

‘Sorrento’ and ‘Tulare’ Walnut Cultivars: Morphological Traits and Phytochemical Enhancement of Their Shell Waste
 ,
,  ,
,  , , and
, , and 
Abstract
1. Introduction
2. Results and Discussion
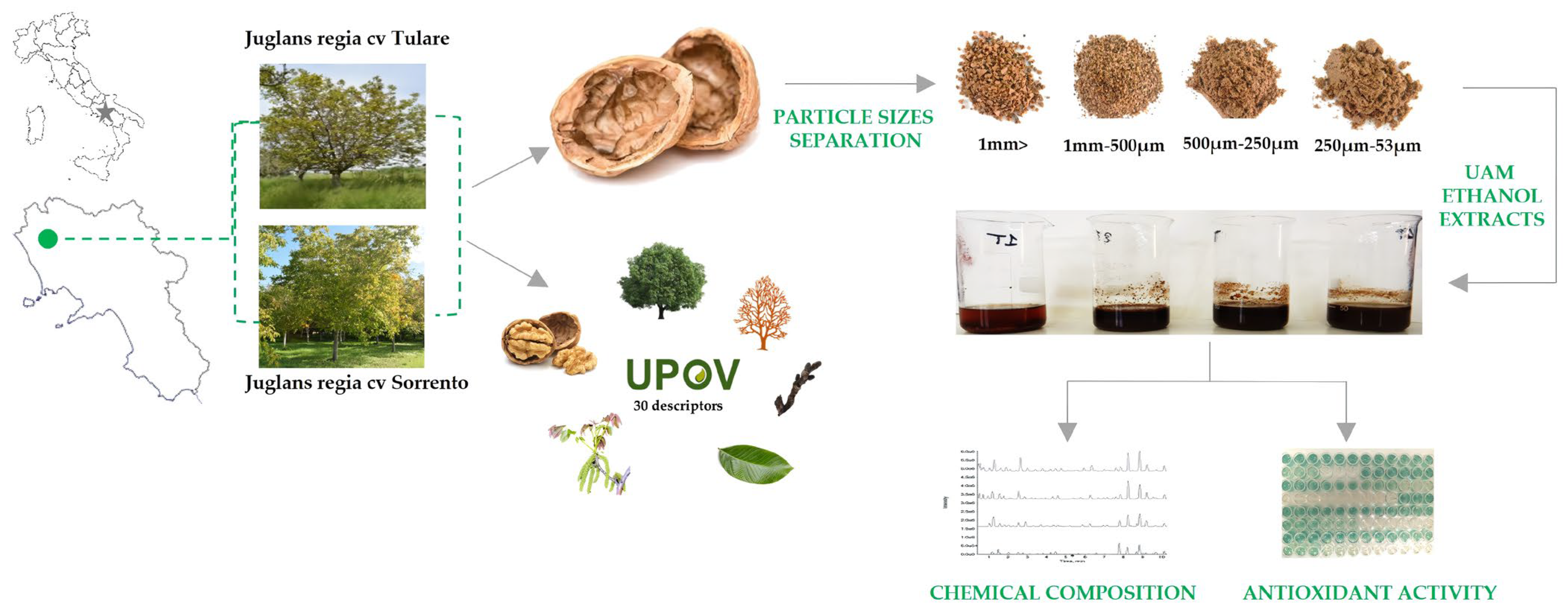
2.1. Morphological and Phenological Traits
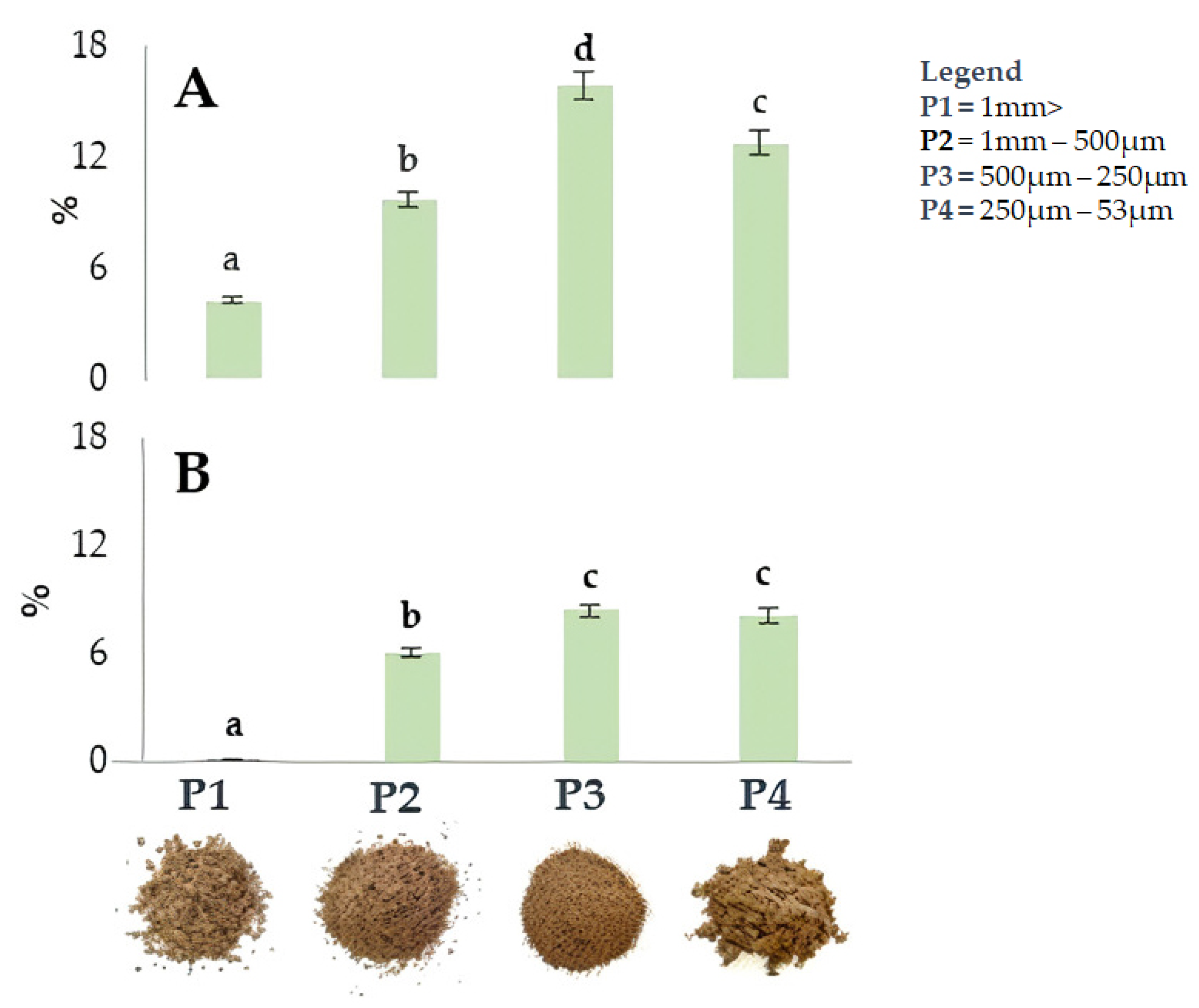
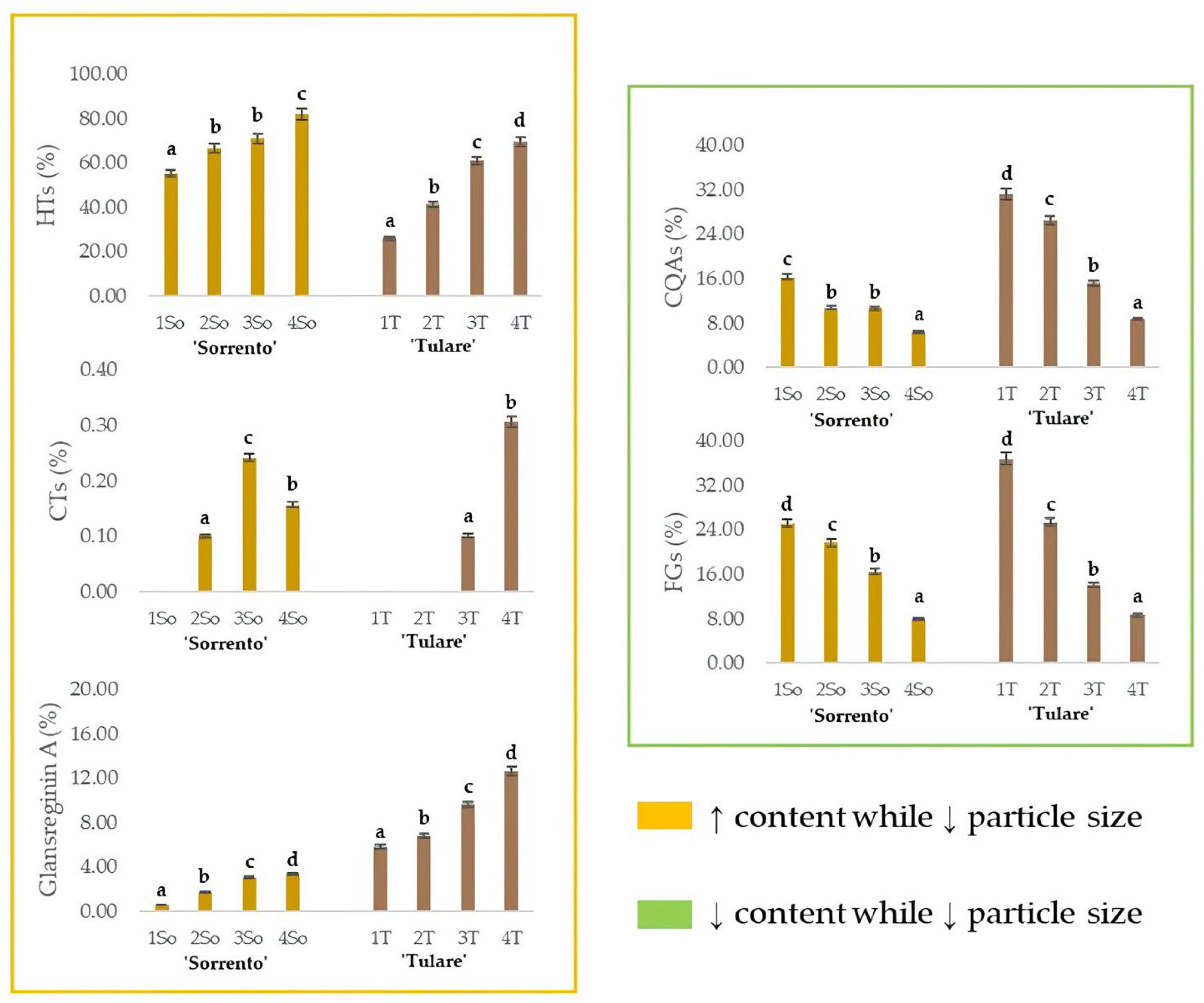
2.2. Extraction of Walnut Endocarp and Bioactive Compounds
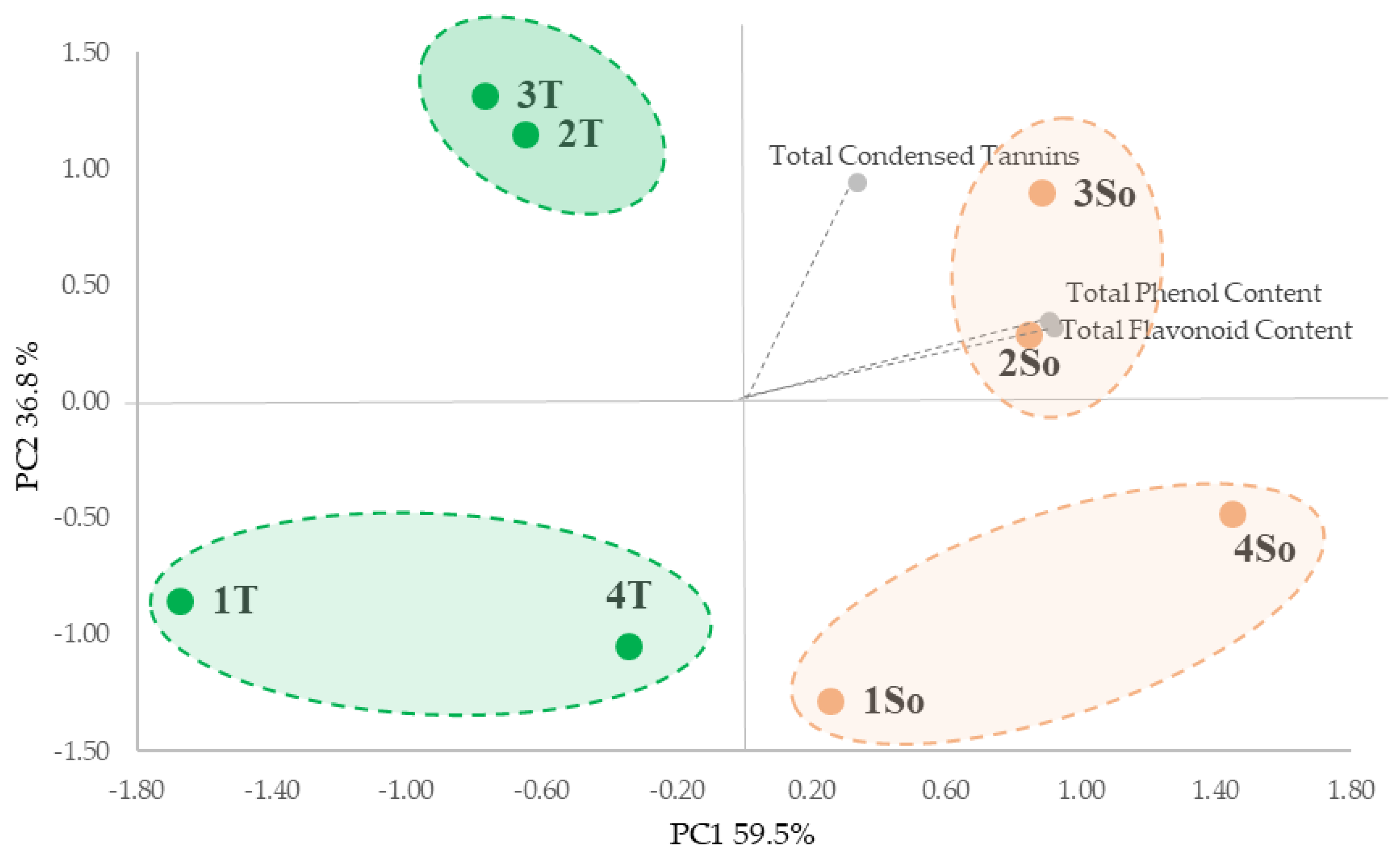
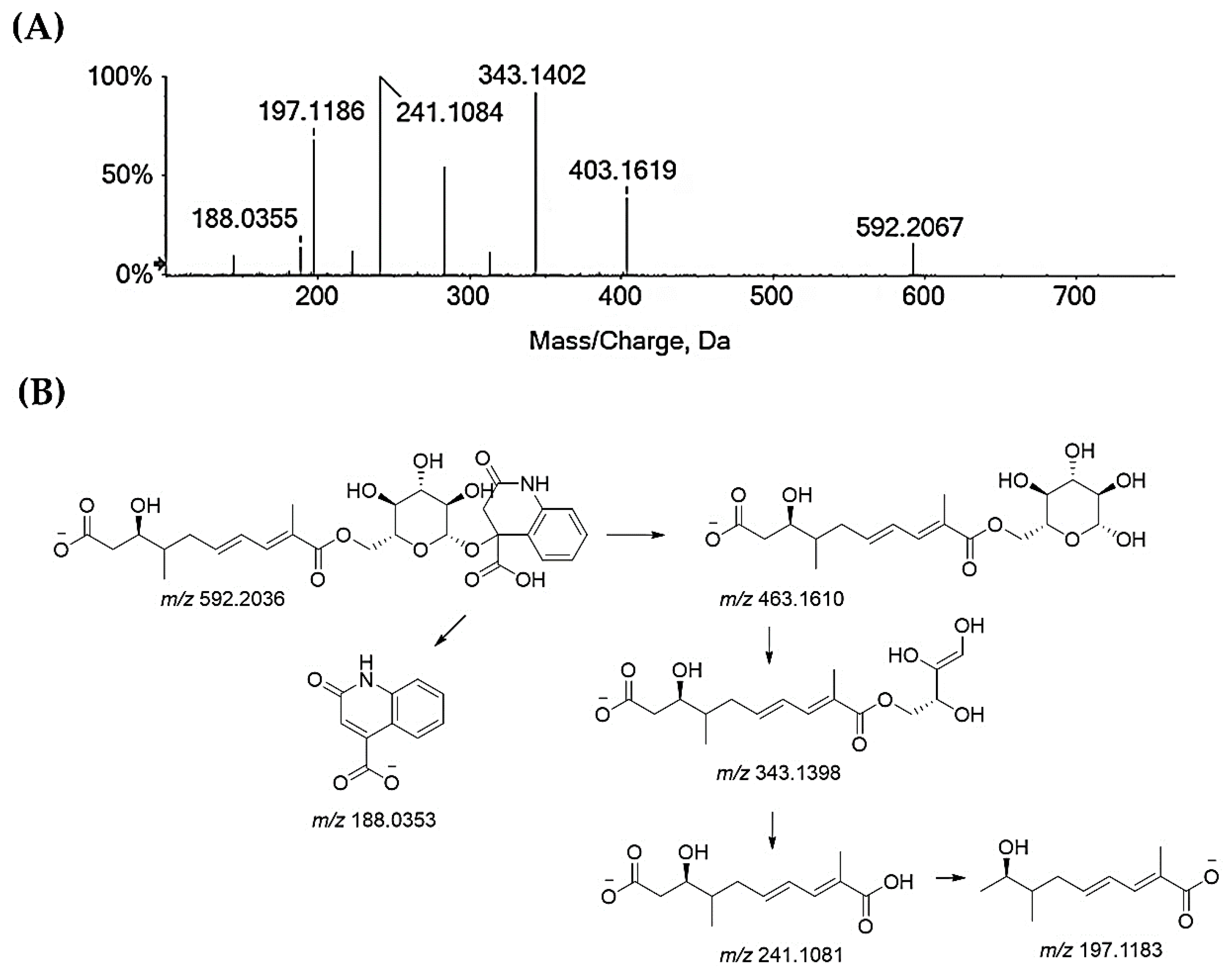
2.3. UHPLC-HR-MS/MS Analysis
3. Materials and Methods
3.1. Walnut Samples
3.2. Extraction of Sorrento and Tulare cv. Walnut Shell
3.3. Determination of DPPH and ABTS Radical Scavenging Capacity
3.4. Determination of Fe(III) Reducing Power
3.5. Determination of Phenol and Flavonoid Content
3.6. Determination of Condensed Tannins
3.7. UHPLC-ESI-QqTOF-MS/MS Analysis
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mc Granahan, G.; Leslie, C. Walnuts (Juglans). Acta Hortic. 1991, 290, 907–974. [Google Scholar] [CrossRef]
- Avanzato, D.; Mc Granahan, G.H.; Vahdati, K.; Botu, M.; Iannamico, L.; Assche, J. Following walnut footprints (Juglans regia L.): Cultivation and culture, folklore and history, traditions and uses. J. Hortic. Sci. 2014, 17, 442. [Google Scholar]
- Shah, R.A.; Bakshi, P.; Sharma, N.; Jasrotia, A.; Itoo, H.; Gupta, R.; Singh, A. Diversity Assessment and selection of superior Persian walnut (Juglans regia L.) trees of seedling origin from north-Western Himalayan region. Resour. Environ. Sustain. 2021, 3, 100015. [Google Scholar] [CrossRef]
- Bottema, S. The Holocene history of walnut, sweet-chestnut, manna-ash and plane tree in the Eastern Mediterranean. Pallas 2000, 52, 35–59. [Google Scholar]
- Pollegioni, P.; Del Lungo, S.; Müller, R.; Woeste, K.E.; Chiocchini, F.; Clark, J.; Hemery, G.E.; Mapelli, S.; Villani, F.; Malvolti, M.E. Biocultural diversity of common walnut (Juglans regia L.) and sweet chestnut (Castanea sativa Mill.) across Eurasia. Ecol. Evol. 2020, 10, 11192–11216. [Google Scholar] [CrossRef] [PubMed]
- Beer, R.; Kaiser, F.; Schmidt, K.; Ammann, B.; Carraro, G.; Grisa, E.; Tinner, W. Vegetation history of the Walnut Forests in Kyrgyzstan (Central Asia): Natural or anthropogenic origin? Quat. Sci. Rev. 2008, 27, 621–632. [Google Scholar] [CrossRef]
- Fürst, S.; Schulz, S. Natural landscape or anthropogenic environment? A case study on the ‘wild’ fruit and walnut forests in Southern Kyrgyzstan. In Utilisation and Management of Natural Resources in Kyrgyzstan, 1st ed.; Dörre, A., Schütte, S., Eds.; Berlin Geographical Papers: Berlin, Germany, 2014; pp. 71–87. [Google Scholar]
- Pollegioni, P.; Woeste, K.E.; Chiocchini, F.; Del Lungo, S.; Olimpieri, I.; Tortolano, V.; Clark, J.; Hemery, G.E.; Mapelli, S.; Malvolti, M.E. Ancient humans influenced the current spatial genetic structure of common walnut populations in Asia. PLoS ONE 2015, 10, e0135980. [Google Scholar] [CrossRef]
- Ducci, F.; De Rogatis, A.; Proietti, R. Protezione delle risorse genetiche di Juglans regia L. Ann. Ist. Sper. Selv. 1997, 25/26, 35–55. [Google Scholar]
- Pollegioni, P.; Woeste, K.; Chiocchini, F.; Del Lungo, S.; Ciolfi, M.; Olimpieri, I.; Tortolano, V.; Clark, J.; Hemery, G.E.; Mapelli, S.; et al. Rethinking the history of common walnut (Juglans regia L.) in Europe: Its origins and human interactions. PLoS ONE 2017, 12, e0172541. [Google Scholar] [CrossRef]
- Persic, M.; Mikulic-Petkovsek, M.; Slatnar, A.; Solar, A.; Veberic, R. Changes in phenolic profiles of red-colored pellicle walnut and hazelnut kernel during ripening. Food Chem. 2018, 252, 349–355. [Google Scholar] [CrossRef]
- Bayazit, S.; Sumbul, A. Determination of fruit quality and fatty acid composition of Turkish walnut (Juglans regia) cultivars and genotypes grown in subtropical climate of eastern Mediterranean region. Int. J. Agric. Biol. 2012, 14, 419–424. [Google Scholar]
- Li, Q.; Yin, R.; Zhang, Q.; Wang, X.; Hu, X.; Gao, Z.; Duan, Z. Chemometrics Analysis on the Content of Fatty Acid Compositions in Different Walnut (Juglans regia L.) Varieties. Eur. Food Res. Technol. 2017, 243, 2235–2242. [Google Scholar] [CrossRef]
- Jia, Y.; Yuan, B.; Yang, Y.; Zheng, C.; Zhou, Q. Flavor characteristics of peeled walnut kernels under two-steps roasting processes. Food Chem. 2023, 423, 136290. [Google Scholar] [CrossRef] [PubMed]
- Hama, J.R.; Omer, R.A.; Rashid, R.S.M.; Mohammad, N.-E.-A.; Thoss, V. The Diversity of Phenolic Compounds along Defatted Kernel, Green Husk and Leaves of Walnut (Juglans regia L.). Anal. Chem. Lett. 2016, 6, 35–46. [Google Scholar] [CrossRef]
- Sánchez-González, C.; Ciudad, C.J.; Noé, V.; Izquierdo-Pulido, M. Health Benefits of Walnut Polyphenols: An Exploration beyond Their Lipid Profile. Crit. Rev. Food Sci. Nutr. 2017, 57, 3373–3383. [Google Scholar] [CrossRef]
- Rusu, M.E.; Gheldiu, A.-M.; Mocan, A.; Moldovan, C.; Popa, D.-S.; Tomuta, I.; Vlase, L. Process optimization for improved phenolic compounds recovery from walnut (Juglans regia L.) septum: Phytochemical profile and biological activities. Molecules 2018, 23, 2814. [Google Scholar] [CrossRef]
- Wang, J.; Liu, J.; John, A.; Jiang, Y.; Zhu, H.; Yang, B.; Wen, L. Structure identification of walnut peptides and evaluation of cellular antioxidant activity. Food Chem. 2022, 388, 132943. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, H.; Tanaka, J.; Kikuchi, M.; Fukuda, T.; Ito, H.; Hatano, T.; Yoshida, T. Walnut Polyphenols Prevent Liver Damage Induced by Carbon Tetrachloride and D-Galactosamine: Hepatoprotective Hydrolyzable Tannins in the Kernel Pellicles of Walnut. J. Agric. Food Chem. 2008, 56, 4444–4449. [Google Scholar] [CrossRef]
- Miao, F.; Shan, C.; Ning, D. Walnut oil alleviates lps-induced intestinal epithelial cells injury by inhibiting TLR4/myd88/NF-ΚB pathway activation. J. Food Biochem. 2021, 45, e13955. [Google Scholar] [CrossRef]
- Altemimi, A.B.; Al-haliem, S.M.; Alkanan, Z.T.; Mohammed, M.J.; Hesarinejad, M.A.; Najm, M.A.; Bouymajane, A.; Cacciola, F.; Abedelmaksoud, T.G. Exploring the phenolic profile, antibacterial, and antioxidant properties of walnut leaves (Juglans regia L.). Food Sci. Nutr. 2023, 11, 6845–6853. [Google Scholar] [CrossRef]
- Ros, E.; Izquierdo-Pulido, M.; Sala-Vila, A. Beneficial effects of walnut consumption on human health. Curr. Opin. Clin. Nutr. Metab. Care 2018, 21, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Esselun, C.; Dieter, F.; Sus, N.; Frank, J.; Eckert, G.P. Walnut oil reduces AΒ levels and increases neurite length in a cellular model of early alzheimer disease. Nutr. J. 2022, 14, 1694. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, M.; Shiravi, A.; Vaezi, G.; Sepehri, H.; Hojati, V.; Khori, V. A non-insulin herbal treatment to improve liver tissue in diabetic rats through Gavage of walnut oil enriched with a phytosterol. Cogent Biol. 2019, 5, 1648628. [Google Scholar] [CrossRef]
- Hayes, D.; Angove, M.J.; Tucci, J.; Dennis, C. Walnuts (Juglans regia) Chemical Composition and Research in Human Health. Crit. Rev. Food Sci. Nutr. 2016, 56, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Khir, R.; Pan, Z. Walnuts. In Integrated Processing Technologies for Food and Agricultural By-Products, 1st ed.; Pan, Z., Zhang, R., Zicari, S., Eds.; Academic Press: London, UK, 2019; pp. 391–411. [Google Scholar]
- Chamorro, F.; Carpena, M.; Lourenço-Lopes, C.; Taofiq, O.; Otero, P.; Cao, H.; Xiao, J.; Simal-Gandara, J.; Prieto, M.A. By-products of Walnut (Juglans regia) as a source of bioactive compounds for the formulation of nutraceuticals and Functional Foods. Biol. Life Sci. Forum 2022, 12, 35. [Google Scholar]
- Prasad, R.B.N. Walnuts and pecans. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Trugo, L., Finglas, P.M., Eds.; Academic Press: Oxford, UK, 2003; pp. 6071–6079. [Google Scholar]
- Soto-Madrid, D.; Gutiérrez-Cutiño, M.; Pozo-Martínez, J.; Zúñiga-López, M.C.; Olea-Azar, C.; Matiacevich, S. Dependence of the ripeness stage on the antioxidant and antimicrobial properties of walnut (Juglans regia L.) green husk extracts from industrial by-products. Molecules 2021, 26, 2878. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Wang, S.; Rakita, M.; Wang, Y.; Han, Q.; Xu, Q. Effect of Ultrasound-Assisted Extraction of Phenolic Compounds on the Characteristics of Walnut Shells. Food Nutr. Sci. 2018, 9, 1034–1045. [Google Scholar] [CrossRef]
- Jahanban-Esfahlan, A.; Ostadrahimi, A.; Tabibiazar, M.; Amarowicz, R. A comprehensive review on the chemical constituents and functional uses of walnut (Juglans spp.) husk. Int. J. Mol. Sci. 2019, 20, 3920. [Google Scholar] [CrossRef]
- Herrera, R.; Hemming, J.; Smeds, A.; Gordobil, O.; Willför, S.; Labidi, J. Recovery of bioactive compounds from hazelnuts and walnuts shells: Quantitative–qualitative analysis and chromatographic purification. Biomolecules 2020, 10, 1363. [Google Scholar] [CrossRef]
- Antreich, S.J.; Xiao, N.; Huss, J.C.; Gierlinger, N. A belt for the cell: Cellulosic Wall thickenings and their role in morphogenesis of the 3D puzzle cells in Walnut Shells. J. Exp. Bot. 2021, 72, 4744–4756. [Google Scholar] [CrossRef]
- Song, X.; Guan, W.; Qin, H.; Han, X.; Wu, L.; Ye, Y. Properties of poly(lactic acid)/walnut shell/hydroxyapatite composites prepared with fused deposition modeling. Sci. Rep. 2022, 12, 11563. [Google Scholar] [CrossRef]
- FAO. 2020. Available online: www.fao.org/statistics/en (accessed on 18 November 2022).
- Crisosto, C.H.; Donis-Gonzalez, I.R.; Wang, S.C.; Lampinen, B.D. Removing English walnut (Juglans regia) ready-to-use shelled walnuts consumption barriers. Horticulturae 2023, 9, 891. [Google Scholar] [CrossRef]
- ISTAT. Coltivazioni: Coltivazioni Legnose Fruttifere. 2023. Available online: http://dati.istat.it/Index.aspx?QueryId=33705 (accessed on 27 November 2023).
- Foroni, I.; Rao, R.; Woeste, K.; Gallitelli, M. Characterisation of Juglans regia L. with SSR markers and evaluation of genetic relationships among cultivars and the ‘Sorrento’ Landrace. J. Hortic. Sci. Biotechnol. 2005, 80, 49–53. [Google Scholar] [CrossRef]
- Pollegioni, P.; Woeste, K.; Olimpieri, I.; Marandola, D.; Cannata, F.; Emilia Malvolti, M. Long-Term Human Impacts on Genetic Structure of Italian Walnut Inferred by SSR Markers. Tree Genet. Genomes 2011, 7, 707–723. [Google Scholar] [CrossRef]
- Di Pierro, E.A.; Franceschi, P.; Endrizzi, I.; Farneti, B.; Poles, L.; Masuero, D.; Khomenko, I.; Trenti, F.; Marrano, A.; Vrhovsek, U.; et al. Valorization of traditional Italian walnut (Juglans regia L.) production: Genetic, nutritional and sensory characterization of locally grown varieties in the Trentino region. Plants 2022, 11, 1986. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Zhou, H.; Cao, D.; Yan, F.; Chen, P.; Wang, J.; Woeste, K.; Chen, X.; Fei, Z.; An, H.; et al. Domestication and selection footprints in Persian walnuts (Juglans regia). PLoS Genet. 2022, 18, e1010513. [Google Scholar] [CrossRef] [PubMed]
- Foroni, I.; Woeste, K.; Monti, L.M.; Rao, R. Identification of ‘sorrento’ walnut using simple sequence repeats (SSRS). Genet. Resour. Crop Evol. 2007, 54, 1081–1094. [Google Scholar] [CrossRef]
- Law No. 238 Organic discipline of Wine Cultivation and Wine Production and Trade. Official Gazette of the Italian Republic. 28 December 2016, p. 45. Available online: https://faolex.fao.org/docs/pdf/ita167457.pdf (accessed on 3 January 2024).
- Romano, R.; De Luca, L.; Vanacore, M.; Genovese, A.; Cirillo, C.; Aiello, A.; Sacchi, R. Compositional and Morphological Characterization of ‘Sorrento’ and ‘Chandler’ Walnuts. Foods 2022, 11, 761. [Google Scholar] [CrossRef] [PubMed]
- Fallah, M.; Vahdati, K.; Hasani, D.; Rasouli, M.; Sarikhani, S. Breeding of Persian walnut: Aiming to introduce late-leafing and early-harvesting varieties by targeted hybridization. Sci. Hortic. 2022, 295, 110885. [Google Scholar] [CrossRef]
- McGranahan, G.H.; Forde, H.I.; Snyder, R.G.; Wilbur Reil, G.S.; Hasey, J.; Ramos, D.E. ‘Tulare’ Persian Walnut. Hortscience 1992, 27, 186–187. [Google Scholar] [CrossRef]
- Mali, P.S.; Kumar, P. Optimization of microwave assisted extraction of bioactive compounds from black bean waste and evaluation of its antioxidant and antidiabetic potential in vitro. Food Chem. Advanc. 2023, 3, 100543. [Google Scholar] [CrossRef]
- Ferrara, E.; Pecoraro, M.T.; Cice, D.; Piccolella, S.; Formato, M.; Esposito, A.; Petriccione, M.; Pacifico, S. A joint approach of morphological and UHPLC-HRMS analyses to throw light on the autochthonous ‘verdole’ chestnut for nutraceutical innovation of its waste. Molecules 2022, 27, 8924. [Google Scholar] [CrossRef]
- Orooji, Y.; Han, N.; Nezafat, Z.; Shafiei, N.; Shen, Z.; Nasrollahzadeh, M.; Karimi-Maleh, H.; Luque, R.; Bokhari, A.; Klemeš, J.J. Valorisation of nuts biowaste: Prospects in sustainable bio(nano)catalysts and environmental applications. J. Clean. Prod. 2022, 347, 131220. [Google Scholar] [CrossRef]
- Dzah, C.S.; Duan, Y.; Zhang, H.; Wen, C.; Zhang, J.; Chen, G.; Ma, H. The effects of ultrasound assisted extraction on yield, antioxidant, anticancer and antimicrobial activity of polyphenol extracts: A Review. Food Biosci. 2020, 35, 100547. [Google Scholar] [CrossRef]
- Zhang, Q.-W.; Lin, L.-G.; Ye, W.-C. Techniques for extraction and isolation of natural products: A comprehensive review. Chin. Med. 2018, 13, 20. [Google Scholar] [CrossRef]
- Shen, L.; Pang, S.; Zhong, M.; Sun, Y.; Qayum, A.; Liu, Y.; Rashid, A.; Xu, B.; Liang, Q.; Ma, H.; et al. A comprehensive review of Ultrasonic assisted extraction (UAE) for bioactive components: Principles, advantages, equipment, and Combined Technologies. Ultrason. Sonochem. 2023, 101, 106646. [Google Scholar] [CrossRef]
- Ali, A.; Riaz, S.; Sameen, A.; Naumovski, N.; Iqbal, M.W.; Rehman, A.; Mehany, T.; Zeng, X.-A.; Manzoor, M.F. The disposition of bioactive compounds from fruit waste, their extraction, and analysis using novel technologies: A Review. Processes 2022, 10, 2014. [Google Scholar] [CrossRef]
- Queirós, C.S.G.P.; Cardoso, S.; Lourenço, A.; Ferreira, J.; Miranda, I.; Lourenço, M.J.; Helena Pereira, H. Characterization of walnut, almond, and pine nut shells regarding chemical composition and extract composition. Biomass Convers. Biorefin. 2020, 10, 175–188. [Google Scholar] [CrossRef]
- Wang, H.; Asker, K.; Zhan, C.; Wang, N. Transcriptomic and metabolic analysis of fruit development and identification of genes involved in raffinose and hydrolysable tannin biosynthesis in walnuts. J. Agric. Food Chem. 2021, 69, 8050–8062. [Google Scholar] [CrossRef] [PubMed]
- Regueiro, J.; Sánchez-González, C.; Vallverdú-Queralt, A.; Simal-Gándara, J.; Lamuela-Raventós, R.; Izquierdo-Pulido, M. Comprehensive identification of walnut polyphenols by liquid chromatography coupled to linear ion trap–orbitrap mass spectrometry. Food Chem. 2014, 152, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Labuckas, D.; Maestri, D.; Lamarque, A. Molecular characterization, antioxidant and protein solubility-related properties of polyphenolic compounds from walnut (Juglans regia). Nat. Prod. Commun. 2016, 11, 637–640. [Google Scholar] [CrossRef]
- Wang, P.; Zhong, L.; Yang, H.; Zhu, F.; Hou, X.; Wu, C.; Zhang, R.; Cheng, Y. Comparative analysis of antioxidant activities between dried and fresh walnut kernels by metabolomic approaches. LWT 2022, 155, 112875. [Google Scholar] [CrossRef]
- Sheng, F.; Hu, B.; Jin, Q.; Wang, J.; Wu, C.; Luo, Z. The analysis of phenolic compounds in walnut husk and pellicle by UPLC-Q-orbitrap hrms and HPLC. Molecules 2021, 26, 3013. [Google Scholar] [CrossRef]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhnert, N. Hierarchical scheme for LC-MSn identification of chlorogenic acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef] [PubMed]
- Pacifico, S.; Piccolella, S.; Nocera, P.; Tranquillo, E.; Dal Poggetto, F.; Catauro, M. New insights into phenol and polyphenol composition of Stevia Rebaudiana leaves. J. Pharm. Biomed. Anal. 2019, 163, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Bai, H.; Jing, Y.; Li, W.; Yin, S.; Zhou, H. A pair of taxifolin-3-O-arabinofuranoside isomers from Juglans regia L. Nat. Prod. Res. 2016, 31, 945–950. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, H.; Yu, J.; Wang, J.; Cui, Q.; Hou, L.; Tian, J. Traditional uses, chemical composition, and pharmacological effects of Diaphragma Juglandis Fructus: A Review. J. Ethnopharmacol. 2023, 312, 116440. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Cheng, Y.; Wang, S.; Li, J.; Ren, H.; Qiao, Y.; Li, Q.; Wang, Y. The chemical constituents of diaphragma juglandis fructus and their inhibitory effect on α-glucosidase activity. Molecules 2022, 27, 3045. [Google Scholar] [CrossRef] [PubMed]
- Petrović, M.; Pastor, F.; Đurović, S.; Veljović, S.; Gorjanović, S.; Sredojević, M.; Vukosavljević, P. Evaluation of novel Green Walnut Liqueur as a source of antioxidants: Multi-method approach. J. Food Sci. Technol. 2020, 58, 2160–2169. [Google Scholar] [CrossRef]
- Żurek, N.; Pycia, K.; Pawłowska, A.; Potocki, L.; Kapusta, I.T. Chemical profiling, bioactive properties, and anticancer and antimicrobial potential of Juglans regia L. leaves. Molecules 2023, 28, 1989. [Google Scholar] [CrossRef]
- Ho, K.-V.; Hsieh, H.-Y.; Roy, A.; Foote, S.; McDonald, P.; Coggeshall, M.V.; Ito, H.; Lei, Z.; Sumner, L.W.; Stewart, G.C.; et al. Quantification and characterization of biological activities of Glansreginin A in black walnuts (Juglans nigra). Sci. Rep. 2023, 13, 18860. [Google Scholar] [CrossRef]
- Haramiishi, R.; Okuyama, S.; Yoshimura, M.; Nakajima, M.; Furukawa, Y.; Ito, H.; Amakura, Y. Identification of the characteristic components in walnut and anti-inflammatory effect of Glansreginin A as an indicator for quality evaluation. Biosci. Biotechnol. Biochem. 2020, 84, 187–197. [Google Scholar] [CrossRef]
- Ho, K.-V.; Lei, Z.; Sumner, L.; Coggeshall, M.; Hsieh, H.-Y.; Stewart, G.; Lin, C.-H. Identifying antibacterial compounds in black walnuts (Juglans nigra) using a metabolomics approach. Metabolites 2018, 8, 58. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Zhang, Q.; Jiang, M.; Chen, M.; Xu, X.; Wang, D.; Pan, Y.; Liu, X. Systematic characterization of the metabolites of defatted walnut powder extract in vivo and screening of the mechanisms against NAFLD by UPLC-Q-exactive orbitrap MS combined with network pharmacology. J. Ethnopharmacol. 2022, 285, 114870. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Chen, D.; Cao, L.; Zhao, S. Effects of pressed degreased walnut meal extracts on lipid metabolism in postnatally monosodium glutamate-induced mice and 3T3-L1 preadipocytes. J. Funct. Foods 2017, 31, 89–96. [Google Scholar] [CrossRef]
- UPOV. Walnut (Juglans regia L.). In Guidelines for the Conduct of Tests for Distinctness, Uniformity and Stability; TG/125/7; International Union for the Protection of New Varieties of Plants (UPOV): Geneva, Switzerland, 2017; p. 34. [Google Scholar]
- Formato, M.; Piccolella, S.; Zidorn, C.; Pacifico, S. UHPLC-HRMS Analysis of Fagus sylvatica (Fagaceae) Leaves: A Renewable Source of Antioxidant Polyphenols. Antioxidants 2021, 10, 1140. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | RT (min) | Tentative Assignment | Formula | [M-H]− Found (m/z) | Error (ppm) | RDB |
|---|---|---|---|---|---|---|
| 1 | 0.432 | Hexahydroxydiphenoyl-D- hexoside isomer 1 | C20H18O14 | 481.0637 | 2.7 | 12 |
| 2 | 0.640 | Bis-HHDP-hexose isomer 1 | C34H24O22 | 783.0686 | 0.7 | 23 |
| 3 | 0.687 | Glucogallin | C13H16O10 | 331.0671 | 1.3 | 6 |
| 4 | 0.750 | Gallic acid | C13H16O10 | 169.0142 | 4.5 | 5 |
| 5 | 0.933 | Hexahydroxydiphenoyl-D- hexoside isomer 2 | C20H18O14 | 481.0624 | 3.4 | 12 |
| 6 | 1.00 | Digalloyl hexose isomer 1 | C20H20O14 | 483.0801 | 4.3 | 11 |
| 7 | 1.042 | Bis-HHDP-hexose isomer 2 | C34H24O22 | 783.0718 | 4.0 | 23 |
| 8 | 1.317 | Galloyl-HHDP-hexose isomer 1 | C20H26O23 | 633.0778 | −2.2 | 8 |
| 9 | 1.517 | Bis-HHDP-hexose isomer 3 | C34H24O22 | 783.0728 | 5.3 | 23 |
| 10 | 1.892 | Digalloyl hexose isomer 2 | C20H20O14 | 483.0799 | 3.9 | 11 |
| 11 | 2.092 | 3-O-Caffeoylquinic acid | C16H18O9 | 353.0880 | 0.5 | 8 |
| 12 | 2.604 | Galloyl-HHDP-hexose isomer 2 | C20H26O23 | 633.0768 | −3.8 | 8 |
| 13 | 2.619 | Bis-HHDP-hexose isomer 4 | C34H24O22 | 783.0710 | 3.0 | 23 |
| 14 | 2.866 | Coumaroylquinic acid isomer 1 | C16H18O8 | 337.0921 | −2.3 | 8 |
| 15 | 2.952 | Coumaroylquinic acid isomer 2 | C16H18O8 | 337.0936 | 2.1 | 8 |
| 16 | 3.522 | Procyanidin B2 isomer 1 | C30H26O12 | 577.1368 | 2.9 | 18 |
| 17 | 3.745 | Procyanidin B2 isomer 2 | C30H26O12 | 577.1375 | 4.1 | 18 |
| 18 | 3.766 | Digalloyl-HHDP-hexose isomer 1 | C27H30O27 | 785.0902 | −0.6 | 13 |
| 19 | 4.099 | 4-O-caffeoylquinic acid | C16H18O9 | 353.0889 | 2 | 8 |
| 20 | 4.831 | Galloyl-HHDP-hexose isomer 3 | C20H26O23 | 633.0764 | −4.4 | 8 |
| 21 | 5.587 | Coumaroylquinic acid isomer 3 | C16H18O8 | 337.0915 | −4.1 | 8 |
| 22 | 6.720 | Digalloyl-HHDP-hexose isomer 2 | C27H30O27 | 785.0876 | −3.3 | 13 |
| 23 | 7.588 | Ellagic acid hexoside | C20H16O13 | 463.0498 | −4.4 | 13 |
| 24 | 7.611 | Ellagic acid pentosyl hexoside | C25H24O17 | 595.0913 | −4.7 | 14 |
| 25 | 7.640 | Taxifolin−3-O-pentoside isomer 1 | C20H20O11 | 435.0946 | 3.0 | 11 |
| 26 | 7.856 | Taxifolin−3-O-pentoside isomer 2 | C20H20O11 | 435.0933 | −0.4 | 11 |
| 27 | 8.179 | Quercetin galloyl hexoside | C28H24O16 | 615.0981 | −1.7 | 17 |
| 28 | 8.192 | Ellagic acid pentoside isomer 1 | C19H14O12 | 433.0423 | 2.4 | 12 |
| 29 | 8.201 | Ellagic acid pentoside isomer 2 | C19H14O12 | 433.0429 | 3.8 | 12 |
| 30 | 8.272 | Ellagic acid | C14H6O8 | 300.9990 | 3.0 | 12 |
| 31 | 8.804 | Glansreginin A | C28H35NO13 | 592.2036 | 2.8 | 12 |
| 32 | 8.927 | Quercetin-3-O-deoxyhexoside | C21H20O11 | 447.0933 | 2.3 | 12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrara, E.; Cice, D.; Piccolella, S.; Esposito, A.; Petriccione, M.; Pacifico, S. ‘Sorrento’ and ‘Tulare’ Walnut Cultivars: Morphological Traits and Phytochemical Enhancement of Their Shell Waste. Molecules 2024, 29, 805. https://doi.org/10.3390/molecules29040805
Ferrara E, Cice D, Piccolella S, Esposito A, Petriccione M, Pacifico S. ‘Sorrento’ and ‘Tulare’ Walnut Cultivars: Morphological Traits and Phytochemical Enhancement of Their Shell Waste. Molecules. 2024; 29(4):805. https://doi.org/10.3390/molecules29040805
Chicago/Turabian StyleFerrara, Elvira, Danilo Cice, Simona Piccolella, Assunta Esposito, Milena Petriccione, and Severina Pacifico. 2024. "‘Sorrento’ and ‘Tulare’ Walnut Cultivars: Morphological Traits and Phytochemical Enhancement of Their Shell Waste" Molecules 29, no. 4: 805. https://doi.org/10.3390/molecules29040805
APA StyleFerrara, E., Cice, D., Piccolella, S., Esposito, A., Petriccione, M., & Pacifico, S. (2024). ‘Sorrento’ and ‘Tulare’ Walnut Cultivars: Morphological Traits and Phytochemical Enhancement of Their Shell Waste. Molecules, 29(4), 805. https://doi.org/10.3390/molecules29040805