

Egg White Protein–Proanthocyanin Complexes Stabilized Emulsions: Investigation of Physical Stability, Digestion Kinetics, and Free Fatty Acid Release Dynamics



 and
and
Abstract

1. Introduction
2. Results and Discussion
2.1. Alterations in Protein Structure
2.2. Variations of Amino Acid Side Chain Groups
2.3. Fourier Transform Infrared Spectroscopy (FTIR) and X-ray Diffraction (XRD) Analysis
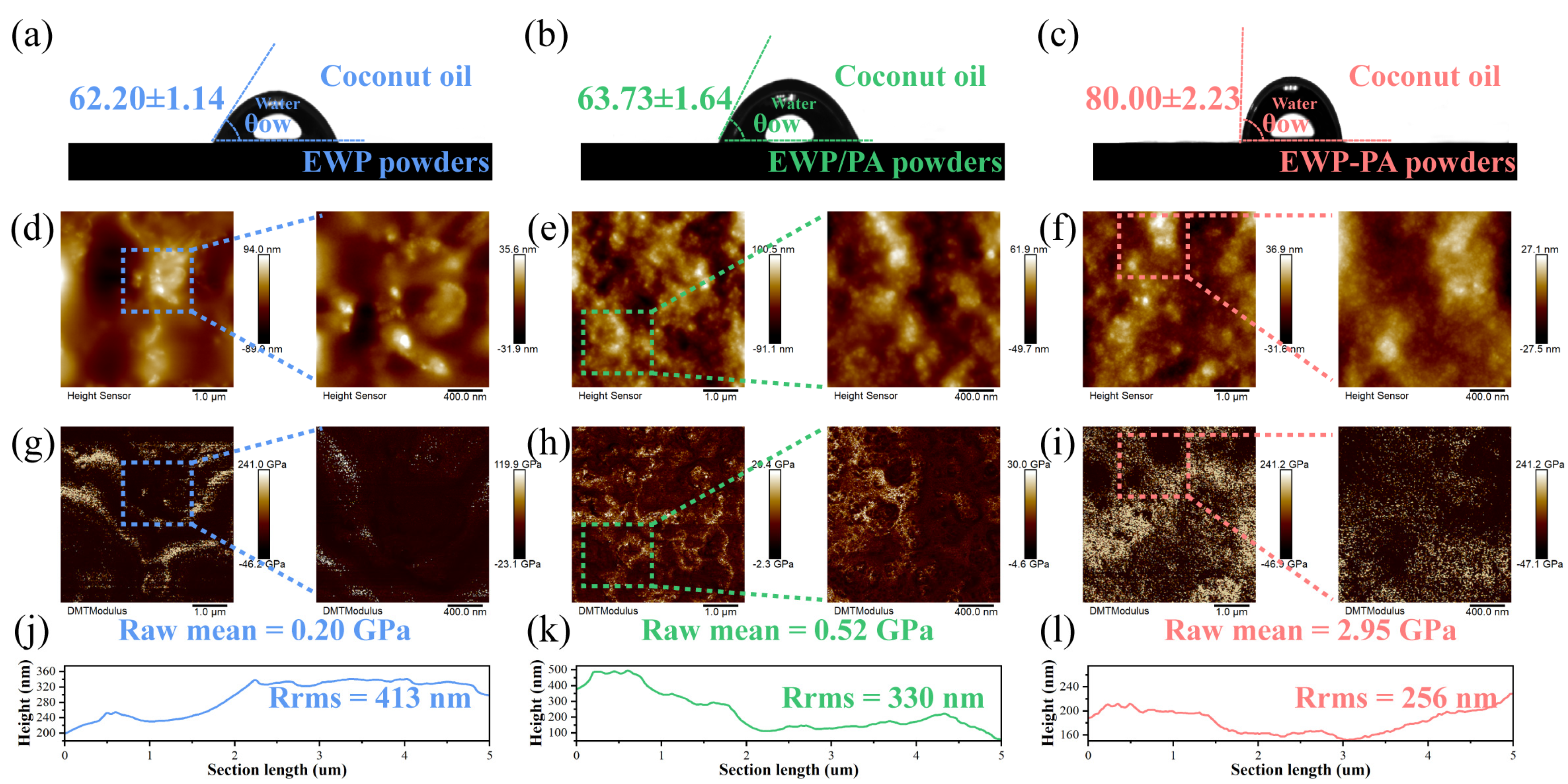
2.4. Wettability and AFM Analysis
2.5. Microstructure Observation
2.6. Emulsion Stability
2.7. Emulsion Digestive Kinetics and FFA Release Kinetics
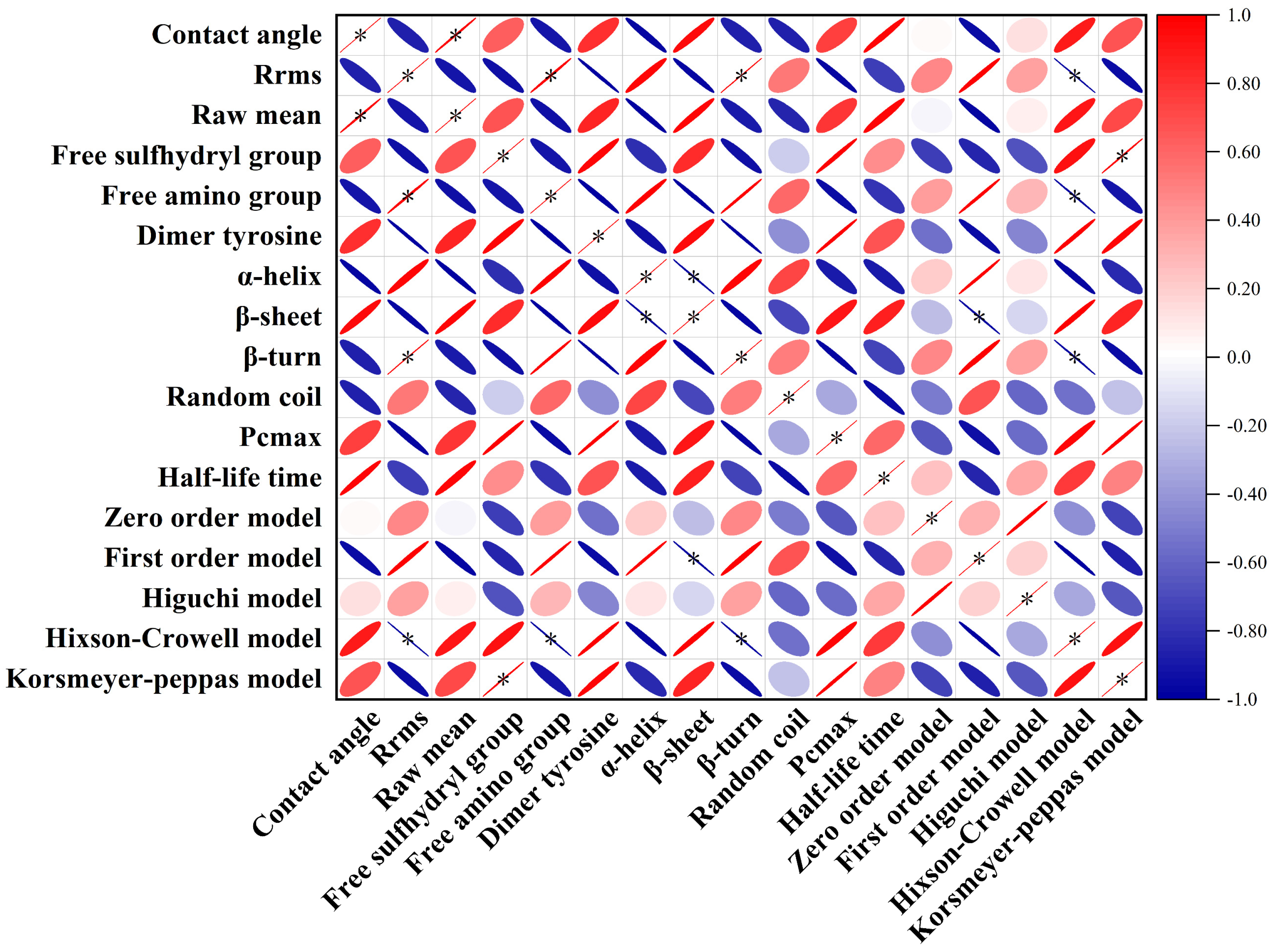
2.8. Correlation Analysis
3. Materials and Methods
3.1. Materials
3.2. Formulation of Egg White Protein–Proanthocyanin Complexes
3.3. Determination of Protein Structural Modifications
3.3.1. Intrinsic Fluorescence Spectroscopy
3.3.2. Surface Hydrophobicity
3.3.3. Circular Dichroism (CD) Spectroscopy
3.4. Alterations of Amino Acid Side Chain Groups in Proteins
3.4.1. Dimer Tyrosine
3.4.2. Free Sulfhydryl Group
3.4.3. Free Amino Group
3.5. FTIR
3.6. XRD
3.7. Three-Phase Contact Angle
3.8. AFM
3.9. Preparation of Emulsion
3.10. Microscopic Observations
3.10.1. Optical Microscope
3.10.2. Cryo-SEM
3.11. Stability Evaluation
3.11.1. Storage Stability
3.11.2. pH Stability
3.12. Emulsion Digestion Kinetics
3.13. FFA Release Kinetics
3.14. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lian, Z.; Yang, S.; Cheng, L.; Liao, P.; Dai, S.; Tong, X.; Tian, T.; Wang, H.; Jiang, L. Emulsifying properties and oil–water interface properties of succinylated soy protein isolate: Affected by conformational flexibility of the interfacial protein. Food Hydrocoll. 2023, 136, 108224. [Google Scholar] [CrossRef]
- Yang, M.; Liu, J.; Yang, X.; Li, S.; Li, C.; Liu, B.; Ma, S.; Liu, X.; Du, Z.; Zhang, T.; et al. Effect of glycation degree on the in vitro simulated gastrointestinal digestion: A promising formulation for egg white gel with controlled digestibility. Food Chem. 2021, 349, 129096. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Niu, F.; Gu, L.; Li, X.; Yang, H.; Zhou, B.; Wang, J.; Su, Y.; Yang, Y. Formation of fibrous or granular egg white protein microparticles and properties of the integrated emulsions. Food Hydrocoll. 2016, 61, 477–486. [Google Scholar] [CrossRef]
- Razi, S.M.; Fahim, H.; Amirabadi, S.; Rashidinejad, A. An overview of the functional properties of egg white proteins and their application in the food industry. Food Hydrocoll. 2023, 135, 108183. [Google Scholar] [CrossRef]
- Dong, W.; Zhang, X.; Ding, L.; Liu, C.; Ai, M.; Jin, Y.; Isobe, K.; Handa, A.; Cai, Z. Enhancement of emulsification properties by modulation of egg white protein fibril structure with different heating times. Food Hydrocoll. 2023, 135, 08203. [Google Scholar] [CrossRef]
- Lu, F.; Chi, Y.; Chi, Y. High-temperature glycosylation of saccharides to modify molecular conformation of egg white protein and its effect on the stability of high internal phase emulsions. Food Res. Int. 2024, 176, 113825. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, Y.; Duan, W.; Wang, Q.; An, F.; Luo, P.; Huang, Q. Ball-milling is an effective pretreatment of glycosylation modified the foaming and gel properties of egg white protein. J. Food End. 2022, 319, 110908. [Google Scholar] [CrossRef]
- Liu, G.; Sun, P.; Yan, J.; Shao, P.; Feng, S. Regulation of Nanoliposome Rigidity and Bioavailability of Oligomeric Proanthocyanidin with Phytosterols Containing Different C3 Branches. ACS Appl. Mater. Interfaces 2023, 5, 43414–43430. [Google Scholar] [CrossRef]
- Zhu, F. Proanthocyanidins in cereals and pseudocereals. Crit. Rev. Food Sci. 2018, 59, 1521–1533. [Google Scholar] [CrossRef]
- Qi, Q.; Chu, M.; Yu, X.; Xie, Y.; Li, Y.; Du, Y.; Liu, X.; Zhang, Z.; Shi, J.; Yan, N. Anthocyanins and proanthocyanidins: Chemical structures, food sources, bioactivities, and product development. Food Rev. Int. 2023, 39, 4581–4609. [Google Scholar] [CrossRef]
- Bartzoka, E.D.; Lange, H.; Mosesso, P.; Crestini, C. Synthesis of nano- and microstructures from proanthocyanidins, tannic acid and epigallocatechin-3-O-gallate for active delivery. Green Chem. 2017, 19, 5074–5091. [Google Scholar] [CrossRef]
- Sintara, M.; Li, L.; Cunningham, D.G.; Prior, R.L.; Wu, X.; Chang, T. Single-Laboratory Validation for Determination of Total Soluble Proanthocyanidins in Cranberry Using 4-Dimethylaminocinnamaldehyde. J. AOAC Int. 2018, 101, 805–809. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Huang, F.; Xie, B.; Sun, Z.; McClements, D.J.; Deng, Q. Fabrication and characterization of whey protein isolates- lotus seedpod proanthocyanin conjugate: Its potential application in oxidizable emulsions. Food Chem. 2021, 346, 128680. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Zhao, X.; Xu, X. Comparisons between Myofibrillar Protein–Luteolin Conjugates Fabricated through Covalent and Noncovalent Modification. J. Agric. Food Chem. 2023, 71, 9908–9921. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.; Liao, P.; Wang, Y.; Tian, T.; Tong, X.; Lyu, B.; Cheng, L.; Miao, L.; Qi, W.; Jiang, L.; et al. Soy protein isolate-catechin non-covalent and covalent complexes: Focus on structure, aggregation, stability and in vitro digestion characteristics. Food Hydrocoll. 2023, 135, 108108. [Google Scholar] [CrossRef]
- Wang, W.; Sun, C.; Mao, L.; Ma, P.; Liu, F.; Yang, J.; Gao, Y. The biological activities, chemical stability, metabolism and delivery systems of quercetin: A review. Trends Food Sci. Technol. 2016, 56, 21–38. [Google Scholar] [CrossRef]
- Li, S.; Wang, X.; Zhang, X.; Zhang, H.; Li, S.; Zhou, J.; Fan, L. Interactions between zein and anthocyanins at different pH: Structural characterization, binding mechanism and stability. Food Res. Int. 2023, 166, 112552. [Google Scholar] [CrossRef]
- Ke, C.; Liu, B.; Dudu, O.E.; Zhang, S.; Meng, L.; Wang, Y.; Wei, W.; Cheng, J.; Yan, T. Modification of structural and functional characteristics of casein treated with quercetin via two interaction modes: Covalent and non-covalent interactions. Food Hydrocoll. 2023, 137, 108394. [Google Scholar] [CrossRef]
- Wang, N.; Ma, Z.; Ma, L.; Zhang, Y.; Zhang, K.; Ban, Q.; Wang, X. Synergistic modification of structural and functional characteristics of whey protein isolate by soybean isoflavones non-covalent binding and succinylation treatment: A focus on emulsion stability. Food Hydrocoll. 2023, 144, 108994. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, M.; Zhou, X.; Zang, H.; Zhang, R.; Yang, H.; Jin, S.; Feng, X.; Shan, A. Oxidative stability and gelation properties of myofibrillar protein from chicken breast after post-mortem frozen storage as influenced by phenolic compound-pterostilbene. Int. J. Biol. Macromol. 2022, 221, 1271–1281. [Google Scholar] [CrossRef]
- Zhang, X.; Yin, C.; Hao, J.; Ma, D.; Li, Z.; Li, Y.; Qi, B. Improving the biological activity, functional properties, and emulsion stability of soybean meal hydrolysate via covalent conjugation with polyphenol. Food Chem. 2023, 422, 136255. [Google Scholar] [CrossRef] [PubMed]
- Günal-Köroğlu, D.; Lorenzo, J.M.; Capanoglu, E. Plant-Based Protein-Phenolic Interactions: Effect on different matrices and in vitro gastrointestinal digestion. Food Res. Int. 2023, 173, 113269. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Yue, X.; Shao, J.-H. The adsorption characteristics of 2D fibril and 3D hydrogel aggregates at the O/W interface combining molecular dynamics simulation. Food Hydrocoll. 2022, 128, 107537. [Google Scholar] [CrossRef]
- Wang, L.; Liu, L.; Dong, B.; Zhao, H. Peroxidase-Mediated In Situ Fabrication of Multi-Stimuli-Responsive and Dynamic Protein Nanogels from Tyrosine-Conjugated Biodynamer and Ovablumin. ACS Macro Lett. 2019, 8, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J. The oxidative environment and protein damage. Biochimica BBA Proteins Proteom. 2005, 1703, 93–109. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Li, F.; Wu, W. Effects of rice bran rancidity on the oxidation and structural characteristics of rice bran protein. LWT Food Sci. Technol. 2020, 120, 108943. [Google Scholar] [CrossRef]
- Xu, B.; Chen, J.; Chitrakar, B.; Li, H.; Wang, J.; Wei, B.; Zhou, C.; Ma, H. Effects of flat sweep frequency and pulsed ultrasound on the activity, conformation and microstructure of mushroom polyphenol oxidase. Ultrason. Sonochem. 2022, 82, 105908. [Google Scholar] [CrossRef]
- Hao, L.; Sun, J.; Pei, M.; Zhang, G.; Li, C.; Li, C.; Ma, X.; He, S.; Liu, L. Impact of non-covalent bound polyphenols on conformational, functional properties and in vitro digestibility of pea protein. Food Chem. 2022, 383, 132623. [Google Scholar] [CrossRef]
- Zhang, X.; Ma, D.; Yin, C.; Li, Z.; Hao, J.; Li, Y.; Zhang, S. The biological activity, functionality, and emulsion stability of soybean meal hydrolysate–proanthocyanidin conjugates. Food Chem. 2024, 432, 137159. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, C.; Xing, S.; Chen, Y.; Su, W.; Wang, H.; Tan, M. Sea bass protein-polyphenol complex stabilized high internal phase of algal oil Pickering emulsions to stabilize astaxanthin for 3D food printing. Food Chem. 2023, 417, 135824. [Google Scholar] [CrossRef]
- Sun, J.; Huang, Y.; Liu, T.; Jing, H.; Zhang, F.; Obadi, M.; Xu, B. Evaluation of crossing-linking sites of egg white protein–polyphenol conjugates: Fabricated using a conventional and ultrasound-assisted free radical technique. Food Chem. 2022, 386, 132606. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wei, Z.; Xue, C. Pickering emulsions stabilized by zein–gallic acid composite nanoparticles: Impact of covalent or non-covalent interactions on storage stability, lipid oxidation and digestibility. Food Chem. 2023, 408, 135254. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Gao, X.; Su, Y.; Li, J.; Chang, C.; Gu, L.; Yang, Y. Fabrication of egg white protein/phosphatidylcholine-EGCG co-assembled nanoparticles with sustained release in simulated gastrointestinal digestion and their transcellular permeability in Caco-2 cultures. Food Hydrocoll. 2023, 143, 108906. [Google Scholar] [CrossRef]
- Tong, X.; Cao, J.; Tian, T.; Lyu, B.; Miao, L.; Lian, Z.; Cui, W.; Liu, S.; Wang, H.; Jiang, L. Changes in structure, rheological property and antioxidant activity of soy protein isolate fibrils by ultrasound pretreatment and EGCG. Food Hydrocoll. 2022, 122, 107084. [Google Scholar] [CrossRef]
- Dai, W.; Zhou, L.; Gu, S.; Wang, W.; Xu, Z.; Zhou, X.; Ding, Y. Preparation and characterization of chitosan films incorporating epigallocatechin gallate: Microstructure, physicochemical, and bioactive properties. Int. J. Biol. Macromol. 2022, 211, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yi, X.; Zhang, Z.; Ding, B.; Li, Z.; Luo, Y. High internal phase Pickering emulsions stabilized by tannic acid-ovalbumin complexes: Interfacial property and stability. Food Hydrocoll. 2022, 125, 107332. [Google Scholar] [CrossRef]
- Chen, S.; Du, Y.; Zhang, H.; Wang, Q.; Gong, Y.; Chang, R.; Zhang, J.; Zhang, J.; Yuan, Y.; Liu, B.; et al. The lipid digestion behavior of oil-in-water Pickering emulsions stabilized by whey protein microgels of various rigidities. Food Hydrocoll. 2022, 130, 107735. [Google Scholar] [CrossRef]
- Nikiforidis, C.V.; Scholten, E. High internal phase emulsion gels (HIPE-gels) created through assembly of natural oil bodies. Food Hydrocoll. 2015, 43, 283–289. [Google Scholar] [CrossRef]
- Yi, D.; Jeong, G.; Seo, J.-H.; Yoo, M.J.; Yang, H. Carbon Dots with Tailored Surface Wettability as Pickering Emulsifiers. ACS Appl. Nano Mater. 2022, 5, 10258–10267. [Google Scholar] [CrossRef]
- Li, R.; Xue, H.; Gao, B.; Liu, H.; Han, T.; Hu, X.; Tu, Y.; Zhao, Y. Physicochemical properties and digestibility of thermally induced ovalbumin–oil emulsion gels: Effect of interfacial film type and oil droplets size. Food Hydrocoll. 2022, 131, 107747. [Google Scholar] [CrossRef]
- Huang, X.; Yan, C.; Lin, M.; He, C.; Xu, Y.; Huang, Y.; Zhou, Z. The effects of conjugation of walnut protein isolate with polyphenols on protein solubility, antioxidant activity, and emulsifying properties. Food Res. Int. 2022, 161, 111910. [Google Scholar] [CrossRef]
- Zhang, S.; Feng, X.; Huang, Y.; Wang, Y.; Chen, H.; Yu, Y.; Ma, L.; Zhang, Y.; Dai, H. Facile isolation of cellulose nanofibrils from agro-processing residues and its improved stabilization effect on gelatin emulsion. Int. J. Biol. Macromol. 2022, 216, 272–281. [Google Scholar] [CrossRef]
- Fan, Y.; Liu, Y.; Gao, L.; Zhang, Y.; Yi, J. Oxidative stability and in vitro digestion of menhaden oil emulsions with whey protein: Effects of EGCG conjugation and interfacial cross-linking. Food Chem. 2018, 265, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Lan, H.; Wang, H.; Chen, C.; Hu, W.; Ai, C.; Chen, L.; Teng, H. Flavonoids and gastrointestinal health: Single molecule for multiple roles. Crit. Rev. Food Sci. 2023, 1–19. [Google Scholar] [CrossRef]
- Liu, C.; Guo, Y.; Cheng, Y.; Qian, H. A colon-targeted delivery system of torularhodin encapsulated in electrospinning microspheres, and its co-metabolic regulation mechanism of gut microbiota. Food Hydrocoll. 2023, 135, 108189. [Google Scholar] [CrossRef]
- Yang, M.; Liu, J.; Guo, J.; Yang, X.; Liu, C.; Zhang, M.; Li, Y.; Zhang, H.; Zhang, T.; Du, Z. Tailoring the physicochemical stability and delivery properties of emulsions stabilized by egg white microgel particles via glycation: Role of interfacial particle network and digestive metabolites. Food Hydrocoll. 2022, 131, 107833. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, T.; Jiang, H.; Liu, J.; Wang, E.; Zhang, M.; Liu, X. pH-induced egg white protein foaming properties enhancement: Insight into protein structure and quantitative proteomic analysis at protein adsorption layer. Food Hydrocoll. 2023, 144, 109060. [Google Scholar] [CrossRef]
- Yang, Q.; Wang, Y.; Yang, M.; Liu, X.; Lyu, S.; Liu, B.; Liu, J.; Zhang, T. Effect of glycation degree on the structure and digestion properties of ovalbumin: A study of amino acids and peptides release after in vitro gastrointestinal simulated digestion. Food Chem. 2022, 373, 131331. [Google Scholar] [CrossRef]
- Yan, X.; He, Y.; Bai, X.; McClements, D.J.; Chen, S.; Liu, X.; Liu, F. Development and application of hydrophilic-hydrophobic dual-protein Pickering emulsifiers: EGCG-modified caseinate-zein complexes. Food Res. Int. 2022, 157, 111451. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.; Li, Z.; Li, Y.; Hao, Y.; Du, Z.; Liu, X.; Shang, X.; Liu, J.; Zhang, T. Aggregation of egg white peptides (EWP) induced by proanthocyanidins: A promising fabrication strategy for EWP emulsion. Food Chem. 2023, 400, 134019. [Google Scholar] [CrossRef]
- Cheng, L.; Ye, A.; Hemar, Y.; Singh, H. Modification of the interfacial structure of droplet-stabilised emulsions during in vitro dynamic gastric digestion: Impact on in vitro intestinal lipid digestion. J. Colloid Interf. Sci. 2022, 608, 1286–1296. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Zero Order (R2) | First Order (R2) | Higuchi (R2) | Hixson–Corwell (R2) | Korsmeyer–Peppas (R2) |
|---|---|---|---|---|---|
| EWP emulsion | 0.28528 | 0.99454 | 0.90398 | −2.41088 | 0.96785 |
| EWP/PA emulsion | −0.68737 | 0.98931 | 0.55448 | −1.78145 | 0.98466 |
| EWP–PA emulsion | −0.13539 | 0.98082 | 0.78625 | −1.16254 | 0.98954 |
| Oil | 0.80189 | 0.99805 | 0.97808 | −1.44173 | 0.98062 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Li, S.; Yang, M.; Li, Y.; Liu, X.; Shang, X.; Liu, J.; Du, Z.; Yu, T. Egg White Protein–Proanthocyanin Complexes Stabilized Emulsions: Investigation of Physical Stability, Digestion Kinetics, and Free Fatty Acid Release Dynamics. Molecules 2024, 29, 743. https://doi.org/10.3390/molecules29030743
Zhang T, Li S, Yang M, Li Y, Liu X, Shang X, Liu J, Du Z, Yu T. Egg White Protein–Proanthocyanin Complexes Stabilized Emulsions: Investigation of Physical Stability, Digestion Kinetics, and Free Fatty Acid Release Dynamics. Molecules. 2024; 29(3):743. https://doi.org/10.3390/molecules29030743
Chicago/Turabian StyleZhang, Ting, Shanglin Li, Meng Yang, Yajuan Li, Xuanting Liu, Xiaomin Shang, Jingbo Liu, Zhiyang Du, and Ting Yu. 2024. "Egg White Protein–Proanthocyanin Complexes Stabilized Emulsions: Investigation of Physical Stability, Digestion Kinetics, and Free Fatty Acid Release Dynamics" Molecules 29, no. 3: 743. https://doi.org/10.3390/molecules29030743
APA StyleZhang, T., Li, S., Yang, M., Li, Y., Liu, X., Shang, X., Liu, J., Du, Z., & Yu, T. (2024). Egg White Protein–Proanthocyanin Complexes Stabilized Emulsions: Investigation of Physical Stability, Digestion Kinetics, and Free Fatty Acid Release Dynamics. Molecules, 29(3), 743. https://doi.org/10.3390/molecules29030743