shi, N.; Bu, X.; Zhang, M.; Wang, B.; Xu, X.; Shi, X.; Hussain, D.; Xu, X.; Chen, D. Current Sample Preparation Methodologies for Determination of Catecholamines and Their Metabolites. Molecules 2022, 27, 2702. [Google Scholar] [CrossRef] [PubMed]
- Ebadnezhad, H.; Afshar Mogaddam, M.R.; Mohebbi, A.; Farajzadeh, M.A.; Nemati, M.; Torbati, M. Combination of temperature-assisted ternary phase homogenous liquid-liquid extraction with deep eutectic solvent-based dispersive liquid-liquid microextraction for the extraction of phytosterols from cow milk and cream samples. J. Sep. Sci. 2021, 44, 1482–1489. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, L.; Tamiji, Z.; Khoshayand, M.R. Applications and opportunities of experimental design for the dispersive liquid-liquid microextraction method—A review. Talanta 2018, 190, 335–356. [Google Scholar] [CrossRef] [PubMed]
- Mansour, F.R.; Khairy, M.A. Pharmaceutical and biomedical applications of dispersive liquid-liquid microextraction. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2017, 1061–1062, 382–391. [Google Scholar] [CrossRef]
- Notardonato, I.; Avino, P. Dispersive Liquid–Liquid Micro Extraction: An Analytical Technique Undergoing Continuous Evolution and Development—A Review of the Last 5 Years. Separations 2024, 11, 203. [Google Scholar] [CrossRef]
- Homaie, O.; Afshar Mogaddam, M.R.; Tamizi, E.; Nemati, M. Comparison of organic and deep eutectic solvents based dispersive liquid-liquid microextraction for the analysis of phytosterols in cow milk combined with high-performance liquid chromatography-ultraviolet detector. J. Sep. Sci. 2021, 44, 4167–4180. [Google Scholar] [CrossRef]
- Sun, J.; Zhao, X.E.; Dang, J.; Sun, X.; Zheng, L.; You, J.; Wang, X. Rapid and sensitive determination of phytosterols in functional foods and medicinal herbs by using UHPLC-MS/MS with microwave-assisted derivatization combined with dual ultrasound-assisted dispersive liquid–liquid microextraction. J. Sep. Sci. 2017, 40, 725–732. [Google Scholar] [CrossRef]
- Slavin, M.; Yu, L.L. A single extraction and HPLC procedure for simultaneous analysis of phytosterols, tocopherols and lutein in soybeans. Food Chem. 2012, 135, 2789–2795. [Google Scholar] [CrossRef]
- Yuan, C.; Ju, Y.; Jin, R.; Ren, L.; Liu, X. Simultaneous HPLC–DAD Analysis of Tocopherols, Phytosterols, and Squalene in Vegetable Oil Deodorizer Distillates. Chromatographia 2015, 78, 273–278. [Google Scholar] [CrossRef]
- Novak, A.; Gutiérrez-Zamora, M.; Domenech, L.; Suñé-Negre, J.M.; Miñarro, M.; García-Montoya, E.; Llop, J.M.; Ticó, J.R.; Pérez-Lozano, P. Development and validation of a simple high-performance liquid chromatography analytical method for simultaneous determination of phytosterols, cholesterol and squalene in parenteral lipid emulsions. Biomed. Chromatogr. 2018, 32, e4084. [Google Scholar] [CrossRef]
- Aljohar, H.I.; Al-Hossaini, A.M.; Alzammay, S.A.; Alhabardi, S.; Maher, H.M.; Ahmed, A.R. Development and comprehensive greenness assessment of HPLC method for quality control of β-sitosterol in pharmaceutical ointments with trio-color coded evaluation. BMC Chem. 2025, 19, 14. [Google Scholar] [CrossRef] [PubMed]
- Gachumi, G.; Demelenne, A.; Poudel, A.; Dallal Bashi, Z.; El-Aneed, A. Novel Fast Chromatography-Tandem Mass Spectrometric Quantitative Approach for the Determination of Plant-Extracted Phytosterols and Tocopherols. Molecules 2021, 26, 1402. [Google Scholar] [CrossRef]
- Nabeshima, K.; Mizutani, A.; Misawa, E.; Tanaka, M.; Yamauchi, K.; Abe, F. Validation of an LC-MS/MS method for the quantitation of phytosterols derived from Aloe vera gel. MethodsX 2022, 9, 101642. [Google Scholar] [CrossRef] [PubMed]
- Mo, S.; Dong, L.; Hurst, W.J.; van Breemen, R.B. Quantitative analysis of phytosterols in edible oils using APCI liquid chromatography-tandem mass spectrometry. Lipids 2013, 48, 949–956. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Llatas, G.; Alegría, A.; Barberá, R.; Cilla, A. Current methodologies for phytosterol analysis in foods. Microchem. J. 2021, 168, 106377. [Google Scholar] [CrossRef]
- Saraiva, D.; Semedo, R.; Castilho Mda, C.; Silva, J.M.; Ramos, F. Selection of the derivatization reagent--the case of human blood cholesterol, its precursors and phytosterols GC-MS analyses. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2011, 879, 3806–3811. [Google Scholar] [CrossRef]
- García-Llatas, G.; Vidal, C.; Cilla, A.; Barberá, R.; Lagarda, M.J. Simultaneous quantification of serum phytosterols and cholesterol precursors using a simple gas chromatographic method. Eur. J. Lipid Sci. Technol. 2012, 114, 520–526. [Google Scholar] [CrossRef]
- Nestola, M. Multidimensional High-Performance Liquid Chromatography–Gas Chromatography (HPLC-GC) Hyphenation Techniques for Food Analysis in Routine Environments. Ph.D. Thesis, Universität Duisburg-Essen, Duisburg, Germany, 2016. [Google Scholar]
- dos Santos, M.; Roehrs, M.; de Pereira, C.; Freitag, R.; de Bairros, A. Analysis of phytosterols in plants and derived products by gas chromatography—A short critical review. Austin Chromatogr 2014, 1, 4. [Google Scholar]
- Chen, Y.Z.; Kao, S.Y.; Jian, H.C.; Yu, Y.M.; Li, J.Y.; Wang, W.H.; Tsai, C.W. Determination of cholesterol and four phytosterols in foods without derivatization by gas chromatography-tandem mass spectrometry. J. Food Drug Anal. 2015, 23, 636–644. [Google Scholar] [CrossRef]
- Uchikata, T.; Matsubara, A.; Nishiumi, S.; Yoshida, M.; Fukusaki, E.; Bamba, T. Development of oxidized phosphatidylcholine isomer profiling method using supercritical fluid chromatography/tandem mass spectrometry. J. Chromatogr. A 2012, 1250, 205–211. [Google Scholar] [CrossRef]
- Parr, M.K.; Langley, G.J. Supercritical fluid chromatography. Drug Test. Anal. 2024, 16, 678–679. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Tang, G.; Zhang, T.; Fillet, M.; Crommen, J.; Jiang, Z. Supercritical fluid chromatography in traditional Chinese medicine analysis. J. Pharm. Biomed. Anal. 2018, 147, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Jayantha, J.; Rad, F.M.; Vidanarachchi, J.; Bergquist, J.; Ubhayasekera, S.K.A. A fast ultra performance supercritical fluid chromatography-tandem mass spectrometric method for profiling of targeted phytosterols. J. Chromatogr. B 2023, 1225, 123737. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.J.; Gao, Y.; Xu, X.; Zhao, M.L.; Xi, B.N.; Shu, Y.; Li, C.; Shen, Y. In Situ Rapid Analysis of Squalene, Tocopherols, and Sterols in Walnut Oils Based on Supercritical Fluid Chromatography-Quadrupole Time-of-Flight Mass Spectrometry. J. Agric. Food Chem. 2023, 71, 16371–16380. [Google Scholar] [CrossRef]
- Vrbková, B.; Roblová, V.; Yeung, E.S.; Preisler, J. Determination of sterols using liquid chromatography with off-line surface-assisted laser desorption/ionization mass spectrometry. J. Chromatogr. A 2014, 1358, 102–109. [Google Scholar] [CrossRef]
- Nie, R.; Zhang, Y.; Zhang, H.; Jin, Q.; Wu, G.; Wang, X. Effect of different processing methods on physicochemical properties, chemical compositions and in vitro antioxidant activities of Paeonia lactiflora Pall seed oils. Food Chem. 2020, 332, 127408. [Google Scholar] [CrossRef]
- Orozco-Solano, M.; Ruiz-Jiménez, J.; Luque de Castro, M.D. Ultrasound-assisted extraction and derivatization of sterols and fatty alcohols from olive leaves and drupes prior to determination by gas chromatography-tandem mass spectrometry. J. Chromatogr. A 2010, 1217, 1227–1235. [Google Scholar] [CrossRef]
- Dai, F.J.; Hsu, W.H.; Huang, J.J.; Wu, S.C. Effect of pigeon pea (Cajanus cajan L.) on high-fat diet-induced hypercholesterolemia in hamsters. Food Chem. Toxicol. 2013, 53, 384–391. [Google Scholar] [CrossRef]
- Lv, W.J.; Huang, J.Y.; Lin, J.; Ma, Y.M.; He, S.Q.; Zhang, Y.W.; Wang, T.Z.; Cheng, K.; Xiong, Y.; Sun, F.G.; et al. Phytosterols Alleviate Hyperlipidemia by Regulating Gut Microbiota and Cholesterol Metabolism in Mice. Oxid. Med. Cell. Longev. 2023, 2023, 6409385. [Google Scholar] [CrossRef]
- Demonty, I.; Ras, R.T.; van der Knaap, H.C.M.; Duchateau, G.S.M.J.E.; Meijer, L.; Zock, P.L.; Geleijnse, J.M.; Trautwein, E.A. Continuous Dose-Response Relationship of the LDL-Cholesterol–Lowering Effect of Phytosterol Intake12. J. Nutr. 2009, 139, 271–284. [Google Scholar] [CrossRef]
- Richelle, M.; Enslen, M.; Hager, C.; Groux, M.; Tavazzi, I.; Godin, J.P.; Berger, A.; Métairon, S.; Quaile, S.; Piguet-Welsch, C.; et al. Both free and esterified plant sterols reduce cholesterol absorption and the bioavailability of beta-carotene and alpha-tocopherol in normocholesterolemic humans. Am. J. Clin. Nutr. 2004, 80, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Vivancos, M.; Moreno, J.J. Effect of resveratrol, tyrosol and beta-sitosterol on oxidised low-density lipoprotein-stimulated oxidative stress, arachidonic acid release and prostaglandin E2 synthesis by RAW 264.7 macrophages. Br. J. Nutr. 2008, 99, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Zhang, F.; Shen, M.; Jia, S.; Xie, J. Phytosterols suppress phagocytosis and inhibit inflammatory mediators via ERK pathway on LPS-triggered inflammatory responses in RAW264. 7 macrophages and the correlation with their structure. Foods 2019, 8, 582. [Google Scholar] [CrossRef] [PubMed]
- Aldini, R.; Micucci, M.; Cevenini, M.; Fato, R.; Bergamini, C.; Nanni, C.; Cont, M.; Camborata, C.; Spinozzi, S.; Montagnani, M. Antiinflammatory effect of phytosterols in experimental murine colitis model: Prevention, induction, remission study. PLoS ONE 2014, 9, e108112. [Google Scholar] [CrossRef]
- Pradhan, N.; Parbin, S.; Kausar, C.; Kar, S.; Mawatwal, S.; Das, L.; Deb, M.; Sengupta, D.; Dhiman, R.; Patra, S.K. Paederia foetida induces anticancer activity by modulating chromatin modification enzymes and altering pro-inflammatory cytokine gene expression in human prostate cancer cells. Food Chem. Toxicol. 2019, 130, 161–173. [Google Scholar] [CrossRef]
- Ali, H.; Dixit, S.; Ali, D.; Alqahtani, S.M.; Alkahtani, S.; Alarifi, S. Isolation and evaluation of anticancer efficacy of stigmasterol in a mouse model of DMBA-induced skin carcinoma. Drug Des. Devel. Ther. 2015, 9, 2793–2800. [Google Scholar] [CrossRef]
- Huang, J.; Xu, M.; Fang, Y.-J.; Lu, M.-S.; Pan, Z.-Z.; Huang, W.-Q.; Chen, Y.-M.; Zhang, C.-X. Association between phytosterol intake and colorectal cancer risk: A case–control study. Br. J. Nutr. 2017, 117, 839–850. [Google Scholar] [CrossRef]
- Ward, M.G.; Li, G.; Barbosa-Lorenzi, V.C.; Hao, M. Stigmasterol prevents glucolipotoxicity induced defects in glucose-stimulated insulin secretion. Sci. Rep. 2017, 7, 9536. [Google Scholar] [CrossRef]
- Ponnulakshmi, R.; Shyamaladevi, B.; Vijayalakshmi, P.; Selvaraj, J. In silico and in vivo analysis to identify the antidiabetic activity of beta sitosterol in adipose tissue of high fat diet and sucrose induced type-2 diabetic experimental rats. Toxicol. Mech. Methods 2019, 29, 276–290. [Google Scholar] [CrossRef]
- Gao, F.; Wang, G.; Wang, L.; Guo, N. Phytosterol nutritional supplement improves pregnancy and neonatal complications of gestational diabetes mellitus in a double-blind and placebo-controlled clinical study. Food Funct. 2017, 8, 424–428. [Google Scholar] [CrossRef]
- Oh, J.H.; Choi, J.S.; Nam, T.J. Fucosterol from an Edible Brown Alga Ecklonia stolonifera Prevents Soluble Amyloid Beta-Induced Cognitive Dysfunction in Aging Rats. Mar. Drugs 2018, 16, 368. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Aggarwal, P.; Khan, F.; Khodve, G.; Padhy, D.S.; Yadav, P.; Banerjee, S. β-sitosterol Protects against Aluminium Chloride-mediated Neurotoxicity. Curr. Alzheimer Res. 2023, 20, 29–37. [Google Scholar] [PubMed]
- Ference, B.A.; Ginsberg, H.N.; Graham, I.; Ray, K.K.; Packard, C.J.; Bruckert, E.; Hegele, R.A.; Krauss, R.M.; Raal, F.J.; Schunkert, H.; et al. Low-density lipoproteins cause atherosclerotic cardiovascular disease. 1. Evidence from genetic, epidemiologic, and clinical studies. A consensus statement from the European Atherosclerosis Society Consensus Panel. Eur. Heart J. 2017, 38, 2459–2472. [Google Scholar] [CrossRef] [PubMed]
- Ras, R.T.; Geleijnse, J.M.; Trautwein, E.A. LDL-cholesterol-lowering effect of plant sterols and stanols across different dose ranges: A meta-analysis of randomised controlled studies. Br. J. Nutr. 2014, 112, 214–219. [Google Scholar] [CrossRef]
- Abumweis, S.S.; Barake, R.; Jones, P.J. Plant sterols/stanols as cholesterol lowering agents: A meta-analysis of randomized controlled trials. Food Nutr. Res. 2008, 52, 1811. [Google Scholar] [CrossRef]
- Rocha, M.; Banuls, C.; Bellod, L.; Jover, A.M.; Victor, V.; Hernandez-Mijares, A. A review on the role of phytosterols: New insights into cardiovascular risk. Curr. Pharm. Des. 2011, 17, 4061–4075. [Google Scholar] [CrossRef]
- Malinowski, J.M.; Gehret, M.M. Phytosterols for dyslipidemia. Am. J. Health Syst. Pharm. 2010, 67, 1165–1173. [Google Scholar] [CrossRef]
- Poznyak, A.V.; Nikiforov, N.G.; Markin, A.M.; Kashirskikh, D.A.; Myasoedova, V.A.; Gerasimova, E.V.; Orekhov, A.N. Overview of OxLDL and its impact on cardiovascular health: Focus on atherosclerosis. Front. Pharmacol. 2021, 11, 613780. [Google Scholar] [CrossRef]
- Jiang, H.; Zhou, Y.; Nabavi, S.M.; Sahebkar, A.; Little, P.J.; Xu, S.; Weng, J.; Ge, J. Mechanisms of oxidized LDL-mediated endothelial dysfunction and its consequences for the development of atherosclerosis. Front. Cardiovasc. Med. 2022, 9, 925923. [Google Scholar] [CrossRef]
- Hulthe, J.; Fagerberg, B. Circulating oxidized LDL is associated with subclinical atherosclerosis development and inflammatory cytokines (AIR Study). Arterioscler. Thromb. Vasc. Biol. 2002, 22, 1162–1167. [Google Scholar] [CrossRef]
- Pardridge, W.M. Blood-brain barrier biology and methodology. J. Neurovirol. 1999, 5, 556–569. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, H.; Wang, Y.; Griffiths, W.J.; Levey, A.I.; Pikuleva, I.; Liang, S.H.; Haider, A. Brain cholesterol and Alzheimer’s disease: Challenges and opportunities in probe and drug development. Brain 2024, 147, 1622–1635. [Google Scholar] [CrossRef]
- Refolo, L.M.; Malester, B.; LaFrancois, J.; Bryant-Thomas, T.; Wang, R.; Tint, G.S.; Sambamurti, K.; Duff, K.; Pappolla, M.A. Hypercholesterolemia accelerates the Alzheimer’s amyloid pathology in a transgenic mouse model. Neurobiol. Dis. 2000, 7, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Wang, L.; Shao, P.; Sun, P.; Yang, C.S. A review on chemical and physical modifications of phytosterols and their influence on bioavailability and safety. Crit. Rev. Food. Sci. Nutr. 2022, 62, 5638–5657. [Google Scholar] [CrossRef]
- Quilliot, D.; Boman, F.; Creton, C.; Pelletier, X.; Floquet, J.; Debry, G. Phytosterols have an unfavourable effect on bacterial activity and no evident protective effect on colon carcinogenesis. Eur. J. Cancer Prev. 2001, 10, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Oyeyinka, S.A.; Xu, W.; Ma, Y.; Zhou, S. In vitro bioaccessibility and physicochemical properties of phytosterol linoleic ester synthesized from soybean sterol and linoleic acid. LWT-Food Sci. Technol. 2018, 92, 265–271. [Google Scholar] [CrossRef]
- Hukkinen, M.; Mutanen, A.; Nissinen, M.; Merras-Salmio, L.; Gylling, H.; Pakarinen, M.P. Parenteral plant sterols accumulate in the liver reflecting their increased serum levels and portal inflammation in children with intestinal failure. J. Parenter. Enteral. Nutr. 2017, 41, 1014–1022. [Google Scholar] [CrossRef]
| Source of Phytosterols | Models | Experimental Design | Major Findings | Conclusions | References |
|---|---|---|---|---|---|
| Cajanus cajan L. (pigeon pea) oil rich in phytosterols | High-fat-diet (HFD)-induced dyslipidemia in Syrian hamsters | Hamsters were fed an HFD containing 20%, 40%, and 80% Cajanus cajan L. oil for 8 weeks. | C. cajan L. oil improved lipid profiles and glucose tolerance in HFD-induced hyperlipidemic hamsters by upregulating cholesterol 7 alpha-hydroxylase (CYP7A1) and carnitine palmitoyltransferase 1 (CPT-1) expression, enhancing bile acid synthesis and β-oxidation, and reducing hepatic lipid accumulation. | C. cajan L. oil-derived phytosterols can modulate lipid metabolism and serve as potential therapeutic agents against metabolic disorders like hyperlipidemia and type 2 diabetes. | [89] |
| Phytosterol, containing 40% β-sitosterol, 20% campesterol, and over 14% stigmasterol | HFD-induced hyperlipidemia in C57BL/6J mice | Phytosterol (100 mg/kg) was administered from week 4 of a 12-week HFD. | Phytosterol improved lipid profiles, enhanced hepatic antioxidant activity, and modulated key genes, such as CYP7A1, sterol 12-alpha-hydroxylase (CYP8B1), sterol 27-hydroxylase (CYP27A1), and oxysterol 7-alpha-hydroxylase (CYP7B1). Phytosterols also regulated cholesterol metabolism pathways and modulated the composition of the gut microbiota. | Phytosterols improved antioxidant function and reduced hyperlipidemia by modulating cholesterol metabolism and shifting bile acid synthesis to the alternative pathway. | [90] |
| Dietary phytosterols | Clinical trials | 6805 participants consumed 2.15 g of plant sterols per day for 21 to 182 days. | Phytosterol supplementation dose-dependently reduced LDL-C by ~9% at 2 g/day, with effects plateauing around 3 g/day. No significant differences were found between sterols and stanols or between esterified and free forms. | This meta-analysis supports phytosterols as an effective LDL-C-lowering strategy, with further study needed on dosing and form. | [91] |
| Plant-based sterol | Clinical trials | 26 men consumed low-fat milk with or without 2.2 g of plant sterols (free sterols or sterol esters) for 1 week. | Both sterol-enriched milks reduced cholesterol absorption by ~60%. The bioavailability of β-carotene and α-tocopherol decreased by approximately 50% and 20%, respectively, with smaller reductions seen in the sterol-free form. | Plant sterols reduce the absorption of cholesterol, β-carotene, and α-tocopherol, with sterol esters having a stronger effect. | [92] |
| β-sitosterol | Oxidized-LDL(oxLDL)-induced RAW 264.7 cells | Cells were pretreated with 10 μM β-sitosterol for 3 h, then exposed to 40 μg/mL oxLDL for 30 min. | β-Sitosterol inhibited H2O2, arachidonic acid (AA), and prostaglandin E2 (PGE2) generation in oxLDL-treated RAW 264.7 cells. The presence of β-sitosterol enhanced the inhibitory effects of tyrosol and resveratrol on oxLDL-induced H2O2, AA, and PGE2 production. | The synergistic effect of β-sitosterol and polyphenol in olive oil may prevent the release of reactive oxygen species (ROS) and inflammatory mediators in oxLDL-treated macrophages. | [93] |
| Ergosterol, stigmasterol, β-sitosterol, campesterol, and ergosterol acetate | Lipopolysaccharide (LPS, 1 μg/mL)-stimulated RAW 264.7 cells | LPS-stimulated RAW 264.7 cells were treated with 25, 50, 100, and 200 μM of phytosterols. | Phytosterols reduced tumor necrosis factor-α (TNF-α), nitric oxide (NO), and related protein expression in LPS-stimulated macrophages, with effects linked to extracellular signal-regulated kinase (ERK) pathway regulation and structural features like C-22 double bonds and C-24 side chains. | Phytosterols have anti-inflammatory potential, warranting further research on their structure–activity relationships. | [94] |
| NRF21 diet | Dextran sodium sulfate (DSS)-induced colitis in Balb/c mice | Mice were fed a phytosterol-enriched diet (400 mg/kg/day) or control diet for 14 days, with 5% DSS given until day 10, which was then replaced by water. | Phytosterol supplementation alleviated DSS-induced colitis by reducing inflammation and oxidative stress, promoting mucosal healing, improving gut motility, and preserving muscle contractions. It also restored cholinergic signaling by upregulating muscarinic receptors impaired by inflammation. | Phytosterol pretreatment alleviates DSS-induced colitis through antioxidant, bile acid, and microbiota modulation, showing potential as a functional food for inflammatory bowel disease (IBD) management. | [95] |
| Lupeol and β-sitosterol isolated from Paederia foetida leaves | PC3 and DU145 prostate cancer cell | PC-3 and DU145 prostate cancer cells were treated with β-sitosterol (80 μM) and lupeol (40 μM). | Lupeol and β-sitosterol reactivated cadherin-1 (CDH1) by suppressing DNA (cytosine-5)-methyltransferase 1 (DNMT1) and histone deacetylase (HDAC), inhibited prostate cancer cell growth and migration, induced apoptosis, and reduced TNF-α expression. | Lupeol and β-sitosterol show anticancer effects in prostate cancer through epigenetic, apoptotic, and anti-inflammatory pathways, highlighting P. foetida’s therapeutic potential. | [96] |
| Stigmasterol isolated from Azadirachta indica | DMBA (7,12-dimethylbenz[a]anthracene)-induced skin carcinoma in Swiss albino mice | DMBA (100 μg/100 μL acetone) and 1% croton oil were applied topically, and stigmasterol (400 mg/kg) was given orally three times weekly for 16 weeks. | Stigmasterol exhibited potent anticancer and antioxidant effects by inhibiting DMBA-induced skin papillomas, reducing oxidative stress, and enhancing antioxidant enzyme activity. It also promoted tissue repair and prevented DNA damage. | Stigmasterol shows strong chemopreventive potential against DMBA-induced skin cancer in mice, supporting its use as a natural anticancer agent. | [97] |
| Dietary phytosterols | Clinical trials | The effects of a daily intake of approximately 322 mg of plant sterols were investigated in colorectal cancer patients. | Higher total phytosterol intake was linked to reduced colorectal cancer risk, while stigmasterol was associated with increased risk, especially in women. Subgroup analysis showed site-specific links, with stronger protective effects in younger individuals. | Total phytosterols reduced colorectal cancer risk, while stigmasterol increased it, especially in women, warranting further study. | [98] |
| Stigmasterol | NS-1 cells and human islets exposed to HGP (30 mM glucose, 0.5 mM palmitate) | Stigmasterol (50 μg/mL) was tested for protecting β-cells from glucolipotoxicity by reducing stress and preserving insulin function. | Stigmasterol protects pancreatic β-cells from glucolipotoxicity by reducing ROS and cholesterol, improving insulin secretion, and lowering apoptosis. Stigmasterol also restores glucose-stimulated insulin secretion in NS-1 cells by inhibiting glucose-induced actin polymerization. | Stigmasterol improves β-cell function under diabetic conditions and may help treat type 2 diabetes, pending further in vivo validation. | [99] |
| β-sitosterol | High-fat-diet- and high-sucrose-diet-induced type 2 diabetes in albino rats | β-Sitosterol (5, 10, 20, and 30 mg/kg) was administered from day 30 of a 60-day HFD. | β-Sitosterol improved glucose and lipid metabolism in diabetic rats by enhancing insulin signaling, increasing glucose transporter 4 (GLUT4), and reducing oxidative stress. | β-Sitosterol may help manage type 2 diabetes by improving insulin signaling and reducing oxidative stress. | [100] |
| Phytosterol-rich margarine spread | Clinical trials | A phytosterol-rich margarine spread (2 g phytosterols per 10 g) was consumed twice daily until late pregnancy. | In patients with gestational diabetes mellitus (GDM), daily consumption of the phytosterol-enriched spread improved maternal lipid and glucose metabolism, as evidenced by reductions in triacylglycerol (TAG), total cholesterol (TC), low-density lipoprotein (LDL), insulin levels, and the homeostatic model assessment of insulin resistance (HOMA-IR), alongside increases in high-density lipoprotein (HDL) and the quantitative insulin sensitivity check index (QUICKI). Neonatal complications, including low birth weight and hypoglycemia, were also reduced. | Phytosterol-enriched spread may improve maternal metabolic profiles and reduce neonatal complications in diabetic patients. | [101] |
| Fucosterol isolated from Eisenia stolonifera | Amyloid-β (Aβ)1–42-induced cognitive impairment in aged rats | Aged rats received fucosterol (10 μmol/h) for 4 weeks and Aβ1–42 (25 mg/kg) for 7 days to induce cognitive impairment. | Fucosterol protected against Aβ1–42-induced endoplasmic reticulum stress (ER) and cognitive impairment by restoring brain-derived neurotrophic factor (BDNF) expression and activating the tropomyosin receptor kinase B–extracellular signal-regulated kinase 1/2 (TrkB-ERK1/2) signaling pathway, which is essential for neuronal survival and memory function. | Fucosterol’s potential as a neuroprotective agent for age-related neurodegenerative diseases is suggested. | [102] |
| β-Sitosterol | AlCl3-induced cognitive impairment in C57BL/6 mice | AlCl3 (10 mg/kg) and β-sitosterol (25 mg/kg) were administered for 21 days. | β-Sitosterol improves cognitive function in Alzheimer’s models by crossing the blood–brain barrier (BBB), reducing Aβ and acetylcholinesterase (AChE) levels, enhancing memory, and boosting brain glutathione (GSH), with potential Aβ clearance via β-site amyloid precursor protein-cleaving enzyme 1 (BACE1) inhibition or secretase activation. | β-Sitosterol prevents AlCl3-induced cognitive impairment and shows potential for Alzheimer’s disease (AD) management through its neuroprotective, anticholinesterase, and anti-inflammatory effects, though its mechanisms require further investigation. | [103] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, B.-H.; Truong, V.-L.; Jeong, W.-S. Phytosterols: Extraction Methods, Analytical Techniques, and Biological Activity. Molecules 2025, 30, 2488. https://doi.org/10.3390/molecules30122488
Yoon B-H, Truong V-L, Jeong W-S. Phytosterols: Extraction Methods, Analytical Techniques, and Biological Activity. Molecules. 2025; 30(12):2488. https://doi.org/10.3390/molecules30122488
Chicago/Turabian StyleYoon, Byoung-Hoon, Van-Long Truong, and Woo-Sik Jeong. 2025. "Phytosterols: Extraction Methods, Analytical Techniques, and Biological Activity" Molecules 30, no. 12: 2488. https://doi.org/10.3390/molecules30122488
APA StyleYoon, B.-H., Truong, V.-L., & Jeong, W.-S. (2025). Phytosterols: Extraction Methods, Analytical Techniques, and Biological Activity. Molecules, 30(12), 2488. https://doi.org/10.3390/molecules30122488