
Sea Cucumber and Blueberry Extracts Suppress Inflammation and Reduce Acute Lung Injury through the Regulation of NF-κB/MAPK/JNK Signaling Pathway in Lipopolysaccharide-Treated C57BL/6 Mice

 , , ,
, , ,  ,
, 

Abstract
1. Introduction
2. Results
2.1. Body Weight of Experimental Mice
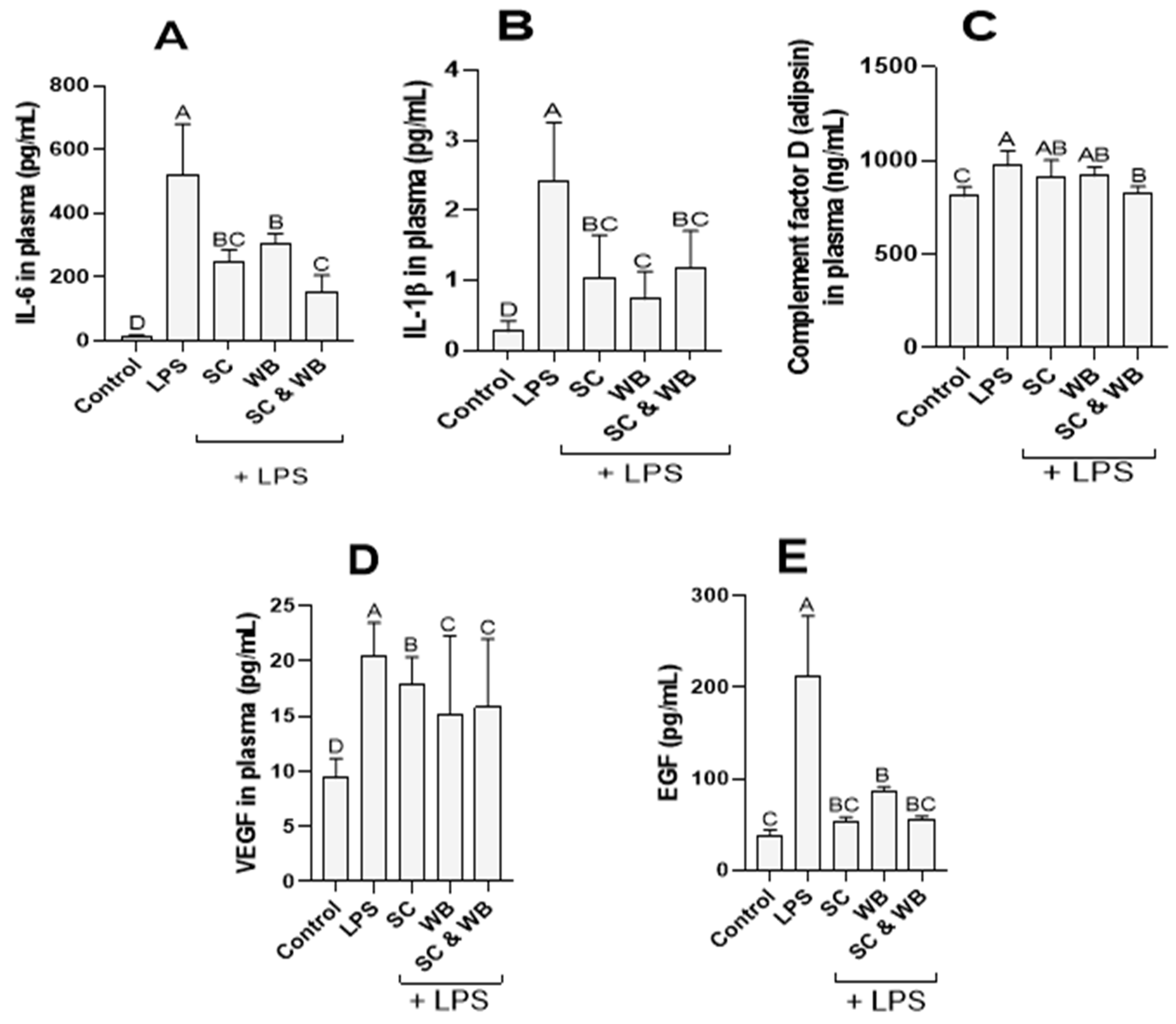
2.2. Effect of Sea Cucumber and Wild Blueberry Extracts on IL-6, IL-1β, Vascular Endothelial Growth Factor (VEGF), and Epidermal Growth Factor (EGF) Levels
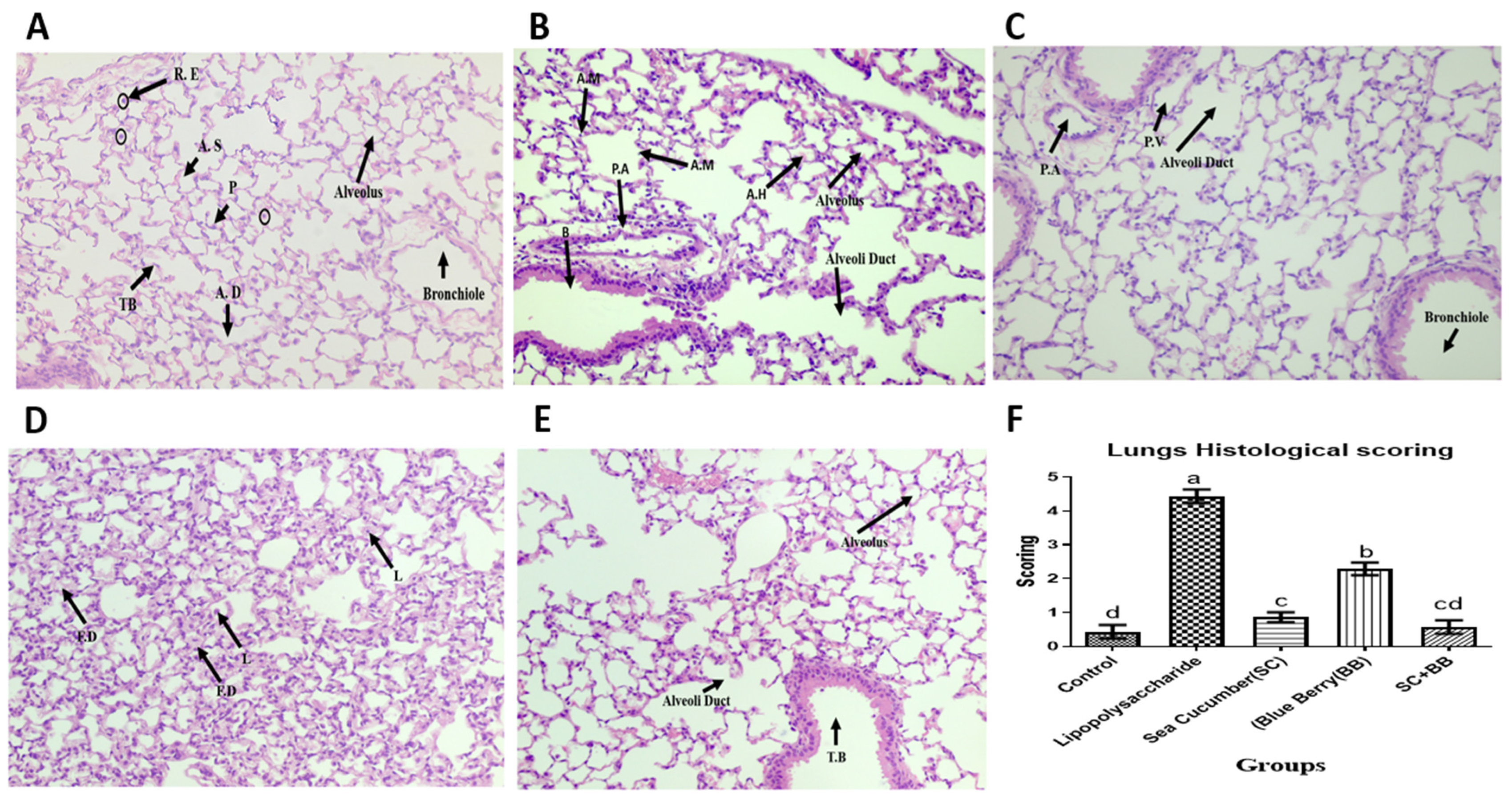
2.3. Sea Cucumber and Wild Blueberry Attenuate Histopathological Signs of Experimental ALI
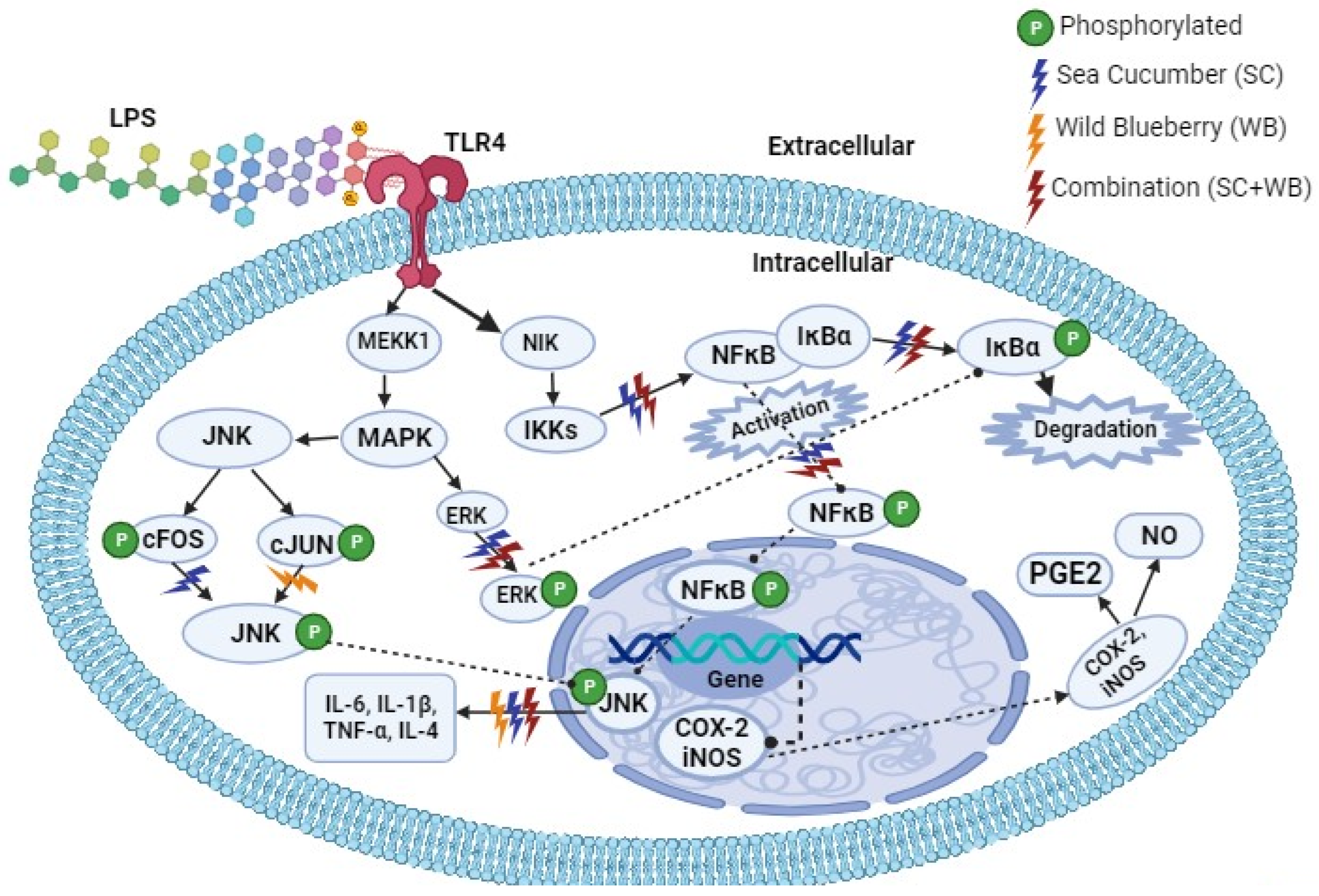
2.4. Supplementation of Sea Cucumber and Wild Blueberry Extracts Attenuated LPS-Induced Inflammation via NF-κB and MAPK Signaling Pathways
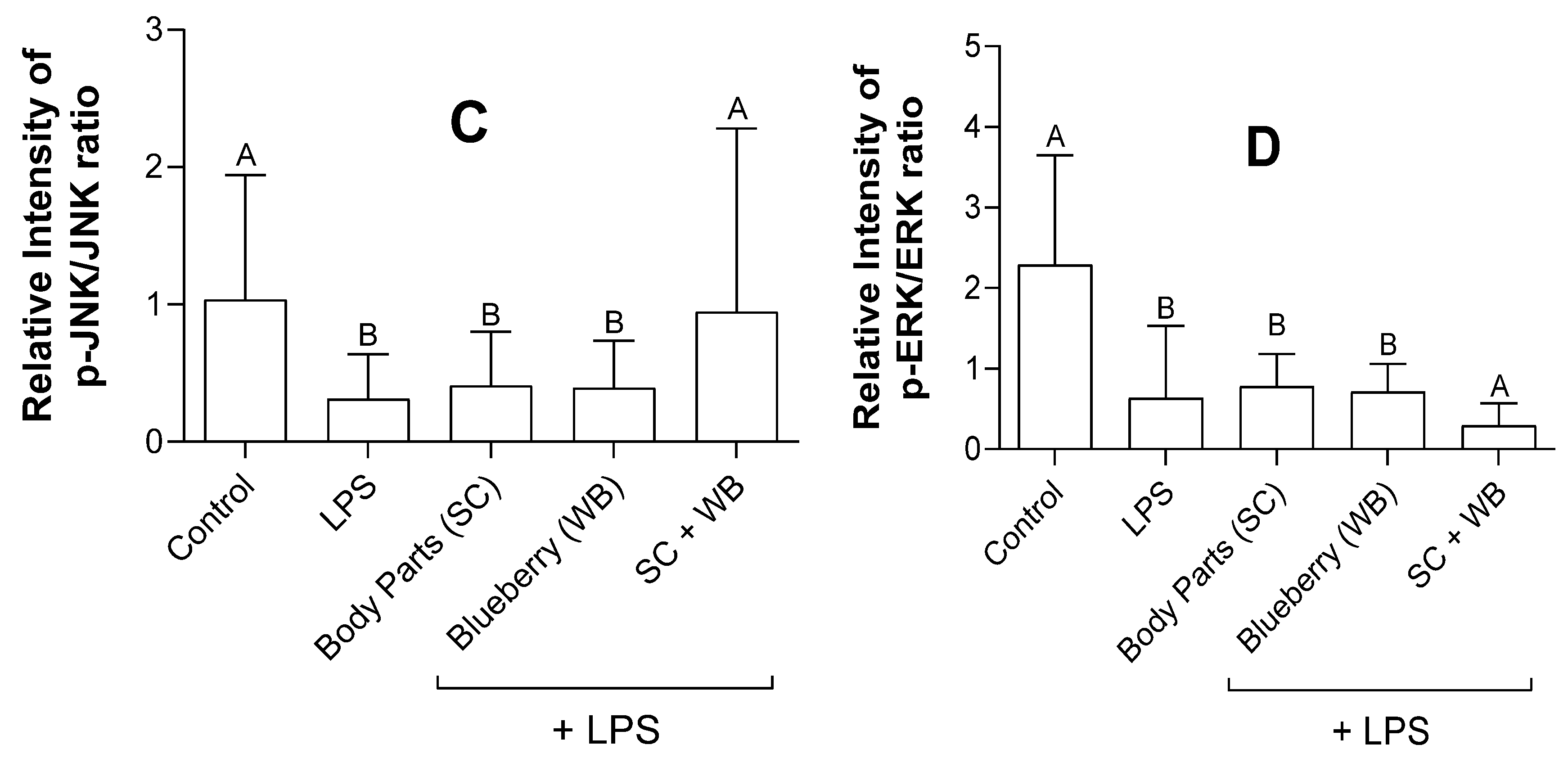
2.5. Sea Cucumber and Wild Blueberry Extracts Reduced NF-κB and MAPK Expression While the Combined Extracts Increased JNK Expression
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Experimental Animals
4.3. Extraction of Sea Cucumber Materials and Purification of Saponins
4.4. Extraction Procedure for Wild Blueberry
4.5. Quantification of Frondoside A Using Ultra-High-Performance Liquid Chromatography-Electrospray Ionization-Mass Spectrometry (UPLC-ESI-MS)
4.6. Quantification of Total Anthocyanins by the pH Differential Method
4.7. Diet Preparation by Palletization of Sea Cucumber and Blueberry Extracts
4.8. Experimental Model of Acute Lung Injury (ALI)
4.9. Luminex® Multiplex and Enzyme-Linked Immunosorbent (ELISA) Assays
4.10. Lung Histological and Histopathological Examination
4.11. Total RNA Isolation and qRT-PCR Analysis
4.12. Western Blotting Techniques for Protein Expression
4.13. Statistical Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnson, E.R.; Matthay, M.A. Acute lung injury: Epidemiology, pathogenesis, and treatment. J. Aerosol Med. Pulm. Drug Deliv. 2010, 23, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Matuschak, G.M.; Lechner, A.J. Acute lung injury and the acute respiratory distress syndrome: Pathophysiology and treatment. Mo. Med. 2010, 107, 252–258. [Google Scholar] [PubMed]
- Rezk, A.R.; Mohamed, M.A.; Elkenawy, M.H.; Bakry, N.; Omran, A. High-frequency oscillatory ventilation versus conventional ventilation in pediatric patients with acute lung injury: Outcomes evaluation. Egypt. Pediatr. Assoc. Gaz. 2022, 70, 36. [Google Scholar] [CrossRef]
- Matthay, M.A.; Zemans, R.L. The acute respiratory distress syndrome: Pathogenesis and treatment. Annu. Rev. Pathol. 2011, 6, 147–163. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Zhang, S.; Yang, Y.; Yao, J.-Q.; Tang, W.-F.; Lyon, C.J.; Hu, T.Y.; Wan, M.-H. Extracellular vesicles in the pathogenesis and treatment of acute lung injury. Mil. Med. Res. 2022, 9, 61. [Google Scholar] [CrossRef] [PubMed]
- Aslan, A.; Aslan, C.; Zolbanin, N.M.; Jafari, R. Acute respiratory distress syndrome in COVID-19: Possible mechanisms and therapeutic management. Pneumonia 2021, 13, 14. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.; Khurram Syed, S.; Fatima, M.; Shaukat, S.; Saadullah, M.; Alqahtani, A.M.; Alqahtani, T.; Bin Emran, T.; Alamri, A.H.; Barkat, M.Q.; et al. Acute Respiratory Distress Syndrome and COVID-19: A Literature Review. J. Inflamm. Res. 2021, 14, 7225–7242. [Google Scholar] [CrossRef]
- Bossardi Ramos, R.; Adam, A.P. Molecular Mechanisms of Vascular Damage During Lung Injury. Adv. Exp. Med. Biol. 2021, 1304, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Semple, J.W.; Rebetz, J.; Kapur, R. Transfusion-associated circulatory overload and transfusion-related acute lung injury. Blood 2019, 133, 1840–1853. [Google Scholar] [CrossRef]
- Vassiliou, A.G.; Kotanidou, A.; Dimopoulou, I.; Orfanos, S.E. Endothelial Damage in Acute Respiratory Distress Syndrome. Int. J. Mol. Sci. 2020, 21, 8793. [Google Scholar] [CrossRef] [PubMed]
- Grommes, J.; Soehnlein, O. Contribution of neutrophils to acute lung injury. Mol. Med. 2011, 17, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Dean, D.A. Gene Therapy for Acute Respiratory Distress Syndrome. Front. Physiol. 2022, 12, 786255. [Google Scholar] [CrossRef] [PubMed]
- Montazersaheb, S.; Hosseiniyan Khatibi, S.M.; Hejazi, M.S.; Tarhriz, V.; Farjami, A.; Ghasemian Sorbeni, F.; Farahzadi, R.; Ghasemnejad, T. COVID-19 infection: An overview on cytokine storm and related interventions. Virol. J. 2022, 19, 92. [Google Scholar] [CrossRef]
- Darif, D.; Hammi, I.; Kihel, A.; El Idrissi Saik, I.; Guessous, F.; Akarid, K. The pro-inflammatory cytokines in COVID-19 pathogenesis: What goes wrong? Microb. Pathog. 2021, 153, 104799. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Abbas, A.; Lehmann, C.; Rupasinghe, H.P.V. Antiviral and Anti-Inflammatory Plant-Derived Bioactive Compounds and Their Potential Use in the Treatment of COVID-19-Related Pathologies. J. Xenobiotics 2022, 12, 289–306. [Google Scholar] [CrossRef] [PubMed]
- Calfee, C.S.; Matthay, M.A. Nonventilatory treatments for acute lung injury and ARDS. Chest 2007, 131, 913–920. [Google Scholar] [CrossRef] [PubMed]
- Meade, M.O.; Cook, D.J.; Guyatt, G.H.; Slutsky, A.S.; Arabi, Y.M.; Cooper, D.J.; Davies, A.R.; Hand, L.E.; Zhou, Q.; Thabane, L.; et al. Ventilation Strategy Using Low Tidal Volumes, Recruitment Maneuvers, and High Positive End-Expiratory Pressure for Acute Lung Injury and Acute Respiratory Distress Syndrome: A Randomized Controlled Trial. JAMA 2008, 299, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Heimberg, D.H.; Illg, Z.; Corser, W.D. Quality Improvement Intervention associated with Improved Lung Protective Ventilation Settings in an Emergency Department. Spartan Med. Res. J. 2022, 7, 29603. [Google Scholar] [CrossRef] [PubMed]
- Bosma, K.J.; Taneja, R.; Lewis, J.F. Pharmacotherapy for prevention and treatment of acute respiratory distress syndrome: Current and experimental approaches. Drugs 2010, 70, 1255–1282. [Google Scholar] [CrossRef] [PubMed]
- Cavaillon, J.-M.; Singer, M.; Skirecki, T. Sepsis therapies: Learning from 30 years of failure of translational research to propose new leads. EMBO Mol. Med. 2020, 12, e10128. [Google Scholar] [CrossRef] [PubMed]
- Keddissi, J.I.; Youness, H.A.; Jones, K.R.; Kinasewitz, G.T. Fluid management in Acute Respiratory Distress Syndrome: A narrative review. Can. J. Respir. Ther. 2019, 55, 1–8. [Google Scholar] [CrossRef]
- Seitz, K.P.; Caldwell, E.S.; Hough, C.L. Fluid management in ARDS: An evaluation of current practice and the association between early diuretic use and hospital mortality. J. Intensive Care 2020, 8, 78. [Google Scholar] [CrossRef] [PubMed]
- He, Y.-Q.; Zhou, C.-C.; Yu, L.-Y.; Wang, L.; Deng, J.-l.; Tao, Y.-L.; Zhang, F.; Chen, W.-S. Natural product derived phytochemicals in managing acute lung injury by multiple mechanisms. Pharmacol. Res. 2021, 163, 105224. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y. Efficacy of statin therapy in patients with acute respiratory distress syndrome/acute lung injury: A systematic review and meta-analysis. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3190–3198. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.-Y.; Zhou, T.-Y.; Zhang, Z.-D.; Liu, H.-Y.; Zheng, Z.-Y.; Xie, H.-Q. Current therapeutic strategies for respiratory diseases using mesenchymal stem cells. MedComm 2021, 2, 351–380. [Google Scholar] [CrossRef] [PubMed]
- Sadeghian Chaleshtori, S.; Mokhber Dezfouli, M.R.; Jabbari Fakhr, M. Mesenchymal stem/stromal cells: The therapeutic effects in animal models of acute pulmonary diseases. Respir. Res. 2020, 21, 110. [Google Scholar] [CrossRef] [PubMed]
- Behnke, J.; Kremer, S.; Shahzad, T.; Chao, C.-M.; Böttcher-Friebertshäuser, E.; Morty, R.E.; Bellusci, S.; Ehrhardt, H. MSC Based Therapies—New Perspectives for the Injured Lung. J. Clin. Med. 2020, 9, 682. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Dave, D.; Shahidi, F. Northern Sea Cucumber (Cucumaria frondosa): A Potential Candidate for Functional Food, Nutraceutical, and Pharmaceutical Sector. Mar. Drugs 2020, 18, 274. [Google Scholar] [CrossRef] [PubMed]
- Fagbohun, O.F.; Joseph, J.S.; Oriyomi, O.V.; Rupasinghe, H.P.V. Saponins of North Atlantic Sea Cucumber: Chemistry, Health Benefits, and Future Prospectives. Mar. Drugs 2023, 21, 262. [Google Scholar] [CrossRef] [PubMed]
- Sivapragasam, N.; Neelakandan, N.; Rupasinghe, H.P.V. Potential health benefits of fermented blueberry: A review of current scientific evidence. Trends Food Sci. Technol. 2023, 132, 103–120. [Google Scholar] [CrossRef]
- Sajwani, F.H. Frondoside A is a potential anticancer agent from sea cucumbers. J. Cancer Res. Ther. 2019, 15, 953–960. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Kundu, N.; Collin, P.D.; Goloubeva, O.; Fulton, A.M. Frondoside A inhibits breast cancer metastasis and antagonizes prostaglandin E receptors EP4 and EP2. Breast Cancer Res. Treat. 2012, 132, 1001–1008. [Google Scholar] [CrossRef] [PubMed]
- Janakiram, N.B.; Mohammed, A.; Rao, C.V. Sea Cucumbers Metabolites as Potent Anti-Cancer Agents. Mar. Drugs 2015, 13, 2909–2923. [Google Scholar] [CrossRef] [PubMed]
- Adrian, T.E.; Collin, P. The Anti-Cancer Effects of Frondoside A. Mar. Drugs 2018, 16, 64. [Google Scholar] [CrossRef] [PubMed]
- Karki, R.; Sharma, B.R.; Tuladhar, S.; Williams, E.P.; Zalduondo, L.; Samir, P.; Zheng, M.; Sundaram, B.; Banoth, B.; Malireddi, R.K.S.; et al. Synergism of TNF-α and IFN-γ Triggers Inflammatory Cell Death, Tissue Damage, and Mortality in SARS-CoV-2 Infection and Cytokine Shock Syndromes. Cell 2021, 184, 149–168.e117. [Google Scholar] [CrossRef]
- Zhang, L.; Zhu, X.-Z.; Badamjav, R.; Zhang, J.-Z.; Kou, J.-P.; Yu, B.-Y.; Li, F. Isoorientin protects lipopolysaccharide-induced acute lung injury in mice via modulating Keap1/Nrf2-HO-1 and NLRP3 inflammasome pathways. Eur. J. Pharmacol. 2022, 917, 174748. [Google Scholar] [CrossRef]
- Yang, S.; Yuan, H.Q.; Hao, Y.M.; Ren, Z.; Qu, S.L.; Liu, L.S.; Wei, D.H.; Tang, Z.H.; Zhang, J.F.; Jiang, Z.S. Macrophage polarization in atherosclerosis. Clin. Chim. Acta 2020, 501, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wu, X.; Han, G.; Shao, B.; Lin, L.; Jiang, S. Altered M1/M2 polarization of alveolar macrophages is involved in the pathological responses of acute silicosis in rats in vivo. Toxicol. Ind. Health 2022, 38, 810–818. [Google Scholar] [CrossRef] [PubMed]
- Mangalmurti, N.; Hunter, C.A. Cytokine Storms: Understanding COVID-19. Immunity 2020, 53, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, C.; Alizadeh-Tabrizi, N.; Hall, S.; Faridi, S.; Euodia, I.; Holbein, B.; Zhou, J.; Chappe, V. Anti-Inflammatory Effects of the Iron Chelator, DIBI, in Experimental Acute Lung Injury. Molecules 2022, 27, 4036. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, J.N.; Lucas, R.; Verin, A.D. The Acute Respiratory Distress Syndrome: Mechanisms and Perspective Therapeutic Approaches. Austin J. Vasc. Med. 2015, 2, 1009. [Google Scholar] [PubMed]
- Torres Acosta, M.A.; Singer, B.D. Pathogenesis of COVID-19-induced ARDS: Implications for an ageing population. Eur. Respir. J. 2020, 56, 2002049. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef] [PubMed]
- Parameswaran, N.; Patial, S. Tumor necrosis factor-α signaling in macrophages. Crit. Rev. Eukaryot. Gene Expr. 2010, 20, 87–103. [Google Scholar] [CrossRef]
- Guo, Y.; Hu, K.; Li, Y.; Lu, C.; Ling, K.; Cai, C.; Wang, W.; Ye, D. Targeting TNF-α for COVID-19: Recent Advanced and Controversies. Front. Public. Health 2022, 10, 833967. [Google Scholar] [CrossRef] [PubMed]
- Mortaz, E.; Tabarsi, P.; Jamaati, H.; Dalil Roofchayee, N.; Dezfuli, N.K.; Hashemian, S.M.; Moniri, A.; Marjani, M.; Malekmohammad, M.; Mansouri, D.; et al. Increased Serum Levels of Soluble TNF-α Receptor Is Associated With ICU Mortality in COVID-19 Patients. Front. Immunol. 2021, 12, 592727. [Google Scholar] [CrossRef] [PubMed]
- Jang, D.I.; Lee, A.H.; Shin, H.Y.; Song, H.R.; Park, J.H.; Kang, T.B.; Lee, S.R.; Yang, S.H. The Role of Tumor Necrosis Factor Alpha (TNF-α) in Autoimmune Disease and Current TNF-α Inhibitors in Therapeutics. Int. J. Mol. Sci. 2021, 22, 2719. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Zheng, S.G. Hall of Fame among Pro-inflammatory Cytokines: Interleukin-6 Gene and Its Transcriptional Regulation Mechanisms. Front. Immunol. 2016, 7, 236681. [Google Scholar] [CrossRef] [PubMed]
- Markiewski, M.M.; Lambris, J.D. The role of complement in inflammatory diseases from behind the scenes into the spotlight. Am. J. Pathol. 2007, 171, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Merle, N.S.; Church, S.E.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement System Part I—Molecular Mechanisms of Activation and Regulation. Front. Immunol. 2015, 6, 134383. [Google Scholar] [CrossRef] [PubMed]
- Elieh Ali Komi, D.; Shafaghat, F.; Kovanen, P.T.; Meri, S. Mast cells and complement system: Ancient interactions between components of innate immunity. Allergy 2020, 75, 2818–2828. [Google Scholar] [CrossRef]
- Dunkelberger, J.R.; Song, W.-C. Complement and its role in innate and adaptive immune responses. Cell Res. 2010, 20, 34–50. [Google Scholar] [CrossRef] [PubMed]
- Barnes, T.C.; Anderson, M.E.; Moots, R.J. The many faces of interleukin-6: The role of IL-6 in inflammation, vasculopathy, and fibrosis in systemic sclerosis. Int. J. Rheumatol. 2011, 2011, 721608. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Yang, J.; Huang, Q.; Zhang, Y.; Peng, C.; Zhang, Y.; He, Y.; Shi, J.; Li, W.; Hu, J.; et al. Biodistribution and pulmonary toxicity of intratracheally instilled graphene oxide in mice. NPG Asia Mater. 2013, 5, e44. [Google Scholar] [CrossRef]
- Spragg, R.G.; Bernard, G.R.; Checkley, W.; Curtis, J.R.; Gajic, O.; Guyatt, G.; Hall, J.; Israel, E.; Jain, M.; Needham, D.M.; et al. Beyond mortality: Future clinical research in acute lung injury. Am. J. Respir. Crit. Care Med. 2010, 181, 1121–1127. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, N.M.; Meliton, A.Y.; Meliton, L.N.; Dudek, S.M.; Leff, A.R. Secretory group V phospholipase A2 regulates acute lung injury and neutrophilic inflammation caused by LPS in mice. Am. J. Physiol. Lung Cell Mol. Physiol. 2009, 296, L879–L887. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.L.; Xie, Q.M.; Shen, Y.H.; Jiang, J.J.; Chen, Y.Y.; Yao, H.Y.; Zhou, J.Y. Mannose prevents lipopolysaccharide-induced acute lung injury in rats. Inflamm. Res. 2008, 57, 104–110. [Google Scholar] [CrossRef]
- Asti, C.; Ruggieri, V.; Porzio, S.; Chiusaroli, R.; Melillo, G.; Caselli, G.F. Lipopolysaccharide-induced lung injury in mice. I. Concomitant evaluation of inflammatory cells and haemorrhagic lung damage. Pulm. Pharmacol. Ther. 2000, 13, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Dong, C.; Yuan, M.; Gao, G.; Wang, S.; Liu, X.; Han, H.; Li, B. Instilled air promotes lipopolysaccharide-induced acute lung injury. Exp. Ther. Med. 2014, 7, 816–820. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, Y.; Franco, C.M.M. Acetylated Triterpene Glycosides and Their Biological Activity from Holothuroidea Reported in the Past Six Decades. Mar. Drugs 2016, 14, 147. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, Y.; Zhang, W.; Franco, C. Discovery of novel saponins from the viscera of the sea cucumber Holothuria lessoni. Mar. Drugs 2014, 12, 2633–2667. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, Y.; Zhang, W.; Franco, C.M.M. Distribution of Saponins in the Sea Cucumber Holothuria lessoni; the Body Wall Versus the Viscera, and Their Biological Activities. Mar. Drugs 2018, 16, 423. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, Y.; Zhang, W.; Chataway, T.; Franco, C. Structure elucidation of five novel isomeric saponins from the viscera of the sea cucumber Holothuria lessoni. Mar. Drugs 2014, 12, 4439–4473. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, Y.; Franco, C.M.M. Structure Elucidation of New Acetylated Saponins, Lessoniosides A, B, C, D, and E, and Non-Acetylated Saponins, Lessoniosides F and G, from the Viscera of the Sea Cucumber Holothuria lessoni. Mar. Drugs 2015, 13, 597–617. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.C.; Tsai, Y.F.; Pan, Y.L.; Hwang, T.L. Understanding the role of neutrophils in acute respiratory distress syndrome. Biomed. J. 2021, 44, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Bao, Z.; Ye, Q.; Gong, W.; Xiang, Y.; Wan, H. Humanized monoclonal antibody against the chemokine CXCL-8 (IL-8) effectively prevents acute lung injury. Int. Immunopharmacol. 2010, 10, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Zhang, J.; Wang, H.; Xu, J.; He, J.; Liu, L.; Zhang, T.; Chen, R.; Kang, J. Phenolic Acid Profiling, Antioxidant, and Anti-Inflammatory Activities, and miRNA Regulation in the Polyphenols of 16 Blueberry Samples from China. Molecules 2017, 22, 312. [Google Scholar] [CrossRef]
- Soccio, M.; Laus, M.N.; Flagella, Z.; Pastore, D. Assessment of Antioxidant Capacity and Putative Healthy Effects of Natural Plant Products Using Soybean Lipoxygenase-Based Methods. Overview. Mol. 2018, 23, 3244. [Google Scholar] [CrossRef] [PubMed]
- Roy, N.; Barnett, M.; Knoch, B.; Dommels, Y.; McNabb, W. Nutrigenomics applied to an animal model of Inflammatory Bowel Diseases: Transcriptomic analysis of the effects of eicosapentaenoic acid- and arachidonic acid-enriched diets. Mutat. Res. 2007, 622, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Belmiro, C.L.; Castelo-Branco, M.T.; Melim, L.M.; Schanaider, A.; Elia, C.; Madi, K.; Pavão, M.S.; de Souza, H.S. Unfractionated heparin and new heparin analogues from ascidians (chordate-tunicate) ameliorate colitis in rats. J. Biol. Chem. 2009, 284, 11267–11278. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Weng, J.; Yu, L.; Huang, Q.; Jiang, Y.; Guo, X. Role of TLR4-p38 MAPK-Hsp27 signal pathway in LPS-induced pulmonary epithelial hyperpermeability. BMC Pulm. Med. 2018, 18, 178. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Durst, R.; Wrolstad, R. AOAC 2005.02: Total Monomeric Anthocyanin Pigment Content of Fruit Juices, Beverages, Natural Colorants, and Wines-pH Differential Method. In Official Methods of Analysis of AOAC International; AOAC: Rockville, MD, USA, 2005; pp. 37–39. [Google Scholar]
- Sandoval-Ramírez, B.A.; Catalán, Ú.; Fernández-Castillejo, S.; Rubió, L.; Macià, A.; Solà, R. Anthocyanin Tissue Bioavailability in Animals: Possible Implications for Human Health. A Systematic Review. J. Agric. Food Chem. 2018, 66, 11531–11543. [Google Scholar] [CrossRef] [PubMed]
- Amararathna, M.; Hoskin, D.W.; Rupasinghe, H.P.V. Cyanidin-3-O-Glucoside-Rich Haskap Berry Administration Suppresses Carcinogen-Induced Lung Tumorigenesis in A/JCr Mice. Molecules 2020, 25, 3823. [Google Scholar] [CrossRef]
- Wu, Y.; Yang, J.; Xu, C.; Li, Q.; Ma, Y.; Zhao, S.; Zhuang, J.; Shen, F.; Wang, Q.; Feng, F.; et al. Sea cucumber (Acaudina leucoprocta) peptides extended the lifespan and enhanced antioxidant capacity via DAF-16/DAF-2/SOD-3/OLD-1/PEPT-1 in Caenorhabditis elegans. Front. Nutr. 2022, 9, 1065145. [Google Scholar] [CrossRef]
- Dakrory, A.I.; Fahmy, S.R.; Soliman, A.M.; Mohamed, A.S.; Amer, S.A.M. Protective and Curative Effects of the Sea Cucumber Holothuria atra Extract against DMBA-Induced Hepatorenal Diseases in Rats. BioMed Res. Int. 2015, 2015, 563652. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.C. Drug combination studies and their synergy quantification using the Chou-Talalay method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Na, K.; Li, K.; Sang, T.; Wu, K.; Wang, Y.; Wang, X. Anticarcinogenic effects of water extract of sporoderm-broken spores of Ganoderma lucidum on colorectal cancer in vitro and in vivo. Int. J. Oncol. 2017, 50, 1541–1554. [Google Scholar] [CrossRef] [PubMed]
- Matute-Bello, G.; Downey, G.; Moore, B.B.; Groshong, S.D.; Matthay, M.A.; Slutsky, A.S.; Kuebler, W.M. An official American Thoracic Society workshop report: Features and measurements of experimental acute lung injury in animals. Am. J. Respir. Cell Mol. Biol. 2011, 44, 725–738. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Groups | Days of Observation | Percentage (%) | |
|---|---|---|---|---|
| Day 0 (g) | Day 7 (g) | |||
| Body Weight (g) | 1—Control | 25.04 ± 0.66 | 25.18 ± 0.69 | 0.52 ± 0.22 |
| 2—LPS | 26.03 ± 0.49 | 26.34 ± 0.46 | 0.85 ± 0.41 | |
| 3—LPS + BP | 25.83 ± 0.35 | 25.59 ± 0.25 | 0.75 ± 0.29 | |
| 4—LPS + WB | 24.28 ± 0.45 | 24.72 ± 0.46 | 1.84 ± 0.42 * | |
| 5—LPS + BP + WB | 24.56 ± 0.64 | 25.43 ± 0.59 | 3.85 ± 0.79 ** | |
| Source of Extract | Bioactive Biomarker Compound in the Extract | The Concentration of the Biomarker Compound in the Extract | Dosage of Extract (7 Days) | Reference Used for the Selected Animal Dose |
|---|---|---|---|---|
| Sea cucumber (Body parts/internal organs) | Frondoside A (Saponin) | 0.36 mg frondoside A/g DW extract | 27.7 mg extract/mouse/day | Adrian and Collin [34] |
| Wild blueberry | Anthocyanins (Polyphenol) | 18.95 mg P3GE/g DW extract | 50.7 mg extract/mouse/day | Sandoval-Ramírez, Catalán [73] |
| Gene | Forward | Reverse | Accession No. |
|---|---|---|---|
| GAPDH | GGGAAGCCCATCACCATCTT | GCCTTCTCCATGGTGGTGAA | NM_008084.3 |
| ACTIN | CTCTGGCTCCTAGCACCATGAAGA | GTAAAACGCAGCTCAGTAACAGTCCG | NM_007393.5 |
| NF-κB | AAGAACAGAGACCGCTGGTG | CAGGTTCTGCATCCCCTCTG | XM_006509023.5 |
| IL-6 | CTGCAAGAGACTTCCATCCAG | AGTGGTATAGACAGGTCTGTTGG | NM_031168 |
| TLR4 | ACTCAGCAAAGTCCCTGATGACA | AGGTGGTGTAAGCCATGCCA | NM_021297.3 |
| p38 (MAPK14) | GCCGCTTAGTCACATACCACT | GTCCCCGTCAGACGCATTAT | NM_001357724.1 |
| JNK1 (MAPK8) | CTTCAGAAGCAGAAGCCCCA | TGTGCTAAAGGAGACGGCTG | NM_016700.4 |
| ERK1 (MAPK3) | ACACTGGCTTTCTGACGGAG | TGATGCGCTTGTTTGGGTTG | NM_011952.2 |
| ERK2 (MAPK1) | TTGCTTTCTCTCCCGCACAA | AGCCCTTGTCCTGACCAATTT | NM_011949.3 |
| Jun (AP-1) | AAGAAGCTCACAAGTCCGGG | GAGGGCATCGTCGTAGAAGG | NM_010591.2 |
| FOS (AP-1) | TGTTCCTGGCAATAGCGTGT | TCAGACCACCTCGACAATGC | NM_010234.3 |
| IL-1β | GAAATGCCACCTTTTGACAGTG | TGGATGCTCTCATCAGGACAG | NM_008361 |
| TNF-α | CAGGCGGTGCCTATGTCTC | CGATCACCCCGAAGTTCAGTAG | NM_013693 |
| IL-4 | CGACTGCACAGCAGTTCCA | CTCTGGTTGGCTTCCTTCACA | NM_172348 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fagbohun, O.F.; Thilakarathna, W.P.D.W.; Zhou, J.; Lehmann, C.; Jiao, G.; Rupasinghe, H.P.V. Sea Cucumber and Blueberry Extracts Suppress Inflammation and Reduce Acute Lung Injury through the Regulation of NF-κB/MAPK/JNK Signaling Pathway in Lipopolysaccharide-Treated C57BL/6 Mice. Molecules 2024, 29, 1511. https://doi.org/10.3390/molecules29071511
Fagbohun OF, Thilakarathna WPDW, Zhou J, Lehmann C, Jiao G, Rupasinghe HPV. Sea Cucumber and Blueberry Extracts Suppress Inflammation and Reduce Acute Lung Injury through the Regulation of NF-κB/MAPK/JNK Signaling Pathway in Lipopolysaccharide-Treated C57BL/6 Mice. Molecules. 2024; 29(7):1511. https://doi.org/10.3390/molecules29071511
Chicago/Turabian StyleFagbohun, Oladapo F., Wasitha P. D. W. Thilakarathna, Juan Zhou, Christian Lehmann, Guangling Jiao, and H. P. Vasantha Rupasinghe. 2024. "Sea Cucumber and Blueberry Extracts Suppress Inflammation and Reduce Acute Lung Injury through the Regulation of NF-κB/MAPK/JNK Signaling Pathway in Lipopolysaccharide-Treated C57BL/6 Mice" Molecules 29, no. 7: 1511. https://doi.org/10.3390/molecules29071511
APA StyleFagbohun, O. F., Thilakarathna, W. P. D. W., Zhou, J., Lehmann, C., Jiao, G., & Rupasinghe, H. P. V. (2024). Sea Cucumber and Blueberry Extracts Suppress Inflammation and Reduce Acute Lung Injury through the Regulation of NF-κB/MAPK/JNK Signaling Pathway in Lipopolysaccharide-Treated C57BL/6 Mice. Molecules, 29(7), 1511. https://doi.org/10.3390/molecules29071511